Abstract
Brazil’s Atlantic Forest biome is currently undergoing forest loss due to repeated episodes of devastation. In this biome, bees perform the most frequent pollination system. Over the last decade, network analysis has been extensively applied to the study of plant–pollinator interactions, as it provides a consistent view of the structure of plant–pollinator interactions. The aim of this study was to use palynological studies to obtain an understanding of the relationship between floral visitor bees and the pioneer plant S. didymum in a fragment of the Atlantic Forest, and also learn about the other plants that interact to form this network. Five hundred bees were collected from 32 species distributed into five families: Andrenidae, Apidae, Colletidae, Megachilidae, and Halictidae. The interaction network consisted of 21 bee species and 35 pollen types. The Solanum-type bee species with the highest number of interactions were Anthrenoides sp. 1, Augochlora sp. 2, and Augochloropsis notophos, representing 71.78% of their interactions. Augochloropsis notophos and Augochlora sp. 2 were the only common species in the flowers of S. didymum. Given the results of our study, we conclude that Solanum is an important source of pollen grains for several native bee species, mainly for the solitary species that are more diverse in the south of Brazil. Moreover, our results indicate that bees from the families Halictidae (A. notophos, Augochlora) and Andrenidae (Anthrenoides) are the pollinators of S. didymum.
1. Introduction
In the Atlantic Forest, the most frequent pollination system is accomplished by bees, which mainly look for pollen and nectar in the various plant species, which are used as nutritional sources [1]. This is a mutualistic relationship, in which the plants provide food for pollinators, and in return receive the benefits of pollen transfer [2].
Several groups of bees have morphological, physiological, and behavioral adaptations that enable them to collect floral resources efficiently. Among these adaptations, there is a specialized behavior for pollen collection by the vibration of flowers with poricidal anthers [3], such as the Solanum [4]. This feature requires pollinators to show vibratile behavior (buzz-pollinating) [3] to mechanically remove a larger amount of pollen grains, and a more efficient pollination of their flowers [5]. Its flowers do not offer nectar as a reward, and are therefore dependent on buzz-pollinating bees. Only some bees are capable of vibrating the flowers (buzzing) in order to collect their pollen [6].
The genus Solanum is one of the largest among the angiosperms, comprising approximately 1500 species and maintaining a wide diversity in South America [7]. As most Solanaceae, these plants have an important role as colonizers of open and disturbed sites such as pastures, clearings, forest edges, and roadsides, thus contributing to the regeneration process of altered areas [8].
Solanum didymum Dunal is one of the Solanum species that occurs in the understory of the mixed Araucaria Forest (Araucaria Forest), an Atlantic Forest plant physiognomy [9] that is one of the main plant formations in southern Brazil [10]. Araucaria Forest communities are characterized by the presence of woody species that are phytogeographically related to Austral–Antartic and Andean floras [11,12]. The most physiognomically important tree species is Araucaria angustifolia. Some other typical species found in these forests are Podocarpus lambertii, Drimys brasiliensis, Dicksonia sellowiana, and several species of Myrtaceae, Melastomataceae, Lauraceae, and Solanaceae.
The only information in the literature about Solanum didymum is that it is a selective hygrophytic and heliophytic bush [13]. Therefore, by using palynological studies, the aim was to determine the interaction network between native bees and S. didymum flowers in a fragment of a mixed Ombrophilous Forest (Araucaria Forest), and also their indirect interaction with the other plants in the same forest.
The network theory has substantially helped understandings of the community structure of mutualisms by revealing some invariant properties of these systems [14], and has been widely used to study plant–pollinator interactions [15], as it provides a consistent view of the structure of plant–pollinator interactions [16]. Besides this analysis, this study aimed to obtain an understanding of the relationship between floral visitors’ bees and the pioneer plant S. didymum in a fragment of the Atlantic Forest, using palynological studies, and with this tool also to know the other plants that interact to form this network.
2. Materials and Methods
2.1. Study Area
The study area is in a protected area called the Araucarias Municipal Park (Parque Municipal das Araucárias—PMA) in Guarapuava, Paraná, BR (25°21′06″ S and 51°28′08″ W). The average altitude is 1070 m [17]. It has an area of approximately 104 hectares, and its vegetation comprises mixed Araucaria Forest (42.75%), gallery forest (10.09%), fields (6.8%), wetlands (7.13%), and altered areas (33.23%).
According to the Köppen climate classification, the climate is the Cfb type, moist with no dry season, and with an average temperature in the warmer months below 22 °C and severe frosts [18].
2.2. Sample Design
As S. didymum is a small herbaceous plant that occurs in groups, we studied one of these groups in the understory of the Araucaria Forest fragment of the Araucarias Municipal Park in Guarapuava (PR), southern Brazil. The bees were captured three times per week from September to November 2013, which corresponds to the period of flowering of S. didymum in this park. The collection time was established after observing the floral receptivity, which occurred from 9am to 4pm. The bees were collected directly from the flowers using transparent plastic containers.
2.3. Pollen Collection
Pollen grain material was retrieved from the floral visiting bees’ bodies using 70% alcohol. From this material, the slides were prepared for observation using an optical microscope from the Instituto de Botânica (São Paulo state) based on the method proposed by Erdtman [19].
Bees from the same species collected on the same day and time were grouped. The pollen grains were simultaneously removed from their bodies to prepare one slide to represent the occurrence of that species on that day and at that time.
Plants in a flowering state, or presenting flower buds that were found in the proximity, were collected to prepare the slides, which helped identify pollen types found on the bees’ bodies.
The pollen types found were analyzed qualitatively and quantitatively, and the first 1000 grains per slide were counted. Afterwards, the percentage occurrence of each pollen type was observed, according to the Vergeron classifications [20].
The identified and acetolyzed pollen grains prepared for light microscopy (LM) were digitally photographed using an OLYMPUS BX 50 photomicroscope coupled to a video camera, and a computer using the CellSens software for Windows from the Botanical Institute of São Paulo.
2.4. Assemblage of Floral Bee Visitors of S. didymum
The frequency of occurrence (FO) and species dominance were calculated for each bee species obtained. Frequency of occurrence is the percentage of the number of collections with a given species, and was calculated as FO = (F/N) × 100 [21], where “F” is the number of collections with the species, and “N” is the total number of collections performed. The bee species were classified as primary (FO > 50%), secondary (FO = 25–50%) or accidental (FO < 25%).
The species dominance of bees (D) was calculated as D = (d/n) × 100 (Palma, 1975), where “d” is the abundance of a specific species, and “n” is the total abundance. The species were classified as dominant (D > 5%), accessory (D = 2.5–5%), or accidental (D < 2.5%). According to Palma [22], the FO and D indices, when used together, group and determine the species as common, intermediary, or rare.
2.5. Interaction Network
The interaction network was built between the bees found in S. didymum flowers, the pollen types found in PMA and in these bees’ bodies, and the pollen types of plants that occur in this park.
The network size was calculated through the formula M = ap, where M is the maximum number of possible interactions, a is the number of bees, and p is the number of pollen types.
Connectance (C = E/(ap)) measures the ratio between the number of observed interactions (E) and the number of possible interactions, given that p is the number of plants (pollen types), and a is the number of bee species in the network. To transform these values into percentages, the results were multiplied by 100 [23]. The connectance is a qualitative measure of the network specialization, and also represents the density of interactions in a network. Thus, a highly specialized community presents a low value of connectance [23,24].
In qualitative interaction networks (binary matrix), the average degree (ҟ) was determined, which is the average number of observed connections for species of plants or bees [25,26]. From the adjacency matrix, which was built with data regarding the presence and absence of plant and bee species, Eulerian graphs were prepared using R software version 3.0.1. The evaluation of the network’s degree of nesting, based on the adjacency matrix, was performed using the metrical base NODF (Nestedness Metric Based on Overlap and Decreasing Fill) [27] for more consistent statistical properties. The analyses were produced using ANINHADO software version 3.0 [28].
3. Results
Five hundred bees from 32 species distributed in five families were collected from its flowers: Andrenidae, Apidae, Colletidae, Halictidae, and Megachilidae. The highest species richness was for the families Halictidae, Andrenidae, and Apidae (Table 1).

Table 1.
Number of bee species on flowers of Solanum didymum (N) and their respective dominance indices (FO: species frequency of occurrence; D: species dominance).
The families with the highest number of individuals were Halictidae and Andrenidae. Most of the species were classified as rare. Only Augochloropsis notophos Vachal and Augochlora sp. 2 were common species in the flowers of S. didymum. Anthrenoides sp. 1, Anthrenoides sp. 2, Ceratalictus sp., Augochloropsis sp. A and Augochlora sp. 1 were classified as intermediate species (Table 1).
Out of the 500 bees collected, only 306 participated in the pollen analysis, because they contained a pollen load in their bodies. Two hundred and forty nine slides comprised the network analysis, which recorded 64,442 pollen grains distributed in 35 pollen types of 26 families (Figure 1, Figure 2 and Figure 3).
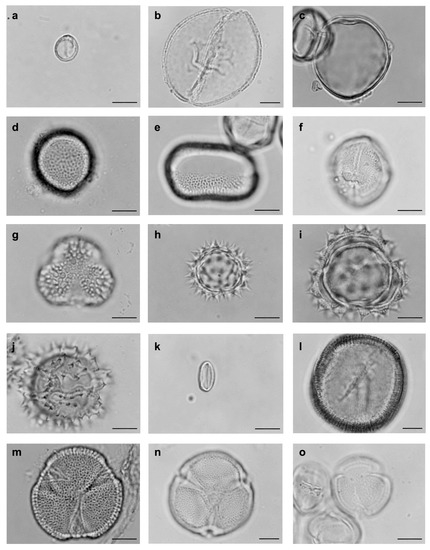
Figure 1.
Pollen types from the bodies of floral visiting bees of Solanum dydimum at the Araucarias Municipal Park in Guarapuava, Paraná state, Brazil. (a) Angiosperms, Magnoliids. Piperaceae, Piper. (b–e) Angiosperms, Monocotyledons. (b) Arecaceae, Butia. (c) Poaceae. (d) Typhaceae, Typha. (e) Monocotyledon type. (f–o) Angiosperms. Eudicotyledons. (f) Anacardiaceae, Schinus. (g) Aquifoliaceae, Ilex. (h) Asteraceae, Baccharis. (i) Asteraceae, Stenachenium. (j) Asteraceae, Vernonia. (k) Begoniaceae, Begonia. (l) Convolvulaceae, Jacquemontia. (m) Dilleniaceae, Davilla. (n) Erythroxylaceae, Erythroxylum. (o) Euphorbiaceae, Sebastiania. (bar = 10 µm).
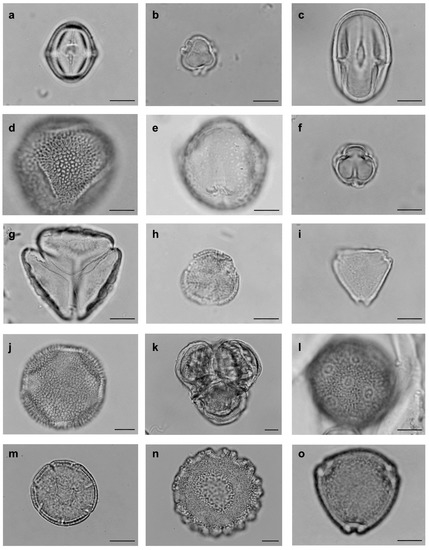
Figure 2.
Pollen types from floral visiting bees’ bodies of Solanum dydimum at Araucarias Municipal Park in Guarapuava, Paraná state, Brazil. (a–o) Angiosperms, Eudicotyledons. (a) Fabaceae, Dalbergia. (b) Fabaceae, Platymiscium. (c) Fabaceae, Senna. (d) Fabaceae, Senna 2. (e) Fabaceae, Sesbania. (f) Fabaceae, Stylosanthes. (g) Loranthaceae, Struthanthus. (h) Menispermaceae. (i) Myrtaceae, Myrcia. (j) Nyctaginaceae, Neea. (k) Onagraceae, Ludwigia. (l) Phytolaccaceae. (m) Rubiaceae, Borreria. (n) Rubiaceae, Diodia. (o) Rubiaceae, Guettarda. (bar = 10 µm).
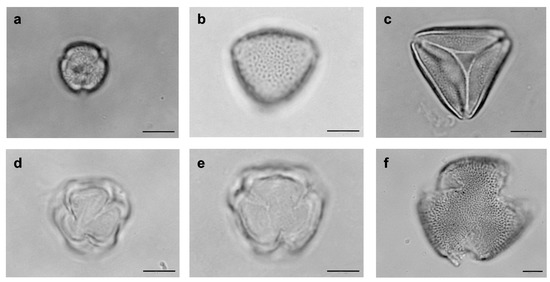
Figure 3.
Pollen types from floral visiting bees’ bodies of Solanum dydimum at Araucarias Municipal Park in Guarapuava, Paraná state, Brazil. (a–f) Angiosperms, Eudicotyledons. (a) Rutaceae, Zanthoxylum. (b) Sapindaceae, Allophylus. (c) Sapindaceae, Matayba. (d) Solanaceae, Solanum. (e) Solanaceae, Solanum didymun (esxicatte). (f) Turneraceae, Turnera. (bar = 10 µm).
The interaction network consisted of 21 bee species (a) and 35 pollen types (p), which enabled 735 interactions (M), and 147 interactions observed between the plant species (p) and the bee species (a) (c = 0.20 (20%)).
Sixteen bee species (66.67%) had interactions with five or less pollen types, and seven (33.33%) bee species had seven or more interactions.
Among the most abundant floral bee visitor species of S. didymum that comprise the network, Anthrenoides sp. 1 (Andrenidae/intermediate) interacted with 23 pollen types found, followed by Augochlora sp. 2 (Halictidae/common) and Augochloropsis notophos (Halictidae/common), which interacted with 22 and 20 pollen types, respectively. From the species considered intermediate, Augochloropsis sp. A (Halictidae) interacted with 16 pollen types, Anthrenoides sp. 2 (Andrenidae) with seven types, and Augochlora sp. 1 (Halictidae) with five types. For the rare species, the number of interactions was below five, except for Augochloropsis cfr sparrilis Vachal (Halictidae), which interacted with seven pollen types, and Halictilus sp. (Halictidae), which interacted with five types. There was no interaction of any bee species with all of the pollen types found.
The bee species with the highest number of interactions with the Solanum type were Anthrenoides sp. 1 (Andrenidae/intermediate), Augochlora sp. 2 (Halictidae/common) and Augochloropsis notophos (Halictidae/common) respectively, representing 71.78% of their interactions. Augochloropsis sp. B (Halictidae/rare), Ceratina (Crewella) sp. (Apidae/rare), Exomalopsis (Exomalopsis) vernoniae Schrottky (Apidae/rare), Plebeia emerina Friese (Apidae/rare), Anthrenoides sp. 3 (Andrenidae/rare), Augochloropsis chloera Moure (Halictidae/rare), Xylocopini (Apidae/rare), A. cfr sparsilis (Halictidae/rare), Colletes argentinus Friese (Colletidae/rare), Augochlorella sp. 2 (Halictidae/rare), Indet.1 (Halictidae/rare) and Pseudaugochlora cfr graminea Fabricius (Halictidae/rare) were the bee species that had fewer interactions with this pollen type (4.80% of interactions).
Concerning the plants, out of the 35 pollen types found, only three interacted with 10 or more bee species, and Solanum was the pollen type that had the highest number of interactions with the different bee species, and therefore a more general type. Nine pollen types (T. Ilex, T. Butia, T. Stenachaerum, T. Jaquemontia, T. Stylosanthes, T. Senna, T. Struthanthus, Pytolacaceae, and T. Monocotyledon) had interactions with only one bee species, and therefore was more specialized.
The average degree (ҟ) of interaction between plant species was 4.26, and the network of degree distribution was heterogeneous, following the truncated power law, and the same happened with bees (ҟ = 7.2).
The bipartite graph (Figure 4) and the NODF = 64.5 index indicated a nested network pattern, with most of the interactions concentrated in a few species.
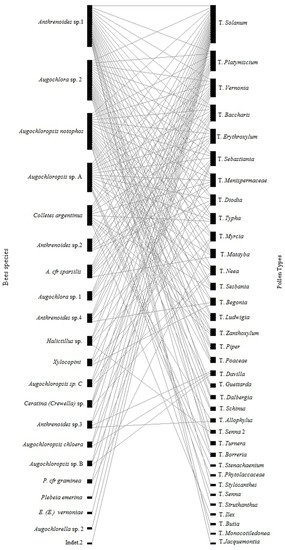
Figure 4.
Eulerian graph representing the interaction networks between pollen types and floral visitor bees of Solanum didymum. The binary matrix qualitatively demonstrates the interactions between species, where A. cfr sparsilis = Augochloropsis cfr sparsilis, P. cfr graminea = Pseudaugochlora cfr gramínea, E. (E.) vernoniae = Exomalopsis (Exomalopsis) vernoniae, Indet. 1 = Indeterminate1, T. = Type.
4. Discussion
The bees of the family Halictidae were the richest and most abundant in S. didymum flowers, and corresponded to the common species in their flowers (A. notophos and Augochlora sp. 2). From the five species classified as intermediate, three belong to this family (Ceratalictus sp., Augochloropsis sp. A, and Augochlora sp. 1), and two belong to the family Andrenidae (Anthrenoides sp. 1 and Anthrenoides sp. 2). The network analysis results also show a strong interaction between these bees and the pollen type Solanum. In several studies on bee communities in southern Brazil, the family Halictidae has shown to be the most diverse, or similar in diversity to that of the family Apidae [29,30,31].
Gonçalves and Melo [30] conducted a standardized survey of bees visiting several species of blooming plants in an area covered by natural grasslands in the Vila Velha State Park, State of Paraná, southern Brazil, from October 2002 to October 2003. Augochlorini and Halictini presented 32 and 26 species, respectively. According to these authors, the numbers are mainly due to the genera Augochlora, Augochloropsis, and Dialictus, which have a high number of species and dense populations.
In our study, Anthrenoides sp. 1 was the most generalist species, interacting with 23 pollen types. This result is interesting, because it suggests that these bees are polylectic, whereas Freitas and Sazima [32], in high-altitude areas in the southeast of the Brazil, found different results. They observed that the main pollinators of Viola species were female bees belonging to the genus Anthrenoides, which search mainly for pollen [32]. According to them, these bees seem to be oligolectic, and obtain large amounts of pollen from Viola by vibrating the flowers or by moving the hook repeatedly back and forth. From the 124 plant species surveyed as part of a study of pollination biology at the community level in the high-altitude grasslands of Serra da Bocaina in southeast Brazil, [33], these bees were observed only on flowers of V. cerasifolia A.St.-Hil. Anthers of Viola, which have dehisce longitudinally, as a function as a single poricidal anther, due to the intimate contact of anthers and the arrangement of the connective projections [32].
Other species that were generalists in our study were Augochlora sp. 2 (22 interactions) and A. notophos (20 interactions), which also interacted strongly with S. didymium. Species of Augochloropisis were in the flowers of Solanum paniculatum L. in a study carried out in the northeast of Brazil, but not those of the Augochlora genus that interacted with several families of plants, least with the Solanaceae species [34].
On the island of Santa Catarina, also in southern Brazil, the broad spectrum of pollen sources characterized Augochlora (A.) esox (Vachal) as a polylectic forager. The pollen from the female’s scopae and from the brood cells included 18 distinct types [35]. Pollen from the families Asteraceae and Melastomataceae were the most frequent, and those from Solanaceae were not registered. For Moure and Hurd [36], the polylectic habit is a common trait in halictid bees, and may help to explain the wide geographical distribution of this species.
If the bees of the Halictidae family are actually polylectic, it seems that there are exceptions, because Dalmazzo and Vossler [37] used pollen analyses in the nests of Augochlora amphitrite and bee flower visitation, and found that the Ludwigia type dominated pollen counts. Based on these results, they concluded that Ludwigia is the most important pollen source in the diet of this bee.
Ludwigia was also one of the pollen types present in the body of the bees collected in the flowers of S. dydimum, but in a small quantity. Their flowers produce pollen grains in large tetrads, with the presence of viscin threads [38], and only specialist bees with long rigid hairs—with few branched in their scopa—and rapid body and leg movements can be the potential pollinators of these plants [39,40], which explains the presence of oligolectic bees in the flowers of Ludwigia in the south and southeast of Brazil [41,42,43,44,45,46].
The relationship of Ludwigia with oligolectic bees was verified in southern Brazil by Buschini et al. [47] when they analyzed the pollen in the Megachile (Moureapis) sp nests. The pollen of Ludwigia peruviana (L.) H. Hara (36%) and Ludwigia sericea (Cambessides) H. Hara (63.6%) accounted for 99.6% of the total amount of pollen collected, and they concluded that this is an oligolectic bee species, and that these plants are their important pollen source. However, when Gonçalves and Buschini [48] collected bees in the flowers of these plants and analyzed the pollen in their bodies, they saw that these plants are generalist in their interactions with bee species, and that the same occurs for most of the bees in relation to these plants. Augochlora amphitrite was among the generalist bee species in their network, interacting with many types of pollen, and not just with Ludwigia type, as observed by Dalmazzo and Vossler [37]. Only the Megachilidae species, which largely depend on the Ludwigia pollen grains, have oligolectic habits.
Another oligolectic bee species that occurs in Guarapuava and has only been recorded from Brazil [49] is Centris tarsata Smith [50,51]. This species uses exclusively Solanum pollen to feed offspring. An analysis of the pollen spectrum based on samples from its nests indicated the presence of 20 pollen types, but the family Solanaceae was visited most often (71%). Solanum americanum Mill. (28.6%) and Sol. variabile Mart. (42.4%) were the primary pollen sources for C. tarsata in Guarapuava [52]. No pollen of S. dydimum was found, because this bee nested only in grassland areas [50,51], and, as previously commented, S. dydimum occurs only in the forest areas in this park.
Another network study carried out in the Guarapuava region, mapping the interactions between Eugenia uniflora L. flowers and floral visitor bees, found that although the most abundant species was Apis mellifera (Linnaeus), the subfamily with the highest number of species was Halictidae (23 species). Andrenidae was represented by just one species (Anthrenoides paolae (Urban)) [53,54]. Similar to S. dydimum, E. uniflora is abundant in the understory of the Araucaria Forest, but their interaction networks were quite different, and no pollen type of Solanum was present in the networks of E. uniflora.
Besides the Solanum pollen type, Platymiscium (Leguminosae), Vernonia (Asteraceae), Baccharis (Asteraceae), and Erythroxylom (Erythroxylaceae) were the most abundant pollen types in the network. Platymiscium was the second most frequent pollen type, but, although it has poricidal anthers, in most of the studies on melittophilus plants, bees are seldom collected on the flowers of these plants, and those of the family Asteraceae are one of the most visited [30,31,34,55,56,57,58].
In the collection of bees visiting several species of blooming plants in the Park of Vila Velha, 150 kms from Guarapuava, Asteraceae was the richest family (43 species), the one that received the most bee visits (55%), and that housed the largest number of species of bees (80 species) [30]. These authors did not record any bees on the flowers of Platymiscium and Erythroxylum. The high number of bee visits to plants of this family may be due to the abundance and richness of this family in the tropics, the accessibility of the resources in their flowers for collection [59], its widespread entomophilic syndrome, the dominance of herbs in secondary fields [60], or presenting massive blooms.
Flowers of Erythroxylum campestre St. Hil., E. suberosum St. Hil., and E. tortuosum Mart were indistinctly visited by 14 wasp species, 14 bee species, and two diptera in the in the central–western region of Brazil. Brachygastra, Polistes, Polybia, and Pepsis wasps were considered effective pollinators of these plants, based on their efficiency in contacting the stigmas, while Trigona spinipes (Fabr.) and Apis mellifera were considered occasional pollinators. Augochloropsis aphrodite was frequent only on the flowers of E. campestre, and Augochloropsis cleopatra (Schrottky) on the flowers of E. suberosum and E. tortuosum These plants are distylic; that is, they have flowers with long styles (pin) as well as short ones (thrum). Their flowers are similar, being small, sweet-scented, creamish, and diurnal, as well as nectar producers (sugar concentration about 20.2%) [61]. In the northeast of Brazil, Erythroxylon catingae Plowman was one of the plants most frequently visited by bees, and A. mellifera was the species of bees most common in its flowers [34].
Given the results of our study, we conclude that Solanum is an important source of pollen grains for several native bee species, mainly for the solitary species that are more diverse in the south of Brazil. Although the behavior of the bees on the flowers of S. didymum was not followed, our results indicate that the bees of the families Halictidae (A. notophos, Augochlora) and Andrenidae (Anthrenoides) are the pollinators of this plant.
Acknowledgments
We would like to thank CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for the scholarship and the Graduate Program in Evolutionary Biology at the Universidade Estadualdo Centro-Oeste (UNICENTRO), Paraná state, Brazil. We would also like to thank Gabriel A. R. Melo from the Universidade Federal do Paraná (UFPR), Paraná state, Brazil, for identifying the bees.
Author Contributions
Francieli Lando carried out field research, drafted the manuscript and performed the analyses. Priscila Rudiak Lustosa helped collect field data. Maria LuisaTunes Buschini and Cyntia Fernandes Pinto da Luz were responsible for the direction and correction of the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Araújo, J.L.O.; Quirino, Z.G.M.; Gandelha Neto, P.C.; Araújo, A.C. Síndromes de polinização ocorrentes em uma área de Mata Atlântica, Paraíba, Brasil. Biotemas 2009, 22, 83–94. [Google Scholar] [CrossRef]
- Torezan-Silingard, H.M. Flores e animais: Uma introdução à história natural dapolinização. In Eologia das Interações Plantas-Animais: Uma Abordagem Ecológico-Evolutiva; Technical Books Editora: Rio de Janeiro, Brazil, 2012; pp. 111–139. [Google Scholar]
- Buchmann, S.L.; Hurley, J.P. A biophysical model for buzz pollination in angiosperms. J. Theor. Biol. 1978, 72, 639–657. [Google Scholar] [CrossRef]
- Olmstead, R.; Palmer, J.D. Implications for the phylogeny, classification, and biogeography of Solanum from cpDNA restriction site variation. Syst. Bot. 1997, 22, 19–29. [Google Scholar] [CrossRef]
- Slaa, E.J.; Sánchez Chaves, L.A.; Malagodi-Braga, K.S.; Hofstede, F.E. Stingless bees in applied pollination: Practice and perspectives. Apidologie 2006, 37, 293–315. [Google Scholar] [CrossRef]
- Buchmann, S.L. Buzz pollination in angiosperms. In Handbook of Experimental Pollination Biology; Van Nostrand Reinhold Company: New York, NY, USA, 1983; pp. 63–113. [Google Scholar]
- Smith, S.D.; Knapp, S. The natural history of reproduction in Solanum and Lycianthes (Solanaceae) in a subtropical moist forest. Bull. Nat. Hist. Mus. Lond. (Bot.) 2002, 32, 125–136. [Google Scholar] [CrossRef]
- Tabarelli, M.; Mantovani, W.; Peres, C.A. Effects of habitat fragmentation on Plant guild structure in the montane Atlantic forest of southeastern Brazil. Biol. Conserv. 1999, 91, 119–127. [Google Scholar] [CrossRef]
- Oliveira-Filho, A.T.; Fontes, M.A.L. Patterns of Floristic Differentiation among Atlantic Forests in Southeastern Brazil and the Influence of Climate. Biotropica 2000, 32, 793–810. [Google Scholar] [CrossRef]
- Duarte, L.S.; Dillenburg, L.R. Ecophysiological responses of Araucaria angustifolia (Araucariaceae) seedlings to different irradiance levels. Aust. J. Bot. 2000, 48, 531–537. [Google Scholar] [CrossRef]
- Rambo, B.O. elemento andino pinhal rio-grandense. Anais Bot. Herb. “Barbosa Rodrigues” 1951, 3, 3–39. [Google Scholar]
- Waechter, J.L. Padrões fitogeográficos na flora atual do Rio Grande do Sul. Ciênc. Amb. 2002, 24, 93–108. [Google Scholar]
- Smith, L.B.; Downs, R. Solanaceas. Flora Ilustrada Catarinense; Sola: Itajaí, Brazil, 1966. [Google Scholar]
- Jordano, P.; Bascompte, J.; Olesen, J.M. Invariant properties in coevolutionary networks of plant-animal interactions. Ecol. Lett. 2003, 6, 69–81. [Google Scholar] [CrossRef]
- Petanidou, T.; Kallimanis, A.S.; Tzanopoulos, J.; Sgardelis, S.P.; Pantis, J.D. Long-term observation of a pollination network: Fluctuation in species and interactions, relative invariance of network structure and implications for estimates of specialization. Ecol. Lett. 2008, 11, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Jordano, P.; Bascompte, J.; Olesen, J.M. The ecological consequences of complex topology and nested structure in pollination webs. In Plant–Pollinator Interactions, from Specialization to Generalization; University of Chicago Press: Chicago, IL, USA, 2006; pp. 173–199. [Google Scholar]
- Secretaria Municiapal do Meio Ambiente—SEMAFLOR. Prefeitura Municipal de Guarapuava. Available online: htpp://www.prefeituramunicipaldeguarapuava (accessed on 7 March 2013).
- Maack, R. Geografia física do Estado do Paraná, 2nd ed.; José Olympio/Sec. da cultura e do esporte do Governo do Estado do Paraná: Rio de Janeiro, Brazil, 1981.
- Erdtman, G. The acetolizes method: A revised description. Svensk Bot. Tidskr. 1960, 54, 561–564. [Google Scholar]
- Vergeron, P. Interprétation statistique des resultants en matièred’analyse pollinique des miels. Ann. Abeille 1964, 7, 349–364. [Google Scholar] [CrossRef]
- Silveira Neto, S.; Nakano, O.; Barbin, D.; Villa Nova, N.A. Manual de Ecologia dos Insetos; Ceres: Piracicaba, Brazil, 1976. [Google Scholar]
- Palma, S. Contribuciónal studio de lossifonoforos encontrados frente a la costa de Valparaiso. In Memorias del II Simpósio Latino Americano Sobre Oceanografia Biológica; Universidad de Oriente: Cumaná, Venezuela, 1975; pp. 119–133. [Google Scholar]
- Jordano, P. Patterns of mutualistic interactions in pollination and seed dispersal: Connectance, dependence asymmetries and coevolution. Am. Nat. 1987, 129, 657–677. [Google Scholar] [CrossRef]
- Blüthgen, N. Why network analysis is often disconnected from community ecology: A critique and an ecologist’s guide. Basic Appl. Ecol. 2010, 11, 185–195. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 1–12. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Blüthgen, N.; Fründ, J.; Vázquez, D.; Menzel, F. What do interaction Network metrics tell us about specialization and biological traits? Ecology 2008, 89, 3387–3399. [Google Scholar]
- Almeida-Neto, M.; Guimarães, P.; Guimarães, P.R.; Loyola, R.D.; Ulrich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Guimarães, P.R.; Guimarães, P. Improving the analyses of nestedness for large sets of matrices. Environ. Model. Softw. 2006, 21, 1512–1513. [Google Scholar] [CrossRef]
- Bortoli, C.; Laroca, S. Melissocenologia no Terceiro Planalto Paranaense. I: Abundância relativa das abelhas silvestres (Apoidea) de um biótopo urbano de Guarapuava (PR, Brasil). Acta Biol. Parana. 1997, 26, 51–86. [Google Scholar] [CrossRef]
- Gonçalves, R.B.; Melo, G.A.R. A comunidade de abelhas (Hymenoptera, Apidae) em uma área restrita de campo natural no Parque Estadual de Vila Velha, Paraná: Diversidade, fenologia e fontes florais de alimento. Rev. Bras. Entomol. 2005, 49, 557–571. [Google Scholar] [CrossRef]
- Gonçalves, R.B.; Melo, G.A.R. A assembléia de abelhas (Hymenoptera, Apidae) de uma área restrita de campos naturais do Parque Estadual de Vila Velha, Paraná e comparações com áreas de campos e cerrado. Pap. Avulsos Zool. 2009, 49, 163–181. [Google Scholar]
- Freitas, L.; Sazima, M. Floral biology and pollination mechanisms in two Viola species-from nectar to pollen flowers? Ann. Bot. 2003, 91, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Freitas, L. Biologia da polinização em campos de altitude no Parque Nacional da Serra da Bocaina, SP. Ph.D. Thesis, Universidade Estadual de Campinas, São Paulo, Brazil, 2002. [Google Scholar]
- Aguiar, C.M.L. Utilização de recursos florais por abelhas (Hymenoptera, Apoidea) em uma área de Caatinga (Itatim, Bahia, Brasil). Rev. Bras. Zool. 2003, 20, 457–467. [Google Scholar] [CrossRef]
- Zillikens, A.; Steiner, S.; Mihalkó, Z. Nests of Augochlora (A.) esox in Bromeliads, a Previously Unknown site for Sweat Bees (Hymenoptera: Halictidae). Stud. Neotrop. Fauna Environ. 2001, 36, 137–142. [Google Scholar] [CrossRef]
- Moure, J.S.; Hurd, P.D. An Annotated Catalog of the Halictid Bees of the Western Hemisphere (Hymenoptera: Halictidae); Smithsonian Institution: Washington, DC, USA, 1987. [Google Scholar]
- Dalmazzo, M.; Vossler, F.G. Assessment of the pollen diet in a wood-dwelling augochlorine bee (Halictidae) using different approaches. Apidologie 2015, 46, 478–488. [Google Scholar] [CrossRef]
- Hesse, M. An exine architeture model for viscin threads. Grana 1984, 23, 69–75. [Google Scholar] [CrossRef]
- Cruden, R.W. Pollen-ovule ratio, pollen size and the ratio of stigmatic area to the pollen-bearing area of the pollinator: An hypothesis. Evolution 1981, 35, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Gimenes, M. Pollinating bees and other visitors of Ludwigia elegans (Onagraceae) flowers at a tropical site in Brazil. Stud. Neotrop. Fauna Environ. 1997, 32, 81–88. [Google Scholar] [CrossRef]
- Schlindwein, C. Are oligolectic bees always the most effective pollinators? In Solitary Bees. Conservation, Rearing and Management for Pollination; Imprensa Universitária: Fortaleza, CE, Brazil, 2004; pp. 231–240. [Google Scholar]
- Michener, C.D. Biogeography of the bees. Ann. Mo. Bot. Gard. 1979, 66, 277–347. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; The Johns Hopkins University Press: Baltimore, MD, USA, 2000. [Google Scholar]
- Gimenes, M.; Benedito-Silva, A.A.; Marques, M.D. Chronobiologic aspects of a coadaptive process: The interaction of Ludwigia elegans flowers and its more frequent bee visitors. Chronobiol. Int. 1993, 10, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Gimenes, M.; Benedito-Silva, A.A.; Marques, M.D. Circadian rhythms of pollen and nectar collection by bees on the flowers of Ludwigia elegans. Biol. Rhythm Res. 1996, 27, 281–290. [Google Scholar] [CrossRef]
- Gimenes, M. Interaction between visiting bees (Hymenoptera, Apoidea) and flowers of Ludwigia elegans (Camb.) Hara (Onagraceae) during the year in two different areas in São Paulo, Brazil. Braz. J. Biol. 2003, 63, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Buschini, M.L.T.; Rigon, J.; Cordeiro, J. Plants used by Megachile (Moureapis) sp. (Hymenoptera: Megachilidae) in the provisioning of their nests. Braz. J. Biol. 2009, 69, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, L.; Buschini, M.L.T. Bees diversity and their interaction networks with Ludwigia sericea (Cambessides) H. Hara and Ludwigia peruviana (L.) H. Hara (Onagraceae) flowers in Atlantic Forest Area in Southern Brazil. Sociobiology 2017, 64, 57–68. [Google Scholar] [CrossRef]
- Aguiar, C.M.L.; Garófalo, A.G. Nesting biology of Centris (Hemisiella) tarsata Smith (Hymenoptera, Apidae, Centridini). Rev. Bras. Zool. 2004, 21, 477–486. [Google Scholar] [CrossRef]
- Buschini, M.L.T. Species diversity and community structure in trap-nesting bees in Southern Brazil. Apidologie 2006, 37, 58–66. [Google Scholar] [CrossRef]
- Buschini, M.L.T.; Wolff, L.L. Nesting biology of Centris (Hemisiella) tarsata Smith in SouthernBrazil (Hymenoptera, Apidae, Centridini). Braz. J. Biol. 2006, 66, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, L.; Silva, C.I.; Buschini, M.L.T. Collection of Pollen Grains by Centris (Hemisiella) tarsata Smith (Apidae: Centridini): Is C. tarsata an Oligolectic or Polylectic Species? Zool. Stud. 2012, 51, 195–203. [Google Scholar]
- Diniz, M.E.R.; Buschini, M.L.T. Pollen analysis and interaction networks of floral visitor bees of Eugenia uniflora L. (Myrtaceae), in Atlantic Forest areas in southern Brazil. Arthropod Plant Interact. 2015, 9, 623–632. [Google Scholar] [CrossRef]
- Diniz, M.E.R.; Buschini, M.L.T. Diversity of flower visiting bees of Eugenia uniflora L. (Myrtaceae) in fragments of Atlantic Forest in South Brazil. Sociobiology 2016, 63, 982–990. [Google Scholar] [CrossRef]
- Antonini, Y.; Martins, R.P. The Flowering-Visiting Bees at the Ecological Station of the Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil. Neotrop. Entomol. 2003, 32, 565–575. [Google Scholar] [CrossRef]
- Imperatriz-Fonseca, V.L.; Santos, I.A.; Santos-Filho, P.S.; Engels, W.; Ramalho, M.; Wilms, W.; Aguilar, J.B.; Pinheiro-Machado, C.A.; Alves, D.A.; Kleinert, A.M.P. Checklist das abelhas e plantas melitófilas no Estado de São Paulo, Brasil. Biota Neotrop. 2011, 11, 632–655. [Google Scholar] [CrossRef]
- Santos, G.M.; Aguiar, C.M.L.; Mello, M.A.R. Flower-visiting guild associated with the Caatinga flora: Trophic interaction networks formed by social bees and social wasps with plants. Apidologie 2010, 41, 466–475. [Google Scholar] [CrossRef]
- Santos, K.C.B.S.; Gonçalvez, A.B.; Cereda, M.P. Polens importantes na flora apícola em uma região de Cerrado em Campo Grande-MS. Rev. Biol. Neotrop. 2015, 12, 81–85. [Google Scholar] [CrossRef]
- Faria-Mucci, G.M.; Melo, M.A.; Campos, L.A.O. A fauna de abelhas (Hymenoptera, Apoidea) e plantas utilizadas como fonte de recursos florais, em um ecossistema de campos rupestris em Lavras Novas, Minas Gerais, Brasil. In Apoidea Neotropica: Homenagem aos 90 Anos de Jesus Santiago Moure; Editora UNESC: Criciúma, SC, Brazil, 2003; pp. 241–256. [Google Scholar]
- Sakagami, S.F.; Laroca, S.; Moure, J.S. Wild bees biocenotics in São José dos Pinhais (PR), South Brazil. Preliminary report. J. Fac. Sci. Hokkaido Univ. Ser. VI Zool. 1967, 16, 253–291. [Google Scholar]
- Barros, M.A.G. Sistemas reprodutivos e polinizac: Ao em especies simpatricas de Erythroxylum P. Br. (Erythroxylaceae) do Brasil. Rev. Bras. Bot. 1998, 21, 159–166. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).