
Hyperthermal Reactions in DNA Triggered by 1–20 eV Electrons: Absolute Cross Sections for Crosslinks, Strand Breaks, Clustered Damages and Base Modifications

 and
and
Abstract
1. Introduction
2. Incorporating LEE Cross Sections into Monte Carlo Codes
3. Mathematical Model to Generate Absolute DNA Damage Cross Sections from Effective Yields
4. Results and Discussion
4.1. ACSs for DNA Damages Induced by 1–20 eV LEEs
4.2. Comparison of Damage ACSs for DNA to Those of Arg-DNA Complexes
4.3. Hyperthermal Reactions in DNA Triggered by 1–20 eV Electrons
5. Summary
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACS | Absolute cross section |
| Arg-DNA | Arginine-DNA |
| BD | Base damage |
| BD-CLs | Base damage related CLs |
| BER | Base excision repair |
| CL | Crosslink |
| CS | Cross section |
| DEA | Dissociative electron attachment |
| DSB | Double strand break |
| HER | High energy radiation |
| LEE | Low-energy electron |
| LS | Loss of supercoiled |
| MC | Monte Carlo |
| NDCD | Non-DSB cluster damages |
| PF | Protection factor |
| SE | Secondary electron |
| SSB | Single strand break |
| TA | Transient anion |
| OND | Other non-detected |
References
- Zheng, Y.; Sanche, L. Effective and absolute cross sections for low-energy (1–30 eV) electron interactions with condensed biomolecules. Appl. Phys. Rev. 2018, 5, 021302. [Google Scholar] [CrossRef]
- Lafosse, A.; Bertin, M.; Azria, R. Electron driven processes in ices: Surface functionalization and synthesis reactions. Prog. Surf. Sci. 2009, 84, 177–198. [Google Scholar] [CrossRef]
- Leung, L.; Lim, T.; Polanyi, J.C.; Hofer, W.A. Molecular calipers control atomic separation at a metal surface. Nano Lett. 2011, 11, 4113–4117. [Google Scholar] [CrossRef]
- Zhang, Y.; He, S.; Guo, W.; Hu, Y.; Huang, J.; Mulcahy, J.R.; Wei, W.D. Surface-plasmon-driven hot electron photochemistry. Chem. Rev. 2018, 118, 2927–2954. [Google Scholar] [CrossRef] [PubMed]
- Bohler, E.; Warneke, J. Control of chemical reactions and synthesis by low-energy electrons. Chem. Soc. Rev. 2013, 42, 9219–9231. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Z.; Hu, J.; Tian, S.X. Electron impact with the liquid-vapor interface. Accounts Chem. Res. 2022, 55, 3071–3079. [Google Scholar] [CrossRef]
- Kundu, S.; Prabhudesai, V.S.; Krishnakumar, E. Electron induced reactions in condensed mixtures of methane and ammonia. Phys. Chem. Chem. Phys. 2017, 19, 25723–25733. [Google Scholar] [CrossRef]
- Bespalov, I.; Zhang, Y.; Haitjema, J.; Tromp, R.M.; van der Molen, S.J.; Brouwer, A.M.; Jobst, J.; Castellanos, S. Key role of very low energy electrons in tin-based molecular resists for extreme ultraviolet nanolithography. ACS Appl. Mater. Interfaces 2020, 12, 9881–9889. [Google Scholar] [CrossRef]
- Roman, B.J.; Shubert-Zuleta, S.A.; Milliron, D.J. Tunable optical response of plasmonic metal oxide nanocrystals. MRS Bull. 2024, 49, 1032–1044. [Google Scholar] [CrossRef]
- Sanche, L. Electronic aging and related electron interactions in thin-film dielectrics. IEEE Trans. Electr. Insul. 1993, 28, 789–819. [Google Scholar] [CrossRef]
- Martel, R.; Avouris, P.; Lyo, I.W. Molecularly adsorbed oxygen species on Si(111)-(7×7): STM-Induced dissociative attachment studies. Science 1996, 272, 385–388. [Google Scholar] [CrossRef]
- Sloan, P.A.; Palmer, R.E. Two-electron dissociation of single molecules by atomic manipulation at room temperature. Nature 2005, 434, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.D.; Wang, C.; Fredin, L.A.; Lin, P.A.; Shimomoto, L.; Lezec, H.J.; Sharma, R. Site-selective CO disproportionation mediated by localized surface plasmon resonance excited by electron beam. Nat. Mater. 2019, 18, 614–619. [Google Scholar] [CrossRef]
- Zhou, J.; Wei, T.; An, X. Combining non-thermal plasma technology with photocatalysis: A critical review. Phys. Chem. Chem. Phys. 2023, 25, 1538–1545. [Google Scholar] [CrossRef] [PubMed]
- Hayes Griffith, O.; Engel, W. Historical perspective and current trends in emission microscopy, mirror electron microscopy and low-energy electron microscopy: An introduction to the proceedings of the second international symposium and workshop on emission microscopy and related techniques. Ultramicroscopy 1991, 36, 1–28. [Google Scholar]
- Di, W.; Rowntree, P.; Sanche, L. Energy-selective reaction of the hydrogen-passivated Si surface with carbon tetrafluoride via dissociative electron attachment. Phys. Rev. B 1995, 52, 16618–16622. [Google Scholar] [CrossRef]
- Hla, S.; Bartels, L.; Meyer, G.; Rieder, K. Inducing all steps of a chemical reaction with the scanning tunneling microscope tip: Towards single molecule engineering. Phys. Rev. Lett. 2000, 85, 2777–2780. [Google Scholar] [CrossRef]
- Arumainayagam, C.R.; Lee, H.; Nelson, R.B.; Haines, D.R.; Gunawardane, R.P. Low-energy electron induced reactions in condensed matter. Surf. Sci. Rep. 2010, 65, 1–44. [Google Scholar] [CrossRef]
- Jurczyk, J.; Brewer, C.R.; Hawkins, O.M.; Polyakov, M.N.; Kapusta, C.; McElwee-White, L.; Utke, I. Focused electron beam-induced deposition and post-growth purification using the heteroleptic Ru complex (η3-C3H5)Ru(CO)3Br. ACS Appl. Mater. Interfaces 2019, 11, 28164–28171. [Google Scholar] [CrossRef]
- Thorman, R.M.; Matsuda, S.J.; McElwee-White, L.; Fairbrother, D.H. Identifying and rationalizing the differing surface reactions of low-energy electrons and ions with an organometallic precursor. J. Phys. Chem. Lett. 2020, 11, 2006–2013. [Google Scholar] [CrossRef]
- Utke, I.; Swiderek, P.; Höflich, K.; Madajska, K.; Jurczyk, J.; Martinović, P.; Szymańska, I.B. Coordination and organometallic precursors of group 10 and 11: Focused electron beam induced deposition of metals and insight gained from chemical vapour deposition, atomic layer deposition, and fundamental surface and gas phase studies. Coord. Chem. Rev. 2022, 458, 213851. [Google Scholar] [CrossRef]
- Pimblott, S.M.; Laverne, J.A. Production of low-energy electrons by ionizing radiation. Radiat. Phys. Chem. 2007, 76, 1244–1247. [Google Scholar] [CrossRef]
- Cobut, V.; Frongillo, Y.; Patau, J.P.; Goulet, T.; Fraser, M.J.; Jay-Gerin, J.P. Monte Carlo simulation of fast electron and proton tracks in liquid water. Radiat. Phys. Chem. 1998, 51, 229–243. [Google Scholar]
- Lu, Q.B.; Sanche, L. Effects of cosmic rays on atmospheric chlorofluorocarbon dissociation and ozone depletion. Phys. Rev. Lett. 2001, 87, 078501. [Google Scholar] [CrossRef]
- Bennett, C.J.; Kaiser, R.I. On the formation of glycolaldehyde (HCOCH2OH) and methyl formate (HCOOCH3) in interstellar ice analogs. Astrophys. J. 2007, 661, 899–909. [Google Scholar] [CrossRef]
- Esmaili, S.; Bass, A.D.; Cloutier, P.; Sanche, L.; Huels, M.A. Glycine formation in CO2:CH4:NH3 ices induced by 0–70 eV electrons. J. Chem. Phys. 2018, 148, 164702. [Google Scholar] [CrossRef]
- Sajeev, Y. Prebiotic chemical origin of biomolecular complementarity. Commun. Chem. 2023, 6, 259. [Google Scholar] [CrossRef]
- Wu, Q.T.; Anderson, H.; Watkins, A.K.; Arora, D.; Barnes, K.; Padovani, M.; Shingledecker, C.N.; Arumainayagam, C.R.; Battat, J. Role of low-energy (<20 eV) secondary electrons in the extraterrestrial synthesis of prebiotic molecules. ACS Earth Space Chem. 2024, 8, 79–88. [Google Scholar]
- Wu, C.; Makiuchi, Y.; Chen, C. High-energy electron beam lithography for nanoscale fabrication. In Lithography; Wang, M., Ed.; InTech: Rijeka, Croatia, 2010; pp. 241–267. [Google Scholar]
- Tebyani, A.; Schramm, S.; Hesselberth, M.; Boltje, D.; Jobst, J.; Tromp, R.; van der Molen, S. Low energy electron microscopy at cryogenic temperatures. Ultramicroscopy 2023, 253, 113815. [Google Scholar] [CrossRef]
- Zhao, J.; Yu, X.; Shentu, X.; Li, D. The application and development of electron microscopy for three-dimensional reconstruction in life science: A review. Cell Tissue Res. 2024, 396, 1–18. [Google Scholar] [CrossRef]
- Penabeï, S.; Meesungnoen, J.; Jay-Gerin, J.-P. Assessment of cystamine’s radioprotective/antioxidant ability under high-dose-rate irradiation: A Monte Carlo multi-track chemistry simulation study. Antioxidants 2023, 12, 776. [Google Scholar] [CrossRef]
- Bepari, M.I.; Meesungnoen, J.; Jay-Gerin, J.-P. Early and transient formation of highly acidic pH spikes in water radiolysis under the combined effect of high dose rate and high linear energy transfer. Radiation 2023, 3, 165–182. [Google Scholar] [CrossRef]
- Sedmidubska, B.; Kocisek, J. Interaction of low-energy electrons with radiosensitizers. Phys. Chem. Chem. Phys. 2024, 26, 9112–9136. [Google Scholar] [CrossRef] [PubMed]
- Jay-Gerin, J.-P. Fundamentals of Water Radiolysis. Encyclopedia 2025, 5, 38. [Google Scholar] [CrossRef]
- Alizadeh, E.; Sanche, L. Precursors of solvated electrons in radiation biology. Chem. Rev. 2012, 112, 5578–5602. [Google Scholar] [CrossRef]
- Freneau, A.; Santos, M.; Voisin, P.; Tang, N.; Vizcarra, M.B.; Villagrasa, C.; Roy, L.; Vaurijoux, A.; Gruel, G. Relation between DNA double-strand breaks and energy spectra of secondary electrons produced by different X-ray energies. Int. J. Radiat. Biol. 2018, 94, 1075–1084. [Google Scholar] [CrossRef]
- Sridharan, D.; Chappell, L.; Whalen, M.; Cucinotta, F.; Pluth, J. Defining the biological effectiveness of components of high-LET track structure. Radiat. Res. 2015, 184, 105–119. [Google Scholar] [CrossRef]
- Kamanin, D.; Matthies, A.; Kormilitsyn, M. New approach to MA transmutation based on low-energy electron accelerators. In Atoms for Prosperity: Updating Eisenhowers Global Vision for Nuclear Energy; American Nuclear Society: New Orleans, LA, USA, 2003; pp. 620–621. [Google Scholar]
- Wang, X.L.; Xu, Z.Y.; Luo, W.; Lu, H.Y.; Zhu, Z.C.; Yan, X.Q. Transmutation prospect of long-lived nuclear waste induced by high-charge electron beam from laser plasma accelerator. Phys. Plasmas 2017, 24, 093105. [Google Scholar] [CrossRef]
- Siragusa, M.; Baiocco, G.; Fredericia, P.; Friedland, W.; Groesser, T.; Ottolenghi, A.; Jensen, M. The COOLER code: A novel analytical approach to calculate subcellular energy deposition by internal electron emitters. Radiat. Res. 2017, 188, 204–220. [Google Scholar] [CrossRef]
- van Dorp, W.F.; Beyer, A.; Mainka, M.; Golzhauser, A.; Hansen, T.W.; Wagner, J.B.; Hagen, C.W.; De Hosson, J. Focused electron beam induced processing and the effect of substrate thickness revisited. Nanotechnology 2013, 24, 345301. [Google Scholar] [CrossRef]
- Rezaee, M.; Hill, R.; Jaffray, D. The exploitation of low-energy electrons in cancer treatment. Radiat. Res. 2017, 188, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Goodhead, D.T.; O’Neill, P.; Menzel, H.G. Microdosimetry: An Interdisciplinary Approach; The Royal Society of Chemistry Press: Cambridge, UK, 1997. [Google Scholar]
- Ptasinska, S.; Varella, M.; Khakoo, M.A.; Slaughter, D.S.; Denifl, S. Electron scattering processes: Fundamentals, challenges, advances, and opportunities. Eur. Phys. J. D 2022, 76, 179. [Google Scholar] [CrossRef]
- Narayanan, J.; Tripathi, D.; Verma, P.; Adhikary, A.; Dutta, A.K. Secondary electron attachment-induced radiation damage to genetic materials. ACS Omega 2023, 8, 10669–10689. [Google Scholar] [CrossRef]
- Alizadeh, E.; Ptasińska, S.; Sanche, L. Transient anions in radiobiology and radiotherapy: From gaseous biomolecules to condensed organic and biolomolecular solids. In Radiation Effects in Materials; Monteiro, W.A., Ed.; IntechOpen: London, UK, 2016; pp. 179–231. [Google Scholar]
- International Atomic Energy Agency. Nuclear Data Section. Atomic and Molecular Data for Radiotherapy and Radiation Research: Final Report of a Co-Ordinated Research Programme; IAEA-TECDOC-799; IAEA: Vienna, Austria, 1995. [Google Scholar]
- Rogers, D. Fifty years of Monte Carlo simulations for medical physics. Phys. Med. Biol. 2006, 51, R287–R301. [Google Scholar] [CrossRef]
- Rucinski, A.; Biernacka, A.; Schulte, R. Applications of nanodosimetry in particle therapy planning and beyond. Phys. Med. Biol. 2021, 66, 24TR01. [Google Scholar] [CrossRef]
- Kyriakou, I.; Sakata, D.; Tran, H.N.; Perrot, Y.; Shin, W.G.; Lampe, N.; Zein, S.; Bordage, M.C.; Guatelli, S.; Villagrasa, C.; et al. Review of the Geant4-DNA simulation toolkit for radiobiological applications at the cellular and DNA Level. Cancers 2021, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Plante, I. A review of simulation codes and approaches for radiation chemistry. Phys. Med. Biol. 2021, 66, 03TR02. [Google Scholar] [CrossRef]
- Matsuya, Y.; Kai, T.; Sato, T.; Ogawa, T.; Hirata, Y.; Yoshii, Y.; Parisi, A.; Liamsuwan, T. Track-structure modes in particle and heavy ion transport code system (PHITS): Application to radiobiological research. Int. J. Radiat. Biol. 2022, 98, 148–157. [Google Scholar] [CrossRef]
- Hahn, M.B. Accessing radiation damage to biomolecules on the nanoscale by particle-scattering simulations. J. Phys. Commun. 2023, 7, 042001. [Google Scholar] [CrossRef]
- Goodhead, D.T.; Weinfeld, M. Clustered DNA damage and its complexity: Tracking the history. Radiat. Res. 2024, 202, 385–407. [Google Scholar] [CrossRef]
- Taleei, R.; Rahmanian, S.; Nikjoo, H. Modelling cellular response to ionizing radiation: Mechanistic, semi-mechanistic, and phenomenological approaches—A historical perspective. Radiat. Res. 2024, 202, 143–160. [Google Scholar] [CrossRef]
- Dong, Y.; Gao, Y.; Liu, W.; Gao, T.; Zheng, Y.; Sanche, L. Clustered DNA damage induced by 2–20 eV electrons and transient anions: General mechanism and correlation to cell death. J. Phys. Chem. Lett. 2019, 10, 2985–2990. [Google Scholar] [CrossRef]
- Gao, Y.; Zheng, Y.; Sanche, L. Low-Energy electron damage to condensed-phase DNA and its constituents. Int. J. Mol. Sci. 2021, 22, 7879. [Google Scholar] [CrossRef] [PubMed]
- Boulanouar, O.; Fromm, M.; Bass, A.D.; Cloutier, P.; Sanche, L. Absolute cross section for loss of supercoiled topology induced by 10 eV electrons in highly uniform /DNA/1,3-diaminopropane films deposited on highly ordered pyrolitic graphite. J. Chem. Phys. 2013, 139, 055104. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Chen, S.; Dong, Y.; Cloutier, P.; Zheng, Y.; Sanche, L. Absolute cross-sections for DNA strand breaks and crosslinks induced by low energy electrons. Phys. Chem. Chem. Phys. 2016, 18, 32762–32771. [Google Scholar] [CrossRef]
- Brodeur, N.; Cloutier, P.; Bass, A.D.; Bertrand, G.; Hunting, D.J.; Grandbois, M.; Sanche, L. Absolute cross section for DNA damage induced by low-energy (10 eV) electrons: Experimental refinements and sample characterization by AFM. J. Chem. Phys. 2018, 149, 164904. [Google Scholar] [CrossRef] [PubMed]
- Boguszewska, K.; Szewczuk, M.; Kaźmierczak-Barańska, J.; Karwowski, B.T. The similarities between human mitochondria and bacteria in the context of structure, genome, and base excision repair system. Molecules 2020, 25, 2857. [Google Scholar] [CrossRef]
- Rezaee, M.; Cloutier, P.; Bass, A.D.; Michaud, M.; Hunting, D.J.; Sanche, L. Absolute cross section for low-energy-electron damage to condensed macromolecules: A case study of DNA. Phys. Rev. E 2012, 86, 031913. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, Y.; Zhuang, P.; Fu, X.; Zheng, Y.; Sanche, L. Role of transient anions in chemoradiation therapy: Base modifications, cross-links, and cluster damages induced to cisplatin-DNA complexes by 1–20 eV electrons. J. Phys. Chem. B 2020, 120, 3315–3325. [Google Scholar] [CrossRef]
- Wang, X.; Liao, H.; Liu, W.; Shao, Y.; Zheng, Y.; Sanche, L. DNA protection against damages induced by low-energy electrons: Absolute cross sections for arginine—DNA complexes. J. Phys. Chem. Lett. 2023, 14, 5674–5680. [Google Scholar] [CrossRef]
- Nikjoo, H.; Emfietzoglou, D.; Liamsuwan, T.; Taleei, R.; Liljequist, D.; Uehara, S. Radiation track, DNA damage and response—A review. Rep. Prog. Phys. 2016, 79, 116601. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.G.; Bordage, M.C.; Emfietzoglou, D.; Kyriakou, I.; Sakata, D.; Min, C.H.; Lee, S.; Guatelli, S.; Incerti, S. Development of a new Geant4-DNA electron elastic scattering model for liquid-phase water using the ELSEPA code. J. Appl. Phys. 2018, 124, 224901. [Google Scholar] [CrossRef]
- Liamsuwan, T.; Emfietzoglou, D.; Uehara, S.; Nikjoo, H. Microdosimetry of low-energy electrons. Int. J. Radiat. Biol. 2012, 88, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Plante, I.; Cucinotta, F. Cross sections for the interactions of 1 eV–100 MeV electrons in liquid water and application to Monte-Carlo simulation of HZE radiation tracks. New J. Phys. 2009, 11, 063047. [Google Scholar] [CrossRef]
- Sanz, A.; Fuss, M.; Muñoz, A.; Blanco, F.; Limão-Vieira, P.; Brunger, M.; Buckman, S.; García, G. Modelling low energy electron and positron tracks for biomedical applications. Int. J. Radiat. Biol. 2012, 88, 71–76. [Google Scholar] [CrossRef]
- Kyriakou, I.; Emfietzoglou, D.; Ivanchenko, V.; Bordage, M.C.; Guatelli, S.; Lazarakis, P.; Tran, H.N.; Incerti, S. Microdosimetry of electrons in liquid water using the low-energy models of Geant4. J. Appl. Phys. 2017, 122, 024303. [Google Scholar] [CrossRef]
- Emfietzoglou, D.; Papamichaela, G.; Nikjoo, H. Monte Carlo electron track structure calculations in liquid water using a new model dielectric response function. Radiat. Res. 2017, 188, 355–368. [Google Scholar] [CrossRef]
- Margis, S.; Kyriakou, I.; Incerti, S.; Bordage, M.C.; Emfietzoglou, D. Sub-keV corrections to binary encounter cross section models for electron ionization of liquid water with application to the Geant4-DNA Monte Carlo code. Appl. Radiat. Isot. 2023, 194, 110693. [Google Scholar] [CrossRef]
- Tran, H.N.; Archer, J.; Baldacchino, G.; Brown, J.M.C.; Chappuis, F.; Cirrone, G.A.P.; Desorgher, L.; Dominguez, N.; Fattori, S.; Guatelli, S.; et al. Review of chemical models and applications in Geant4-DNA: Report from the ESA BioRad III Project. Med. Phys. 2024, 51, 5873–5889. [Google Scholar] [CrossRef]
- Bernal, M.A.; Bordage, M.C.; Brown, J.M.C.; Davídková, M.; Delage, E.; El Bitar, Z.; Enger, S.A.; Francis, Z.; Guatelli, S.; Ivanchenko, V.N.; et al. Track structure modeling in liquid water: A review of the Geant4-DNA very low energy extension of the Geant4 Monte Carlo simulation toolkit. Phys. Medica 2015, 31, 861–874. [Google Scholar] [CrossRef]
- Tanaka, H.; Brunger, M.J.; Campbell, L.; Kato, H.; Hoshino, M.; Rau, A.R.P. Scaled plane-wave Born cross sections for atoms and molecules. Rev. Mod. Phys. 2016, 88, 025004. [Google Scholar] [CrossRef]
- Thomson, R.M.; Kawrakow, I. Quantum versus classical Monte Carlo simulation of low-energy electron transport in condensed amorphous media. Phys. Medica 2018, 54, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.E.; Green, N.J.B.; Pimblott, S.M. Methods for the simulation of the slowing of low-energy electrons in water. J. Comput. Chem. 2018, 39, 2217–2225. [Google Scholar] [CrossRef]
- Chatzipapas, K.; Papadimitroulas, P.; Emfietzoglou, D.; Kalospyros, S.; Hada, M.; Georgakilas, A.G.; Kagadis, G.C. Ionizing radiation and complex DNA damage: Quantifying the radiobiological damage using Monte Carlo simulations. Cancers 2020, 12, 799. [Google Scholar] [CrossRef] [PubMed]
- de Vera, P.; Abril, I.; Garcia-Molina, R. Excitation and ionisation cross-sections in condensed-phase biomaterials by electrons down to very low energy: Application to liquid water and genetic building blocks. Phys. Chem. Chem. Phys. 2021, 23, 5079–5095. [Google Scholar] [CrossRef]
- Derksen, L.; Pfuhl, T.; Engenhart-Cabillic, R.; Zink, K.; Baumann, K. Investigating the feasibility of TOPAS-nBio for Monte Carlo track structure simulations by adapting GEANT4-DNA examples application. Phys. Med. Biol. 2021, 66, 175023. [Google Scholar] [CrossRef]
- Signorell, R. Electron scattering in liquid water and amorphous ice: A striking resemblance. Phys. Rev. Lett. 2020, 124, 205501. [Google Scholar] [CrossRef]
- Michaud, M.; Wen, A.; Sanche, L. Cross sections for low-energy (1–100 eV) electron elastic and inelastic scattering in amorphous ice. Radiat. Res. 2003, 159, 3–22. [Google Scholar] [CrossRef]
- Nikjoo, H.; O’Neill, P.; Goodhead, D.; Terrissol, M. Computational modelling of low-energy electroninduced DNA damage by early physical and chemical events. Int. J. Radiat. Biol. 1997, 71, 467–483. [Google Scholar] [CrossRef]
- Nikjoo, H.; Uehara, S.; Emfietzoglou, D.; Cucinotta, F.A. Track-structure codes in radiation research. Radiat. Meas. 2006, 41, 1052–1074. [Google Scholar] [CrossRef]
- Zhang, L.; Tan, Z. A new calculation on spectrum of direct DNA damage induced by low-energy electrons. Radiat. Environ. Biophys. 2010, 49, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Friedland, W.; Dingfelder, M.; Kundrát, P.; Peter Jacob, P. Track structures, DNA targets and radiation effects in the biophysical Monte Carlo simulation code PARTRAC. Mutat. Res. 2011, 711, 28–40. [Google Scholar] [CrossRef]
- Nikjoo, H.; Girard, P. A model of the cell nucleus for DNA damage calculations. Int. J. Radiat. Biol. 2012, 88, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.; Hung, T.; Tu, S.; Tung, C. Fast Monte Carlo simulation of DNA damage induction by Auger-electron emission. Int. J. Radiat. Biol. 2014, 90, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Pater, P.; Seuntjens, J.; Naqa, I. On the consistency of Monte Carlo track structure DNA damage simulations. Med. Phys. 2014, 41, 121708. [Google Scholar] [CrossRef]
- Madsen, R.; Akabani, G. Low-energy cross-section calculations of single molecules by electron impact: A classical Monte Carlo transport approach with quantum mechanical description. Phys. Med. Biol. 2014, 59, 2285–2305. [Google Scholar] [CrossRef]
- Bug, M.U.; Baek, W.Y.; Rabus, H.; Villagrasa, C.; Meylan, S.; Rosenfeld, A.B. An electron-impact cross section data set (10 eV–1 keV) of DNA constituents based on consistent experimental data: A requisite for Monte Carlo simulations. Radiat. Phys. Chem. 2017, 130, 459–479. [Google Scholar] [CrossRef]
- Sakata, D.; Belov, O.; Bordage, M.C.; Emfietzoglou, D.; Guatelli, S.; Inaniwa, T.; Ivanchenko, V.; Karamitros, M.; Kyriakou, I.; Lampe, N.; et al. Fully integrated Monte Carlo simulation for evaluating radiation induced DNA damage and subsequent repair using Geant4-DNA. Sci. Rep. 2020, 10, 20788. [Google Scholar] [CrossRef]
- Lamghari, Y.; Lu, H.; Bentourkia, M. DNA damage by radiation as a function of electron energy and interaction at the atomic level with Monte Carlo simulation. Z. Med. Physik 2023, 33, 489–498. [Google Scholar] [CrossRef]
- Rezaee, M.; Adhikary, A. The effects of particle LET and fluence on the complexity and frequency of clustered DNA damage. DNA 2024, 4, 34–51. [Google Scholar] [CrossRef]
- Liu, W.; Tan, Z.; Zhang, L.; Champion, C. Calculation on spectrum of direct DNA damage induced by low energy electrons including dissociative electron attachment. Radiat. Environ. Biophys. 2017, 56, 99–110. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Qiu, R.; Yan, C.; Xie, W.; Zeng, Z.; Tung, C. Comparison of direct DNA strand breaks induced by low energy electrons with different inelastic cross sections. Nucl. Instrum. Methods Phys. Res. B 2013, 311, 27–36. [Google Scholar] [CrossRef]
- Toburen, L. Challenges in Monte Carlo track structure modelling. Int. J. Radiat. Biol. 2012, 88, 2–9. [Google Scholar] [CrossRef]
- Caron, L.G.; Tonzani, S.; Greene, C.H.; Sanche, L. Diffraction in low-energy electron scattering from DNA: Bridging gas phase and solid-state theory. Phys. Rev. A 2008, 78, 042710. [Google Scholar] [CrossRef]
- Kai, T.; Toigawa, T.; Matsuya, Y.; Hirata, Y.; Tezuka, T.; Tsuchida, H.; Yokoya, A. Significant role of secondary electrons in the formation of a multi-body chemical species spur produced by water radiolysis. Sci. Rep. 2024, 14, 24722. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.Y.; Tian, Z.; Qin, N.; Yan, C.; Lai, Y.; Hung, S.H.; Chi, Y.; Jia, X. A new open-source GPU-based microscopic Monte Carlo simulation tool for the calculations of DNA damages caused by ionizing radiation—Part I: Core algorithm and validation. Med. Phys. 2020, 47, 1958–1970. [Google Scholar] [CrossRef]
- Zein, S.A.; Bordage, M.C.; Francis, Z.; Macetti, G.; Genoni, A.; Dal Cappello, C.; Shin, W.G.; Incerti, S. Electron transport in DNA bases: An extension of the Geant4-DNA Monte Carlo toolkit. Nucl. Instrum. Methods Phys. Res. B 2021, 488, 70–82. [Google Scholar] [CrossRef]
- Lemelin, V.; Bass, A.D.; Cloutier, P.; Sanche, L. Low energy (1–19 eV) electron scattering from condensed thymidine (dT) II: Comparison of vibrational excitation cross sections with those of tetrahydrofuran and the recalibrated values of thymine. Phys. Chem. Chem. Phys. 2019, 21, 23818–23825. [Google Scholar] [CrossRef]
- Caron, L.; Bouchiha, D.; Gorfinkiel, J.D.; Sanche, L. Adapting gas phase electron scattering R-matrix calculations to a condensed matter environment. Phys. Rev. A 2007, 76, 032716. [Google Scholar] [CrossRef]
- Seiwert, T.Y.; Salama, J.K.; Vokes, E.E. The concurrent chemoradiation paradigm—General principles. Nat. Clin. Pract. Oncol. 2007, 4, 86–100. [Google Scholar] [CrossRef]
- Hayes, A. Applications of nuclear physics. Rep. Prog. Phys. 2017, 80, 026301. [Google Scholar] [CrossRef]
- Ku, A.; Facca, V.J.; Cai, Z.; Reilly, R.M. Auger electrons for cancer therapy—A review. EJNMMI Radiopharm. Chem. 2019, 4, 27. [Google Scholar] [CrossRef]
- Chen, X.; Karmaker, N.; Cloutier, P.; Bass, A.D.; Zheng, Y.; Sanche, L. Low-energy electron damage to plasmid DNA in thin films: Dependence on substrates, surface density, charging, environment, and uniformity. J. Phys. Chem. B 2022, 126, 5443–5457. [Google Scholar] [CrossRef]
- Meesungnoen, J.; Jay-Gerin, J.P.; Filali-Mouhim, A.; Mankhetkorn, S. Low-energy electron penetration range in liquid water. Radiat. Res. 2002, 158, 657–660. [Google Scholar] [CrossRef]
- Powell, C.J.; Jablonski, A. Progress in quantitative surface analysis by X-ray photoelectron spectroscopy: Current status and perspectives. J. Electron Spectrosc. Relat. Phenom. 2010, 178–179, 331–346. [Google Scholar] [CrossRef]
- Cazaux, J. Mechanisms of charging in electron spectroscopy. J. Electron Spectrosc. Relat. Phenom. 1999, 105, 155–185. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Laval, J.; Boiteux, S. Substrate specificity of the Escherichia Coli endonuclease III: Excision of thymine- and cytosine-derived lesions in DNA produced by radiation-generated free radicals. Biochemistry 1993, 32, 12105–12111. [Google Scholar] [CrossRef]
- Seeberg, E.; Eide, L.; Bjørås, M. The base excision repair pathway. Trends Biochem. Sci. 1995, 20, 391–397. [Google Scholar] [CrossRef]
- Capecchi, M.R. Altering the genome by homologous recombination. Science 1989, 244, 1288–1292. [Google Scholar] [CrossRef]
- Sonoda, E.; Hochegger, H.; Saberi, A.; Taniguchi, Y.; Takeda, S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair. DNA Repair 2006, 5, 1021–1029. [Google Scholar] [CrossRef]
- Asaithamby, A.; Hu, B.; Chen, D.J. Unrepaired clustered DNA lesions induce chromosome breakage in human cells. Biol. Sci. 2011, 108, 8293–8298. [Google Scholar] [CrossRef]
- Martin, F.; Burrow, P.; Cai, Z.; Cloutier, P.; Hunting, D.; Sanche, L. DNA strand breaks induced by 0–4 eV electrons: The role of shape resonances. Phys. Rev. Lett. 2004, 93, 068101. [Google Scholar] [CrossRef]
- Shao, Y.; Dong, Y.; Hunting, D.; Zheng, Y.; Sanche, L. Unified mechanism for the generation of isolated and clustered DNA damages by a single low energy electron. J. Phys. Chem. C 2017, 121, 2466–2472. [Google Scholar] [CrossRef]
- Wityk, P.; Kostrzewa-Nowak, D.; Krawczyk, B.; Michalik, M.; Nowak, R. X-ray and UV radiation damage of dsDNA/protein complexes. Molecules 2021, 26, 3132. [Google Scholar] [CrossRef]
- Luscombe, N.M.; Laskowski, R.A.; Thornton, J.M. Amino acid-base interactions: A three-dimensional analysis of protein-DNA interactions at an atomic level. Nucleic. Acids. Res. 2001, 29, 2860–2874. [Google Scholar] [CrossRef]
- Cheng, A.C.; Frankel, A.D. Ab initio interaction energies of hydrogen-bonded amino acid side chain-nucleic acid base interactions. J. Am. Chem. Soc. 2004, 126, 434–435. [Google Scholar] [CrossRef]
- Barrios, R.; Skurski, P.; Simons, J. Mechanism for damage to DNA by low-energy electrons. J. Phys. Chem. B 2002, 106, 7991–7994. [Google Scholar] [CrossRef]
- Takayanagi, K. Scattering of slow electrons by molecules. Prog. Theor. Phys. Suppl. 1967, 40, 216–248. [Google Scholar] [CrossRef]
- Christophorou, L.G.; Anderson, V.E.; Birks, J.B. Atomic and molecular radiation physics. In Health Physics Division Annual Progress Report for Period Ending; Oak Ridge National Lab.: Oak Ridge, TN, USA, 1971. [Google Scholar]
- Zheng, Y.; Wagner, J.R.; Sanche, L. DNA damage induced by low-energy electrons: Electron transfer and diffraction. Phys. Rev. Lett. 2006, 96, 208101. [Google Scholar] [CrossRef]
- Kumari, B.; Huwaidi, A.; Robert, G.; Cloutier, P.; Bass, A.D.; Sanche, L.; Wagner, J.R. Shape resonances in DNA: Nucleobase release, reduction, and dideoxynucleoside products induced by 1.3 to 2.3 eV electrons. J. Phys. Chem. B 2022, 126, 5175–5184. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Jaruga, P. Mechanisms of free radical-induced damage to DNA. Free Radic. Res. 2012, 46, 382–419. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sevilla, M.D.; Sanche, L. How a single 5 eV electron can induce double-strand breaks in DNA: A time-dependent density functional theory study. J. Phys. Chem. B 2024, 128, 4053–4062. [Google Scholar] [CrossRef]
- Eccles, L.J.; O’Neill, P.; Lomax, M.E. Delayed repair of radiation induced clustered DNA damage: Friend or foe? Mutat. Res. 2011, 711, 134–141. [Google Scholar] [CrossRef]
- Vogel, A.; Noack, J.; Huttman, G.; Paltauf, G. Mechanisms of femtosecond laser nanosurgery of cells and tissues. Appl. Phys. B 2005, 81, 1015–1047. [Google Scholar] [CrossRef]
- Dharmadhikari, A.K.; Bharambe, H.; Dharmadhikari, J.A.; D’Souza, J.S.; Mathur, D. DNA damage by OH radicals produced using intense, ultrashort, long wavelength laser pulses. Phys. Rev. Lett. 2014, 112, 138105. [Google Scholar] [CrossRef]
- Schmalz, M.; Liang, X.X.; Wieser, I.; Gruschel, C.; Muskalla, L.; Stöckl, M.T.; Nitschke, R.; Linz, N.; Leitenstorfer, A.; Vogel, A.; et al. Dissection of DNA damage and repair pathways in live cells by femtosecond laser microirradiation and free-electron modeling. Proc. Natl. Acad. Sci. USA 2023, 120, e2220132120. [Google Scholar] [CrossRef] [PubMed]
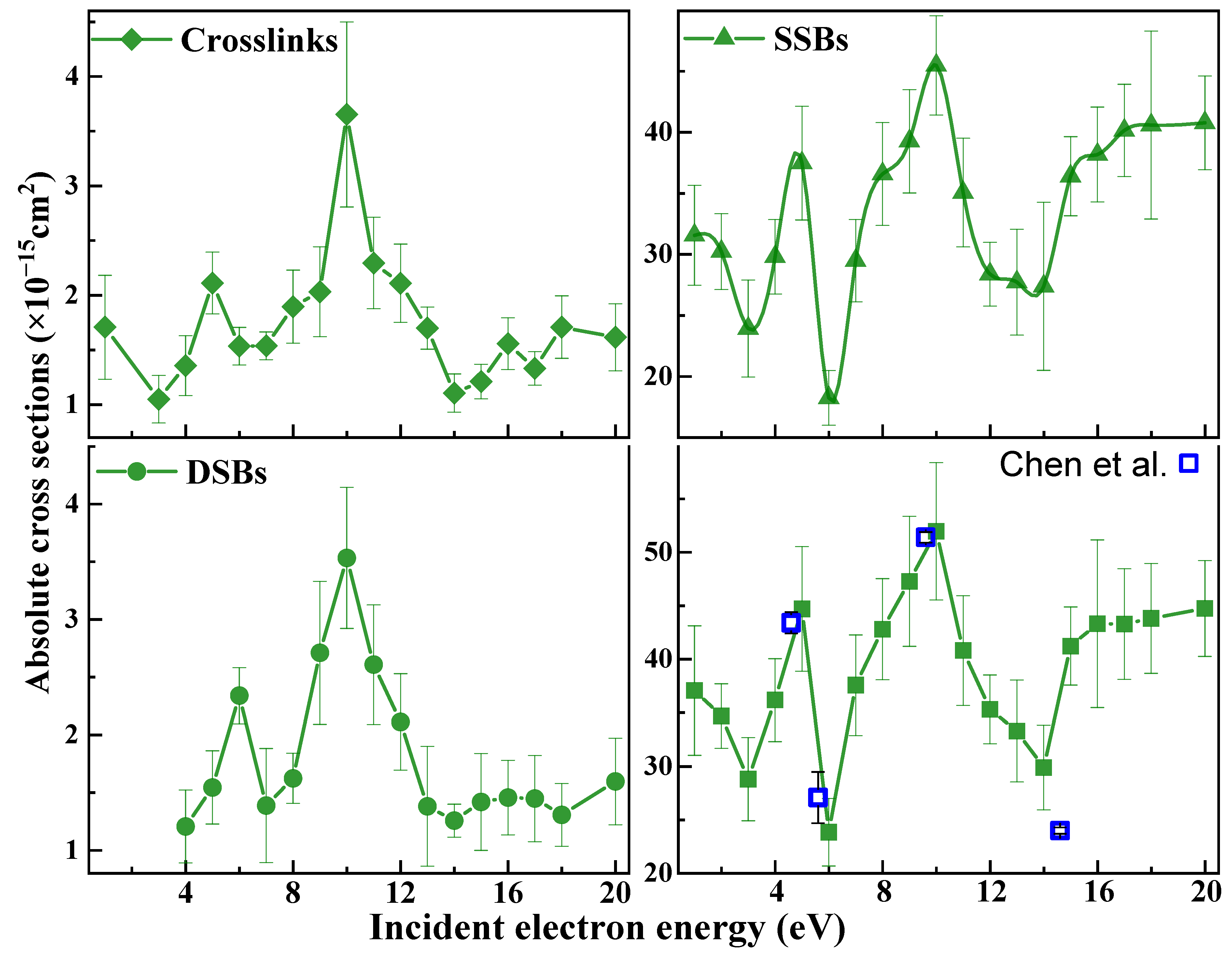
 ) induced by 1−20 eV electrons in 3197 base-pair plasmid DNA. The absolute values are generated from effective yields and a penetration factor f of 0.25 ± 0.02, which is obtained from the ratio of the effective cross section to the ACS (Equation (6)) for loss of the supercoiled configuration induced by 10 eV electron impact on 10, 15, and 20 nm plasmid DNA films [57,64]. The error bars arise principally from the standard deviations in the measured effective yields.
) induced by 1−20 eV electrons in 3197 base-pair plasmid DNA. The absolute values are generated from effective yields and a penetration factor f of 0.25 ± 0.02, which is obtained from the ratio of the effective cross section to the ACS (Equation (6)) for loss of the supercoiled configuration induced by 10 eV electron impact on 10, 15, and 20 nm plasmid DNA films [57,64]. The error bars arise principally from the standard deviations in the measured effective yields.
) induced by 1−20 eV electrons in 3197 base-pair plasmid DNA. The absolute values are generated from effective yields and a penetration factor f of 0.25 ± 0.02, which is obtained from the ratio of the effective cross section to the ACS (Equation (6)) for loss of the supercoiled configuration induced by 10 eV electron impact on 10, 15, and 20 nm plasmid DNA films [57,64]. The error bars arise principally from the standard deviations in the measured effective yields.
) induced by 1−20 eV electrons in 3197 base-pair plasmid DNA. The absolute values are generated from effective yields and a penetration factor f of 0.25 ± 0.02, which is obtained from the ratio of the effective cross section to the ACS (Equation (6)) for loss of the supercoiled configuration induced by 10 eV electron impact on 10, 15, and 20 nm plasmid DNA films [57,64]. The error bars arise principally from the standard deviations in the measured effective yields.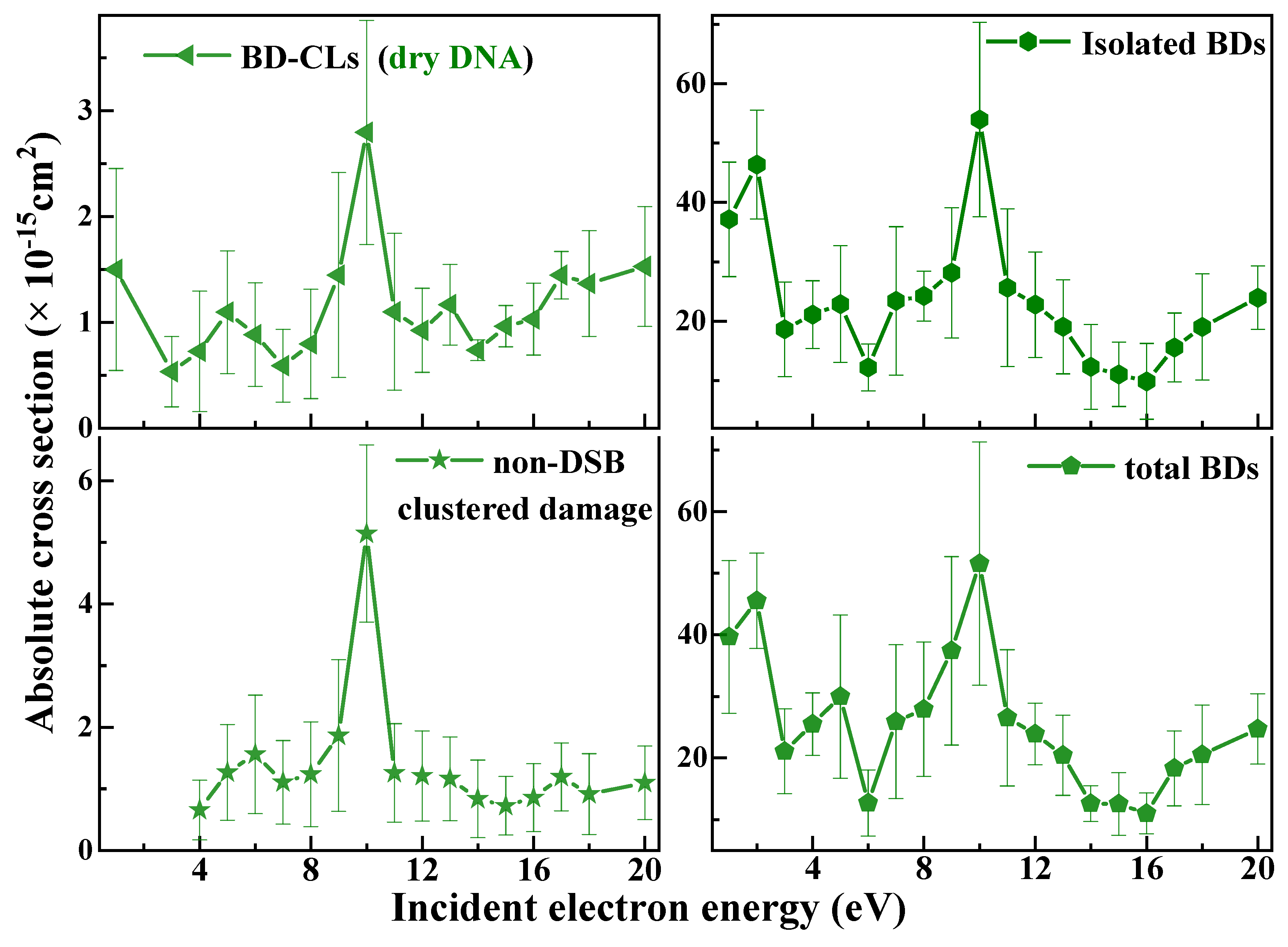
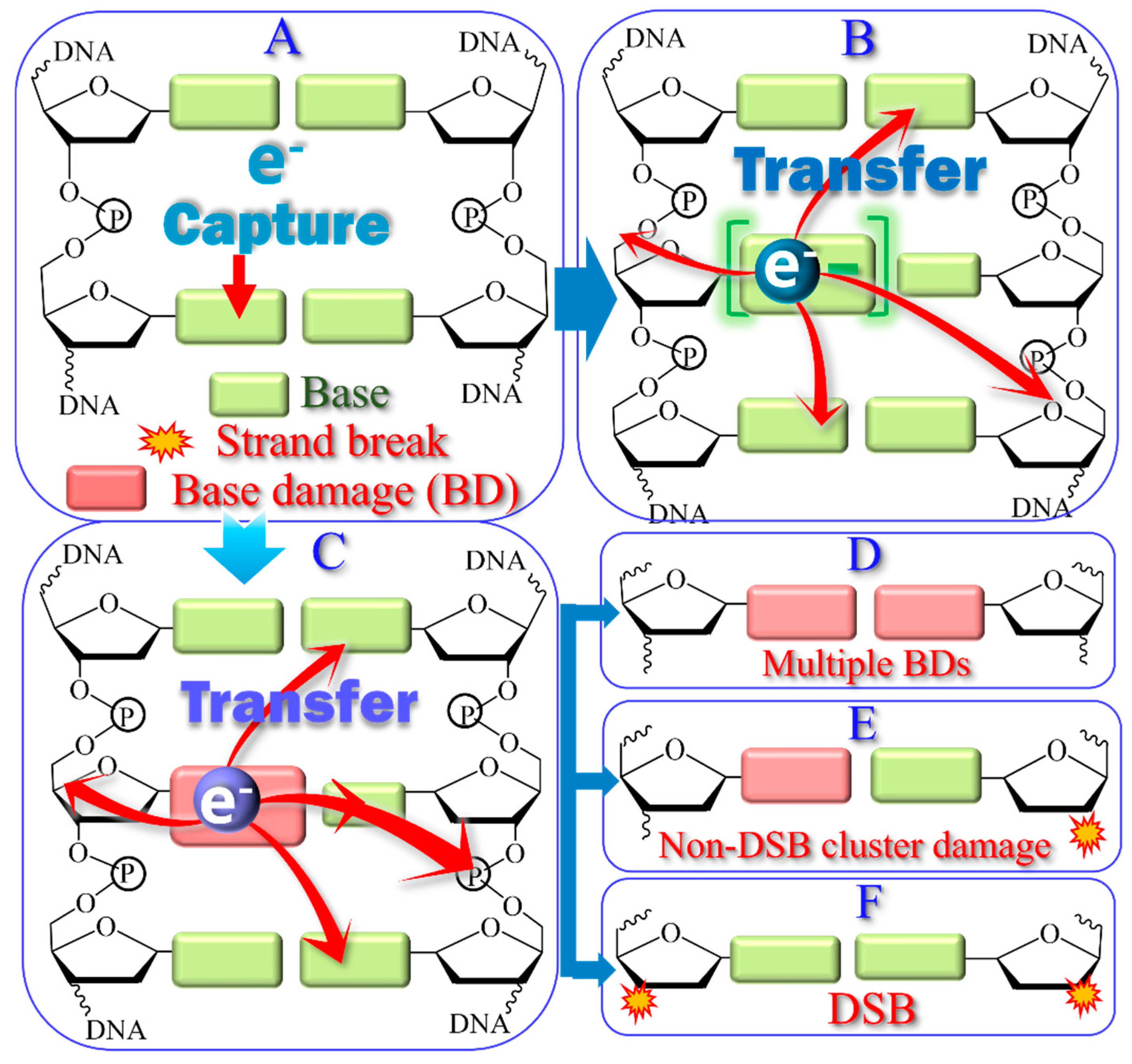

{kind=link}
{kind=link}
{kind=link}
| Energy (eV) | CLs | DSBs | SSBs | LS | BD-CLs | Non-DSB Clustered Damages | Isolated | Total BDs | Total DNA |
|---|---|---|---|---|---|---|---|---|---|
| BDs | Damages | ||||||||
| 1 | 1.7 ± 0.5 | n.d. | 31.6 ± 4.1 | 37.1 ± 6.1 | 1.5 ± 1.0 | n.d. | 37.1 ± 9.6 | 39.6 ± 12.4 | 76.7 ± 13.5 |
| 2 | n.d. | n.d. | 30.2 ± 3.1 | 34.7 ± 3.0 | n.d. | n.d. | 46.4 ± 9.2 | 45.5 ± 7.7 | 80.2 ± 9.4 |
| 3 | 1.0 ± 0.2 | n.d. | 23.9 ± 4.0 | 28.8 ± 3.9 | 0.5 ± 0.3 | n.d. | 18.7 ± 8.0 | 21.1 ± 6.9 | 49.9 ± 7.8 |
| 4 | 1.4 ± 0.3 | 1.2 ± 0.3 | 29.8 ± 3.1 | 36.2 ± 3.9 | 0.7 ± 0.6 | 0.7 ± 0.5 | 21.1 ± 5.7 | 25.5 ± 5.1 | 61.6 ± 6.8 |
| 5 | 2.1 ± 0.3 | 1.5 ± 0.3 | 37.5 ± 4.7 | 44.7 ± 5.8 | 1.1 ± 0.6 | 1.3 ± 0.8 | 22.9 ± 9.8 | 29.9 ± 13.3 | 74.6 ± 14.3 |
| 6 | 1.5 ± 0.2 | 2.3 ± 0.2 | 18.2 ± 2.2 | 23.8 ± 3.2 | 0.9 ± 0.5 | 1.6 ± 1.0 | 12.2 ± 3.9 | 12.7 ± 5.4 | 36.9 ± 5.5 |
| 7 | 1.5 ± 0.1 | 1.4 ± 0.5 | 29.5 ± 3.4 | 37.6 ± 4.7 | 0.6 ± 0.4 | 1.1 ± 0.7 | 23.4 ± 12.5 | 25.9 ± 12.5 | 65.5 ± 13.3 |
| 8 | 1.9 ± 0.3 | 1.6 ± 0.2 | 36.6 ± 4.2 | 42.8 ± 4.7 | 0.8 ± 0.5 | 1.2 ± 0.9 | 24.2 ± 4.2 | 27.9 ± 10.9 | 70.7 ± 12.1 |
| 9 | 2.0 ± 0.4 | 2.7 ± 0.6 | 39.3 ± 4.2 | 47.3 ± 6.1 | 1.5 ± 1.0 | 1.9 ± 1.2 | 28.2 ± 11.0 | 37.4 ± 15.3 | 84.7 ± 16.5 |
| 10 | 3.7 ± 0.8 | 3.5 ± 0.6 | 45.5 ± 4.1 | 52.0 ± 6.4 | 2.9 ± 1.1 | 5.1 ± 1.4 | 54.0 ± 16.4 | 51.5 ± 19.7 | 103.5 ± 21.0 |
| 11 | 2.3 ± 0.4 | 2.6 ± 0.5 | 35.1 ± 4.5 | 40.8 ± 5.1 | 1.1 ± 0.8 | 1.3 ± 0.8 | 25.6 ± 13.3 | 26.5 ± 11.1 | 67.3 ± 12.1 |
| 12 | 2.1 ± 0.4 | 2.1 ± 0.4 | 28.4 ± 2.6 | 35.3 ± 3.2 | 0.9 ± 0.4 | 1.2 ± 0.7 | 22.8 ± 8.9 | 23.9 ± 5.0 | 59.2 ± 6.6 |
| 13 | 1.7 ± 0.2 | 1.4 ± 0.5 | 27.7 ± 4.3 | 33.3 ± 4.7 | 1.2 ± 0.4 | 1.2 ± 0.7 | 19.1 ± 7.9 | 20.4 ± 6.5 | 54.6 ± 7.7 |
| 14 | 1.1 ± 0.2 | 1.3 ± 0.1 | 27.4 ± 6.9 | 29.9 ± 4.0 | 0.8 ± 0.1 | 0.8 ± 0.6 | 12.3 ±7.2 | 12.6 ± 2.9 | 42.5 ± 4.3 |
| 15 | 1.2 ± 0.2 | 1.4 ± 0.4 | 36.4 ± 3.2 | 41.2 ± 3.7 | 1.0 ± 0.2 | 0.7 ± 0.5 | 11.0 ± 5.4 | 12.5 ± 5.1 | 53.7 ± 6.6 |
| 16 | 1.6 ± 0.2 | 1.5 ± 0.3 | 38.2 ± 3.9 | 43.3 ± 7.8 | 1.1 ± 0.4 | 0.9 ± 0.6 | 9.9 ± 6.4 | 11.0 ± 3.3 | 54.3 ± 5.4 |
| 17 | 1.3 ± 0.2 | 1.4 ± 0.4 | 40.2 ± 3.8 | 43.3 ± 5.2 | 1.5 ± 0.3 | 1.2 ± 0.5 | 15.6 ± 5.8 | 18.3 ± 6.1 | 61.6 ± 7.7 |
| 18 | 1.7 ± 0.3 | 1.3 ± 0.3 | 40.6 ± 7.7 | 43.8 ± 5.1 | 1.4 ± 0.5 | 0.9 ± 0.7 | 19.0 ± 8.9 | 20.5 ± 8.1 | 55.3 ± 9.1 |
| 20 | 1.6 ± 0.3 | 1.6 ± 0.4 | 40.8 ± 3.8 | 44.7 ± 4.5 | 1.6 ± 0.6 | 1.1 ± 0.6 | 23.9 ± 5.3 | 24.7 ± 5.7 | 69.4 ± 7.7 |
| 5 * 10 * | 3.9 ± 0.6 | 1.4 ± 0.2 | 8.4 ± 0.7 | 17.3 ± 0.3 | 0.8 ± 0.1 | 0.6 ± 0.3 | 23.0 ± 0.3 | 26.5 ± 1.1 | 45.8 ± 0.7 |
| 5.7 ± 0.6 | 2.7 ± 0.8 | 12.3 ± 1.2 | 20.0 ± 1.0 | 4.3 ± 1.9 | 3.4 ± 2.0 | 28.2 ± 2.9 | 35.1 ± 2.3 | 57.5 ± 2.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Huang, X.; Zhang, W.; Shao, Y.; Cloutier, P.; Zheng, Y.; Sanche, L. Hyperthermal Reactions in DNA Triggered by 1–20 eV Electrons: Absolute Cross Sections for Crosslinks, Strand Breaks, Clustered Damages and Base Modifications. Int. J. Mol. Sci. 2025, 26, 4057. https://doi.org/10.3390/ijms26094057
Dong Y, Huang X, Zhang W, Shao Y, Cloutier P, Zheng Y, Sanche L. Hyperthermal Reactions in DNA Triggered by 1–20 eV Electrons: Absolute Cross Sections for Crosslinks, Strand Breaks, Clustered Damages and Base Modifications. International Journal of Molecular Sciences. 2025; 26(9):4057. https://doi.org/10.3390/ijms26094057
Chicago/Turabian StyleDong, Yanfang, Xin Huang, Wenlu Zhang, Yu Shao, Pierre Cloutier, Yi Zheng, and Léon Sanche. 2025. "Hyperthermal Reactions in DNA Triggered by 1–20 eV Electrons: Absolute Cross Sections for Crosslinks, Strand Breaks, Clustered Damages and Base Modifications" International Journal of Molecular Sciences 26, no. 9: 4057. https://doi.org/10.3390/ijms26094057
APA StyleDong, Y., Huang, X., Zhang, W., Shao, Y., Cloutier, P., Zheng, Y., & Sanche, L. (2025). Hyperthermal Reactions in DNA Triggered by 1–20 eV Electrons: Absolute Cross Sections for Crosslinks, Strand Breaks, Clustered Damages and Base Modifications. International Journal of Molecular Sciences, 26(9), 4057. https://doi.org/10.3390/ijms26094057