
A Major Latex Protein-Encoding Gene from Populus simonii × P. nigra (PsnMLP328) Contributes to Defense Responses to Salt and Cadmium Stress

Abstract
1. Introduction
2. Results
2.1. Cloning and Sequence Analysis of PsnMLP328
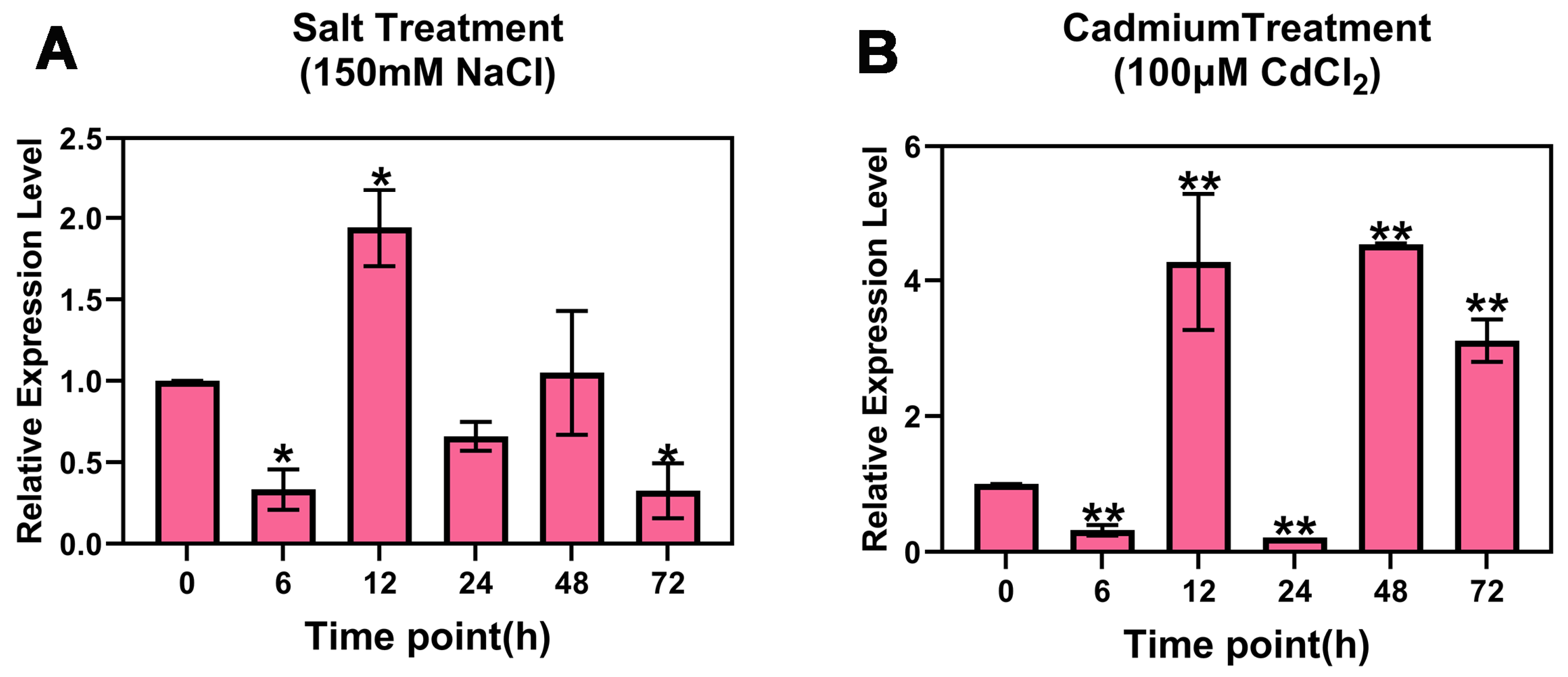
2.2. Response of PsnMLP328 to Salt Stress and Cadmium Stress
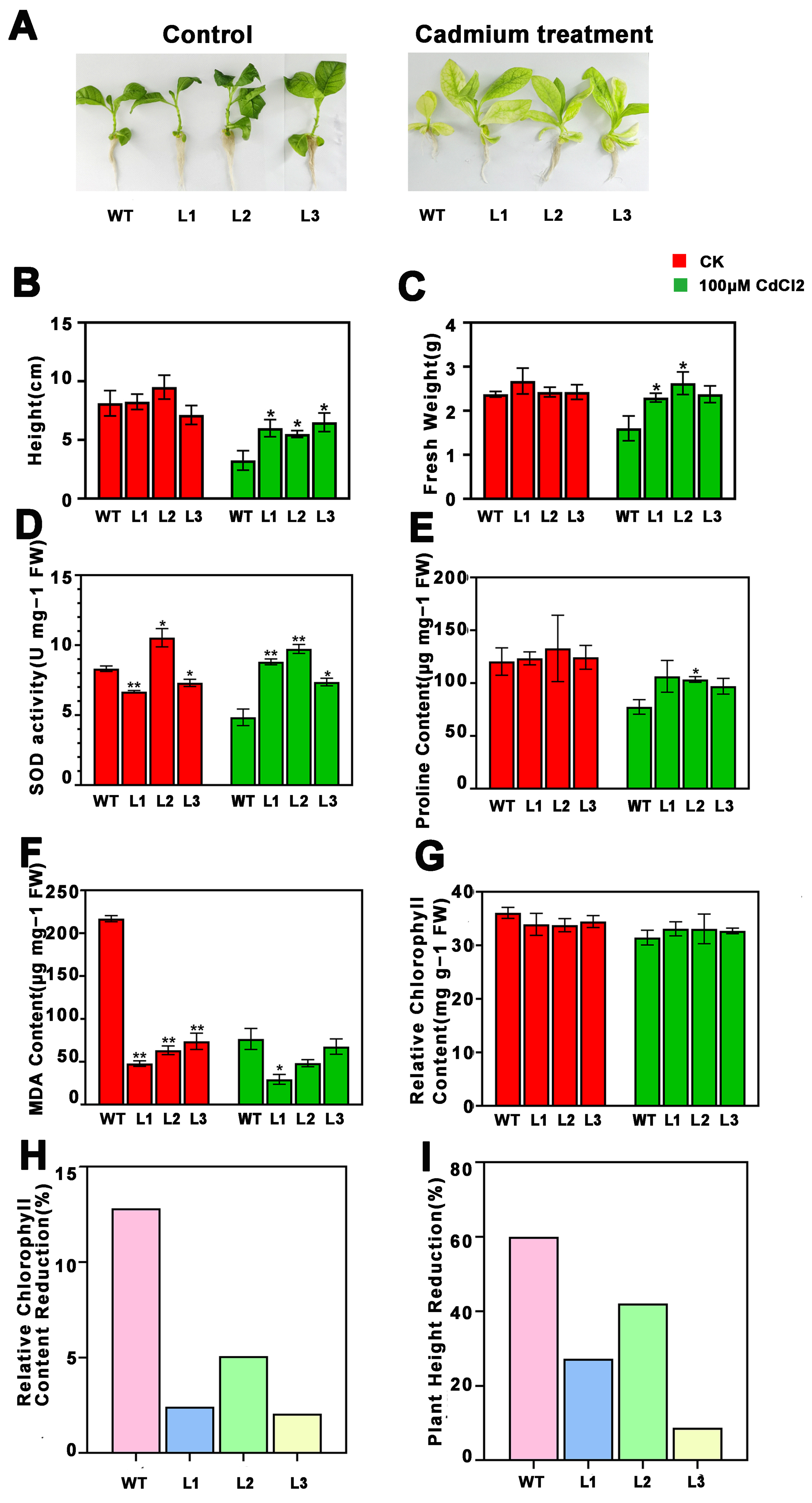
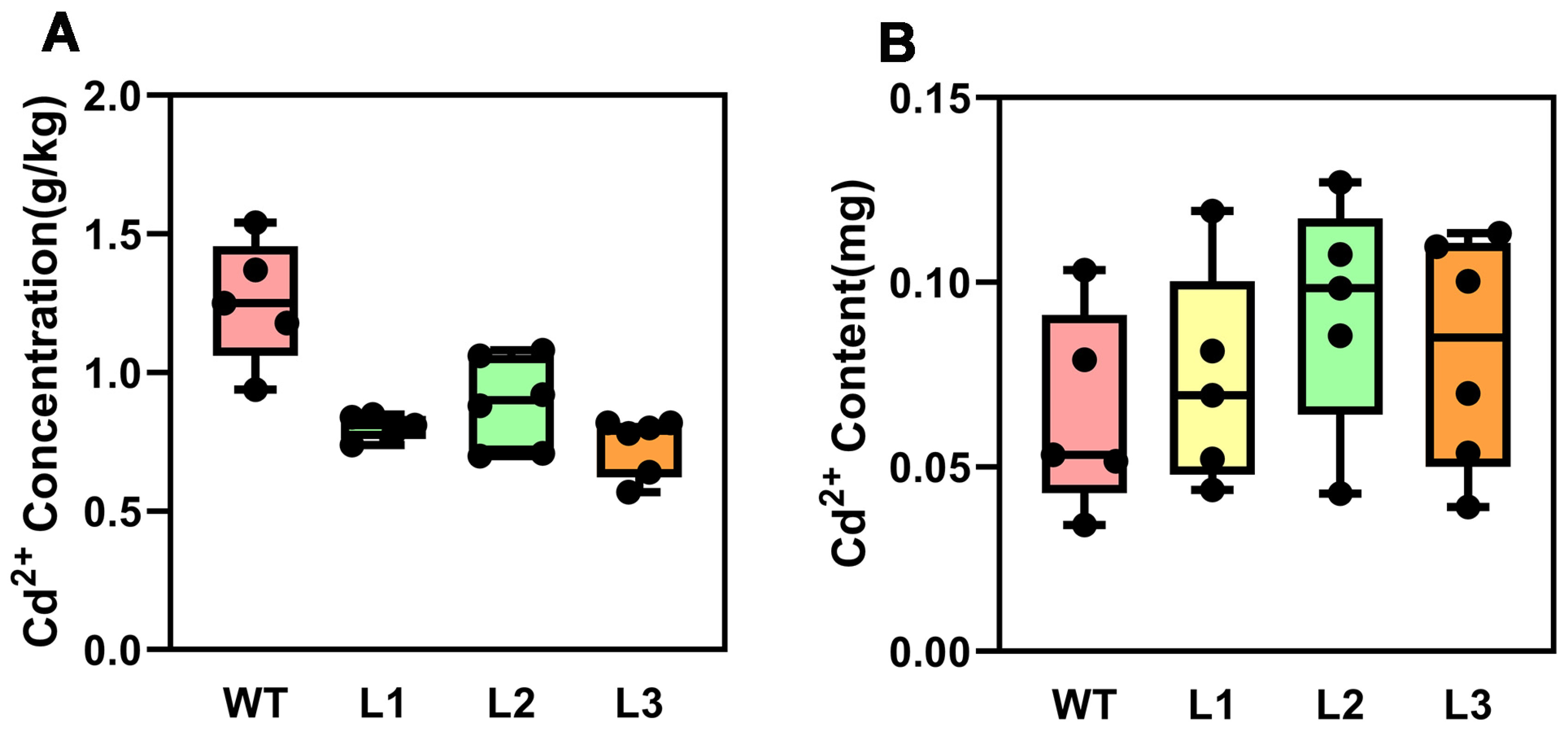
2.3. Overexpression of PsnMLP328 Can Improve Resistance in Transgenic Tobacco Under Salt and Cadmium Stress
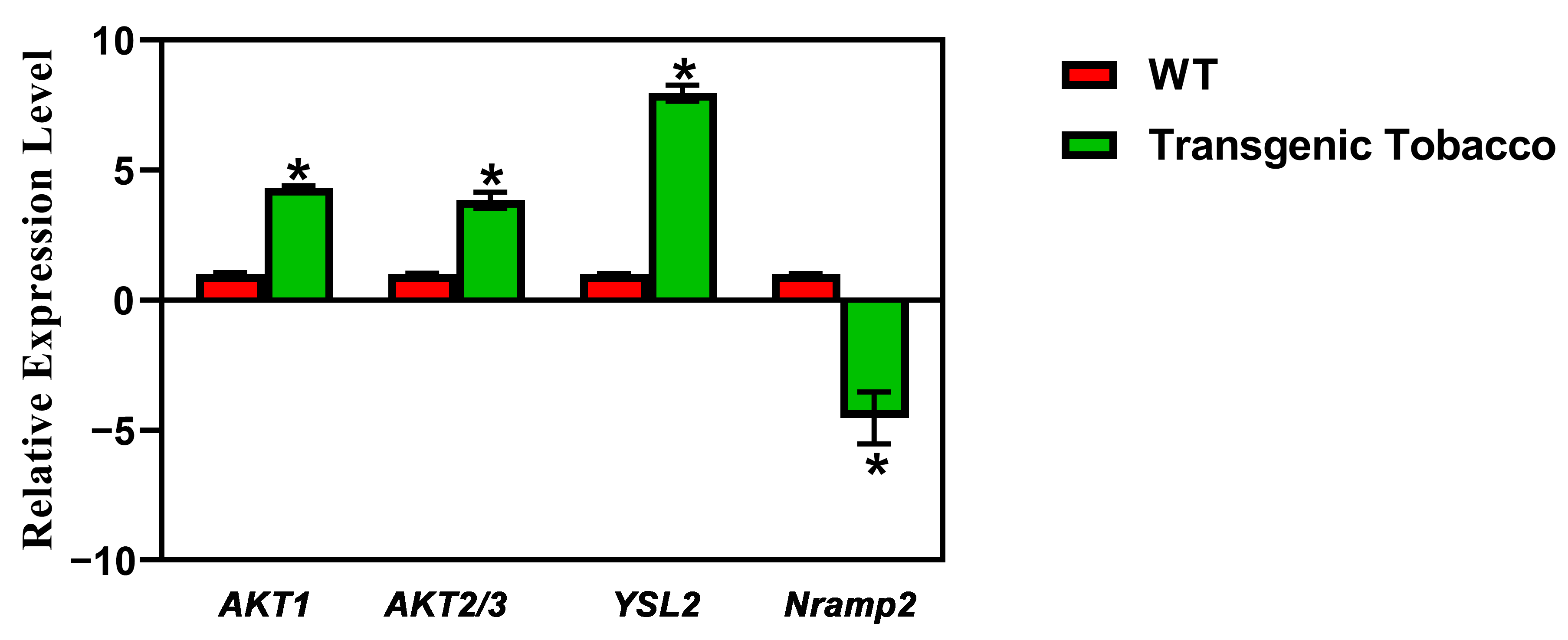
2.4. Expression Analysis of Transgenic Plants and the Prediction of Genes Regulated by PsnMLP328
3. Discussion
3.1. Possible Mechanisms by Which PsnMLP328 Regulates Salt-Stress Resistance in Plants
3.2. Possible Mechanisms by Which PsnMLP328 Regulates Cadmium-Stress Resistance in Plants
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Gene Cloning and Sequence Analysis of PsnMLP328
4.3. Vector Construction and Transformation of Nicotiana tabacum L. cv. Petit Havana SR-1
4.4. Identification of Transgenic Plants
4.5. Plant Treatment and Expression Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, S.; Naushad, M.; Lima, E.C.; Zhang, S.; Shaheen, S.M.; Rinklebe, J. Global soil pollution by toxic elements: Current status and future perspectives on the risk assessment and remediation strategies—A review. J. Hazard. Mater. 2021, 417, 126039. [Google Scholar]
- Litalien, A.; Zeeb, B. Curing the earth: A review of anthropogenic soil salinization and plant-based strategies for sustainable mitigation. Sci. Total Environ. 2020, 698, 134235. [Google Scholar]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Networks in Response to Abiotic Stresses in Arabidopsis and Grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar]
- Ma, L.; Liu, X.; Lv, W.; Yang, Y. Molecular mechanisms of plant responses to salt stress. Front. Plant Sci. 2022, 13, 934877. [Google Scholar]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar]
- Qin, G.; Niu, Z.; Yu, J.; Li, Z.; Ma, J.; Xiang, P. Soil heavy metal pollution and food safety in China: Effects, sources and removing technology. Chemosphere 2021, 267, 129205. [Google Scholar]
- Asare, M.O.; Száková, J.; Tlustoš, P. Mechanisms of As, Cd, Pb, and Zn hyperaccumulation by plants and their effects on soil microbiome in the rhizosphere. Front. Environ. Sci. 2023, 11, 1157415. [Google Scholar]
- Lai, C.W.; Lou, F.; Liu, J.L.; Liu, L.; Zhou, Y.Y.; Shi, S.Y. Progress and Prospects of Research on Cadmium Tolerance Genes in Plants. J. Org. Chem. Res. 2023, 11, 57–74. [Google Scholar]
- Li, Y.; Ding, L.; Zhou, M.; Chen, Z.; Ding, Y.; Zhu, C. Transcriptional Regulatory Network of Plant Cadmium Stress Response. Int. J. Mol. Sci. 2023, 24, 4378. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ma, C.; Ma, Y.; Li, H.; Kang, J.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Cadmium tolerance in six poplar species. Environ. Sci. Pollut. Res. Int. 2013, 20, 163–174. [Google Scholar] [PubMed]
- Clemens, S.; Aarts, M.G.M.; Thomine, S.; Verbruggen, N. Plant science: The key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [PubMed]
- Singh, P.; Mitra, P.; Goyal, T.; Sharma, S.; Sharma, P. Blood lead and cadmium levels in occupationally exposed workers and their effect on markers of DNA damage and repair. Environ. Geochem. Health 2021, 43, 185–193. [Google Scholar]
- Kumar, S.; Sharma, A. Cadmium toxicity: Effects on human reproduction and fertility. Rev. Environ. Health 2019, 34, 327–338. [Google Scholar]
- Noor, I.; Sohail, H.; Akhtar, M.T.; Cui, J.; Lu, Z.; Mostafa, S.; Hasanuzzaman, M.; Hussain, S.; Guo, N.; Jin, B. From stress to resilience: Unraveling the molecular mechanisms of cadmium toxicity, detoxification and tolerance in plants. Sci. Total Environ. 2024, 954, 176462. [Google Scholar] [CrossRef]
- Niu, L.; Li, C.; Wang, W.; Zhang, J.; Scali, M.; Li, W.; Liu, H.; Tai, F.; Hu, X.; Wu, X. Cadmium tolerance and hyperaccumulation in plants—A proteomic perspective of phytoremediation. Ecotoxicol. Environ. Saf. 2023, 256, 114882. [Google Scholar] [CrossRef]
- Yue, C.; Huang, S.Y.; Tu, C.B.; Wu, C.F.; He, Y.; Wang, Z. Research Progress on Phytoremediation of Soil Contaminated by Heavy Metal Cadmium. Mod. Agric. Sci. Technol. 2024, 17, 6. [Google Scholar]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of Cadmium: Physiological, Biochemical, and Molecular Mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef]
- Cao, Y.; Tan, Q.; Zhang, F.; Ma, C.; Xiao, J.; Chen, G. Phytoremediation potential evaluation of multiple Salix clones for heavy metals (Cd, Zn and Pb) in flooded soils. Sci. Total Environ. 2022, 813, 152482. [Google Scholar]
- Qiao, K.; Shan, Q.; Zhang, H.; Lv, F.; Zhou, A. Populus euphratica plant cadmium tolerance PePCR3 improves cadmium tolerance. Tree Physiol. 2023, 43, 1950–1963. [Google Scholar] [PubMed]
- Shi, R.; Liang, L.; Liu, W.; Zeb, A. Kochia scoparia L., a newfound candidate halophyte, for phytoremediation of cadmium-contaminated saline soils. Environ. Sci. Pollut. Res. Int. 2022, 29, 44759–44768. [Google Scholar] [PubMed]
- Hussain, M.S.; Naeem, M.S.; Tanvir, M.A.; Nawaz, M.F.; Abd-Elrahman, A. Eco-physiological evaluation of multipurpose tree species to ameliorate saline soils. Int. J. Phytoremediat. 2021, 23, 969–981. [Google Scholar]
- Zhang, Y.; Sa, G.; Zhang, Y.; Hou, S.; Wu, X.; Zhao, N.; Zhang, Y.; Deng, S.; Deng, C.; Deng, J.; et al. Populus euphratica annexin1 facilitates cadmium enrichment in transgenic Arabidopsis. J. Hazard. Mater. 2021, 405, 124063. [Google Scholar]
- Bedair, H.; Ghosh, S.; Abdelsalam, I.M.; Keerio, A.A.; AlKafaas, S.S. Potential implementation of trees to remediate contaminated soil in Egypt. Environ. Sci. Pollut. Res. Int. 2022, 29, 78132–78151. [Google Scholar]
- He, J.; Li, H.; Luo, J.; Ma, C.; Li, S.; Qu, L.; Gai, Y.; Jiang, X.; Janz, D.; Polle, A.; et al. A Transcriptomic Network Underlies Microstructural and Physiological Responses to Cadmium in Populus × canescens. Plant Physiol. 2013, 162, 424–439. [Google Scholar]
- Fujita, K.; Inui, H. Review: Biological functions of major latex-like proteins in plants. Plant Sci. Int. J. Exp. Plant Biol. 2021, 306, 110856. [Google Scholar] [CrossRef]
- Nessler, C.L.; Kurz, W.G.W.; Pelcher, L.E. Isolation and analysis of the major latex protein genes of opium poppy. Plant Mol. Biol. 1990, 15, 951–953. [Google Scholar] [CrossRef]
- Ipsen, H.; Løwenstein, H. Isolation and immunochemical characterization of the major allergen of birch pollen (Betula verrucosa). J. Allergy Clin. Immunol. 1983, 72, 150–159. [Google Scholar]
- Inui, H.; Sawada, M.; Goto, J.; Yamazaki, K.; Kodama, N.; Tsuruta, H.; Eun, H. A Major Latex-Like Protein Is a Key Factor in Crop Contamination by Persistent Organic Pollutants. Plant Physiol. 2013, 161, 2128–2135. [Google Scholar]
- Nawrot, R.; Lippmann, R.; Matros, A.; Musidlak, O.; Nowicki, G.; Mock, H.P. Proteomic comparison of Chelidonium majus L. latex in different phases of plant development. Plant Physiol. Biochem. 2017, 112, 312–325. [Google Scholar] [PubMed]
- Holmquist, L.; Dölfors, F.; Fogelqvist, J.; Cohn, J.; Kraft, T.; Dixelius, C. Major latex protein-like encoding genes contribute to Rhizoctonia solani defense responses in sugar beet. Mol. Genet. Genom. 2020, 296, 155–164. [Google Scholar]
- Song, L.; Wang, J.; Jia, H.; Kamran, A.; Qin, Y.; Liu, Y.; Hao, K.; Han, F.; Zhang, C.; Li, B.; et al. Identification and functional characterization of NbMLP28, a novel MLP-like protein 28 enhancing Potato virus Y resistance in Nicotiana benthamiana. BMC Microbiol. 2020, 20, 55. [Google Scholar]
- Wang, Y.; Yang, L.; Chen, X.; Ye, T.; Zhong, B.; Liu, R.; Wu, Y.; Chan, Z. Major latex protein-like protein 43(MLP43) functions as a positive regulator during abscisic acid responses and confers drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 421–434. [Google Scholar]
- Liu, H.; Ma, X.; Liu, S.; Du, B.; Cheng, N.; Wang, Y.; Zhang, Y. The Nicotiana tabacum L. major latex protein-like protein 423 (NtMLP423) positively regulates drought tolerance by ABA-dependent pathway. BMC Plant Biol. 2020, 20, 475. [Google Scholar]
- Yao, W.J.; Wang, L.; Zhou, B.R.; Wang, S.J.; Li, R.H.; Jiang, T.B. Over-expression of poplar transcription factor ERF76 gene confers salt tolerance in transgenic tobacco. J. Plant Physiol. 2016, 198, 23–31. [Google Scholar]
- Manikandan, M.; Kannan, V.; Mahalingam, K.; Vimala, A.; Chun, S. Phytoremediation potential of chromium-containing tannery effluent-contaminated soil by native Indian timber-yielding tree species. Prep. Biochem. Biotechnol. 2016, 46, 100–108. [Google Scholar]
- Landmeyer, J.E.; Rock, S.; Freeman, J.L.; Nagle, G.; Samolis, M.; Levine, H.; Cook, A.M.; O’Neill, H. Phytoremediation of slightly brackish, polycyclic aromatic hydrocarbon-contaminated groundwater from 250 ft below land surface: A pilot-scale study using salt-tolerant, endophyte-enhanced hybrid poplar trees at a Superfund site in the Central Valley of California, April–November 2019. Remediat. J. 2020, 31, 73–89. [Google Scholar]
- Shabir, R.; Abbas, G.; Saqib, M.; Shahid, M.; Shah, G.M.; Akram, M.; Niazi, N.K.; Naeem, M.A.; Hussain, M.; Ashraf, F. Cadmium tolerance and phytoremediation potential of acacia (Acacia nilotica L.) under salinity stress. Int. J. Phytoremediat. 2018, 20, 739–746. [Google Scholar]
- Ebone, L.A.; Caverzan, A.; Chavarria, G. Physiologic alterations in orthodox seeds due to deterioration processes. Plant Physiol. Biochem. 2019, 145, 34–42. [Google Scholar]
- Han, D.; Han, J.; Xu, T.; Li, T.; Yao, C.; Wang, Y.; Luo, D.; Yang, G. Isolation and preliminary functional characterization of MxWRKY64, a new WRKY transcription factor gene from Malus xiaojinensis Cheng et Jiang. In Vitr. Cell. Dev. Biol. Plant 2021, 57, 202–213. [Google Scholar]
- Wei, Z.; Ye, J.; Zhou, Z.; Chen, G.; Meng, F.; Liu, Y. Isolation and characterization of PoWRKY, an abiotic stress-related WRKY transcription factor from Polygonatum odoratum. Physiol. Mol. Biol. Plants 2021, 27, 1–9. [Google Scholar] [PubMed]
- Wu, B.; Qi, F.; Liang, Y. Fuels for ROS signaling in plant immunity. Trends Plant Sci. 2023, 28, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar]
- Czégény, G.; Rácz, A. Phenolic peroxidases: Dull generalists or purposeful specialists in stress responses. J. Plant Physiol. 2023, 280, 153884. [Google Scholar]
- Liu, H.; Du, B.; Ma, X.; Wang, Y.; Cheng, N.; Zhang, Y. Overexpression of major latex protein 423 (NtMLP423) enhances the chilling stress tolerance in Nicotiana tabacum. Plant Sci. 2023, 329, 111604. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef]
- Lebaudy, A.; Véry, A.A.; Sentenac, H. K+ channel activity in plants: Genes, regulations and functions. FEBS Lett. 2007, 581, 2357–2366. [Google Scholar] [CrossRef]
- Hirsch, R.E.; Lewis, B.D.; Spalding, E.P.; Sussman, M.R. A role for the AKT1 potassium channel in plant nutrition. Science 1998, 280, 918–921. [Google Scholar]
- Lagarde, D.; Basset, M.; Lepetit, M.; Conejero, G.; Gaymard, F.; Astruc, S.; Grignon, C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. Cell Mol. Biol. 1996, 9, 195–203. [Google Scholar]
- Sandmann, M.; Skłodowski, K.; Gajdanowicz, P.; Michard, E.; Rocha, M.; Gomez-Porras, J.L.; González, W.; Corrêa, L.G.G.; Ramírez-Aguilar, S.J.; Cuin, T.A.; et al. The K+ battery-regulating Arabidopsis K+ channel AKT2 is under the control of multiple post-translational steps. Plant Signal. Behav. 2011, 6, 558–562. [Google Scholar] [PubMed]
- Cuin, T.; Dreyer, I.; Michard, E. The Role of Potassium Channels in Arabidopsis thaliana Long Distance Electrical Signalling: AKT2 Modulates Tissue Excitability While GORK Shapes Action Potentials. Int. J. Mol. Sci. 2018, 19, 926. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Shen, L.; Luan, J.; Zhou, Z.; Guo, D.; Shen, Y.; Jing, W.; Zhang, B.; Zhang, Q.; Zhang, W. Rice shaker potassium channel OsAKT2 positively regulates salt tolerance and grain yield by mediating K+ redistribution. Plant Cell Environ. 2021, 44, 2951–2965. [Google Scholar] [PubMed]
- Zhang, X.; Li, M.; Yang, H.; Li, X.; Cui, Z. Physiological responses of Suaeda glauca and Arabidopsis thaliana in phytoremediation of heavy metals. J. Environ. Manag. 2018, 223, 132–139. [Google Scholar]
- Li, L.Z.; Tu, C.; Peijnenburg, W.J.G.M.; Luo, Y.M. Characteristics of cadmium uptake and membrane transport in roots of intact wheat (Triticum aestivum L.) seedlings. Environ. Pollut. 2017, 221, 351–358. [Google Scholar]
- Rahman, M.F.; Ghosal, A.; Alam, M.F.; Kabir, A.H. Remediation of cadmium toxicity in field peas (Pisum sativum L.) through exogenous silicon. Ecotoxicol. Environ. Saf. 2017, 135, 165–172. [Google Scholar]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar]
- Lian, J.; Zhao, L.; Wu, J.; Xiong, H.; Bao, Y.; Zeb, A.; Tang, J.; Liu, W. Foliar spray of TiO2 nanoparticles prevails over root application in reducing Cd accumulation and mitigating Cd-induced phytotoxicity in maize (Zea mays L.). Chemosphere 2020, 239, 124794. [Google Scholar]
- Dong, Y.J.; Chen, W.F.; Liu, F.Z.; Wan, Y.S. Physiological responses of peanut seedlings to exposure to low or high cadmium concentration and the alleviating effect of exogenous nitric oxide to high cadmium concentration stress. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2019, 154, 405–412. [Google Scholar]
- Siripornadulsil, S.; Traina, S.; Verma, D.P.S.; Sayre, R.T. Molecular Mechanisms of Proline-Mediated Tolerance to Toxic Heavy Metals in Transgenic Microalgae. Plant Cell 2002, 14, 2837–2847. [Google Scholar]
- Sun, R.L.; Zhou, Q.X.; Sun, F.H.; Jin, C.X. Antioxidative defense and proline/phytochelatin accumulation in a newly discovered Cd-hyperaccumulator, Solanum nigrum L. Environ. Exp. Bot. 2007, 60, 468–476. [Google Scholar]
- Tan, L.; Zhu, Y.; Fan, T.; Peng, C.; Wang, J.; Sun, L.; Chen, C. OsZIP7 functions in xylem loading in roots and inter-vascular transfer in nodes to deliver Zn/Cd to grain in rice. Biochem. Biophys. Res. Commun. 2019, 512, 112–118. [Google Scholar] [PubMed]
- Zhao, J.; Yang, W.; Zhang, S.; Yang, T.; Liu, Q.; Dong, J.; Fu, H.; Mao, X.; Liu, B. Genome-wide association study and candidate gene analysis of rice cadmium accumulation in grain in a diverse rice collection. Rice 2018, 11, 61. [Google Scholar]
- Koike, S.; Inoue, H.; Mizuno, D.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem. Plant J. 2004, 39, 415–424. [Google Scholar]
- Wu, X.; Chen, Q.; Chen, L.; Tian, F.; Chen, X.; Han, C.; Mi, J.; Lin, X.; Wan, X.; Jiang, B.; et al. A WRKY transcription factor, PyWRKY75, enhanced cadmium accumulation and tolerance in poplar. Ecotoxicol. Environ. Saf. 2022, 239, 113630. [Google Scholar]
- He, Y.; Liu, M.; Wang, R.; Salam, M.; Yang, Y.; Zhang, Z.; He, Q.; Hu, X.; Li, H. Potassium regulates cadmium toxicity in Microcystis aeruginosa. J. Hazard. Mater. 2021, 413, 125374. [Google Scholar]
- Liu, P.; Jin, Z.; Dai, C.; Guo, L.; Cui, X.; Yang, Y. Potassium enhances cadmium resistance ability of Panax notoginseng by brassinolide signaling pathway-regulated cell wall pectin metabolism. Ecotoxicol. Environ. Saf. 2021, 227, 112906. [Google Scholar] [CrossRef]
- Huang, B.; Liao, Q.; Fu, H.; Ye, Z.; Mao, Y.; Luo, J.; Wang, Y.; Yuan, H.; Xin, J. Effect of potassium intake on cadmium transporters and root cell wall biosynthesis in sweet potato. Ecotoxicol. Environ. Saf. 2023, 250, 114501. [Google Scholar] [CrossRef]
- Topping, J.F. Tobacco transformation. Methods Mol. Biol. 1998, 81, 365–372. [Google Scholar] [CrossRef]
- Kamitani, M.; Kashima, M.; Tezuka, A.; Nagano, A.J. Lasy-Seq: A High-throughput library preparation method for RNA-Seq and its application in the analysis of plant responses to fluctuating temperatures. Sci. Rep. 2019, 9, 7091. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Casein kinase II phosphorylation site | |
| 35–38: | Phosphoserine |
| 106–109: | Phosphoserine |
| N-glycosylation site | |
| 33–36: | N-linked (GlcNAc…) asparagine |
| 79–82: | N-linked (GlcNAc…) asparagine |
| Protein kinase C phosphorylation site | |
| 54–56: | Phosphoserine |
| 97–99: | Phosphoserine |
| 101–103: | Phosphothreonine |
| Gene Name | Log2-Fold Change | p Value | Padj | Description |
|---|---|---|---|---|
| AKT1 | 2.4053718 | 4.21 × 10−5 | 0.001272395 | potassium channel AKT1-like%2C transcript variant X2 |
| AKT2/3 | 1.754015611 | 2.90 × 10−6 | 0.000163073 | potassium channel AKT2/3-like |
| YSL2 | 3.514848502 | 1.58 × 10−12 | 7.44 × 10−10 | metal–nicotianamine transporter YSL2-like%2C transcript variant X1 |
| Nramp2 | −2.125372581 | 0.000164615 | 0.003521159 | metal transporter Nramp2-like |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Wang, L.; Liu, S.; Li, Y.; Sun, Y.; Wu, Q.; Fu, D. A Major Latex Protein-Encoding Gene from Populus simonii × P. nigra (PsnMLP328) Contributes to Defense Responses to Salt and Cadmium Stress. Int. J. Mol. Sci. 2025, 26, 3350. https://doi.org/10.3390/ijms26073350
Sun X, Wang L, Liu S, Li Y, Sun Y, Wu Q, Fu D. A Major Latex Protein-Encoding Gene from Populus simonii × P. nigra (PsnMLP328) Contributes to Defense Responses to Salt and Cadmium Stress. International Journal of Molecular Sciences. 2025; 26(7):3350. https://doi.org/10.3390/ijms26073350
Chicago/Turabian StyleSun, Xin, Lei Wang, Shuang Liu, Yao Li, Yao Sun, Qiong Wu, and Di Fu. 2025. "A Major Latex Protein-Encoding Gene from Populus simonii × P. nigra (PsnMLP328) Contributes to Defense Responses to Salt and Cadmium Stress" International Journal of Molecular Sciences 26, no. 7: 3350. https://doi.org/10.3390/ijms26073350
APA StyleSun, X., Wang, L., Liu, S., Li, Y., Sun, Y., Wu, Q., & Fu, D. (2025). A Major Latex Protein-Encoding Gene from Populus simonii × P. nigra (PsnMLP328) Contributes to Defense Responses to Salt and Cadmium Stress. International Journal of Molecular Sciences, 26(7), 3350. https://doi.org/10.3390/ijms26073350