


Cancer Development and Progression Through a Vicious Cycle of DNA Damage and Inflammation




Abstract
1. Introduction
2. DNA Damage in Inflammation-Related Cancer
2.1. Pathogens
2.1.1. Opisthorchis viverrini
2.1.2. Schistosoma haematobium
2.1.3. Helicobacter pylori
2.1.4. Hepatitis B/C Virus
2.1.5. Human Papillomavirus
2.1.6. Epstein–Barr Virus
2.2. Environmental Factors
2.2.1. Ultraviolet Light
2.2.2. Asbestos and Nanomaterials
2.3. Inflammation-Related Disorders
2.3.1. Oral Lichen Planus/Leukoplakia
2.3.2. Inflammatory Bowel Disease
2.3.3. Gastric Acid Reflux (Barrett’s Esophagus)
2.3.4. Malignant Fibrous Histiocytomas (Undifferentiated Pleomorphic Sarcomas)
2.4. 8-NitroG as a Biomarker Candidate
3. DNA Damage and Cancer Stem Cells
3.1. CSCs in OV-Induced Cholangiocarcinoma
3.2. CSCs in SH-Infected Bladder Cancer Cells
3.3. CSCs in Nasopharyngeal Carcinoma
3.4. CSCs in Barrett’s Esophageal Adenocarcinoma
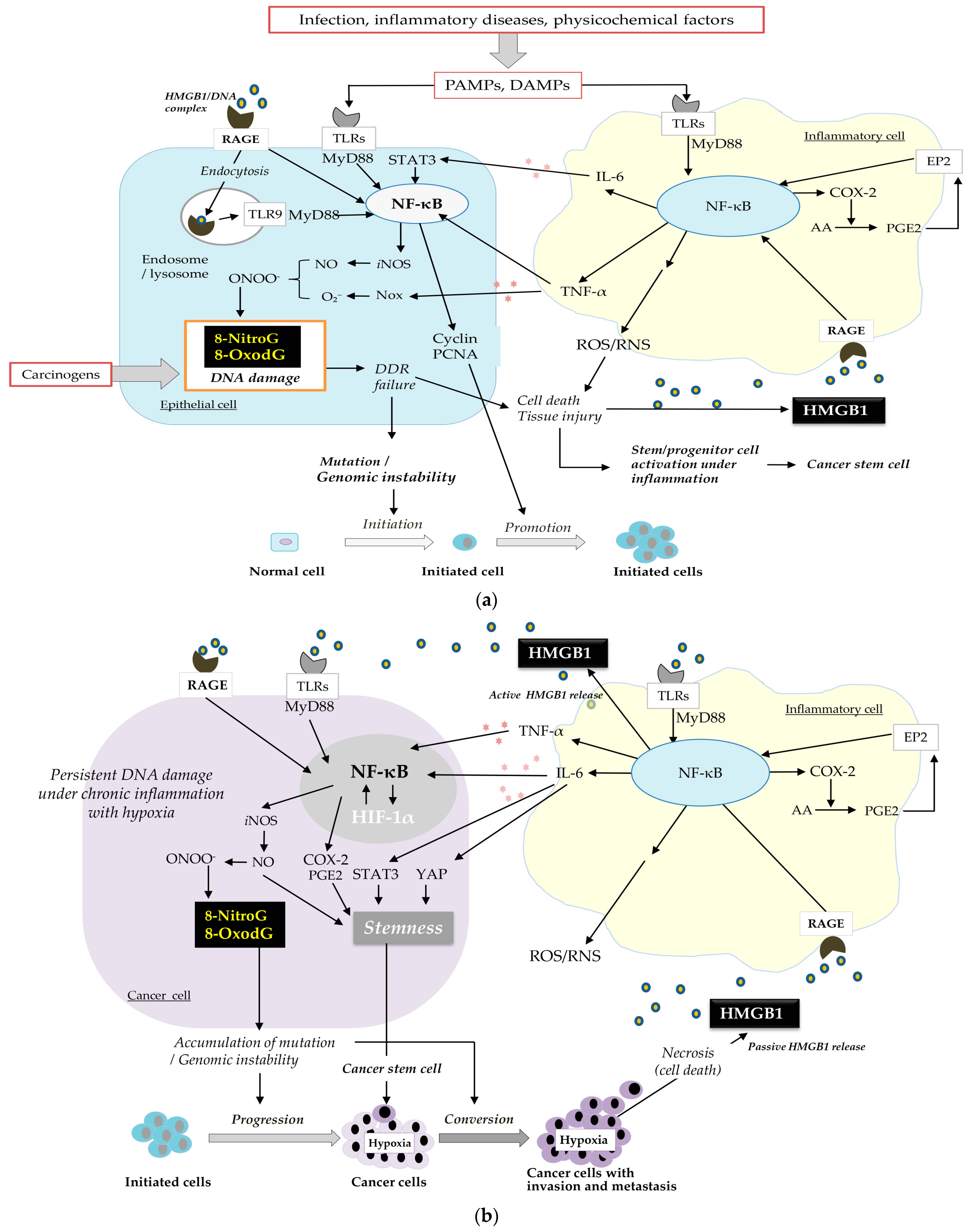
4. Mechanism of Inflammation-Related Carcinogenesis
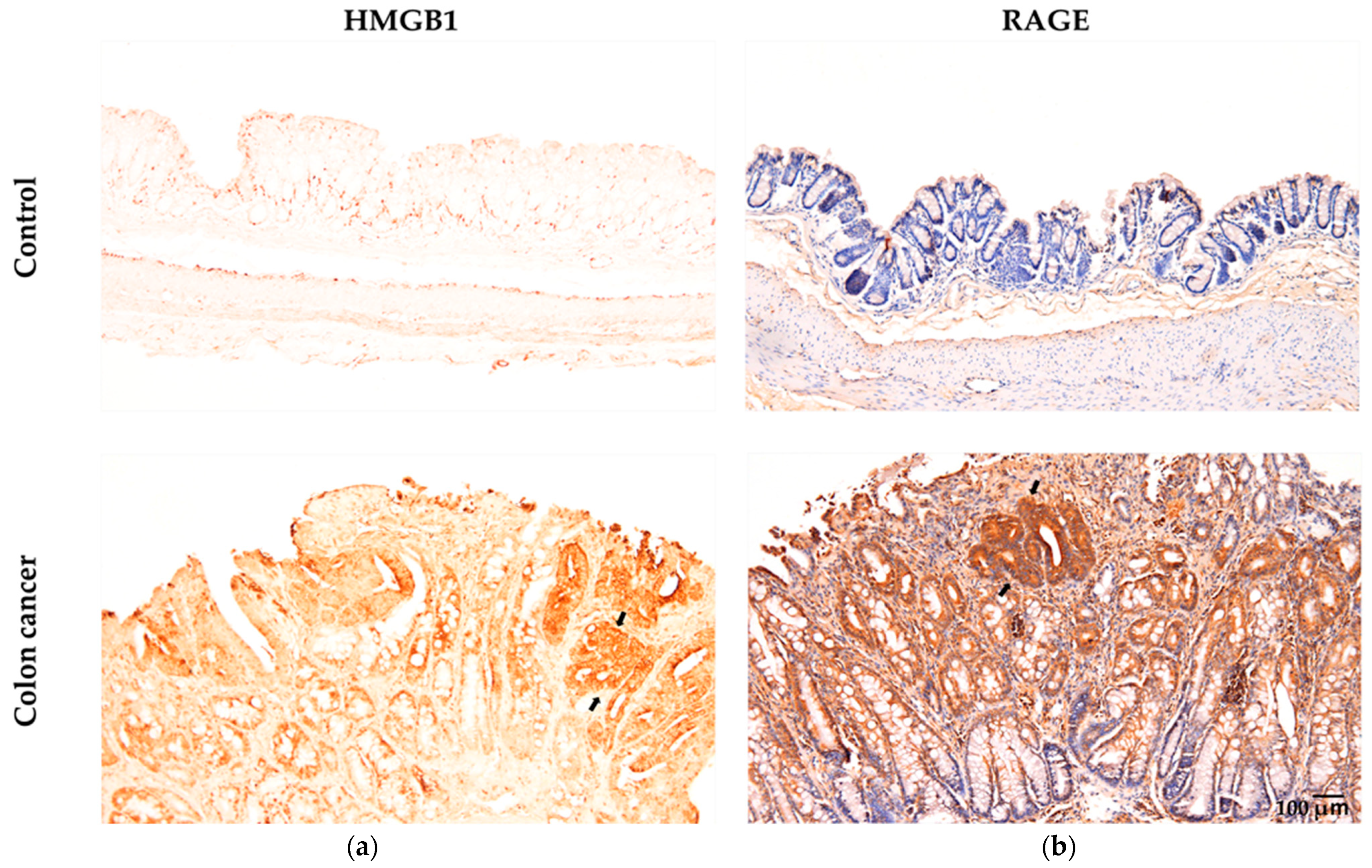
4.1. HMGB1 as a Key Factor in the Mechanism
4.2. Cancer Development and Progression Occur Through a Vicious Cycle of DNA Damage and Inflammation via HMGB1
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 8-oxodG | 8-Oxo-7,8-dihydro-2′-deoxyguanosine |
| 8-nitroG | 8-Nitroguanine |
| AOM | Azoxymethane |
| AA | Arachidonic acid |
| BE | Barrett’s esophagus |
| BEA | Barrett’s esophageal adenocarcinoma |
| CB | Carbon black |
| CCA | Cholangiocarcinoma |
| CD44v9 | CD44 variant 9 |
| CIN | Cervical intraepithelial neoplasia |
| CSC | Cancer stem cell |
| DAMP | Damage-associated molecular pattern |
| DDR | DNA damage response |
| DSS | Dextran sodium sulfate |
| EBV | Epstein–Barr virus |
| EBER | EBV-encoded RNA |
| EMT | Epithelial–mesenchymal transition |
| GL | Glycyrrhizin |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| HCC | Hepatocellular carcinoma |
| HIF | Hypoxia-inducible factor |
| HPV | Human papillomavirus |
| HMGB1 | High mobility group box 1 |
| IBD | Inflammatory bowel disease |
| IHC | Immunohistochemical |
| iNOS | Inducible nitric oxide synthase |
| LMP1 | latent membrane protein 1 |
| MFH | Malignant fibrous histiocytoma |
| NO | Nitric oxide |
| Nox | NADPH oxidase |
| NPC | Nasopharyngeal carcinoma |
| OLP | Oral lichen planus |
| OSCC | Oral squamous cell carcinoma |
| OV | Opisthorchis viverrini |
| PAMP | Pathogen-associated molecular pattern |
| PG | Prostaglandin |
| PPI | Proton pump inhibitor |
| RAGE | Receptor for advanced glycation end products |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| PCNA | Proliferating cell nuclear antigen |
| SH | Schistosoma haematobium |
| SOD | Superoxide dismutase |
| TLR | Toll-like receptor |
| UC | Ulcerative colitis |
| UV | Ultraviolet |
References
- Kawanishi, S.; Ohnishi, S.; Ma, N.; Hiraku, Y.; Murata, M. Crosstalk between DNA Damage and Inflammation in the Multiple Steps of Carcinogenesis. Int. J. Mol. Sci. 2017, 18, 1808. [Google Scholar] [CrossRef] [PubMed]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Hao, S.; Hong, W.; Tergaonkar, V.; Sethi, G.; Tian, Y.; Duan, C. Versatile function of NF-ĸB in inflammation and cancer. Exp. Hematol. Oncol. 2024, 13, 68. [Google Scholar] [PubMed]
- Kawanishi, S.; Hiraku, Y. Oxidative and nitrative DNA damage as biomarker for carcinogenesis with special reference to inflammation. Antioxid. Redox Signal. 2006, 8, 1047–1058. [Google Scholar]
- Hiraku, Y.; Kawanishi, S. Immunohistochemical analysis of 8-nitroguanine, a nitrative DNA lesion, in relation to inflammation-associated carcinogenesis. Methods Mol. Biol. 2009, 512, 3–13. [Google Scholar]
- Pinlaor, S.; Sripa, B.; Ma, N.; Hiraku, Y.; Yongvanit, P.; Wongkham, S.; Pairojkul, C.; Bhudhisawasdi, V.; Oikawa, S.; Murata, M.; et al. Nitrative and oxidative DNA damage in intrahepatic cholangiocarcinoma patients in relation to tumor invasion. World J. Gastroenterol. 2005, 11, 4644–4649. [Google Scholar] [CrossRef]
- Thanan, R.; Pairojkul, C.; Pinlaor, S.; Khuntikeo, N.; Wongkham, C.; Sripa, B.; Ma, N.; Vaeteewoottacharn, K.; Furukawa, A.; Kobayashi, H.; et al. Inflammation-related DNA damage and expression of CD133 and Oct3/4 in cholangiocarcinoma patients with poor prognosis. Free Radic. Biol. Med. 2013, 65, 1464–1472. [Google Scholar]
- Pinlaor, S.; Ma, N.; Hiraku, Y.; Yongvanit, P.; Semba, R.; Oikawa, S.; Murata, M.; Sripa, B.; Sithithaworn, P.; Kawanishi, S. Repeated infection with Opisthorchis viverrini induces accumulation of 8-nitroguanine and 8-oxo-7,8-dihydro-2′-deoxyguanine in the bile duct of hamsters via inducible nitric oxide synthase. Carcinogenesis 2004, 25, 1535–1542. [Google Scholar]
- Pinlaor, S.; Hiraku, Y.; Yongvanit, P.; Tada-Oikawa, S.; Ma, N.; Pinlaor, P.; Sithithaworn, P.; Sripa, B.; Murata, M.; Oikawa, S.; et al. iNOS-dependent DNA damage via NF-kappaB expression in hamsters infected with Opisthorchis viverrini and its suppression by the antihelminthic drug praziquantel. Int. J. Cancer 2006, 119, 1067–1072. [Google Scholar]
- Santos, L.L.; Santos, J.; Gouveia, M.J.; Bernardo, C.; Lopes, C.; Rinaldi, G.; Brindley, P.J.; Costa, J. Urogenital Schistosomiasis-History, Pathogenesis, and Bladder Cancer. J. Clin. Med. 2021, 10, 205. [Google Scholar] [CrossRef]
- Ma, N.; Thanan, R.; Kobayashi, H.; Hammam, O.; Wishahi, M.; El Leithy, T.; Hiraku, Y.; Amroel, K.; Oikawa, S.; Ohnishi, S.; et al. Nitrative DNA damage and Oct3/4 expression in urinary bladder cancer with Schistosoma haematobium infection. Biochem. Biophys. Res. Commun. 2011, 414, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Fei, X.; Li, N.; Xu, X.; Zhu, Y. Macrophage biology in the pathogenesis of Helicobacter pylori infection. Crit. Rev. Microbiol. 2024, 31, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Krzysiek-Maczka, G.; Brzozowski, T.; Ptak-Belowska, A. Helicobacter pylori-activated fibroblasts as a silent partner in gastric cancer development. Cancer Metastasis Rev. 2023, 42, 1219–1256. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Adachi, Y.; Hiraku, Y.; Horiki, N.; Horiike, S.; Imoto, I.; Pinlaor, S.; Murata, M.; Semba, R.; Kawanishi, S. Accumulation of 8-nitroguanine in human gastric epithelium induced by Helicobacter pylori infection. Biochem. Biophys. Res. Commun. 2004, 319, 506–510. [Google Scholar] [CrossRef]
- Acharya, N.; Patel, S.K.; Sahu, S.R.; Kumari, P. ‘PIPs’ in DNA polymerase: PCNA interaction affairs. Biochem. Soc. Trans. 2020, 48, 2811–2822. [Google Scholar] [CrossRef]
- Katsurahara, M.; Kobayashi, Y.; Iwasa, M.; Ma, N.; Inoue, H.; Fujita, N.; Tanaka, K.; Horiki, N.; Gabazza, E.C.; Takei, Y. Reactive nitrogen species mediate DNA damage in Helicobacter pylori-infected gastric mucosa. Helicobacter 2009, 14, 552–558. [Google Scholar] [CrossRef]
- Horiike, S.; Kawanishi, S.; Kaito, M.; Ma, N.; Tanaka, H.; Fujita, N.; Iwasa, M.; Kobayashi, Y.; Hiraku, Y.; Oikawa, S.; et al. Accumulation of 8-nitroguanine in the liver of patients with chronic hepatitis C. J. Hepatol. 2005, 43, 403–410. [Google Scholar] [CrossRef]
- Nosaka, T.; Naito, T.; Hiramatsu, K.; Ohtani, M.; Nemoto, T.; Marusawa, H.; Ma, N.; Hiraku, Y.; Kawanishi, S.; Yamashita, T.; et al. Gene expression profiling of hepatocarcinogenesis in a mouse model of chronic hepatitis B. PLoS ONE 2017, 12, e0185442. [Google Scholar] [CrossRef]
- Wei, F.; Georges, D.; Man, I.; Baussano, I.; Clifford, G.M. Causal attribution of human papillomavirus genotypes to invasive cervical cancer worldwide: A systematic analysis of the global literature. Lancet 2024, 404, 435–444. [Google Scholar] [CrossRef]
- Trugilo, K.P.; Cebinelli, G.C.M.; Castilha, E.P.; Silva, M.R.D.; Berti, F.C.B.; Oliveira, K.B.D. The role of transforming growth factor β in cervical carcinogenesis. Cytokine Growth Factor Rev. 2024, 80, 12–23. [Google Scholar]
- Hiraku, Y.; Tabata, T.; Ma, N.; Murata, M.; Ding, X.; Kawanishi, S. Nitrative and oxidative DNA damage in cervical intraepithelial neoplasia associated with human papilloma virus infection. Cancer Sci. 2007, 98, 964–972. [Google Scholar] [PubMed]
- Wong, K.C.W.; Hui, E.P.; Lo, K.W.; Lam, W.K.J.; Johnson, D.; Li, L.; Tao, Q.; Chan, K.C.A.; To, K.F.; King, A.D.; et al. Nasopharyngeal carcinoma: An evolving paradigm. Nat. Rev. Clin. Oncol. 2021, 18, 679–695. [Google Scholar] [PubMed]
- Ma, N.; Kawanishi, M.; Hiraku, Y.; Murata, M.; Huang, G.W.; Huang, Y.; Luo, D.Z.; Mo, W.G.; Fukui, Y.; Kawanishi, S. Reactive nitrogen species-dependent DNA damage in EBV-associated nasopharyngeal carcinoma: The relation to STAT3 activation and EGFR expression. Int. J. Cancer 2008, 122, 2517–2525. [Google Scholar] [PubMed]
- Yu, Z.W.; Zheng, M.; Fan, H.Y.; Liang, X.H.; Tang, Y.L. Ultraviolet (UV) radiation: A double-edged sword in cancer development and therapy. Mol. Biomed. 2024, 5, 49. [Google Scholar]
- Ma, N.; Ohnishi, S.; Hiraku, Y.; Nishigori, C.; Kawanishi, S. 8-Nitroguanine formation in UV-induced skin tumors in mice. Photomed. Photobiol. 2011, 33, 21–22. [Google Scholar]
- Shariev, A.; Menounos, S.; Laos, A.J.; Laxman, P.; Lai, D.; Hua, S.; Zinger, A.; McRae, C.R.; Casbolt, L.S.; Combes, V.; et al. Skin protective and regenerative effects of RM191A, a novel superoxide dismutase mimetic. Redox Biol. 2021, 38, 101790. [Google Scholar]
- Hiraku, Y.; Sakai, K.; Shibata, E.; Kamijima, M.; Hisanaga, N.; Ma, N.; Kawanishi, S.; Murata, M. Formation of the nitrative DNA lesion 8-nitroguanine is associated with asbestos contents in human lung tissues: A pilot study. J. Occup. Health 2014, 56, 186–196. [Google Scholar]
- Hiraku, Y.; Guo, F.; Ma, N.; Yamada, T.; Wang, S.; Kawanishi, S.; Murata, M. Multi-walled carbon nanotube induces nitrative DNA damage in human lung epithelial cells via HMGB1-RAGE interaction and Toll-like receptor 9 activation. Part. Fibre Toxicol. 2016, 13, 16. [Google Scholar] [CrossRef]
- Ahmed, S.; Kobayashi, H.; Afroz, T.; Ma, N.; Oikawa, S.; Kawanishi, S.; Murata, M.; Hiraku, Y. Nitrative DNA damage in lung epithelial cells exposed to indium nanoparticles and indium ions. Sci. Rep. 2020, 10, 10741. [Google Scholar]
- Chaiyarit, P.; Ma, N.; Hiraku, Y.; Pinlaor, S.; Yongvanit, P.; Jintakanon, D.; Murata, M.; Oikawa, S.; Kawanishi, S. Nitrative and oxidative DNA damage in oral lichen planus in relation to human oral carcinogenesis. Cancer Sci. 2005, 96, 553–559. [Google Scholar]
- Ma, N.; Tagawa, T.; Hiraku, Y.; Murata, M.; Ding, X.; Kawanishi, S. 8-Nitroguanine formation in oral leukoplakia, a premalignant lesion. Nitric Oxide 2006, 14, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Fanizza, J.; Bencardino, S.; Allocca, M.; Furfaro, F.; Zilli, A.; Parigi, T.L.; Fiorino, G.; Peyrin-Biroulet, L.; Danese, S.; D’Amico, F. Inflammatory Bowel Disease and Colorectal Cancer. Cancers 2024, 16, 2943. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Hiraku, Y.; Ma, N.; Kato, T.; Saito, K.; Nagahama, M.; Semba, R.; Kuribayashi, K.; Kawanishi, S. Inducible nitric oxide synthase-dependent DNA damage in mouse model of inflammatory bowel disease. Cancer Sci. 2005, 96, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Saigusa, S.; Araki, T.; Tanaka, K.; Hashimoto, K.; Okita, Y.; Fujikawa, H.; Okugawa, Y.; Toiyama, Y.; Inoue, Y.; Uchida, K.; et al. Identification of patients with developing ulcerative colitis-associated neoplasia by nitrative DNA damage marker 8-nitroguanin expression in rectal mucosa. J. Clin. Gastroenterol. 2013, 47, e80–e86. [Google Scholar] [CrossRef]
- Sachdeva, K.; Natarajan, K.; Iyer, P.G. Improving esophageal cancer screening across the globe: Translating knowledge into action. Indian J. Gastroenterol. Off. J. Indian Soc. Gastroenterol. 2024, 43, 705–716. [Google Scholar] [CrossRef]
- Thanan, R.; Ma, N.; Iijima, K.; Abe, Y.; Koike, T.; Shimosegawa, T.; Pinlaor, S.; Hiraku, Y.; Oikawa, S.; Murata, M.; et al. Proton pump inhibitors suppress iNOS-dependent DNA damage in Barrett’s esophagus by increasing Mn-SOD expression. Biochem. Biophys. Res. Commun. 2012, 421, 280–285. [Google Scholar] [CrossRef]
- Hoki, Y.; Hiraku, Y.; Ma, N.; Murata, M.; Matsumine, A.; Nagahama, M.; Shintani, K.; Uchida, A.; Kawanishi, S. iNOS-dependent DNA damage in patients with malignant fibrous histiocytoma in relation to prognosis. Cancer Sci. 2007, 98, 163–168. [Google Scholar] [CrossRef]
- Hoki, Y.; Murata, M.; Hiraku, Y.; Ma, N.; Matsumine, A.; Uchida, A.; Kawanishi, S. 8-Nitroguanine as a potential biomarker for progression of malignant fibrous histiocytoma, a model of inflammation-related cancer. Oncol. Rep. 2007, 18, 1165–1169. [Google Scholar]
- Luo, Y.; Xiao, J.H. Inflammatory auxo-action in the stem cell division theory of cancer. PeerJ 2023, 11, e15444. [Google Scholar] [CrossRef]
- Frąszczak, K.; Barczyński, B. The Role of Cancer Stem Cell Markers in Ovarian Cancer. Cancers 2023, 16, 40. [Google Scholar] [CrossRef]
- MacLean, M.R.; Walker, O.L.; Arun, R.P.; Fernando, W.; Marcato, P. Informed by Cancer Stem Cells of Solid Tumors: Advances in Treatments Targeting Tumor-Promoting Factors and Pathways. Int. J. Mol. Sci. 2024, 25, 4102. [Google Scholar] [CrossRef] [PubMed]
- Odarenko, K.V.; Zenkova, M.A.; Markov, A.V. The Nexus of Inflammation-Induced Epithelial-Mesenchymal Transition and Lung Cancer Progression: A Roadmap to Pentacyclic Triterpenoid-Based Therapies. Int. J. Mol. Sci. 2023, 24, 17325. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.A.; Zakeri, A.; Desgrosellier, J.S. Chromosomal instability as an architect of the cancer stemness landscape. Front. Cell Dev. Biol. 2024, 12, 1450614. [Google Scholar]
- Suwannakul, N.; Ma, N.; Thanan, R.; Pinlaor, S.; Ungarreevittaya, P.; Midorikawa, K.; Hiraku, Y.; Oikawa, S.; Kawanishi, S.; Murata, M. Overexpression of CD44 Variant 9: A Novel Cancer Stem Cell Marker in Human Cholangiocarcinoma in Relation to Inflammation. Mediat. Inflamm. 2018, 2018, 4867234. [Google Scholar]
- Suwannakul, N.; Ma, N.; Midorikawa, K.; Oikawa, S.; Kobayashi, H.; He, F.; Kawanishi, S.; Murata, M. CD44v9 Induces Stem Cell-Like Phenotypes in Human Cholangiocarcinoma. Front. Cell Dev. Biol. 2020, 8, 417. [Google Scholar]
- Thanan, R.; Murata, M.; Ma, N.; Hammam, O.; Wishahi, M.; El Leithy, T.; Hiraku, Y.; Oikawa, S.; Kawanishi, S. Nuclear localization of COX-2 in relation to the expression of stemness markers in urinary bladder cancer. Mediat. Inflamm. 2012, 2012, 165879. [Google Scholar]
- Wang, S.; Ma, N.; Zhao, W.; Midorikawa, K.; Kawanishi, S.; Hiraku, Y.; Oikawa, S.; Zhang, Z.; Huang, G.; Murata, M. Inflammation-Related DNA Damage and Cancer Stem Cell Markers in Nasopharyngeal Carcinoma. Mediat. Inflamm. 2016, 2016, 9343460. [Google Scholar]
- Thanan, R.; Ma, N.; Hiraku, Y.; Iijima, K.; Koike, T.; Shimosegawa, T.; Murata, M.; Kawanishi, S. DNA Damage in CD133-Positive Cells in Barrett’s Esophagus and Esophageal Adenocarcinoma. Mediat. Inflamm. 2016, 2016, 7937814. [Google Scholar]
- Omidvar, S.; Vahedian, V.; Sourani, Z.; Yari, D.; Asadi, M.; Jafari, N.; Khodavirdilou, L.; Bagherieh, M.; Shirzad, M.; Hosseini, V. The molecular crosstalk between innate immunity and DNA damage repair/response: Interactions and effects in cancers. Pathol. Res. Pract. 2024, 260, 155405. [Google Scholar]
- Jiang, M.; Jia, K.; Wang, L.; Li, W.; Chen, B.; Liu, Y.; Wang, H.; Zhao, S.; He, Y.; Zhou, C. Alterations of DNA damage response pathway: Biomarker and therapeutic strategy for cancer immunotherapy. Acta Pharm. Sinica. B 2021, 11, 2983–2994. [Google Scholar]
- Shi, Y.; Gilkes, D.M. HIF-1 and HIF-2 in cancer: Structure, regulation, and therapeutic prospects. Cell. Mol. Life Sci. CMLS 2025, 82, 44. [Google Scholar] [PubMed]
- Han, Y.; Sheng, W.; Liu, X.; Liu, H.; Jia, X.; Li, H.; Wang, C.; Wang, B.; Hu, T.; Ma, Y. Glycyrrhizin ameliorates colorectal cancer progression by regulating NHEJ pathway through inhibiting HMGB1-induced DNA damage response. Sci. Rep. 2024, 14, 24948. [Google Scholar]
- Lv, G.; Yang, M.; Gai, K.; Jia, Q.; Wang, Z.; Wang, B.; Li, X. Multiple functions of HMGB1 in cancer. Front. Oncol. 2024, 14, 1384109. [Google Scholar]
- Fan, A.; Gao, M.; Tang, X.; Jiao, M.; Wang, C.; Wei, Y.; Gong, Q.; Zhong, J. HMGB1/RAGE axis in tumor development: Unraveling its significance. Front. Oncol. 2024, 14, 1336191. [Google Scholar]
- Taneja, S.; Vetter, S.W.; Leclerc, E. Hypoxia and the Receptor for Advanced Glycation End Products (RAGE) Signaling in Cancer. Int. J. Mol. Sci. 2021, 22, 8153. [Google Scholar] [CrossRef]
- Idoudi, S.; Bedhiafi, T.; Pedersen, S.; Elahtem, M.; Alremawi, I.; Akhtar, S.; Dermime, S.; Merhi, M.; Uddin, S. Role of HMGB1 and its associated signaling pathways in human malignancies. Cell. Signal. 2023, 112, 110904. [Google Scholar]
- Wang, G.; Hiramoto, K.; Ma, N.; Yoshikawa, N.; Ohnishi, S.; Murata, M.; Kawanishi, S. Glycyrrhizin Attenuates Carcinogenesis by Inhibiting the Inflammatory Response in a Murine Model of Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 2609. [Google Scholar] [CrossRef]
- Wang, G.; Hiramoto, K.; Ma, N.; Ohnishi, S.; Morita, A.; Xu, Y.; Yoshikawa, N.; Chinzei, Y.; Murata, M.; Kawanishi, S. Immunohistochemical analyses reveal FoxP3 expressions in spleen and colorectal cancer in mice treated with AOM/DSS, and their suppression by glycyrrhizin. PLoS ONE 2024, 19, e0307038. [Google Scholar]
- Sun, S.; Ma, J.; Xie, P.; Wu, Z.; Tian, X. Hypoxia-responsive miR-141-3p is involved in the progression of breast cancer via mediating the HMGB1/HIF-1α signaling pathway. J. Gene Med. 2020, 22, e3230. [Google Scholar]
- Songjang, W.; Nensat, C.; Jitpewngarm, W.; Jiraviriyakul, A. Potential Serum HMGB1, HSP90, and S100A9 as Metastasis Predictive Biomarkers for Cancer Patients and Relevant Cytokines: A Pilot Study. Int. J. Mol. Sci. 2024, 25, 13232. [Google Scholar] [CrossRef]
- Vladimirova, D.; Staneva, S.; Ugrinova, I. Multifaceted role of HMGB1: From nuclear functions to cytoplasmic and extracellular signaling in inflammation and cancer-Review. Adv. Protein Chem. Struct. Biol. 2025, 143, 271–300. [Google Scholar] [PubMed]
- Ohnishi, S.; Hiramoto, K.; Ma, N.; Kawanishi, S. Chemoprevention by aspirin against inflammation-related colorectal cancer in mice. J. Clin. Biochem. Nutr. 2021, 69, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.R.D.; Oliveira, L.D.D.; Milan, T.M.; Eskenazi, A.P.E.; Bighetti-Trevisan, R.L.; de Almeida, O.G.G.; Amorim, M.L.M.; Squarize, C.H.; Castilho, R.M.; Almeida, L.O.D. Up-regulation of TNF-alpha/NFkB/SIRT1 axis drives aggressiveness and cancer stem cells accumulation in chemoresistant oral squamous cell carcinoma. J. Cell. Physiol. 2024, 239, e31164. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhu, Y.; Shang, L.; Qiu, Y.; Shen, N.; Wang, J.; Adam, T.; Wei, W.; Song, Q.; Li, J.; et al. LncRNA XIST regulates breast cancer stem cells by activating proinflammatory IL-6/STAT3 signaling. Oncogene 2023, 42, 1419–1437. [Google Scholar] [CrossRef]
- Shigdar, S.; Li, Y.; Bhattacharya, S.; O’Connor, M.; Pu, C.; Lin, J.; Wang, T.; Xiang, D.; Kong, L.; Wei, M.Q.; et al. Inflammation and cancer stem cells. Cancer Lett. 2014, 345, 271–278. [Google Scholar] [CrossRef]
- Zhang, Z.; Sha, W. MicroRNA-513b-5p inhibits epithelial mesenchymal transition of colon cancer stem cells through IL-6/STAT3 signaling pathway. Discov. Oncol. 2024, 15, 267. [Google Scholar] [CrossRef]
- Karin, M.; Clevers, H. Reparative inflammation takes charge of tissue regeneration. Nature 2016, 529, 307–315. [Google Scholar] [CrossRef]
- Sadhukhan, P.; Feng, M.; Illingworth, E.; Sloma, I.; Ooki, A.; Matoso, A.; Sidransky, D.; Johnson, B.A., 3rd; Marchionni, L.; Sillé, F.C.; et al. YAP1 induces bladder cancer progression and promotes immune evasion through IL-6/STAT3 pathway and CXCL deregulation. J. Clin. Investig. 2024, 135, e171164. [Google Scholar] [CrossRef]
- Wang, G.; Ma, N.; He, F.; Kawanishi, S.; Kobayashi, H.; Oikawa, S.; Murata, M. Taurine Attenuates Carcinogenicity in Ulcerative Colitis-Colorectal Cancer Mouse Model. Oxid. Med. Cell. Longev. 2020, 2020, 7935917. [Google Scholar] [CrossRef]
- Cheng, R.Y.S.; Burkett, S.; Ambs, S.; Moody, T.; Wink, D.A.; Ridnour, L.A. Chronic Exposure to Nitric Oxide Induces P53 Mutations and Malignant-like Features in Human Breast Epithelial Cells. Biomolecules 2023, 13, 311. [Google Scholar] [CrossRef]
- Kawanishi, S.; Ohnishi, S.; Ma, N.; Hiraku, Y.; Oikawa, S.; Murata, M. Nitrative and oxidative DNA damage in infection-related carcinogenesis in relation to cancer stem cells. Genes. Environ. 2016, 38, 26. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Semenza, G.L. Master Role of Hypoxia in Cancer Progression: Major Insights During ISOTT’s Half-Century. Adv. Exp. Med. Biol. 2024, 1463, 15–20. [Google Scholar] [PubMed]
- Castillo-Rodríguez, R.A.; Trejo-Solís, C.; Cabrera-Cano, A.; Gómez-Manzo, S.; Dávila-Borja, V.M. Hypoxia as a Modulator of Inflammation and Immune Response in Cancer. Cancers 2022, 14, 2291. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Simińska, D.; Gąssowska-Dobrowolska, M.; Listos, J.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Chronic and Cycling Hypoxia: Drivers of Cancer Chronic Inflammation through HIF-1 and NF-κB Activation: A Review of the Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 10701. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y. HMGB1 in inflammation and cancer. J. Hematol. Oncol. 2020, 13, 116. [Google Scholar] [CrossRef]
- Tian, Z.; Zhu, L.; Xie, Y.; Hu, H.; Ren, Q.; Liu, J.; Wang, Q. The mechanism of high mobility group box-1 protein and its bidirectional regulation in tumors. Biomol. Biomed. 2024, 24, 477–485. [Google Scholar] [CrossRef]
- Chu, X.; Tian, W.; Ning, J.; Xiao, G.; Zhou, Y.; Wang, Z.; Zhai, Z.; Tanzhu, G.; Yang, J.; Zhou, R. Cancer stem cells: Advances in knowledge and implications for cancer therapy. Signal Transduct. Target. Ther. 2024, 9, 170. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Agents | Ref. | Samples | 8-NitroG | 8-OxodG | PCNA | p16 | p53 | HIF-1α |
|---|---|---|---|---|---|---|---|---|
| OV | [6] | CCA tissues vs adjacent non-cancer tissues | ** | ** | – | – | – | – |
| H. pylori | [14] | Gastritis with HP vs without HP | ** | *** | ** | – | – | – |
| [16] | Gastritis with HP before/after eradication | ** | * | – | – | – | – | |
| HCV | [17] | Hepatitis C before/after INF (responder) | * | * | – | – | – | – |
| HPV | [21] | CIN1 vs condyloma | * | ns | ** | ns | – | – |
| Leukoplakia | [31] | Leukoplakia vs normal mucosa | *** | *** | ns | – | * | * |
| UC | [34] | UCAC samples vs UC samples | **** | ** | – | – | – | – |
| BE | [36] | BEA samples vs BE samples | * | * | – | – | – | – |
| BE samples before/after PPI | ** | ** | – | – | – | – | ||
| MFH | [38] | Samples from deseased vs living MFH | *** | – | – | – | – | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawanishi, S.; Wang, G.; Ma, N.; Murata, M. Cancer Development and Progression Through a Vicious Cycle of DNA Damage and Inflammation. Int. J. Mol. Sci. 2025, 26, 3352. https://doi.org/10.3390/ijms26073352
Kawanishi S, Wang G, Ma N, Murata M. Cancer Development and Progression Through a Vicious Cycle of DNA Damage and Inflammation. International Journal of Molecular Sciences. 2025; 26(7):3352. https://doi.org/10.3390/ijms26073352
Chicago/Turabian StyleKawanishi, Shosuke, Guifeng Wang, Ning Ma, and Mariko Murata. 2025. "Cancer Development and Progression Through a Vicious Cycle of DNA Damage and Inflammation" International Journal of Molecular Sciences 26, no. 7: 3352. https://doi.org/10.3390/ijms26073352
APA StyleKawanishi, S., Wang, G., Ma, N., & Murata, M. (2025). Cancer Development and Progression Through a Vicious Cycle of DNA Damage and Inflammation. International Journal of Molecular Sciences, 26(7), 3352. https://doi.org/10.3390/ijms26073352