
Mitochondrial Changes Induced by SGLT2i in Lymphocytes from Diabetic Kidney Transplant Recipients: A Pilot Study

 ,
,  , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Results
2.1. Description of the Studied Cohort
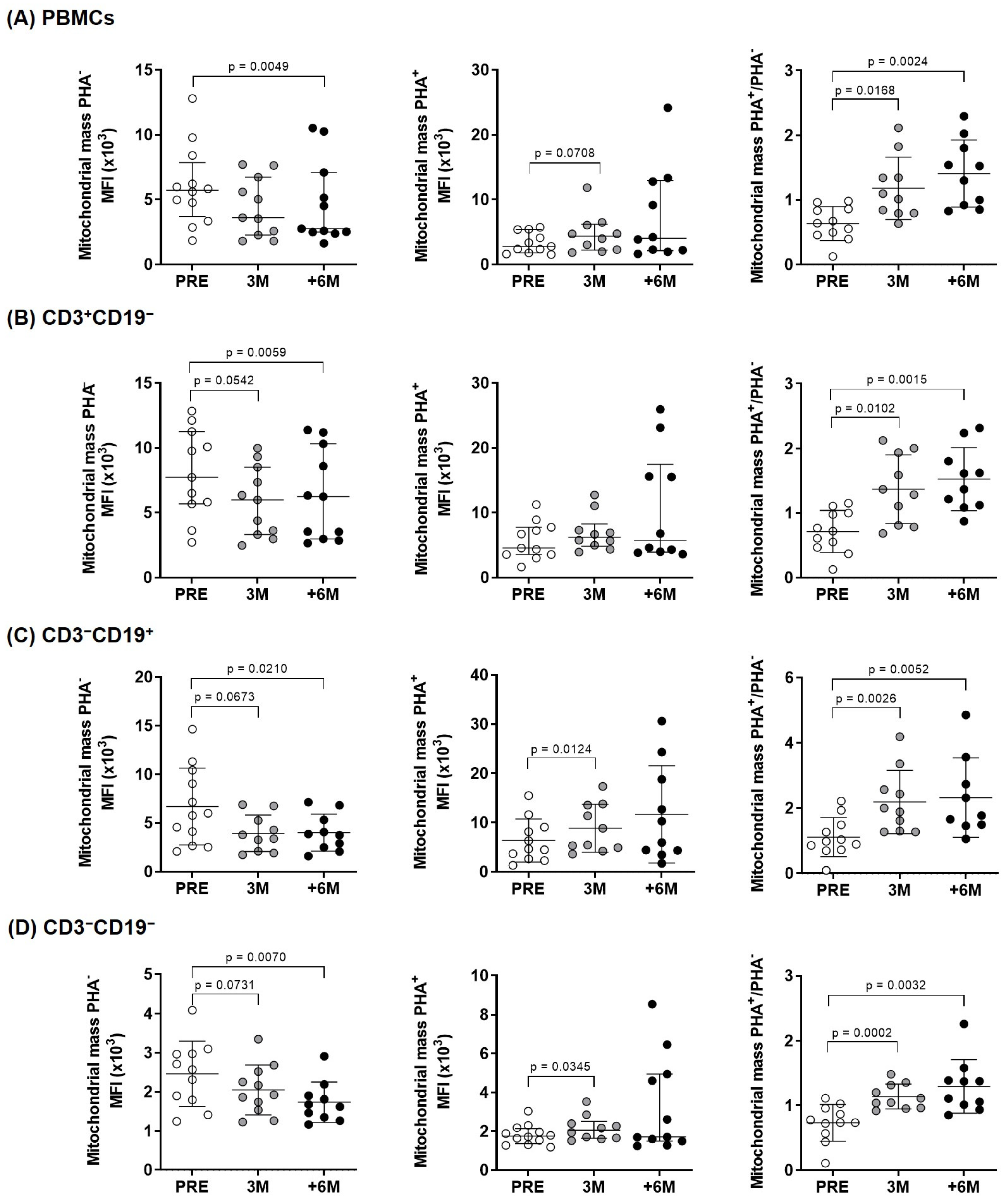
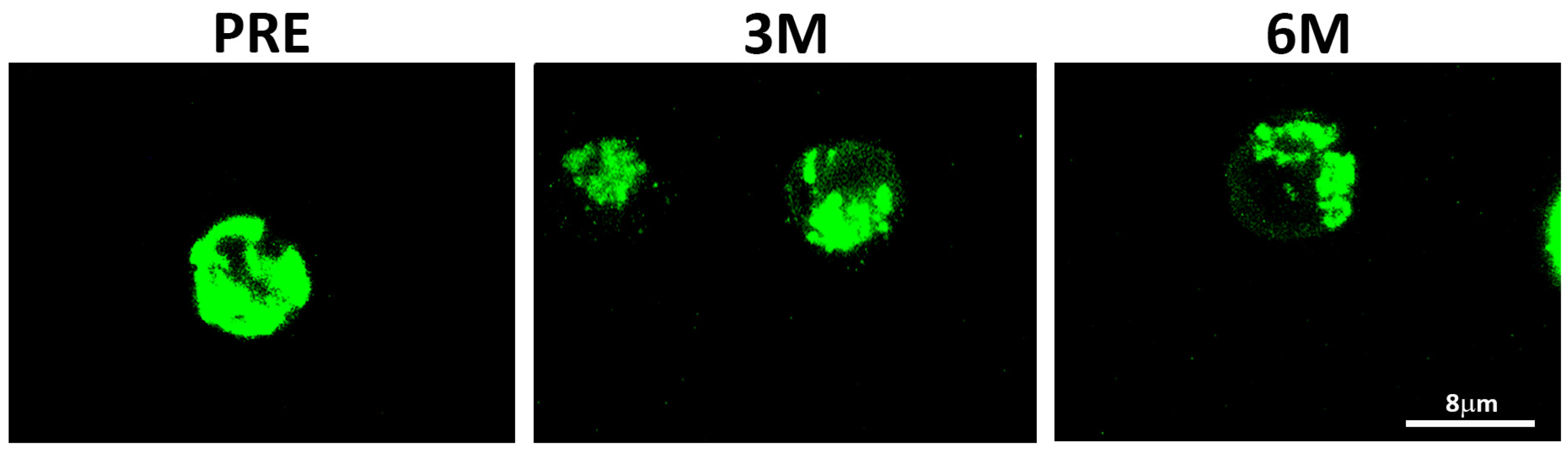
2.2. Upon PHA-Stimulation, SGLT2i Treatment Induces an Increase in Mitochondrial Mass and Membrane Potential, but Not in ROS Content
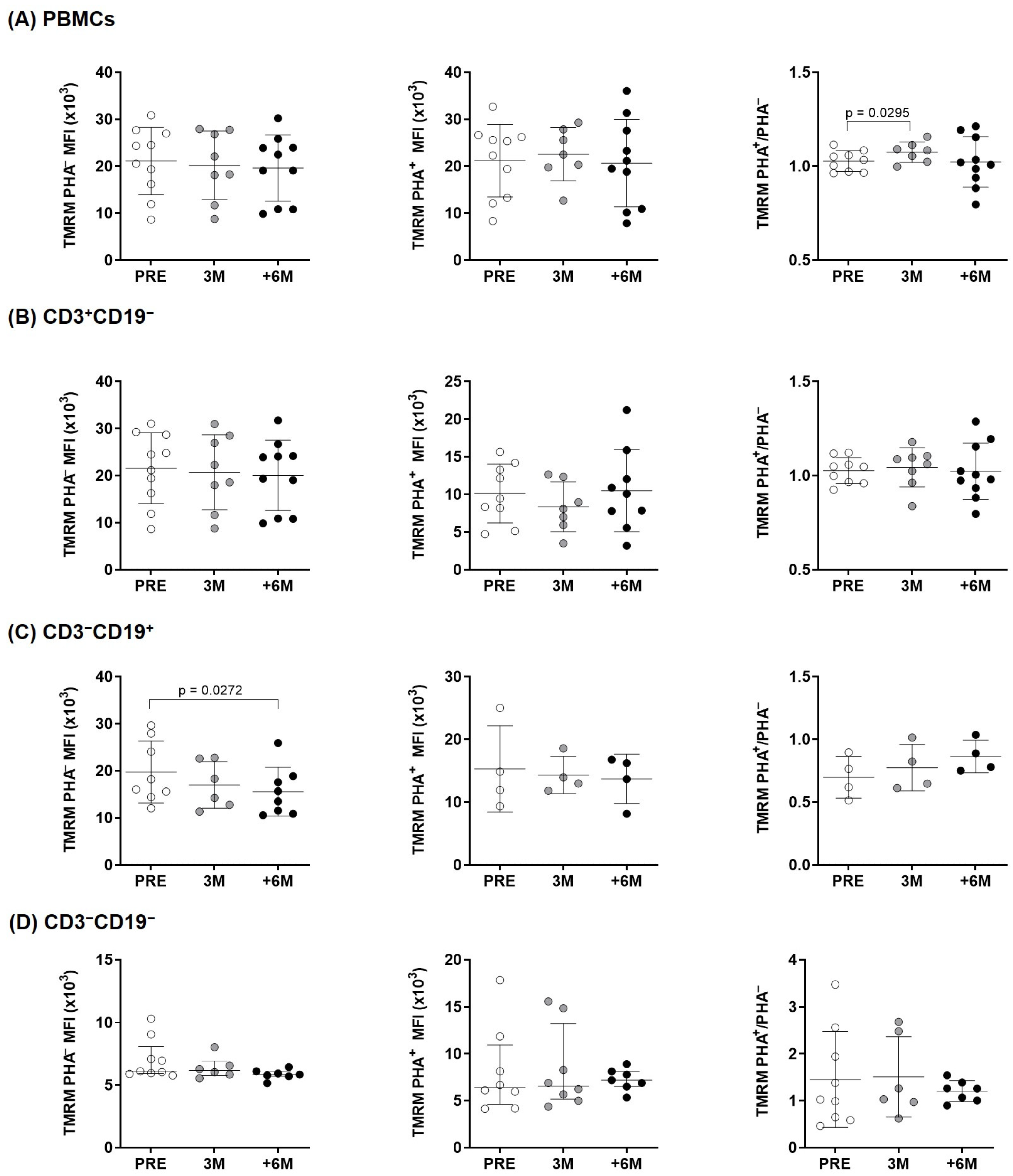
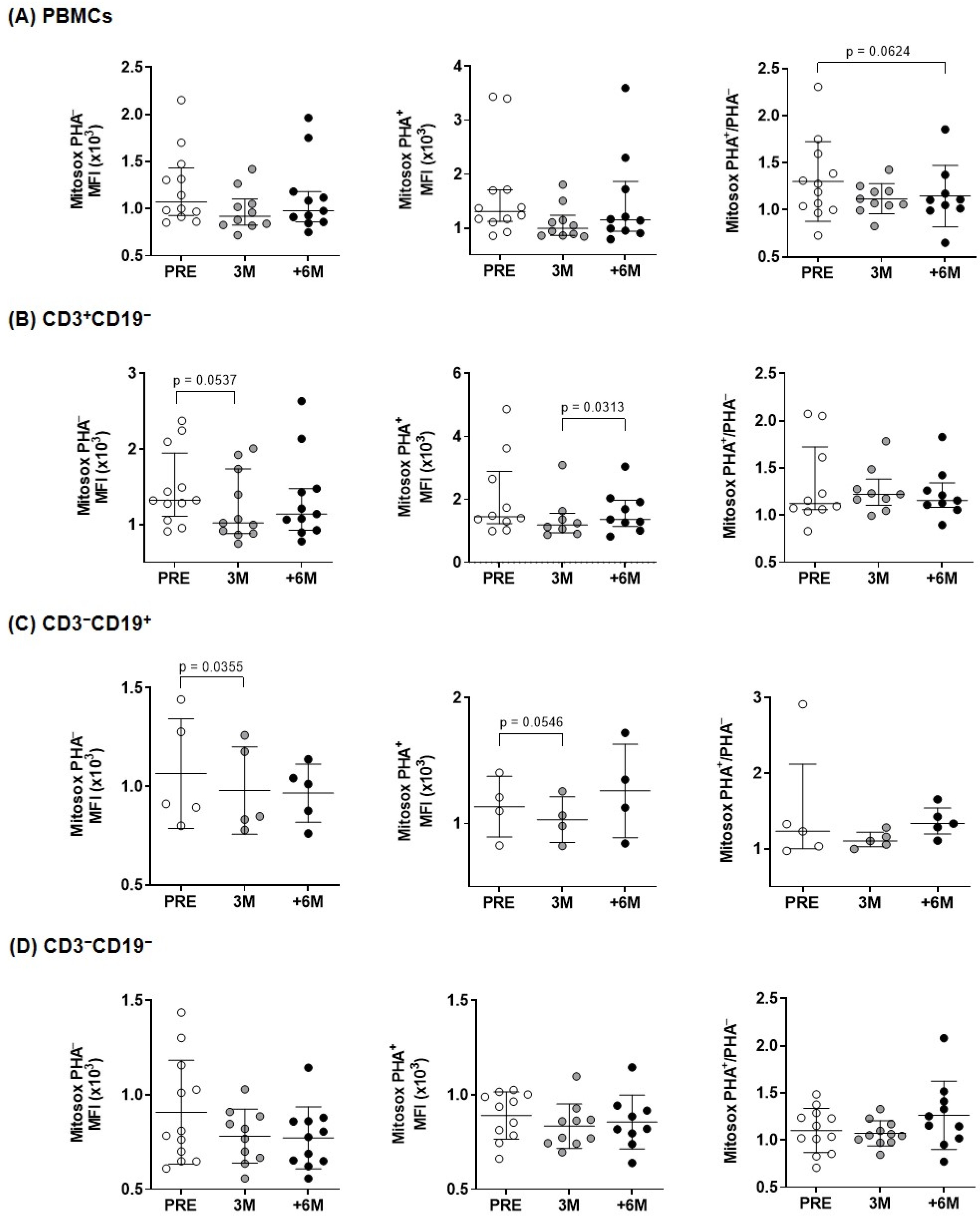
2.3. Mitochondrial Activity After SGLT2i Therapy
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Sample Collection
4.3. Cell Metabolism
4.4. Flow Cytometry
4.5. Confocal Microscopy
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| +6M | Six or more months after the establishment SGLT2i treatment |
| 3M | Three months after the establishment SGLT2i treatment |
| CKD | Chronic kidney disease |
| CVD | Cardiovascular disease |
| DBP | Diastolic blood pressures |
| DMSO | Dimethylsulfoxide |
| ECAR | Extracellular acidification rate |
| ETC | Electron transport chain |
| eGFR | Glomerular filtration rate |
| FCCP | Carbonylcyanide-p-trifluoromethoxyphenylhydrazone |
| PBMCs | Peripheral blood mononuclear cells |
| PHA | Phytohemagglutinin |
| PRE | Before the establishment SGLT2i treatment |
| OCR | Oxygen consumption rate |
| OXPHOS | Oxidative phosphorylation |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| SBP | Systolic blood pressures |
| SGLT2 | Sodium-glucose co-transporter 2 |
| SGLT2i | Sodium-glucose co-transporter 2 inhibitors |
| T2D | Type 2 diabetes |
| TMRM | Tetramethylrhodamine |
References
- Zinman, B.; Wanner, C.; Lachin, J.M.; Fitchett, D.; Bluhmki, E.; Hantel, S.; Mattheus, M.; Devins, T.; Johansen, O.E.; Woerle, H.J.; et al. Empagliflozin, Cardiovascular Outcomes, and Mortality in Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 2117–2128. [Google Scholar] [CrossRef]
- Voors, A.A.; Angermann, C.E.; Teerlink, J.R.; Collins, S.P.; Kosiborod, M.; Biegus, J.; Ferreira, J.P.; Nassif, M.E.; Psotka, M.A.; Tromp, J.; et al. The SGLT2 Inhibitor Empagliflozin in Patients Hospitalized for Acute Heart Failure: A Multinational Randomized Trial. Nat. Med. 2022, 28, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.K.; Strong, J. The Pleiotropic Effects of Sodium-Glucose Cotransporter-2 Inhibitors: Beyond the Glycemic Benefit. Diabetes Ther. 2019, 10, 1771–1792. [Google Scholar] [CrossRef] [PubMed]
- Zannad, F.; Ferreira, J.P.; Pocock, S.J.; Zeller, C.; Anker, S.D.; Butler, J.; Filippatos, G.; Hauske, S.J.; Brueckmann, M.; Pfarr, E.; et al. Cardiac and Kidney Benefits of Empagliflozin in Heart Failure Across the Spectrum of Kidney Function: Insights From EMPEROR-Reduced. Circulation 2021, 143, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial Energetics in the Kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef]
- Forbes, J.M.; Thorburn, D.R. Mitochondrial Dysfunction in Diabetic Kidney Disease. Nat. Rev. Nephrol. 2018, 14, 291–312. [Google Scholar] [CrossRef]
- Lu, Y.P.; Zhang, Z.Y.; Wu, H.W.; Fang, L.J.; Hu, B.; Tang, C.; Zhang, Y.Q.; Yin, L.; Tang, D.E.; Zheng, Z.H.; et al. SGLT2 Inhibitors Improve Kidney Function and Morphology by Regulating Renal Metabolic Reprogramming in Mice with Diabetic Kidney Disease. J. Transl. Med. 2022, 20, 420. [Google Scholar] [CrossRef]
- Rastogi, A.; Januzzi, J.L. Pleiotropic Effects of Sodium-Glucose Cotransporter-2 Inhibitors in Cardiovascular Disease and Chronic Kidney Disease. J. Clin. Med. 2023, 12, 2824. [Google Scholar] [CrossRef]
- Packer, M. Critical Reanalysis of the Mechanisms Underlying the Cardiorenal Benefits of SGLT2 Inhibitors and Reaffirmation of the Nutrient Deprivation Signaling/Autophagy Hypothesis. Circulation 2022, 146, 1383–1405. [Google Scholar] [CrossRef]
- IDF Diabetes Atlas. Available online: https://diabetesatlas.org/citation-usage/ (accessed on 14 January 2025).
- Sharif, A.; Chakkera, H.; de Vries, A.P.J.; Eller, K.; Guthoff, M.; Haller, M.C.; Hornum, M.; Nordheim, E.; Kautzky-Willer, A.; Krebs, M.; et al. International Consensus on Post-Transplantation Diabetes Mellitus. Nephrol. Dial. Transpl. 2024, 39, 531–549. [Google Scholar] [CrossRef]
- Nash, A.; Samoylova, M.; Leuthner, T.; Zhu, M.; Lin, L.; Meyer, J.N.; Brennan, T.V. Effects of Immunosuppressive Medications on Mitochondrial Function. J. Surg. Res. 2020, 249, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.Y.; Chang, L.Y.; Chen, J.Y.; Pan, H.C.; Tseng, C.S.; Chueh, J.S.; Wu, V.C. The Outcomes of SGLT-2 Inhibitor Utilization in Diabetic Kidney Transplant Recipients. Nat. Commun. 2024, 15, 10043. [Google Scholar] [CrossRef]
- Joshi, S.S.; Singh, T.; Newby, D.E.; Singh, J. Sodium-Glucose Co-Transporter 2 Inhibitor Therapy: Mechanisms of Action in Heart Failure. Heart 2021, 107, 1032–1038. [Google Scholar] [CrossRef]
- Ravindran, S.; Munusamy, S. Renoprotective Mechanisms of Sodium-Glucose Co-Transporter 2 (SGLT2) Inhibitors against the Progression of Diabetic Kidney Disease. J. Cell Physiol. 2022, 237, 1182–1205. [Google Scholar] [CrossRef] [PubMed]
- Vallon, V.; Platt, K.A.; Cunard, R.; Schroth, J.; Whaley, J.; Thomson, S.C.; Koepsell, H.; Rieg, T. SGLT2 Mediates Glucose Reabsorption in the Early Proximal Tubule. J. Am. Soc. Nephrol. 2011, 22, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Zügner, E.; Yang, H.C.; Kotzbeck, P.; Boulgaropoulos, B.; Sourij, H.; Hagvall, S.; Elmore, C.S.; Esterline, R.; Moosmang, S.; Oscarsson, J.; et al. Differential In Vitro Effects of SGLT2 Inhibitors on Mitochondrial Oxidative Phosphorylation, Glucose Uptake and Cell Metabolism. Int. J. Mol. Sci. 2022, 23, 7966. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Takagi, S.; Nitta, K.; Kitada, M.; Srivastava, S.P.; Takagaki, Y.; Kanasaki, K.; Koya, D. Renal Protective Effects of Empagliflozin via Inhibition of EMT and Aberrant Glycolysis in Proximal Tubules. JCI Insight 2020, 5, 129034. [Google Scholar] [CrossRef]
- Li, X.; Preckel, B.; Hermanides, J.; Hollmann, M.W.; Zuurbier, C.J.; Weber, N.C. Amelioration of Endothelial Dysfunction by Sodium Glucose Co-Transporter 2 Inhibitors: Pieces of the Puzzle Explaining Their Cardiovascular Protection. Br. J. Pharmacol. 2022, 179, 4047–4062. [Google Scholar] [CrossRef]
- Sridharan, K.; Sivaramakrishnan, G. Sodium Glucose Cotransporter-2 Inhibitors Improve Endothelial Function and Arterial Stiffness in Diabetic Individuals: A Systematic Review and Network Meta-Analysis. Curr. Vasc. Pharmacol. 2025; online ahead of print. [Google Scholar] [CrossRef]
- Buck, M.D.; O’Sullivan, D.; Pearce, E.L. T Cell Metabolism Drives Immunity. J. Exp. Med. 2015, 212, 1345–1360. [Google Scholar] [CrossRef]
- Kishi, S.; Nagasu, H.; Kidokoro, K.; Kashihara, N. Oxidative Stress and the Role of Redox Signalling in Chronic Kidney Disease. Nat. Rev. Nephrol. 2024, 20, 101–119. [Google Scholar] [CrossRef]
- Geltink, R.I.K.; Kyle, R.L.; Pearce, E.L. Unraveling the Complex Interplay Between T Cell Metabolism and Function. Annu. Rev. Immunol. 2018, 36, 461–488. [Google Scholar] [CrossRef] [PubMed]
- Rajasekeran, H.; Lytvyn, Y.; Cherney, D.Z.I. Sodium-Glucose Cotransporter 2 Inhibition and Cardiovascular Risk Reduction in Patients with Type 2 Diabetes: The Emerging Role of Natriuresis. Kidney Int. 2016, 89, 524–526. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Reeves, W.B.; Awad, A.S. Pathophysiology of Diabetic Kidney Disease: Impact of SGLT2 Inhibitors. Nat. Rev. Nephrol. 2021, 17, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Pantanetti, P.; Cangelosi, G.; Alberti, S.; Di Marco, S.; Michetti, G.; Cerasoli, G.; Di Giacinti, M.; Coacci, S.; Francucci, N.; Petrelli, F.; et al. Changes in Body Weight and Composition, Metabolic Parameters, and Quality of Life in Patients with Type 2 Diabetes Treated with Subcutaneous Semaglutide in Real-World Clinical Practice. Front. Endocrinol. 2024, 15, 1394506. [Google Scholar] [CrossRef]
- Doehner, W.; Anker, S.D.; Butler, J.; Zannad, F.; Filippatos, G.; Coats, A.J.S.; Ferreira, J.P.; Henrichmoeller, I.; Brueckmann, M.; Schueler, E.; et al. Uric Acid and SGLT2 Inhibition With Empagliflozin in Heart Failure With Preserved Ejection Fraction: The EMPEROR-Preserved Trial. JACC Hear Fail. 2024, 12, 2057–2070. [Google Scholar] [CrossRef]
- Sun, X.; Wang, G. Renal Outcomes with Sodium-Glucose Cotransporters 2 Inhibitors. Front. Endocrinol. 2022, 13, 1063341. [Google Scholar] [CrossRef]
- Shao, Q.; Meng, L.; Lee, S.; Tse, G.; Gong, M.; Zhang, Z.; Zhao, J.; Zhao, Y.; Li, G.; Liu, T. Empagliflozin, a Sodium Glucose Co-Transporter-2 Inhibitor, Alleviates Atrial Remodeling and Improves Mitochondrial Function in High-Fat Diet/Streptozotocin-Induced Diabetic Rats. Cardiovasc. Diabetol. 2019, 18, 165. [Google Scholar] [CrossRef]
- Lackey, D.E.; Olefsky, J.M. Regulation of Metabolism by the Innate Immune System. Nat. Rev. Endocrinol. 2016, 12, 15–20. [Google Scholar] [CrossRef]
- van Exel, E.; Gussekloo, J.; de Craen, A.J.; Frölich, M.; der Wiel, A.B.-V.; Westendorp, R.G.J. Low Production Capacity of Interleukin-10 Associates with the Metabolic Syndrome and Type 2 Diabetes: The Leiden 85-Plus Study. Diabetes 2002, 51, 1088–1092. [Google Scholar] [CrossRef]
- Jenkins, B.J.; Blagih, J.; Ponce-Garcia, F.M.; Canavan, M.; Gudgeon, N.; Eastham, S.; Hill, D.; Hanlon, M.M.; Ma, E.H.; Bishop, E.L.; et al. Canagliflozin Impairs T Cell Effector Function via Metabolic Suppression in Autoimmunity. Cell Metab. 2023, 35, 1132–1146.e9. [Google Scholar] [CrossRef]
- Bantug, G.R.; Galluzzi, L.; Kroemer, G.; Hess, C. The Spectrum of T Cell Metabolism in Health and Disease. Nat. Rev. Immunol. 2018, 18, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Maejima, Y. SGLT2 Inhibitors Play a Salutary Role in Heart Failure via Modulation of the Mitochondrial Function. Front. Cardiovasc. Med. 2020, 6, 186. [Google Scholar] [CrossRef]
- Canet, F.; Iannantuoni, F.; de Marañon, A.M.; Díaz-Pozo, P.; López-Domènech, S.; Vezza, T.; Navarro, B.; Solá, E.; Falcón, R.; Bañuls, C.; et al. Does Empagliflozin Modulate Leukocyte-Endothelium Interactions, Oxidative Stress, and Inflammation in Type 2 Diabetes? Antioxidants 2021, 10, 1228. [Google Scholar] [CrossRef]
- Iannantuoni, F.; de Marañon, A.M.; Abad-Jiménez, Z.; Canet, F.; Díaz-Pozo, P.; López-Domènech, S.; Morillas, C.; Rocha, M.; Víctor, V.M. Mitochondrial Alterations and Enhanced Human Leukocyte/Endothelial Cell Interactions in Type 1 Diabetes. J. Clin. Med. 2020, 9, 2155. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Jin, L.; Luo, K.; Lim, S.W.; Chung, B.H.; Yang, C.W. Effect of Empagliflozin on Tacrolimus-Induced Pancreas Islet Dysfunction and Renal Injury. Am. J. Transpl. 2017, 17, 2601–2616. [Google Scholar] [CrossRef]
- Koizumi, T.; Watanabe, M.; Yokota, T.; Tsuda, M.; Handa, H.; Koya, J.; Nishino, K.; Tatsuta, D.; Natsui, H.; Kadosaka, T.; et al. Empagliflozin Suppresses Mitochondrial Reactive Oxygen Species Generation and Mitigates the Inducibility of Atrial Fibrillation in Diabetic Rats. Front. Cardiovasc. Med. 2023, 10, 1005408. [Google Scholar] [CrossRef]
- Mizuno, M.; Kuno, A.; Yano, T.; Miki, T.; Oshima, H.; Sato, T.; Nakata, K.; Kimura, Y.; Tanno, M.; Miura, T. Empagliflozin Normalizes the Size and Number of Mitochondria and Prevents Reduction in Mitochondrial Size after Myocardial Infarction in Diabetic Hearts. Physiol. Rep. 2018, 6, e13741. [Google Scholar] [CrossRef]
- Fontecha-barriuso, M.; Martin-sanchez, D.; Martinez-moreno, J.M.; Monsalve, M.; Ramos, A.M.; Sanchez-niño, M.D.; Ruiz-ortega, M.; Ortiz, A.; Sanz, A.B. The Role of PGC-1α and Mitochondrial Biogenesis in Kidney Diseases. Biomolecules 2020, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Lopaschuk, G.D.; Verma, S. Mechanisms of Cardiovascular Benefits of Sodium Glucose Co-Transporter 2 (SGLT2) Inhibitors: A State-of-the-Art Review. JACC. Basic Transl. Sci. 2020, 5, 632–644. [Google Scholar] [CrossRef]
- Packer, M. SGLT2 Inhibitors: Role in Protective Reprogramming of Cardiac Nutrient Transport and Metabolism. Nat. Rev. Cardiol. 2023, 20, 443–462. [Google Scholar] [CrossRef]
- Scheen, A.J.; Delanaye, P. Understanding the Protective Effects of SGLT2 Inhibitors in Type 2 Diabetes Patients with Chronic Kidney Disease. Expert Rev. Endocrinol. Metab. 2022, 17, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Packer, M.; Ferreira, J.P.; Butler, J.; Filippatos, G.; Januzzi, J.L.; González Maldonado, S.; Panova-Noeva, M.; Pocock, S.J.; Prochaska, J.H.; Saadati, M.; et al. Reaffirmation of Mechanistic Proteomic Signatures Accompanying SGLT2 Inhibition in Patients With Heart Failure: A Validation Cohort of the EMPEROR Program. J. Am. Coll. Cardiol. 2024, 84, 1979–1994. [Google Scholar] [CrossRef]
- Bayes-Genis, A.; Iborra-Egea, O.; Spitaleri, G.; Domingo, M.; Revuelta-López, E.; Codina, P.; Cediel, G.; Santiago-Vacas, E.; Cserkóová, A.; Pascual-Figal, D.; et al. Decoding Empagliflozin’s Molecular Mechanism of Action in Heart Failure with Preserved Ejection Fraction Using Artificial Intelligence. Sci. Rep. 2021, 11, 12025. [Google Scholar] [CrossRef]
- Li, H.; Mai, Z.; Yu, S.; Wang, B.; Lai, W.; Chen, G.; Zhou, C.; Liu, J.; Yang, Y.; Chen, S.; et al. Exploring the Pleiotropic Genes and Therapeutic Targets Associated with Heart Failure and Chronic Kidney Disease by Integrating MetaCCA and SGLT2 Inhibitors’ Target Prediction. Biomed. Res. Int. 2021, 2021, 4229194. [Google Scholar] [CrossRef]
- Wang, A.; Li, Z.; Zhuo, S.; Gao, F.; Zhang, H.; Zhang, Z.; Ren, G.; Ma, X. Mechanisms of Cardiorenal Protection With SGLT2 Inhibitors in Patients With T2DM Based on Network Pharmacology. Front. Cardiovasc. Med. 2022, 9, 857952. [Google Scholar] [CrossRef]
- Bettencourt, I.A.; Powell, J.D. Targeting Metabolism as a Novel Therapeutic Approach to Autoimmunity, Inflammation, and Transplantation. J. Immunol. 2017, 198, 999–1005. [Google Scholar] [CrossRef]
- Luna-Marco, C.; Iannantuoni, F.; Hermo-Argibay, A.; Devos, D.; Salazar, J.D.; Víctor, V.M.; Rovira-Llopis, S. Cardiovascular Benefits of SGLT2 Inhibitors and GLP-1 Receptor Agonists through Effects on Mitochondrial Function and Oxidative Stress. Free Radic. Biol. Med. 2024, 213, 19–35. [Google Scholar] [CrossRef]
- Pantanetti, P.; Ronconi, V.; Sguanci, M.; Palomares, S.M.; Mancin, S.; Tartaglia, F.C.; Cangelosi, G.; Petrelli, F. Oral Semaglutide in Type 2 Diabetes: Clinical-Metabolic Outcomes and Quality of Life in Real-World Practice. J. Clin. Med. 2024, 13, 4752. [Google Scholar] [CrossRef]
- Scheen, A.J. SGLT2 Inhibitors as Add-On Therapy to Metformin for People with Type 2 Diabetes: A Review of Placebo-Controlled Trials in Asian versus Non-Asian Patients. Diabetes Metab. Syndr. Obes. 2020, 13, 2765–2779. [Google Scholar] [CrossRef]
- O’Hara, D.V.; Lam, C.S.P.; McMurray, J.J.V.; Yi, T.W.; Hocking, S.; Dawson, J.; Raichand, S.; Januszewski, A.S.; Jardine, M.J. Applications of SGLT2 Inhibitors beyond Glycaemic Control. Nat. Rev. Nephrol. 2024, 20, 513–529. [Google Scholar] [CrossRef]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) Statement: Guidelines for Reporting Observational Studies. J. Clin. Epidemiol. 2008, 61, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pyakurel, A.; Glytsou, C.; Scorrano, L. The Cell Biology of Mitochondrial Membrane Dynamics. Nat. Rev. Mol. Cell Biol. 2020, 21, 204–224. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age [Years, Mean (SD)] | 66.4 (10.2) |
| Male, n (%) | 10 (71.4) |
| Months after transplantation, median (IQR) | 103 (48–205) |
| Pre-transplant T2D, n (%) | 4 (28.6) |
| Pre-transplant T2D duration [years, median (IQR)] | 18 (12.9–36.1) |
| Post-transplant T2D, n (%) | 10 (71.4) |
| Post-transplant T2D duration [months, median (IQR)] | 54.7 (16.8–146.9) |
| Immunosuppression Therapy, n (%) | |
| Tacrolimus | 12 (85.7) |
| Sirolimus | 3 (21.4) |
| MPA (Mycophenolic acid) | 8 (57.1) |
| Steroids | 5 (35.7) |
| Antidiabetic agents, n (%) | |
| Insulin | 4 (28.6) |
| Metformin | 4 (28.6) |
| Insulin and Metformin | 2 (14.3) |
| Non-antidiabetic agents | 4 (28.6) |
| SGLT2i, n (%) | |
| Dapaglifozin | 11 (78.6) |
| Empagliglozin | 3 (21.4) |
| −6M | −3M | PRE | +3M | +6M | p1 | p2 | |
|---|---|---|---|---|---|---|---|
| eGFR (mL/min) | 51.5 (25.9) | 48.1 (20.7) | 44.9 (16.6) | 43.7 (14.5) | 43.1 (16.6) | 0.212 | 0.784 |
| uPCR (mg/mg) | 0.20 (0.14–0.48) | 0.18 (0.12–1.00) | 0.22 (0.11–0.57) | 0.33 (0.12–0.56) | 0.21 (0.19–0.49) | 0.723 | 0.212 |
| Fast glycaemia (mg/dL) | 137 (48) | 134 (36) | 130 (30) | 124 (35) | 131 (28) | 0.560 | 0.863 |
| HbA1c (%) | 6.6 (0.8) | 6.4 (0.5) | 6.9 (0.8) | 6.6 (0.6) | 6.7 (0.9) | 0.051 | 0.305 |
| Total cholesterol (mg/dL) | 156 (40) | 148 (22) | 145 (32) | 149 (33) | 137 (37) | 0.508 | 0.501 |
| Triglycerides (mg/dL) | 147 (30) | 214 (103) | 173 (48) | 149 (50) | 179 (88) | 0.270 | 0.460 |
| Body weight (kg) | 78.0 (23.4) | 82.5 (11.5) | 80.9 (15.6) | 79 (15) | 77.4 (15.0) | 0.058 | 0.002 |
| SBP (mmHg) | 130 (14) | 126 (14) | 136 (16) | 134 (17) | 130 (14) | 0.079 | 0.651 |
| Haemoglobin (g/dL) | 13.4 (1.1) | 13.5 (2.3) | 13.4 (1.7) | 13.5 (1.2) | 13.5 (1.7) | 0.555 | 0.732 |
| Serum potassium (mEq/L) | 4.4 (0.7) | 4.1 (0.3) | 4.1 (0.6) | 4.2 (0.3) | 4.3 (0.3) | 0.371 | 0.298 |
| Serum magnesium (mg/dL) | 1.6 (0.3) | 1.7 (0.1) | 1.6 (0.2) | 1.9 (0.2) | 1.9 (0.2) | 0.711 | 0.004 |
| Serum uric acid (mg/dL) | 6.7 (1.4) | 6.2 (1.0) | 6.2 (1.5) | 6.0 (1.5) | 5.7 (1.4) | 0.357 | 0.157 |
| PRE | 3M | +6M | p | |
|---|---|---|---|---|
| PBMCs PHA− | 5716 (3698–7862) | 3603 (2263–6731) | 2753 (2496–7089) | 0.045 |
| PBMCs PHA+ | 2791 (1780–5385) | 4354 (2207–6178) | 4037 (2147–12,930) | 0.717 |
| PBMCs PHA+/PHA− | 0.64 (0.45–0.86) | 1.05 (0.79–1.46) | 1.41 (0.90–1.85) | 0.013 |
| CD3+CD19− PHA− | 7735 (5682–11,262) | 5994 (3321–8523) | 6244 (2981–10,323) | 0.050 |
| CD3+CD19− PHA+ | 4569 (3557–7760) | 6211 (4825–8254) | 5696 (3888–17,468) | 0.895 |
| CD3+CD19− PHA+/PHA− | 0.72 (0.46–1.00) | 1.32 (0.80–1.95) | 1.49 (1.11–1.91) | 0.004 |
| CD3−CD19+ PHA− | 5896 (3021–10,076) | 3792 (2114–6765) | 3816 (2400–5699) | 0.045 |
| CD3−CD19+ PHA+ | 4641 (2623–9087) | 7157 (4753–13,607) | 8115 (4114–20,177) | 0.121 |
| CD3−CD19+ PHA+/PHA− | 1.01 (0.75–1.82) | 1.94 (1.29–2.76) | 1.76 (1.48–2.94) | 0.004 |
| CD3−CD19− PHA− | 2566 (1792–2971) | 1922 (1536–2517) | 1648 (1326–1973) | 0.169 |
| CD3−CD19− PHA+ | 1756 (1380–2133) | 2053 (1648–2512) | 1701 (1439–4692) | 0.368 |
| CD3−CD19− PHA+/PHA− | 0.75 (0.60–0.97) | 1.08 (0.96–1.32) | 1.24 (0.99–1.43) | 0.005 |
| Mitochondrial Mass in: | Hba1c | Body Weight | Total Cholesterol | LDL Cholesterol | HDL Cholesterol |
|---|---|---|---|---|---|
| PBMCs PHA− | r = −0.070 p = 0.828 | r = −0.552 p = 0.063 | r = 0.300 p = 0.370 | r = 0.231 p = 0.471 | r = 0.238 p = 0.456 |
| PBMCs PHA+ | r = −0.728 p = 0.011 | r = 0.173 p = 0.612 | r = −0.536 p = 0.089 | r = −0.727 p = 0.011 | r = −0.018 p = 0.958 |
| PBMCs PHA+/PHA− | r = 0.228 p = 0.475 | r = 0.764 p = 0.006 | r = −0.773 p = 0.005 | r = −0.891 p < 0.001 | r = −0.224 p = 0.484 |
| CD3+CD19− PHA− | r = −0.229 p = 0.499 | r = −0.400 p = 0.223 | r = 0.188 p = 0.603 | r = 0.073 p = 0.832 | r = 0.328 p = 0.325 |
| CD3+CD19− PHA+ | r = −0.563 p = 0.071 | r = 0.291 p = 0.385 | r = −0.382 p = 0.247 | r = −0.518 p = 0.102 | r = −0.173 p = 0.612 |
| CD3+CD19− PHA+/PHA− | r = 0.128 p = 0.724 | r = 0.809 p = 0.003 | r = −0.709 p = 0.015 | r = −0.827 p = 0.002 | r = −0.155 p = 0.650 |
| CD3−CD19+ PHA− | r = 0.074 p= 0.820 | r = −0.350 p = 0.265 | r = 0.236 p = 0.484 | r = 0.049 p = 0.880 | r = 0.452 p = 0.140 |
| CD3−CD19+ PHA+ | r = −0.577 p = 0.063 | r = 0.455 p = 0.160 | r = −0.545 p = 0.083 | r = −0.727 p = 0.011 | r = 0.055 p = 0.873 |
| CD3−CD19+ PHA+/PHA− | r = −0.629 p = 0.028 | r = 0.720 p = 0.008 | r = −0.745 p = 0.008 | r = −0.552 p = 0.063 | r = −0.434 p = 0.158 |
| CD3−CD19− PHA− | r = −0.269 p = 0.423 | r = −0.100 p = 0.770 | r = −0.042 p = 0.907 | r = 0.182 p = 0.593 | r = 0.018 p = 0.958 |
| CD3−CD19− PHA+ | r = −0.696 p = 0.012 | r = 0.217 p = 0.499 | r = −0.518 p = 0.102 | r = −0.350 p = 0.265 | r = −0.256 p = 0.422 |
| CD3−CD19− PHA+/PHA− | r = −0.330 p = 0.294 | r = 0.764 p = 0.006 | r = −0.54 p = 0.071 | r = −0.427 p = 0.167 | r = −0.371 p = 0.235 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Flores, I.; López-Pastor, A.R.; Gómez-Pinedo, U.; Gómez-Infantes, A.; Espino-Paisán, L.; Calvo Romero, N.; Moreno de la Higuera, M.A.; Rodríguez-Cubillo, B.; Gómez-Delgado, I.; Sánchez-Fructuoso, A.I.; et al. Mitochondrial Changes Induced by SGLT2i in Lymphocytes from Diabetic Kidney Transplant Recipients: A Pilot Study. Int. J. Mol. Sci. 2025, 26, 3351. https://doi.org/10.3390/ijms26073351
Pérez-Flores I, López-Pastor AR, Gómez-Pinedo U, Gómez-Infantes A, Espino-Paisán L, Calvo Romero N, Moreno de la Higuera MA, Rodríguez-Cubillo B, Gómez-Delgado I, Sánchez-Fructuoso AI, et al. Mitochondrial Changes Induced by SGLT2i in Lymphocytes from Diabetic Kidney Transplant Recipients: A Pilot Study. International Journal of Molecular Sciences. 2025; 26(7):3351. https://doi.org/10.3390/ijms26073351
Chicago/Turabian StylePérez-Flores, Isabel, Andrea R. López-Pastor, Ulises Gómez-Pinedo, Andrea Gómez-Infantes, Laura Espino-Paisán, Natividad Calvo Romero, M. Angeles Moreno de la Higuera, Beatriz Rodríguez-Cubillo, Irene Gómez-Delgado, Ana I. Sánchez-Fructuoso, and et al. 2025. "Mitochondrial Changes Induced by SGLT2i in Lymphocytes from Diabetic Kidney Transplant Recipients: A Pilot Study" International Journal of Molecular Sciences 26, no. 7: 3351. https://doi.org/10.3390/ijms26073351
APA StylePérez-Flores, I., López-Pastor, A. R., Gómez-Pinedo, U., Gómez-Infantes, A., Espino-Paisán, L., Calvo Romero, N., Moreno de la Higuera, M. A., Rodríguez-Cubillo, B., Gómez-Delgado, I., Sánchez-Fructuoso, A. I., & Urcelay, E. (2025). Mitochondrial Changes Induced by SGLT2i in Lymphocytes from Diabetic Kidney Transplant Recipients: A Pilot Study. International Journal of Molecular Sciences, 26(7), 3351. https://doi.org/10.3390/ijms26073351