
Chronic Administration with FAD012 (3,5-Dimethyl-4-hydroxycinnamic Acid) Maintains Cerebral Blood Flow and Ameliorates Swallowing Dysfunction After Chronic Cerebral Hypoperfusion in Rats

 , ,
, ,
Abstract

1. Introduction
2. Results
2.1. Body Weights and Survival Rates
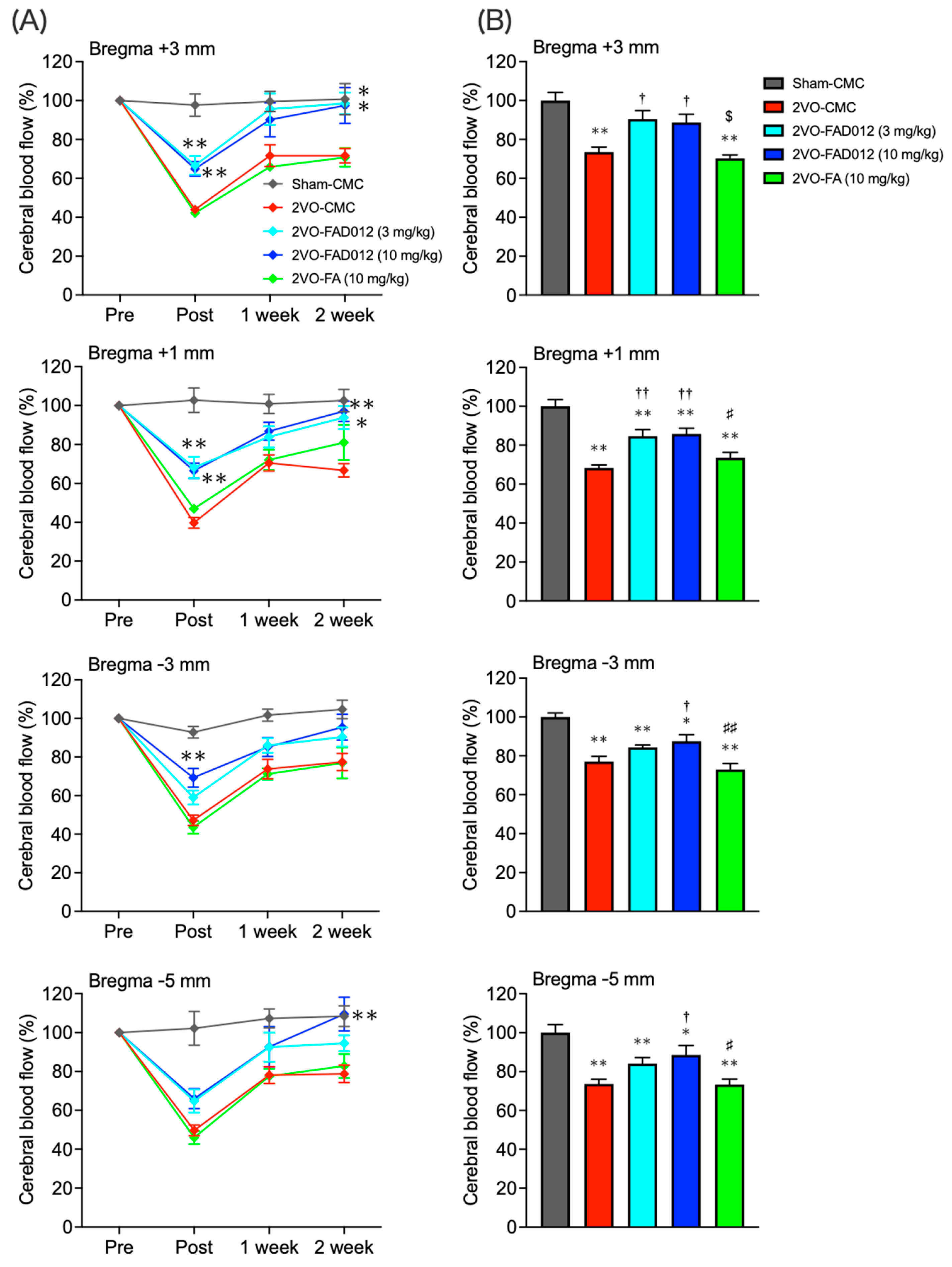
2.2. Changes in Cerebral Blood Flow (CBF) After Bilateral Common Carotid Artery Occlusion (2VO)
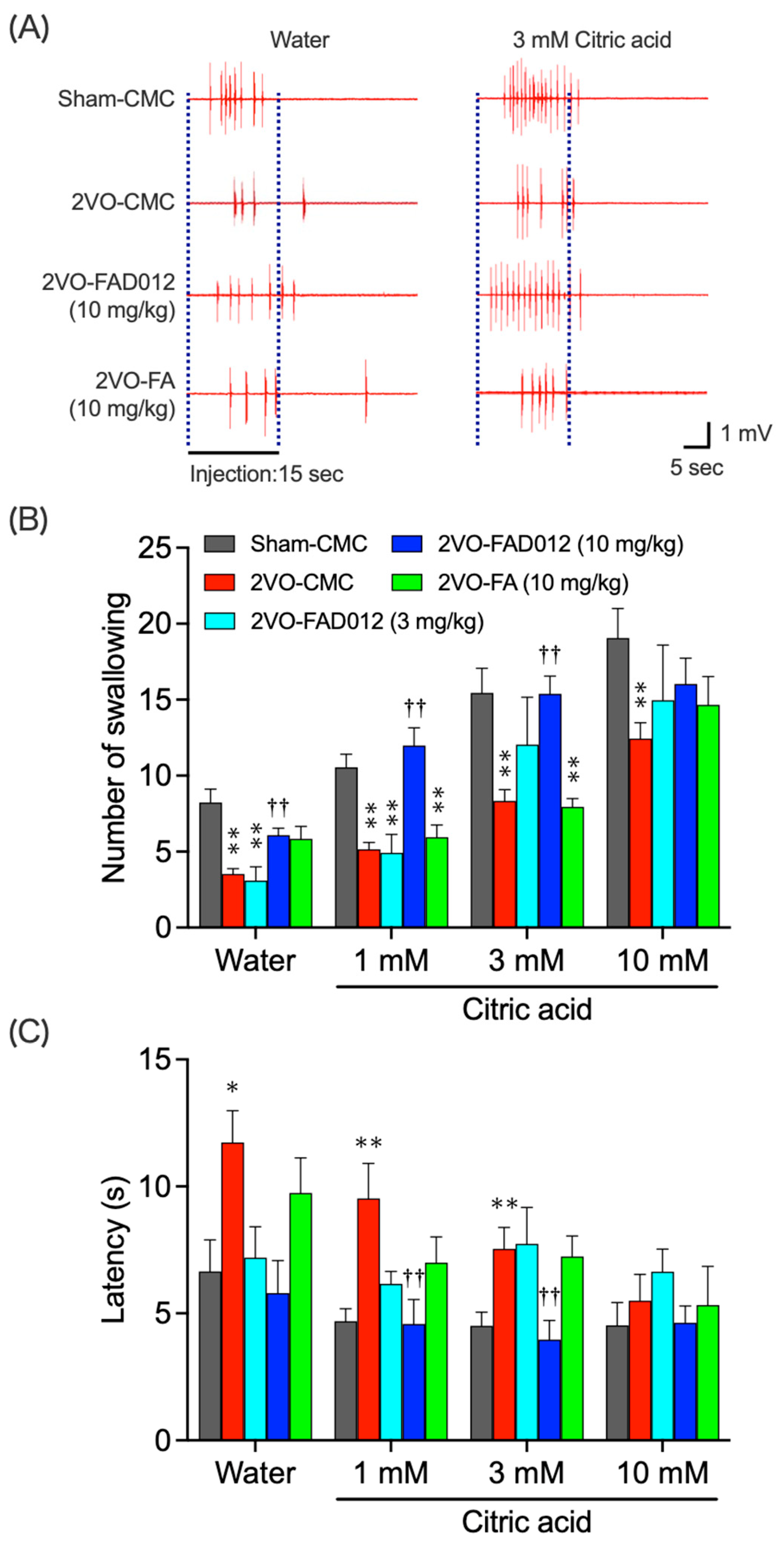
2.3. 2VO-Induced Swallowing Dysfunction
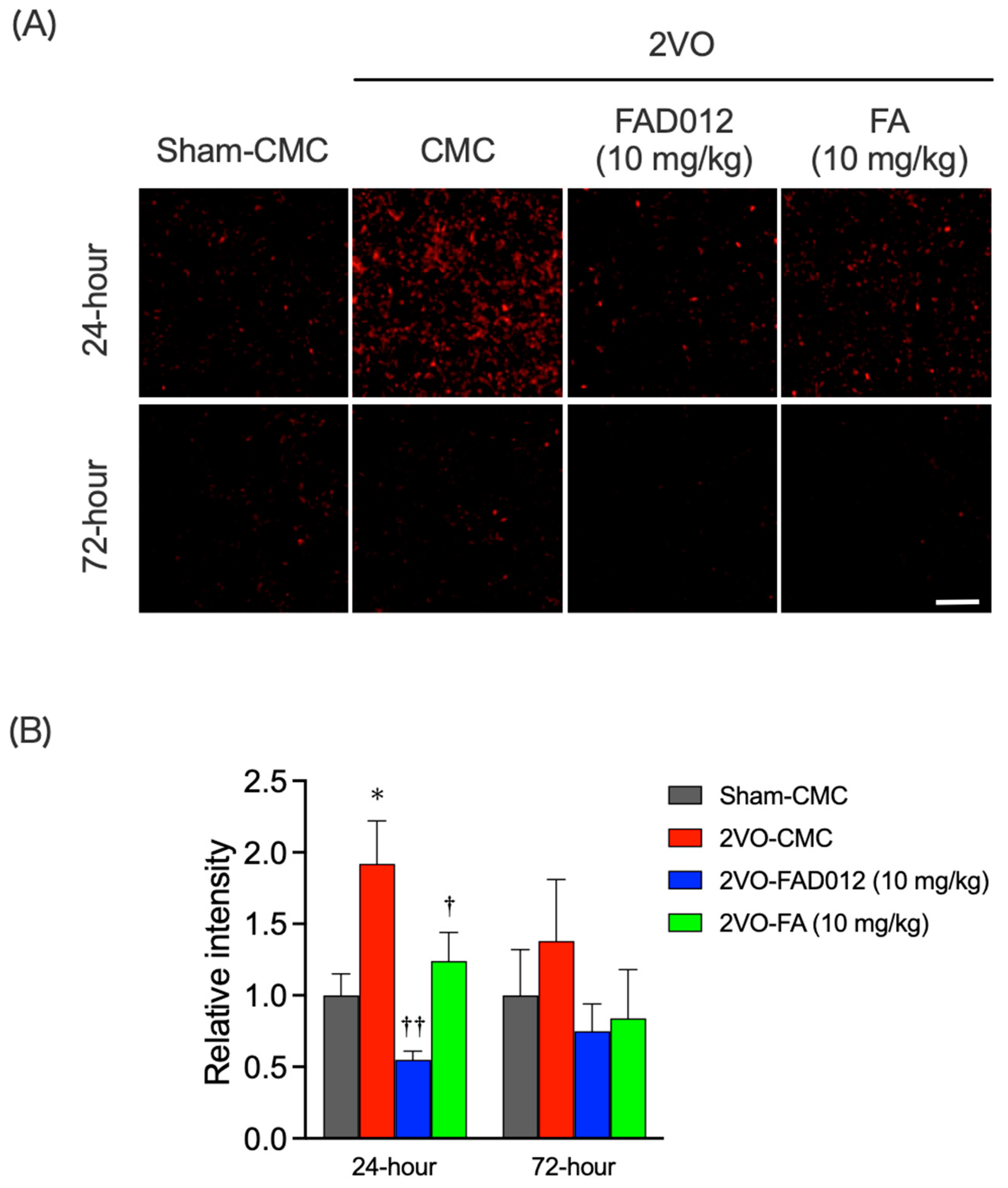
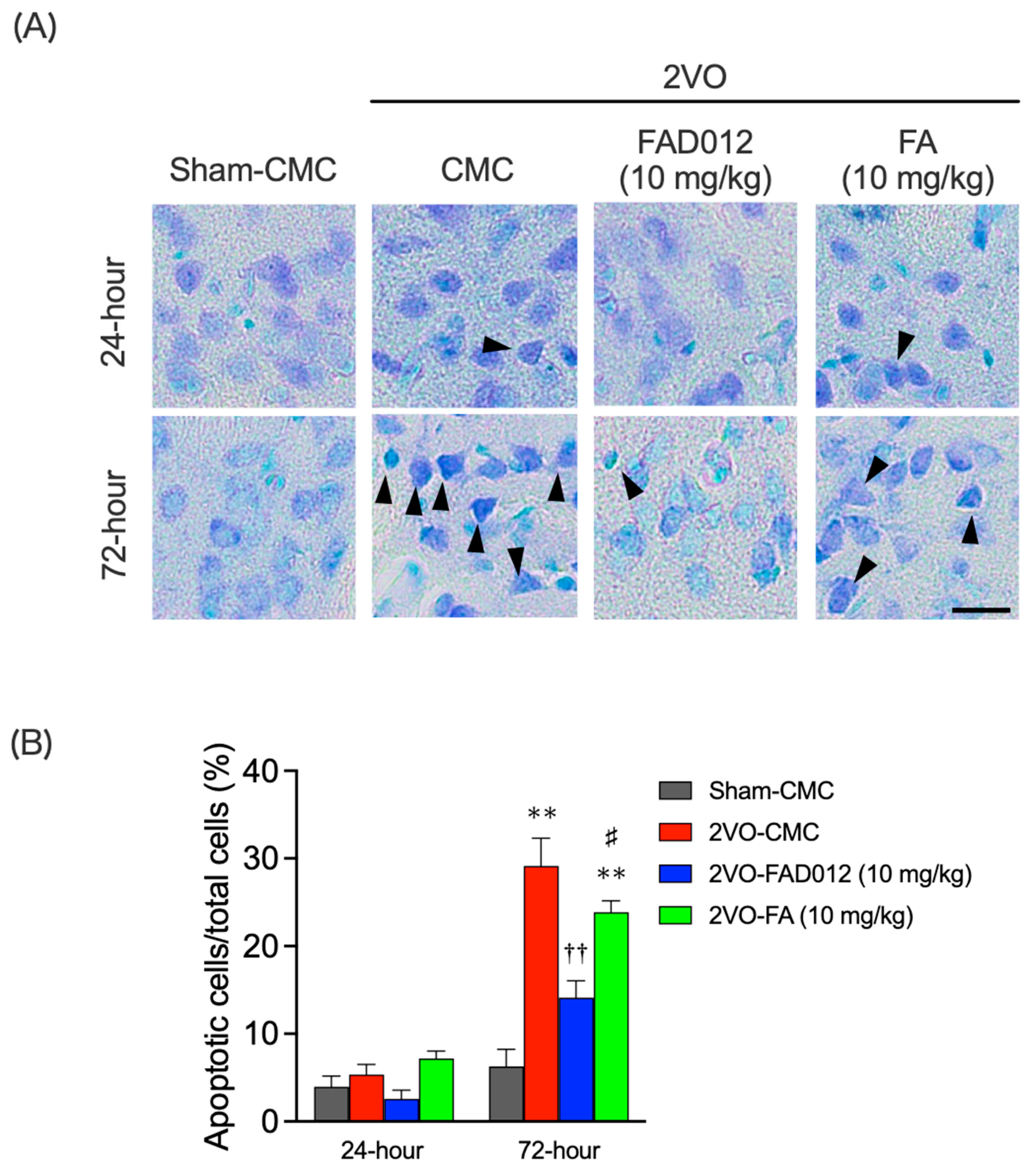
2.4. Oxidative Stress and Apoptotic Cell Death in the Striatum
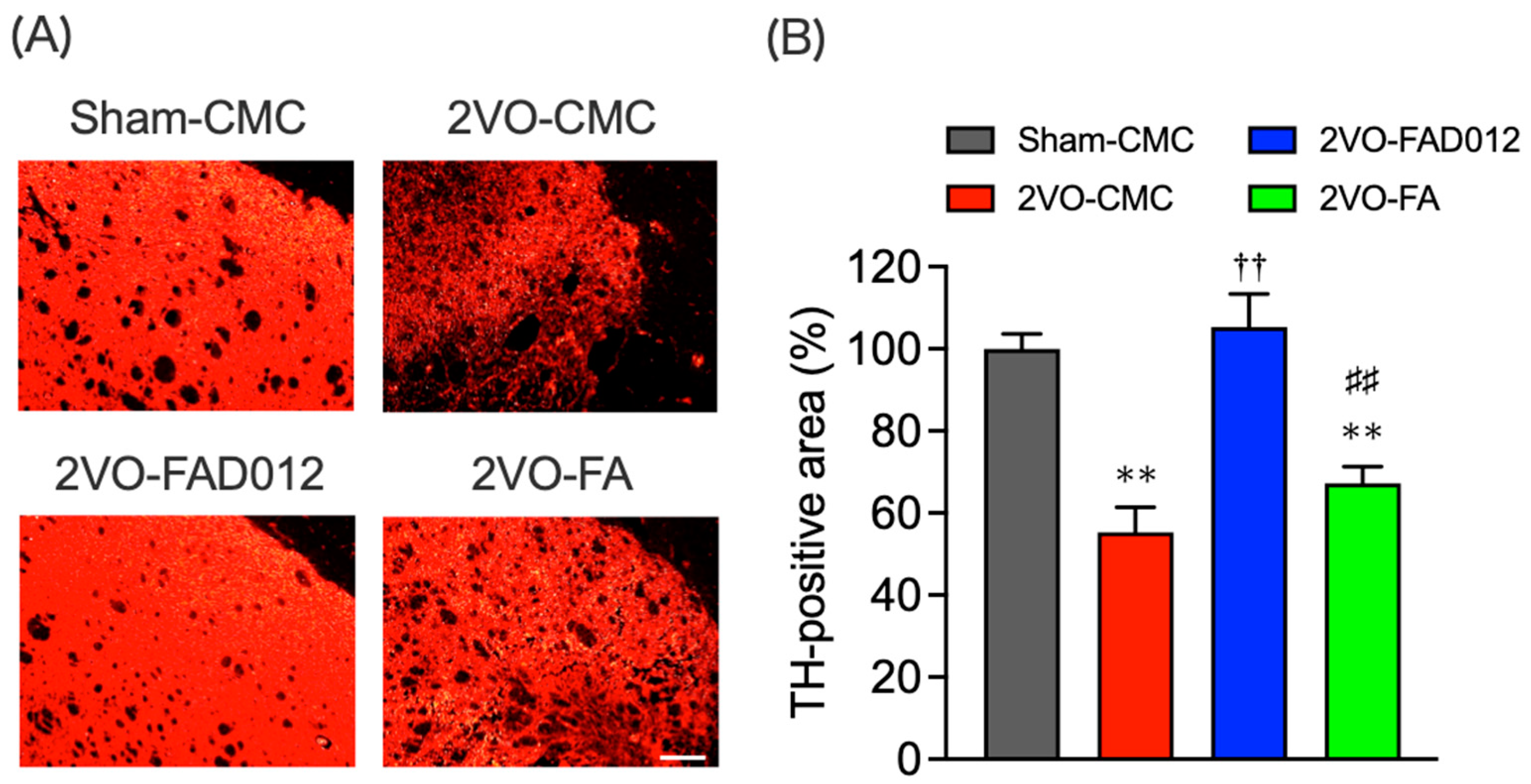
2.5. Tyrosine Hydroxylase (TH) Expression in the Striatum
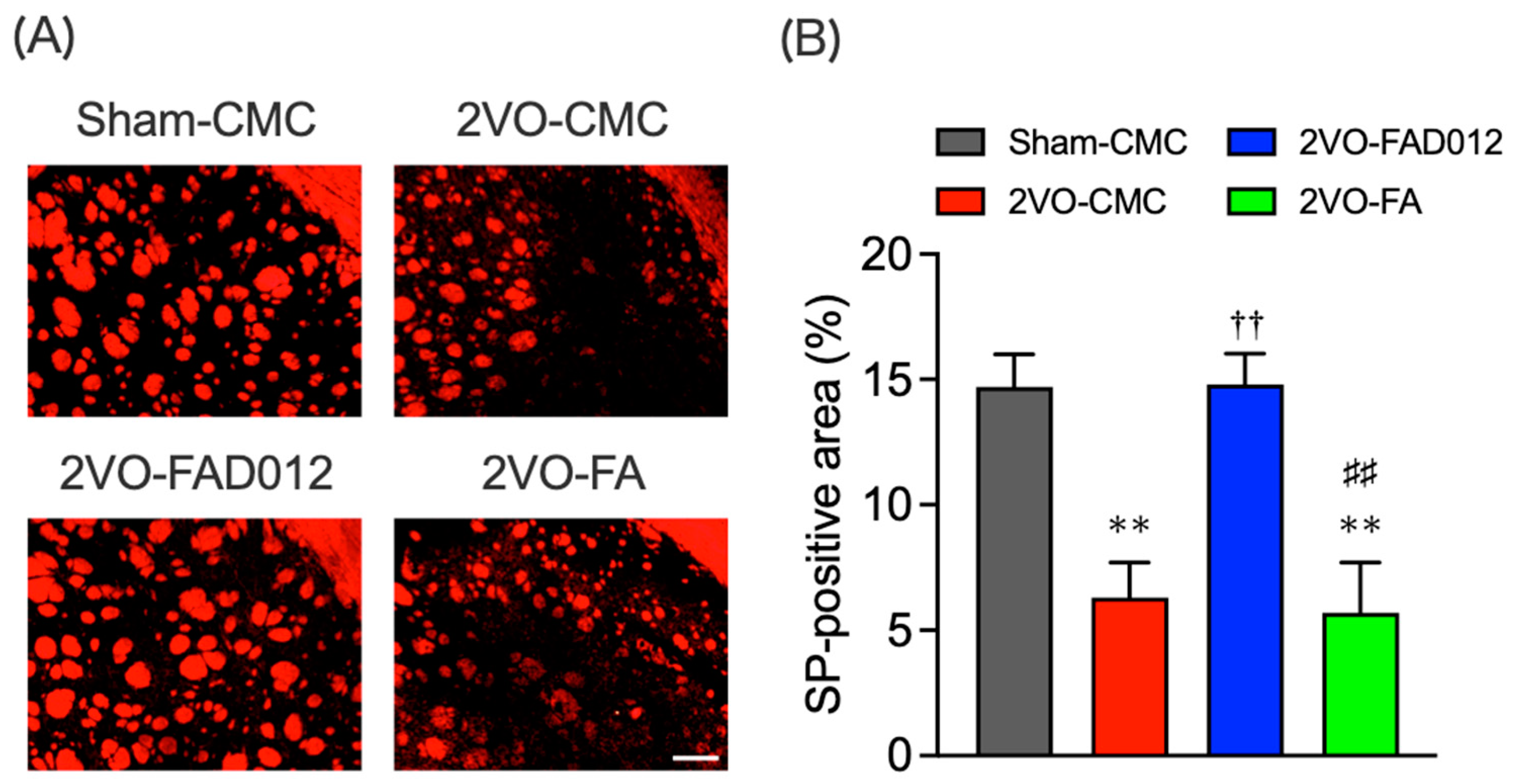
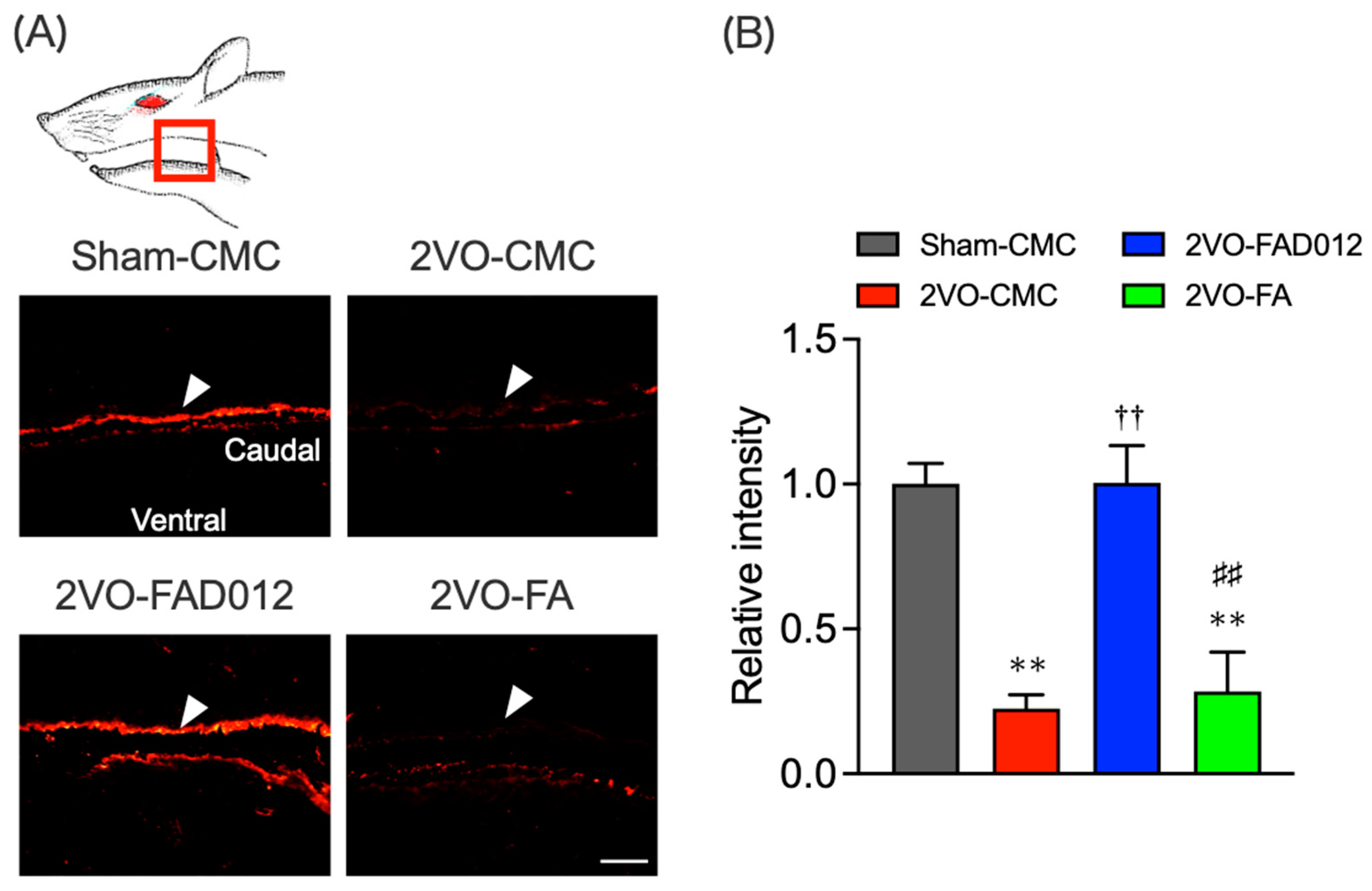
2.6. Substance P (SP) Expression in the Striatum and Laryngopharyngeal Region
3. Discussion
4. Materials and Methods
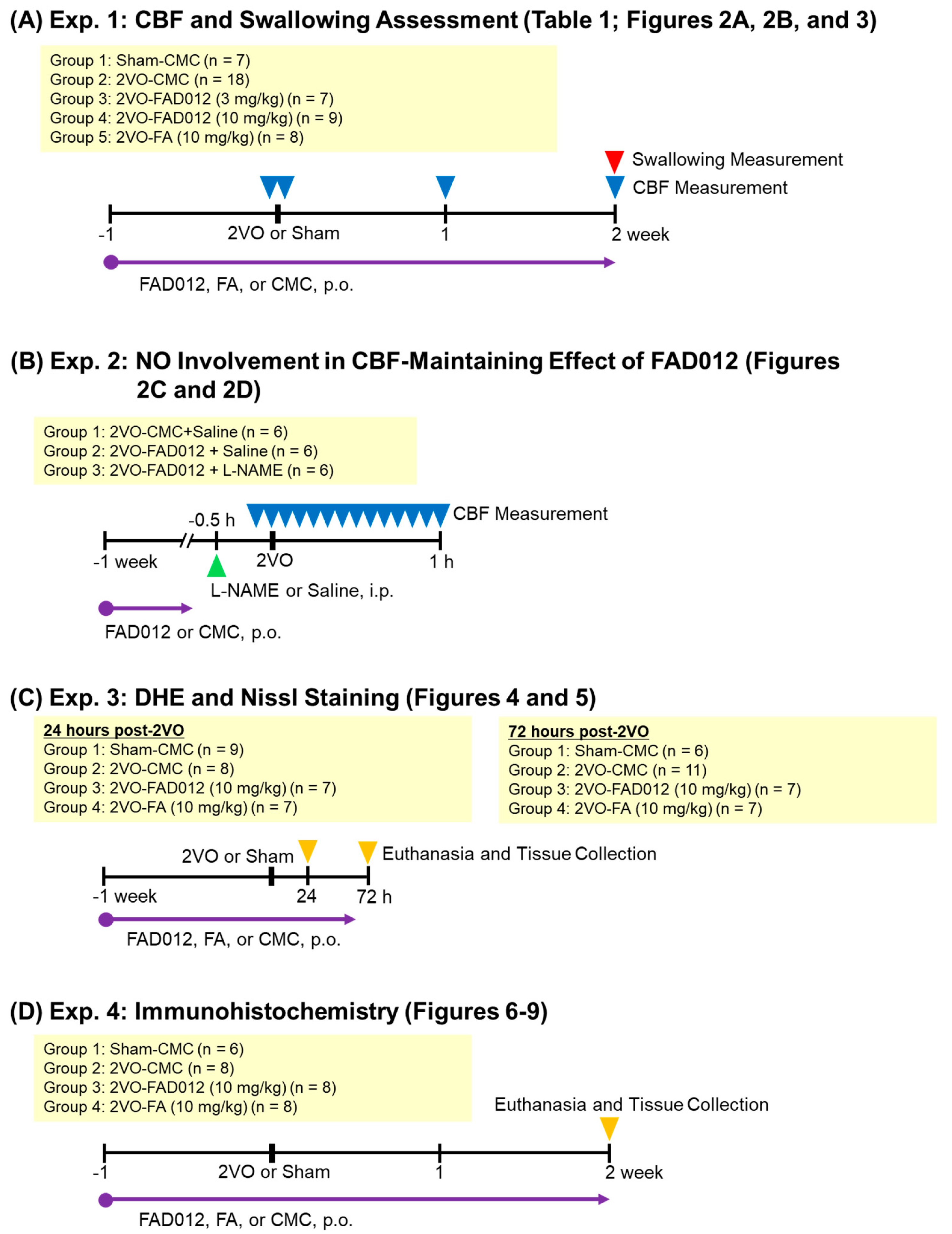
4.1. Experimental Design
4.2. Animals
4.3. 2VO Procedure [13]
4.4. Measurement of CBF [13]
4.5. Measurement of Swallowing [13,14]
4.6. Perfusion Fixation and Coronal Section Preparation [13]
4.7. Evaluation of O2− Production by Dihydroethidium (DHE) Staining [13]
4.8. Nissl Staining [44]
4.9. Immunohistochemistry [13]
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | One-way analysis of variance |
| AUC | Area under the curve |
| BBB | Blood–brain barrier |
| CBF | Cerebral blood flow |
| CMC | Carboxymethyl cellulose |
| CPG | Swallowing central pattern generator |
| DHE | Dihydroethidium |
| EMG | Electromyogram |
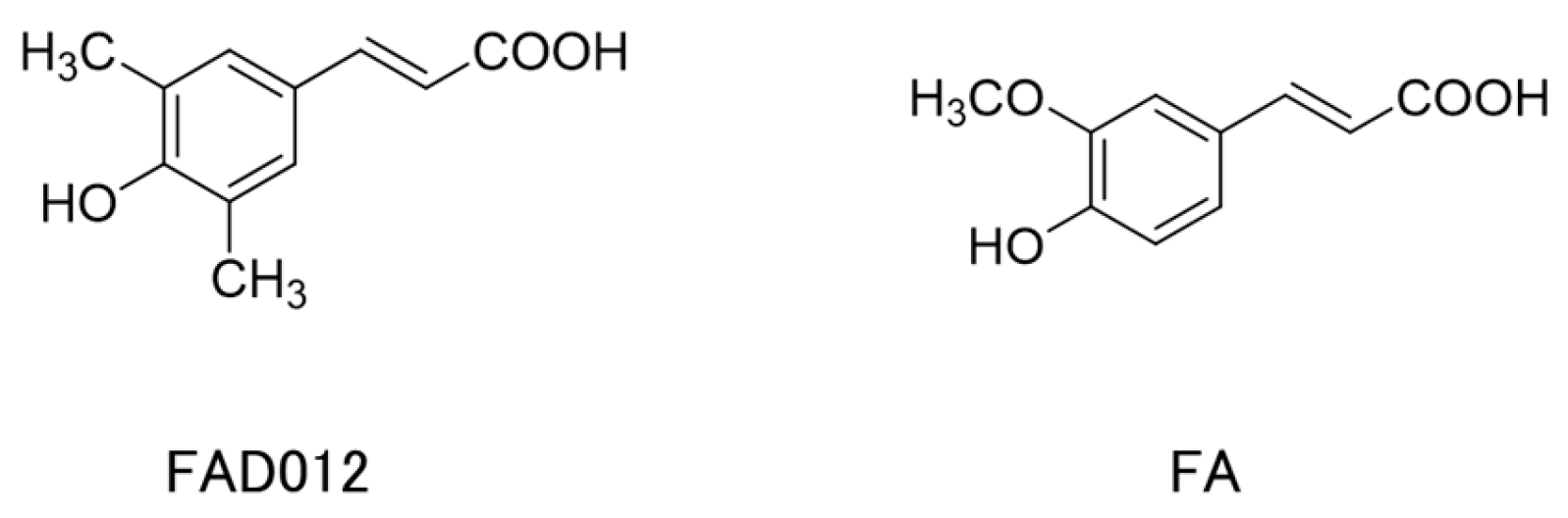
| FA | Ferulic acid; 4-hydroxy-3-methoxycinnamic acid |
| FAD012 | 3,5-Dimethyl-4-hydroxy cinnamic acid |
| HO-1 | Heme oxygenase-1 |
| MCAO | Middle cerebral artery occlusion |
| L-NAME | NG-nitro-L-arginine methyl ester |
| NO | Nitric oxide |
| NOS | NO synthase |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| t-PA | Tissue plasminogen activator |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| ROS | Reactive oxygen species |
| SP | Substance P |
| TH | Tyrosine hydroxylase |
| 2VO | Ligation of bilateral common carotid arteries |
References
- Johnson, C.O.; Nguyen, M.; Roth, G.A.; Nichols, E.; Alam, T.; Abate, D.; Abd-Allah, F.; Abdelalim, A.; Abraha, H.N.; Abu-Rmeileh, N.M.; et al. Global, regional, and national burden of stroke, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.S.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Barone Gibbs, B.; Beaton, A.Z.; Boehme, A.K.; et al. 2024 Heart Disease and Stroke Statistics: A Report of US and Global Data From the American Heart Association. Circulation 2024, 149, e347–e913. [Google Scholar]
- Hankey, G.J. Long-term outcome after ischaemic stroke/transient ischaemic attack. Cerebrovasc. Dis. 2003, 16 (Suppl. S1), 14–19. [Google Scholar] [CrossRef] [PubMed]
- Martino, R.; Foley, N.; Bhogal, S.; Diamant, N.; Speechley, M.; Teasell, R. Dysphagia after stroke: Incidence, diagnosis, and pulmonary complications. Stroke 2005, 36, 2756–2763. [Google Scholar] [CrossRef]
- Cohen, D.L.; Roffe, C.; Beavan, J.; Blackett, B.; Fairfield, C.A.; Hamdy, S.; Havard, D.; McFarlane, M.; McLauglin, C.; Randall, M.; et al. Post-stroke dysphagia: A review and design considerations for future trials. Int. J. Stroke 2016, 11, 399–411. [Google Scholar] [CrossRef]
- Banda, K.J.; Chu, H.; Kang, X.L.; Liu, D.; Pien, L.C.; Jen, H.J.; Hsiao, S.S.; Chou, K.R. Prevalence of dysphagia and risk of pneumonia and mortality in acute stroke patients: A meta-analysis. BMC Geriatr. 2022, 22, 420. [Google Scholar] [CrossRef] [PubMed]
- Balcerak, P.; Corbiere, S.; Zubal, R.; Kägi, G. Post-stroke Dysphagia: Prognosis and Treatment–A Systematic Review of, R.C.T on Interventional Treatments for Dysphagia Following Subacute Stroke. Front. Neurol. 2022, 13, 823189. [Google Scholar] [CrossRef]
- Powers, W.J.; Rabinstein, A.A.; Ackerson, T.; Adeoye, O.M.; Bambakidis, N.C.; Becker, K.; Biller, J.; Brown, M.; Demaerschalk, B.M.; Hoh, B.; et al. Guidelines for the Early Management of Patients with Acute Ischemic Stroke: 2019 Update to the 2018 Guidelines for the Early Management of Acute Ischemic Stroke: A Guideline for Healthcare Professionals from the American Heart Association/American Stroke Association. Stroke 2019, 50, e344–e418. [Google Scholar]
- Miyamoto, S.; Ogasawara, K.; Kuroda, S.; Itabashi, R.; Toyoda, K.; Itoh, Y.; Iguchi, Y.; Shiokawa, Y.; Takagi, Y.; Ohtsuki, T.; et al. Japan Stroke Society Guideline 2021 for the Treatment of Stroke. Int. J. Stroke 2022, 17, 1039–1049. [Google Scholar] [CrossRef]
- Li, D.; Rui, Y.X.; Guo, S.D.; Luan, F.; Liu, R.; Zeng, N. Ferulic acid: A review of its pharmacology, pharmacokinetics and derivatives. Life Sci. 2021, 284, 119921. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Su, S.Y.; Tang, N.Y.; Ho, T.Y.; Chiang, S.Y.; Hsieh, C.L. Ferulic acid provides neuroprotection against oxidative stress-related apoptosis after cerebral ischemia/reperfusion injury by inhibiting ICAM-1 mRNA expression in rats. Brain Res. 2008, 1209, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Barone, E.; Calabrese, V.; Mancuso, C. Ferulic acid and its therapeutic potential as a hormetin for age-related diseases. Biogerontology 2009, 10, 97–108. [Google Scholar] [CrossRef]
- Asano, T.; Matsuzaki, H.; Iwata, N.; Xuan, M.; Kamiuchi, S.; Hibino, Y.; Sakamoto, T.; Okazaki, M. Protective effects of ferulic acid against chronic cerebral hypoperfusion-induced swallowing dysfunction in rats. Int. J. Mol. Sci. 2017, 18, 550. [Google Scholar] [CrossRef]
- Kajii, Y.; Shingai, T.; Kitagawa, J.I.; Takahashi, Y.; Taguchi, Y.; Noda, T.; Yamada, Y. Sour taste stimulation facilitates reflex swallowing from the pharynx and larynx in the rat. Physiol. Behav. 2002, 77, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Ertekin, C.; Aydogdu, I. Neurophysiology of swallowing. Clin. Neurophysiol. Off. J. Int. Fed. Clin. Neurophysiol. 2003, 114, 2226–2244. [Google Scholar]
- Cruz-Sanchez, F.F.; Cardozo, A.; Castejon, C.; Tolosa, E.; Rossi, M.L. Aging and the nigro-striatal pathway. J. Neural Transm. Suppl. 1997, 51, 9–25. [Google Scholar]
- Jia, Y.X.; Sekizawa, K.; Ohrui, T.; Nakayama, K.; Sasaki, H. Dopamine D1 receptor antagonist inhibits swallowing reflex in guinea pigs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 274, R76–R80. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jeon, J.; Lee, Y.; Kim, S.M.; Cheon, M.; Kim, J.Y. Striatal Dopaminergic Loss and Dysphagia in Parkinson Disease. Clin. Nucl. Med. 2023, 48, 143–149. [Google Scholar] [CrossRef]
- Jin, Y.; Sekizawa, K.; Fukushima, T.; Morikawa, M.; Nakazawa, H.; Sasaki, H. Capsaicin desensitization inhibits swallowing reflex in guinea pigs. Am. J. Respir. Crit. Care Med. 1994, 149, 261–263. [Google Scholar] [CrossRef]
- Horner, J.; Massey, E.W. Silent aspiration following stroke. Neurology 1988, 38, 317–319. [Google Scholar] [CrossRef]
- Takizawa, C.; Gemmell, E.; Kenworthy, J.; Speyer, R. A Systematic Review of the Prevalence of Oropharyngeal Dysphagia in Stroke, Parkinson’s Disease, Alzheimer’s Disease, Head Injury, and Pneumonia. Dysphagia 2016, 31, 434–441. [Google Scholar] [PubMed]
- Suttrup, I.; Warnecke, T. Dysphagia in Parkinson’s Disease. Dysphagia 2016, 31, 24–32. [Google Scholar]
- Asano, T.; Xuan, M.; Iwata, N.; Takayama, J.; Hayashi, K.; Kato, Y.; Aoyama, T.; Sugo, H.; Matsuzaki, H.; Yuan, B.; et al. Involvement of the Restoration of Cerebral Blood Flow and Maintenance of eNOS Expression in the Prophylactic Protective Effect of the Novel Ferulic Acid Derivative FAD012 against Ischemia/Reperfusion Injuries in Rats. Int. J. Mol. Sci. 2023, 24, 9663. [Google Scholar] [CrossRef]
- Teramae, H.; Xuan, M.; Yamashita, T.; Takayama, J.; Okazaki, M.; Sakamoto, T. Theoretical study on antioxidant properties of ferulic acid. J. Comput. Chem. Jpn 2018, 17, 150–152. [Google Scholar]
- Osawa, A.; Maeshima, S.; Tanahashi, N. Efficacy of cilostazol in preventing aspiration pneumonia in acute cerebral infarction. J. Stroke Cerebrovasc. Dis. 2013, 22, 857–861. [Google Scholar] [PubMed]
- Ikeda, N.; Inoue, Y.; Ogata, Y.; Murata, I.; Meiyan, X.; Takayama, J.; Sakamoto, T.; Okazaki, M.; Kanamoto, I. Improvement of the Solubility and Evaluation of the Physical Properties of an Inclusion Complex Formed by a New Ferulic Acid Derivative and γ-Cyclodextrin. ACS Omega 2020, 5, 12073–12080. [Google Scholar] [CrossRef]
- Zhang, N.; Miyamoto, N.; Tanaka, R.; Mochizuki, H.; Hattori, N.; Urabe, T. Activation of tyrosine hydroxylase prevents pneumonia in a rat chronic cerebral hypoperfusion model. Neuroscience 2009, 158, 665–672. [Google Scholar]
- Sivam, S.P.; Cox, J. Postnatal administration of D1 dopamine agonist reverses neonatal dopaminergic lesion-induced changes in striatal enkephalin and substance P systems. Brain Res. 2006, 1073–1074, 159–163. [Google Scholar]
- Ren, Z.; Zhang, R.; Li, Y.; Li, Y.; Yang, Z.; Yang, H. Ferulic acid exerts neuroprotective effects against cerebral ischemia/reperfusion-induced injury via antioxidant and anti-apoptotic mechanisms in vitro and in vivo. Int. J. Mol. Med. 2017, 40, 1444–1456. [Google Scholar] [CrossRef]
- Mugundhan, V.; Arthanari, A.; Parthasarathy, P.R. Protective Effect of Ferulic Acid on Acetylcholinesterase and Amyloid Beta Peptide Plaque Formation in Alzheimer’s Disease: An In Vitro Study. Cureus 2024, 16, e54103. [Google Scholar]
- Karademir, Y.; Mackie, A.; Tuohy, K.; Dye, L. Effects of Ferulic Acid on Cognitive Function: A Systematic Review. Mol. Nutr. Food Res. 2024, 68, 2300526. [Google Scholar]
- Wang, F.; Peng, Q.; Liu, J.; Alolga, R.N.; Zhou, W. A novel ferulic acid derivative attenuates myocardial cell hypoxia reoxygenation injury through a succinate dehydrogenase dependent antioxidant mechanism. Eur. J. Pharmacol. 2019, 856, 172417. [Google Scholar] [CrossRef]
- Ronchetti, D.; Borghi, V.; Gaitan, G.; Herrero, J.F.; Impagnatiello, F. NCX 2057, a novel NO-releasing derivative of ferulic acid, suppresses inflammatory and nociceptive responses in in vitro and in vivo models. Br. J. Pharmacol. 2009, 158, 569–579. [Google Scholar] [PubMed]
- Ikeda, J.I.; Kojima, N.; Saeki, K.; Ishihara, M.; Takayama, M. Perindopril increases the swallowing reflex by inhibiting substance P degradation and tyrosine hydroxylase activation in a rat model of dysphagia. Eur. J. Pharmacol. 2015, 746, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Koh, P.O. Ferulic acid modulates nitric oxide synthase expression in focal cerebral ischemia. Lab. Anim. Res. 2012, 28, 273–278. [Google Scholar] [PubMed]
- Ji, H.J.; Hu, J.F.; Wang, Y.H.; Chen, X.Y.; Zhou, R.; Chen, N.H. Osthole improves chronic cerebral hypoperfusion induced cognitive deficits and neuronal damage in hippocampus. Eur. J. Pharmacol. 2010, 636, 96–101. [Google Scholar]
- Niizuma, K.; Yoshioka, H.; Chen, H.; Kim, G.S.; Jung, J.E.; Katsu, M.; Okami, N.; Chan, P.H. Mitochondrial and apoptotic neuronal death signaling pathways in cerebral ischemia. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2010, 1802, 92–99. [Google Scholar]
- Zhai, Y.; Wang, T.; Fu, Y.; Yu, T.; Ding, Y.; Nie, H. Ferulic Acid: A Review of Pharmacology, Toxicology, and Therapeutic Effects on Pulmonary Diseases. Int. J. Mol. Sci. 2023, 24, 8011. [Google Scholar] [CrossRef]
- Kobayashi, H.; Nakagawa, T.; Sekizawa, K.; Arai, H.; Sasaki, H. Levodopa and swallowing reflex. Lancet 1996, 348, 1320–1321. [Google Scholar]
- Arai, T.; Sekizawa, K.; Yoshimi, N.; Toshima, S.; Fujiwara, H. Cabergoline and silent aspiration in elderly patients with stroke. J. Am. Geriatr. Soc. 2003, 51, 1815–1816. [Google Scholar] [CrossRef]
- Hirano, M.; Isono, C.; Sakamoto, H.; Ueno, S.; Kusunoki, S.; Nakamura, Y. Rotigotine Transdermal Patch Improves Swallowing in Dysphagic Patients with Parkinson’s Disease. Dysphagia 2015, 30, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Zhang, N.; Liu, M.; Tanaka, R.; Mizuno, Y.; Urabe, T. Cilostazol protects against brain white matter damage and cognitive impairment in a rat model of chronic cerebral hypoperfusion. Stroke 2006, 37, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Komine-Kobayashi, M.; Tanaka, R.; Liu, M.; Mizuno, Y.; Urabe, T. Edaravone reduces early accumulation of oxidative products and sequential inflammatory responses after transient focal ischemia in mice brain. Stroke 2005, 36, 2220–2225. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotactic Coordinates; Academic Press Inc.: San Diego, CA, USA, 1986. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Relative Body Weights (%) | Survival Rates (%) |
|---|---|---|
| Sham-CMC | 112.6 ± 3.2 | 100 (7/7) |
| 2VO-CMC | 99.2 ± 2.2 * | 66.7 (12/18) |
| 2VO-FAD012 (3 mg/kg) | 102.3 ± 1.3 * | 85.7 (6/7) |
| 2VO-FAD012 (10 mg/kg) | 98.4 ± 1.9 * | 77.8 (7/9) |
| 2VO-FA (10 mg/kg) | 98.8 ± 1.6 * | 87.5 (7/8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asano, T.; Matsuzaki, H.; Xuan, M.; Yuan, B.; Takayama, J.; Sakamoto, T.; Okazaki, M. Chronic Administration with FAD012 (3,5-Dimethyl-4-hydroxycinnamic Acid) Maintains Cerebral Blood Flow and Ameliorates Swallowing Dysfunction After Chronic Cerebral Hypoperfusion in Rats. Int. J. Mol. Sci. 2025, 26, 3277. https://doi.org/10.3390/ijms26073277
Asano T, Matsuzaki H, Xuan M, Yuan B, Takayama J, Sakamoto T, Okazaki M. Chronic Administration with FAD012 (3,5-Dimethyl-4-hydroxycinnamic Acid) Maintains Cerebral Blood Flow and Ameliorates Swallowing Dysfunction After Chronic Cerebral Hypoperfusion in Rats. International Journal of Molecular Sciences. 2025; 26(7):3277. https://doi.org/10.3390/ijms26073277
Chicago/Turabian StyleAsano, Takashi, Hirokazu Matsuzaki, Meiyan Xuan, Bo Yuan, Jun Takayama, Takeshi Sakamoto, and Mari Okazaki. 2025. "Chronic Administration with FAD012 (3,5-Dimethyl-4-hydroxycinnamic Acid) Maintains Cerebral Blood Flow and Ameliorates Swallowing Dysfunction After Chronic Cerebral Hypoperfusion in Rats" International Journal of Molecular Sciences 26, no. 7: 3277. https://doi.org/10.3390/ijms26073277
APA StyleAsano, T., Matsuzaki, H., Xuan, M., Yuan, B., Takayama, J., Sakamoto, T., & Okazaki, M. (2025). Chronic Administration with FAD012 (3,5-Dimethyl-4-hydroxycinnamic Acid) Maintains Cerebral Blood Flow and Ameliorates Swallowing Dysfunction After Chronic Cerebral Hypoperfusion in Rats. International Journal of Molecular Sciences, 26(7), 3277. https://doi.org/10.3390/ijms26073277