
Unveiling the Mystery of the Stimulatory Effects of Arecoline: Its Relevance to the Regulation of Neurotransmitters and the Microecosystem in Multi-Ecological Intestinal Sites



Abstract
1. Introduction
2. Results
2.1. Effects of the Arecoline Solution on Mental State and Spontaneous Activity in Mice
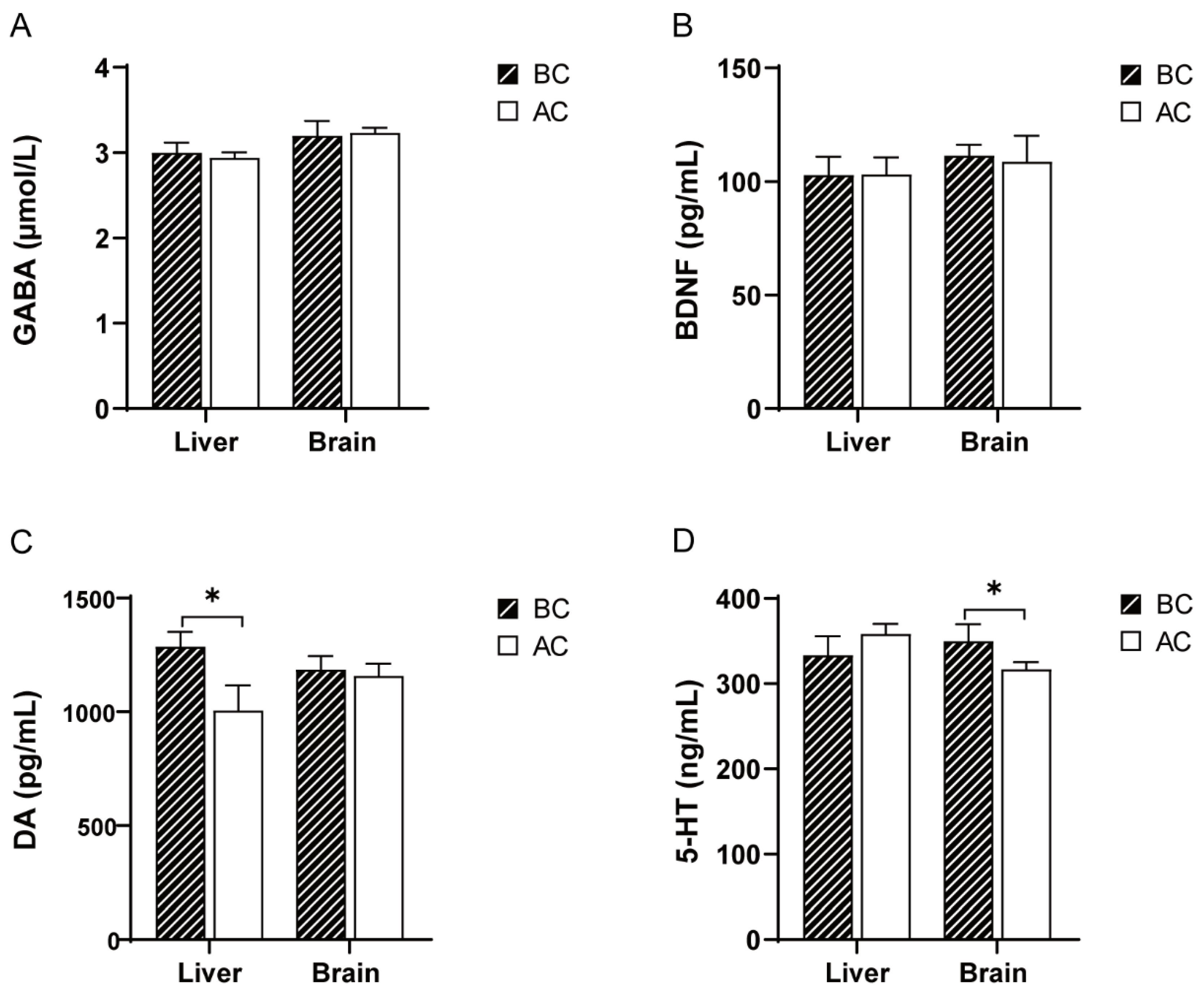
2.2. Effects of the Arecoline Solution on the GABA, DA, 5-HT, and BDNF Levels in the Liver and Brain Tissues of Mice
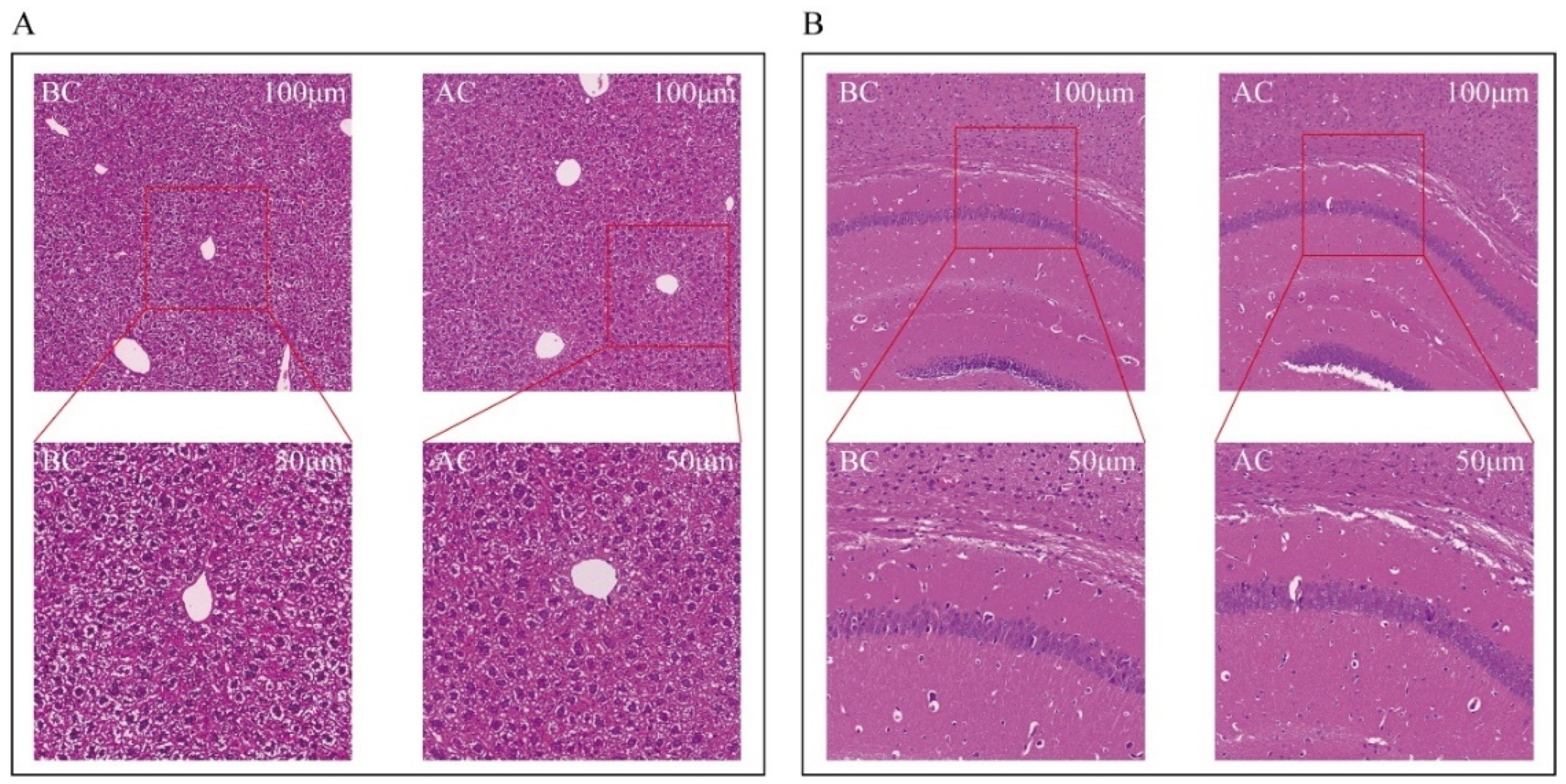
2.3. Effects of the Arecoline Solution on the Morphology of the Liver and Brain Tissues in Mice
2.4. Effects of the Arecoline Solution on the Intestinal Microecosystem of Mice
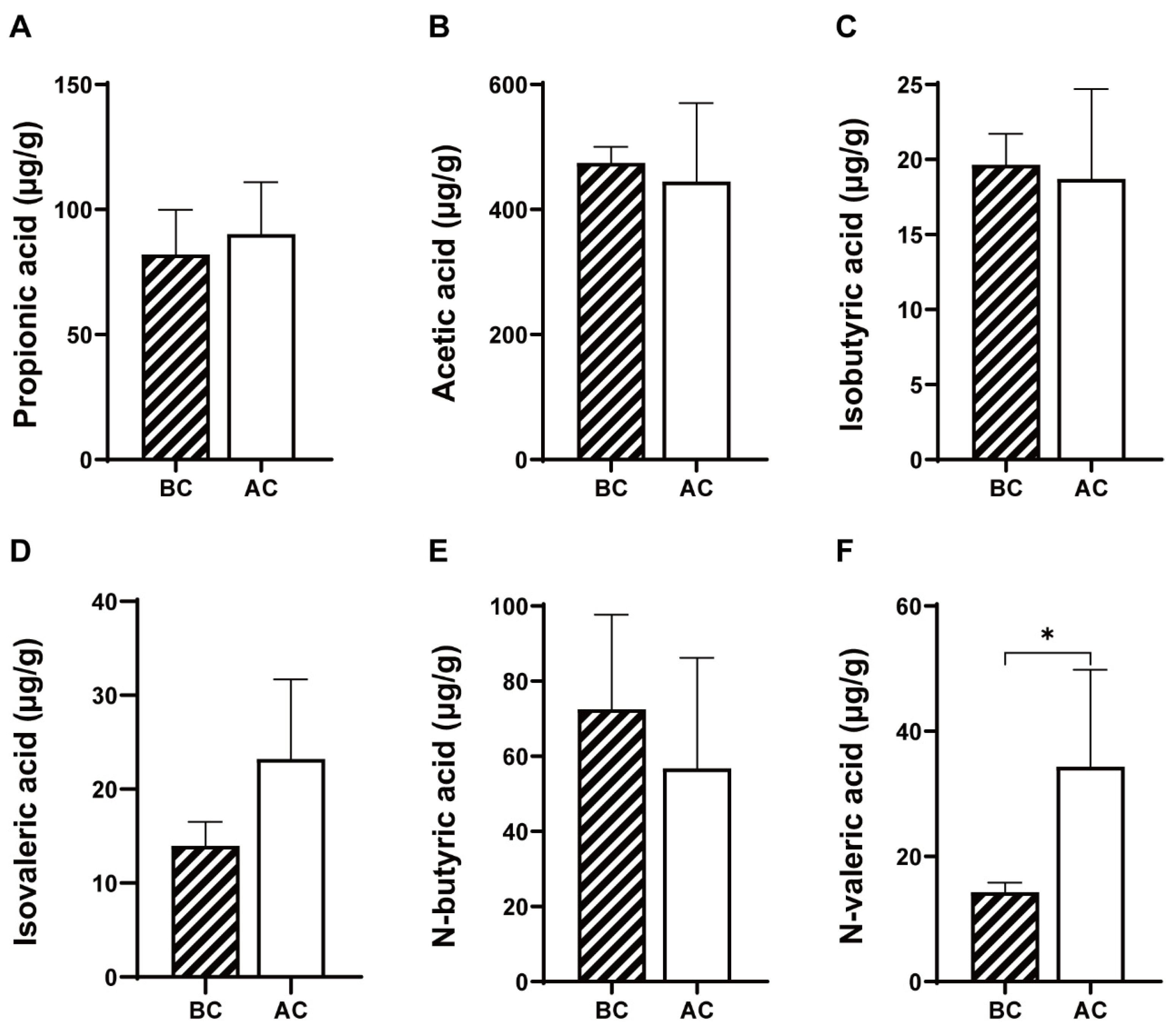
2.4.1. Effects of the Arecoline Solution on SCFAs in the Intestinal Contents of Mice
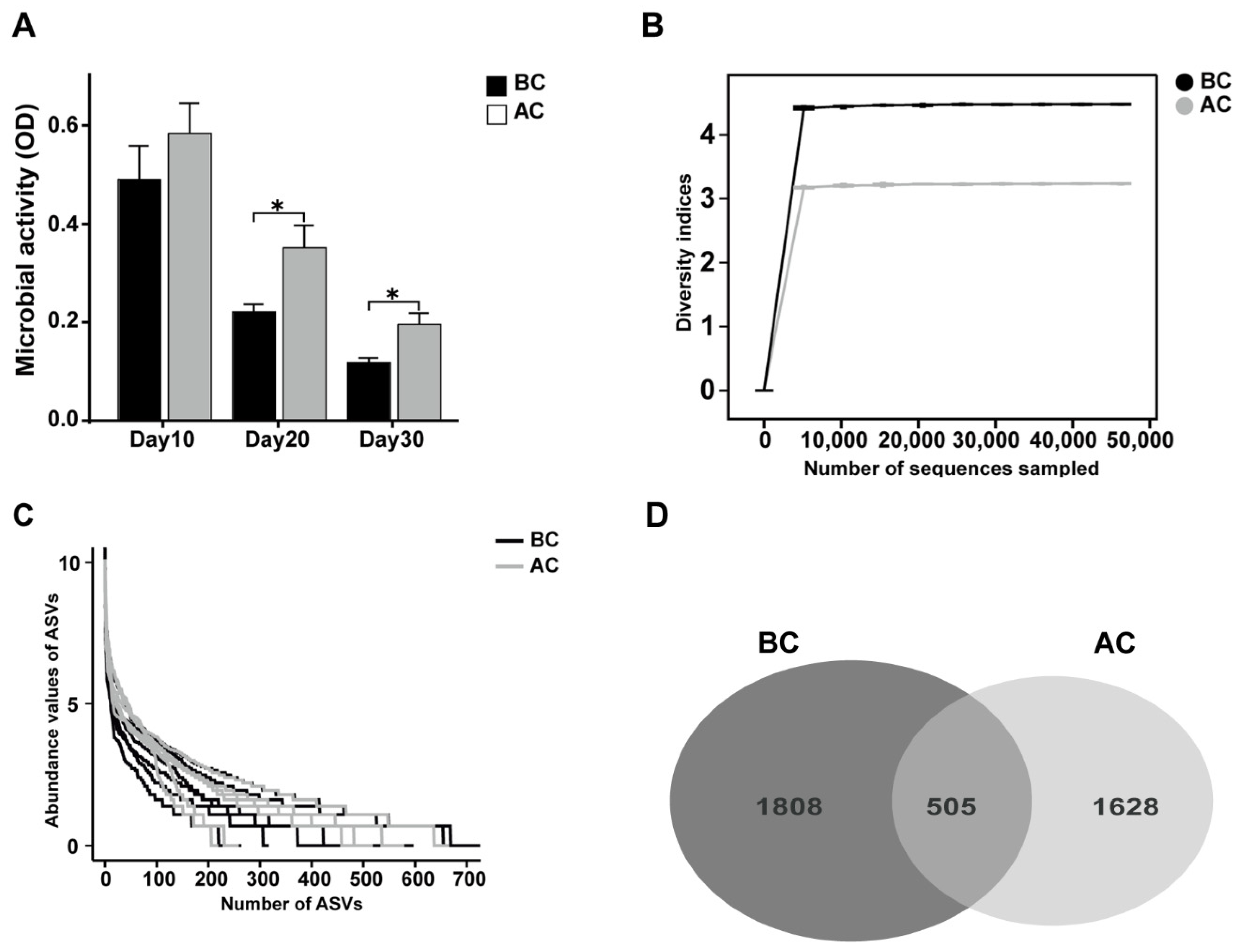
2.4.2. Effects of the Arecoline Solution on Fecal Microbial Activity and ASVs in the Intestinal Mucosal Microbiota in Mice
2.4.3. Effects of the Arecoline Solution on the Species Diversity of the Intestinal Mucosal Microbiota in Mice
2.4.4. Effects of the Arecoline Solution on the Composition and Relative Abundance of the Intestinal Mucosal Microbiota in Mice
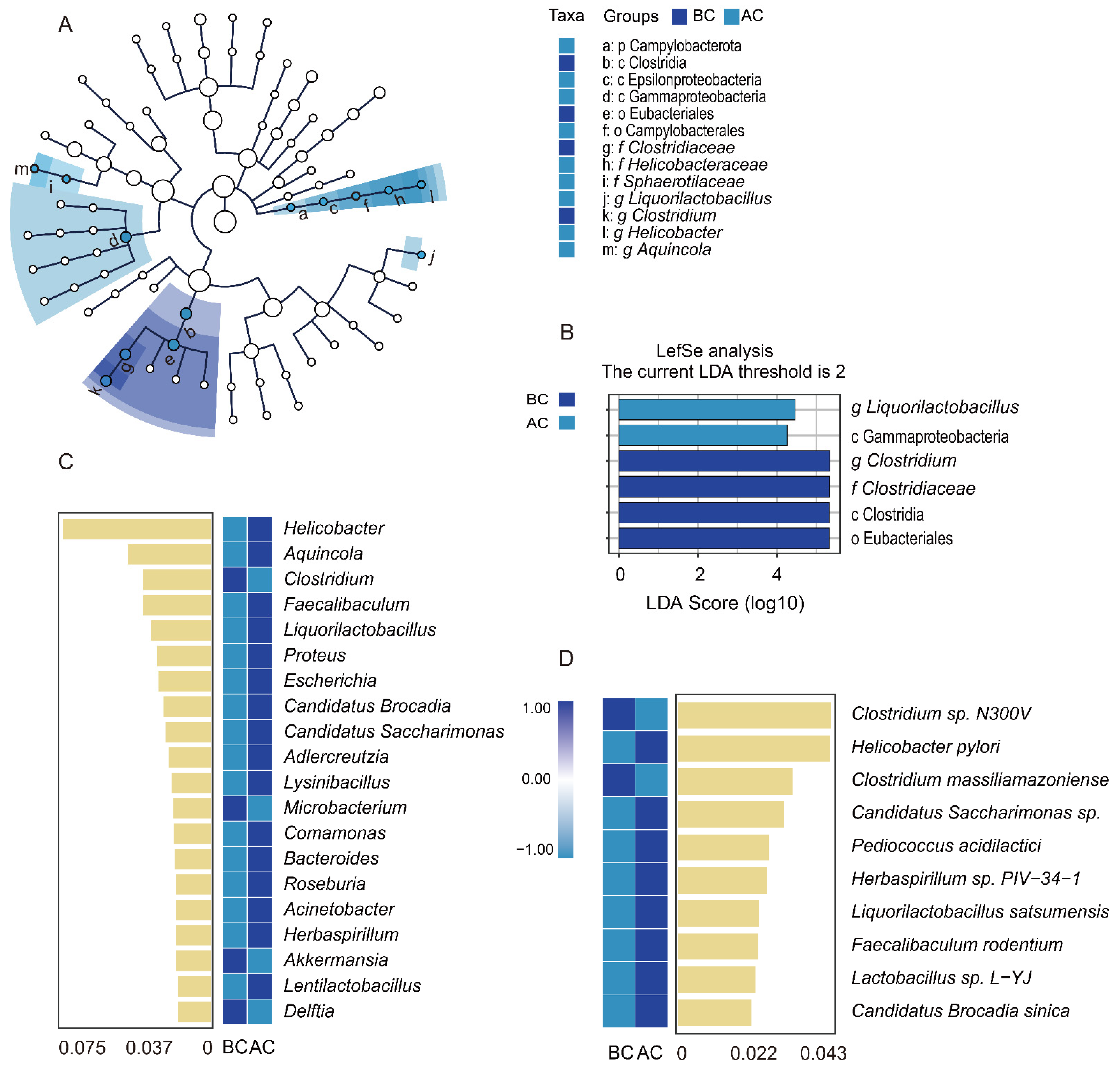
2.4.5. Effects of the Arecoline Solution on Marker Species of the Intestinal Mucosal Microbiota in Mice
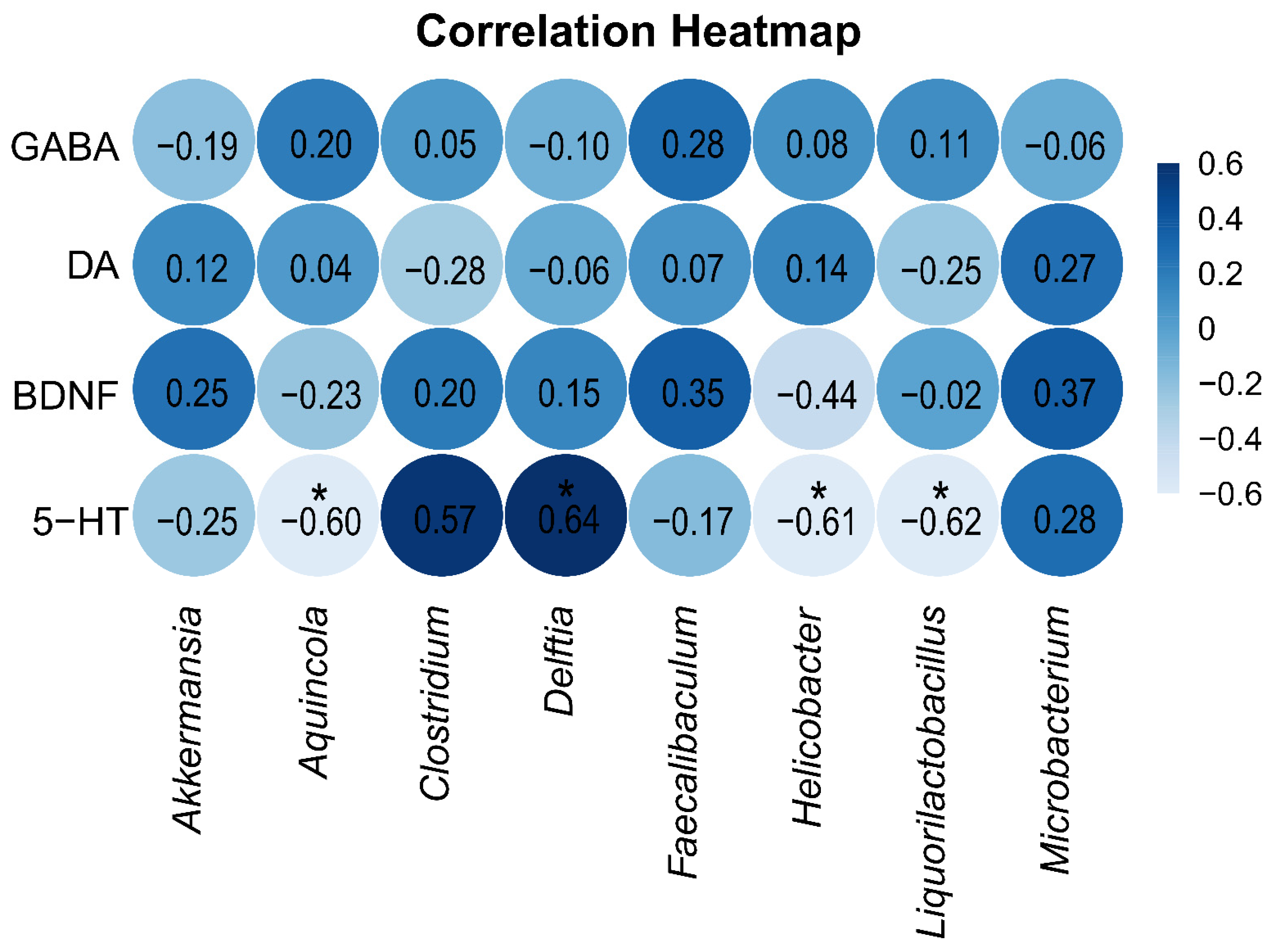
2.5. Correlation Analysis Between GABA, DA, 5-HT, and BDNF in Brain Tissue and the Intestinal Mucosal Microbiota
3. Discussion
3.1. Arecoline Alters Mouse Behavior and Exerts Significant Effects on DA and 5-HT
3.2. Arecoline Enhances Intestinal Microbial Activity and Affects the SCFA Levels in the Intestine
3.3. Arecoline Alters the Structure and Diversity of the Intestinal Mucosal Microbiota in Mice
3.4. Correlation Between the 5-HT Levels in Brain Tissue and Marker Bacterial Genera of the Intestinal Mucosal Microbiota
4. Material and Methods
4.1. Preparation of the Experimental Animals
4.2. Information on Animal Feed
4.3. Pharmaceuticals and Kits
4.4. Animal Grouping and Drug Administration
4.5. Open-Field Test
4.6. Sample Collection
4.7. Detection of GABA, DA, 5-HT, and BDNF in Liver and Brain Tissues
4.8. Histological Analysis of Liver and Brain Tissues
4.9. Fecal Microbial Activity Assay
4.10. Detection of SCFAs in the Intestinal Content
4.11. DNA Extraction, 16S rRNA Gene Amplicon Sequencing, and Sequence Analysis
4.12. Bioinformatics Analysis of High-Throughput Sequencing Data
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chatterjee, N.; Gupte, H.A. Areca nut use among adolescents: How do we prevent and control this problem? J. Glob. Health 2023, 13, 03022. [Google Scholar] [CrossRef]
- Benegal, V.; Rajkumar, R.P.; Muralidharan, K. Does areca nut use lead to dependence? Drug Alcohol. Depend. 2008, 9, 114–121. [Google Scholar] [CrossRef]
- Oliveira, N.G.; Ramos, D.L.; Dinis-Oliveira, R.J. Genetic toxicology and toxicokinetics of arecoline and related areca nut compounds: An updated review. Arch. Toxicol. 2021, 95, 375–393. [Google Scholar] [CrossRef]
- Ansari, A.; Mahmood, T.; Bagga, P.; Shamim, A.; Ahmad, S. Areca catechu: A phytopharmacological legwork. Food Front. 2021, 2, 163–183. [Google Scholar]
- Dangore-Khasbage, S.; Bhowate, R.R.; Khubchandani, M. Chemical Composition of Areca Nut and Its Adverse Effects on Human Health. Cureus 2023, 15, e43739. [Google Scholar] [CrossRef]
- Pfeiffer, C.C.; Beck, R.A.; Goldstein, L. The modification of central nervous system (CNS) function by autonomic drugs. Amine shift responses differentiate be-tween CNS nicotinic and nuscarinic effects. Ann. N. Y. Acad. Sci. 1967, 142, 181–189. [Google Scholar] [CrossRef]
- Volgin, A.D.; Bashirzade, A.; Amstislavskaya, T.G.; Yakovlev, O.A.; Demin, K.A.; Ho, Y.J.; Wang, D.; Shevyrin, V.A.; Yan, D.; Tang, Z.; et al. DARK classics in chemical neuroscience: Arecoline. ACS Chem. Neurosci. 2019, 10, 2176–2185. [Google Scholar] [CrossRef]
- Xiao, B.; Xiao, N.; Peng, M.; Gong, L.; Li, C.; Lin, L. Research on effects of arecoline on refreshing and acute toxicity test. China Mod. Med. 2013, 20, 14–16. [Google Scholar]
- Arias-Carrión, O.; Caraza-Santiago, X.; Salgado-Licona, S.; Salama, M.; Machado, S.; Nardi, A.E.; Menéndez-González, M.; Murillo-Rodríguez, E. Orquestic regulation of neurotransmitters on reward-seeking behavior. Int. Arch. Med. 2014, 7, 29. [Google Scholar] [CrossRef]
- Begum, N.; Mandhare, A.; Tryphena, K.P.; Srivastava, S.; Shaikh, M.F.; Singh, S.B.; Khatri, D.K. Epigenetics in depression and gut-brain axis: A molecular crosstalk. Front. Aging Neurosci. 2022, 14, 1048333. [Google Scholar] [CrossRef]
- Tao, R.; Liu, S.; Crawford, J.; Tao, F. Gut-Brain Crosstalk and the Central Mechanisms of Orofacial Pain. Brain. Sci. 2023, 13, 1456. [Google Scholar] [CrossRef]
- Mahbub, N.U.; Islam, M.M.; Hong, S.T.; Chung, H.J. Dysbiosis of the gut microbiota and its effect on α-synuclein and prion protein misfolding: Consequences for neurodegeneration. Front. Cell. Infect. Microbiol. 2024, 14, 1348279. [Google Scholar] [CrossRef]
- Li, Y.; Luo, Z.Y.; Hu, Y.Y.; Bi, Y.W.; Yang, J.M.; Zou, W.J.; Song, Y.L.; Li, S.; Shen, T.; Li, S.J.; et al. The gut microbiota regulates autism-like behavior by mediating vitamin B6 homeostasis in EphB6-deficient mice. Microbiome 2020, 8, 120. [Google Scholar] [CrossRef]
- Dohnalová, L.; Lundgren, P.; Carty, J.R.E.; Goldstein, N.; Wenski, S.L.; Nanudorn, P.; Thiengmag, S.; Huang, K.P.; Litichevskiy, L.; Descamps, H.C.; et al. A microbiome-dependent gut-brain pathway regulates motivation for exercise. Nature 2022, 612, 739–747. [Google Scholar] [CrossRef]
- Vafadari, B. Stress and the role of the gut-brain axis in the pathogenesis of schizophrenia: A Literature review. Int. J. Mol. Sci. 2021, 22, 9747. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Zhou, M.; Fan, Y.; Xu, L.; Yu, Z.; Wang, S.; Xu, H.; Zhang, J.; Zhang, L.; Liu, W.; Wu, L.; et al. Microbiome and tryptophan metabolomics analysis in adolescent depression: Roles of the gut microbiota in the regulation of tryptophan-derived neurotransmitters and behaviors in human and mice. Microbiome 2023, 11, 145. [Google Scholar] [CrossRef]
- Zhuang, Z.; Yang, R.; Wang, W.; Qi, L.; Huang, T. Associations between gut microbiota and Alzheimer’s disease, major depressive disorder, and schizophrenia. J. Neuroinflammation 2020, 17, 288. [Google Scholar] [CrossRef]
- Ketchesin, K.D.; Becker-Krail, D.; McClung, C.A. Mood-related central and peripheral clocks. Eur. J. Neurosci. 2020, 51, 326–345. [Google Scholar] [CrossRef]
- Hirayama, M.; Mure, L.S.; Le, H.D.; Panda, S. Neuronal reprogramming of mouse and human fibroblasts using transcription factors involved in suprachiasmatic nucleus development. iScience 2024, 27, 109051. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Konturek, T.J.; Martinez, C.; Niesler, B.; van der Voort, I.; Mönnikes, H.; Stengel, A.; Goebel-Stengel, M. The role of brain-derived neurotrophic factor in irritable bowel syndrome. Front. Psychiatry 2021, 11, 531385. [Google Scholar] [CrossRef]
- Ragab, A.E.; Badawy, E.T.; Aboukhatwa, S.M.; Kabbash, A.; Abo El-Seoud, K.A. In vitro characterization of inhibitors for lung A549 and leukemia K562 cell lines from fungal transformation of arecoline supported by in silico docking to M3-mAChR and ADME prediction. Pharmaceuticals 2022, 15, 1171. [Google Scholar] [CrossRef]
- Paydar, A.; Lee, B.; Gangadharan, G.; Lee, S.; Hwang, E.M.; Shin, H.S. Extrasynaptic GABAA receptors in mediodorsal thalamic nucleus modulate fear extinction learning. Mol. Brain 2014, 7, 39. [Google Scholar] [CrossRef]
- Neumann, J.; Hofmann, B.; Dhein, S.; Gergs, U. Role of Dopamine in the Heart in Health and Disease. Int. J. Mol. Sci. 2023, 24, 5042. [Google Scholar] [CrossRef]
- Tavares, G.; Martins, F.O.; Melo, B.F.; Matafome, P.; Conde, S.V. Peripheral Dopamine Directly Acts on Insulin-Sensitive Tissues to Regulate Insulin Signaling and Metabolic Function. Front. Pharmacol. 2021, 12, 713418. [Google Scholar] [CrossRef]
- Li, M.; Zhou, L.; Sun, X.; Yang, Y.; Zhang, C.; Wang, T.; Fu, F. Dopamine, a co-regulatory component, bridges the central nervous system and the immune system. Biomed. Pharmacother. 2022, 145, 112458. [Google Scholar] [CrossRef]
- Császár-Nagy, N.; Bob, P.; Bókkon, I. A Multidisciplinary Hypothesis about Serotonergic Psychedelics. Is it Possible that a Portion of Brain Serotonin Comes from the Gut? J. Integr. Neurosci. 2022, 21, 148. [Google Scholar] [CrossRef]
- Neumann, J.; Hofmann, B.; Dhein, S.; Gergs, U. Cardiac Roles of Serotonin (5-HT) and 5-HT-Receptors in Health and Disease. Int. J. Mol. Sci. 2023, 24, 4765. [Google Scholar] [CrossRef]
- Flaive, A.; Fougère, M.; van der Zouwen, C.I.; Ryczko, D. Serotonergic Modulation of Locomotor Activity from Basal Vertebrates to Mammals. Front. Neural. Circuits 2020, 14, 590299. [Google Scholar] [CrossRef]
- Kang, J.Y.; Kim, D.Y.; Lee, J.S.; Hwang, S.J.; Kim, G.H.; Hyun, S.H.; Son, C.G. Korean Red Ginseng Ameliorates Fatigue via Modulation of 5-HT and Corticosterone in a Sleep-Deprived Mouse Model. Nutrients. 2021, 13, 3121. [Google Scholar] [CrossRef]
- Meeter, M.; Talamini, L.; Schmitt, J.A.; Riedel, W.J. Effects of 5-HT on memory and the hippocampus: Model and data. Neuropsychopharmacology 2006, 31, 712–720. [Google Scholar] [CrossRef]
- Zhou, M.; Li, X.; Liu, J.; Wu, Y.; Tan, Z.; Deng, N. Adenine’s impact on mice’s gut and kidney varies with the dosage administered and relates to intestinal microorganisms and enzyme activities. 3 Biotech 2024, 14, 88. [Google Scholar] [CrossRef]
- Rasouli-Saravani, A.; Jahankhani, K.; Moradi, S.; Gorgani, M.; Shafaghat, Z.; Mirsanei, Z.; Mehmandar, A.; Mirzaei, R. Role of microbiota short-chain fatty acids in the pathogenesis of autoimmune diseases. Biomed. Pharmacother. 2023, 162, 114620. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Luu, M.; Pautz, S.; Kohl, V.; Singh, R.; Romero, R.; Lucas, S.; Hofmann, J.; Raifer, H.; Vachharajani, N.; Carrascosa, L.C.; et al. The short-chain fatty acid pentanoate suppresses autoimmunity by modulating the metabolic-epigenetic crosstalk in lymphocytes. Nat. Commun. 2019, 10, 760. [Google Scholar] [CrossRef]
- Nishida, K.; Sawada, D.; Kuwano, Y.; Tanaka, H.; Rokutan, K. Health Benefits of Lactobacillus gasseri CP2305 Tablets in Young Adults Exposed to Chronic Stress: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2019, 11, 1859. [Google Scholar] [CrossRef]
- Yang, T.; Chen, S.; Qiu, L.; Guo, Q.; Wang, Z.; Jiang, Y.; Bai, H.; Bi, Y.; Chang, G. Effect of high dietary iron on fat deposition and gut microbiota in chickens. Animals 2024, 14, 2254. [Google Scholar] [CrossRef]
- van der Stel, A.X.; Wösten, M.M.S.M. Regulation of respiratory pathways in campylobacterota: A review. Front. Microbiol. 2019, 10, 1719. [Google Scholar] [CrossRef]
- Zhang, B.; Dong, W.; Ma, Z.; Duan, S.; Han, R.; Lv, Z.; Liu, X.; Mao, Y. Hyperbaric oxygen improves depression-like behaviors in chronic stress model mice by remodeling gut microbiota and regulating host metabolism. CNS Neurosci. Ther. 2023, 29, 239–255. [Google Scholar] [CrossRef]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The food-gut axis: Lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol. Rev. 2020, 44, 454–489. [Google Scholar] [CrossRef]
- Larini, I.; Tintori, S.; Gatto, V.; Felis, G.E.; Salvetti, E.; Torriani, S. Comparative genomics reveals the potential biotechnological applications of Liquorilactobacillus nagelii VUCC-R001, a strain isolated from kombucha tea. Food Biosci. 2024, 59, 104001. [Google Scholar] [CrossRef]
- Dürre, P. Physiology and Sporulation in Clostridium. Microbiol. Spectr. 2014, 2, TBS-0010-2012. [Google Scholar] [CrossRef]
- Guo, P.; Zhang, K.; Ma, X.; He, P. Clostridium species as probiotics: Potentials and challenges. J. Anim. Sci. Biotechnol. 2020, 11, 24. [Google Scholar] [CrossRef]
- Jessen, G.L.; Lichtschlag, A.; Struck, U.; Boetius, A. Distribution and composition of thiotrophic mats in the hypoxic zone of the black sea (150–170 m Water Depth, Crimea Margin). Front. Microbiol. 2016, 7, 1011. [Google Scholar] [CrossRef]
- Sharndama, H.C.; Mba, I.E. Helicobacter pylori: An up-to-date overview on the virulence and pathogenesis mechanisms. Braz. J. Microbiol. 2022, 53, 33–50. [Google Scholar] [CrossRef]
- Pirzadeh, M.; Khalili, N.; Rezaei, N. The interplay between aryl hydrocarbon receptor, H. pylori, tryptophan, and arginine in the pathogenesis of gastric cancer. Int. Rev. Immunol. 2022, 41, 299–312. [Google Scholar] [CrossRef]
- Waskito, L.A.; Salama, N.R.; Yamaoka, Y. Pathogenesis of helicobacter pylori infection. Helicobacter 2018, 23, e12516. [Google Scholar] [CrossRef]
- Wu, Y.; Peng, X.; Li, X.; Li, D.; Tan, Z.; Yu, R. Sex hormones influence the intestinal microbiota composition in mice. Front. Microbiol. 2022, 13, 964847. [Google Scholar] [CrossRef]
- Zhou, K.; Deng, N.; Yi, X.; Cai, Y.; Peng, M.; Xiao, N. Baohe pill decoction for diarrhea induced by high-fat and high-protein diet is associated with the structure of lactase-producing bacterial community. Front. Cell. Infect. Microbiol. 2022, 12, 1004845. [Google Scholar] [CrossRef]
- Fang, L.; Shen, J.; Wu, Y.; Tan, Z. Involvement of intestinal mucosal microbiota in adenine-induced liver function injury. 3 Biotech 2025, 15, 6. [Google Scholar] [CrossRef]
- Shen, J.; Fang, L.; Tan, Z.; Xiao, N.; Peng, M. The Effects of Functional Biscuits on Intestinal Mucosal Microbiota Composition, Brain Function, and Antioxidant Activity. Biosci. Microbiota Food Health 2025, 44, 2024-078. [Google Scholar] [CrossRef]
- Liu, J.; Qiao, B.; Cai, Y.; Tan, Z.; Deng, N. Diarrhea accompanies intestinal inflammation and intestinal mucosal microbiota dysbiosis during fatigue combined with a high-fat diet. BMC Microbiol. 2023, 23, 151. [Google Scholar] [CrossRef]
- Zhou, M.; Li, X.; Wang, X.; Deng, N.; Cai, Y.; Tan, Z. The dysfunction in intestinal microorganisms and enzyme activity as significant contributors to diarrhea with kidney-yang deficiency syndrome. Front. Microbiol. 2024, 14, 1324938. [Google Scholar] [CrossRef]
- Schnürer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef]
- Hui, H.; Wu, Y.; Zheng, T.; Zhou, S.; Tan, Z. Bacterial Characteristics in Intestinal Contents of Antibiotic-Associated Diarrhea Mice Treated with Qiweibaizhu Powder. Med. Sci. Monit. 2020, 26, e921771. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Distance Traveled | Average Speed | Time Spent in Fast Movement | Number of Entries into the Center | ||
|---|---|---|---|---|---|
| Day 7 | BC | 1588.07 ± 320.74 | 4.41 ± 0.89 | 7.70 ± 3.55 | 18.80 ± 6.57 |
| AC | 1634.35 ± 357.02 | 4.54 ± 0.99 | 6.45 ± 3.50 | 18.60 ± 4.16 | |
| Day 14 | BC | 1195.87 ± 104.94 | 3.32 ± 0.29 | 5.19 ± 1.89 | 7.80 ± 4.32 |
| AC | 1299.96 ± 252.78 | 3.61 ± 0.70 | 6.21 ± 3.15 | 6.00 ± 3.32 | |
| Day 21 | BC | 1195.87 ± 104.94 | 3.32 ± 0.29 | 4.48 ± 1.12 | 7.40 ± 1.14 |
| AC | 1299.96 ± 252.78 | 3.61 ± 0.70 | 8.30 ± 3.12 * | 14.00 ± 1.41 * | |
| Day 28 | BC | 1002.55 ± 398.32 | 2.78 ± 1.11 | 4.48 ± 2.26 | 3.80 ± 0.45 |
| AC | 977.68 ± 166.76 | 2.72 ± 0.47 | 5.28 ± 1.34 | 8.60 ± 2.30 * | |
| Steps | Conditions |
|---|---|
| Column temperature requirement | Initial Temperature: 100 °C, held for 5 min Ramp to 150 °C at 5 °C/min (no hold) Immediate Ramp to 240 °C at 30 °C/min Final Temperature: 240 °C, held for 30 min |
| Flow rate requirement | 1 mL/min |
| Shunt ratio | 75:1 |
| Carrier gas | Helium |
| Chromatographic column | TG WAX 30 m × 0.25 mm × 0.25 μm |
| Injector | 240 °C |
| Bombardment voltage | 70 eV |
| Single-ion scan mode | Quantitative ion 63, 70 |
| Ion source temperature | 200 °C |
| Connection line temperature | 250 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, J.; Zhou, M.; Xiao, N.; Tan, Z.; Liang, X. Unveiling the Mystery of the Stimulatory Effects of Arecoline: Its Relevance to the Regulation of Neurotransmitters and the Microecosystem in Multi-Ecological Intestinal Sites. Int. J. Mol. Sci. 2025, 26, 3150. https://doi.org/10.3390/ijms26073150
Shen J, Zhou M, Xiao N, Tan Z, Liang X. Unveiling the Mystery of the Stimulatory Effects of Arecoline: Its Relevance to the Regulation of Neurotransmitters and the Microecosystem in Multi-Ecological Intestinal Sites. International Journal of Molecular Sciences. 2025; 26(7):3150. https://doi.org/10.3390/ijms26073150
Chicago/Turabian StyleShen, Junxi, Mengsi Zhou, Nenqun Xiao, Zhoujin Tan, and Xuejuan Liang. 2025. "Unveiling the Mystery of the Stimulatory Effects of Arecoline: Its Relevance to the Regulation of Neurotransmitters and the Microecosystem in Multi-Ecological Intestinal Sites" International Journal of Molecular Sciences 26, no. 7: 3150. https://doi.org/10.3390/ijms26073150
APA StyleShen, J., Zhou, M., Xiao, N., Tan, Z., & Liang, X. (2025). Unveiling the Mystery of the Stimulatory Effects of Arecoline: Its Relevance to the Regulation of Neurotransmitters and the Microecosystem in Multi-Ecological Intestinal Sites. International Journal of Molecular Sciences, 26(7), 3150. https://doi.org/10.3390/ijms26073150