
Lessons from RatA: Why the Basics in Molecular Biology Are Still Crucial!


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Overexpression of ratA Leads to the Production of Two RatA Variants
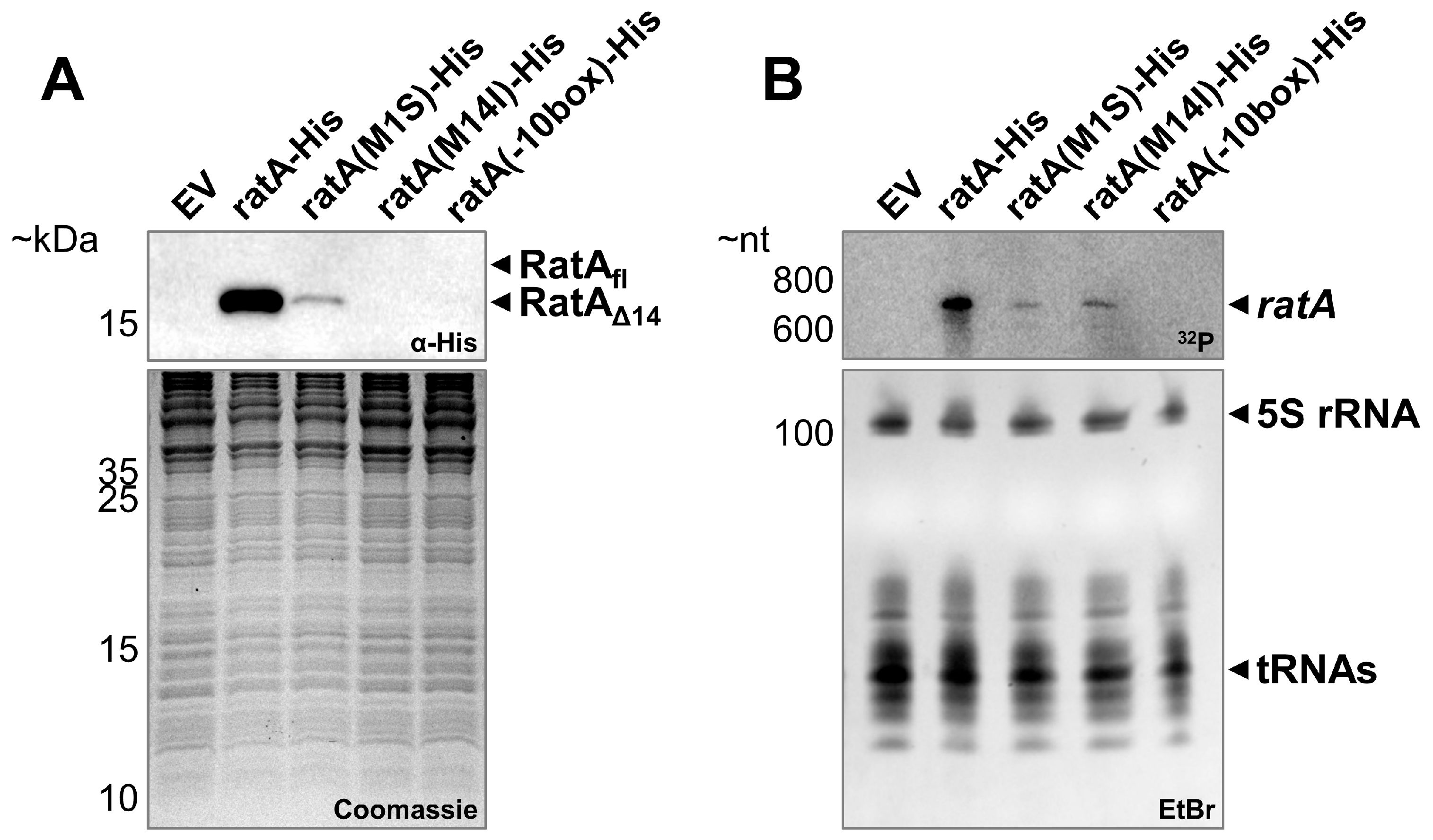
2.2. Full-Length ratA Is Responsible for the Observed Toxic Effect
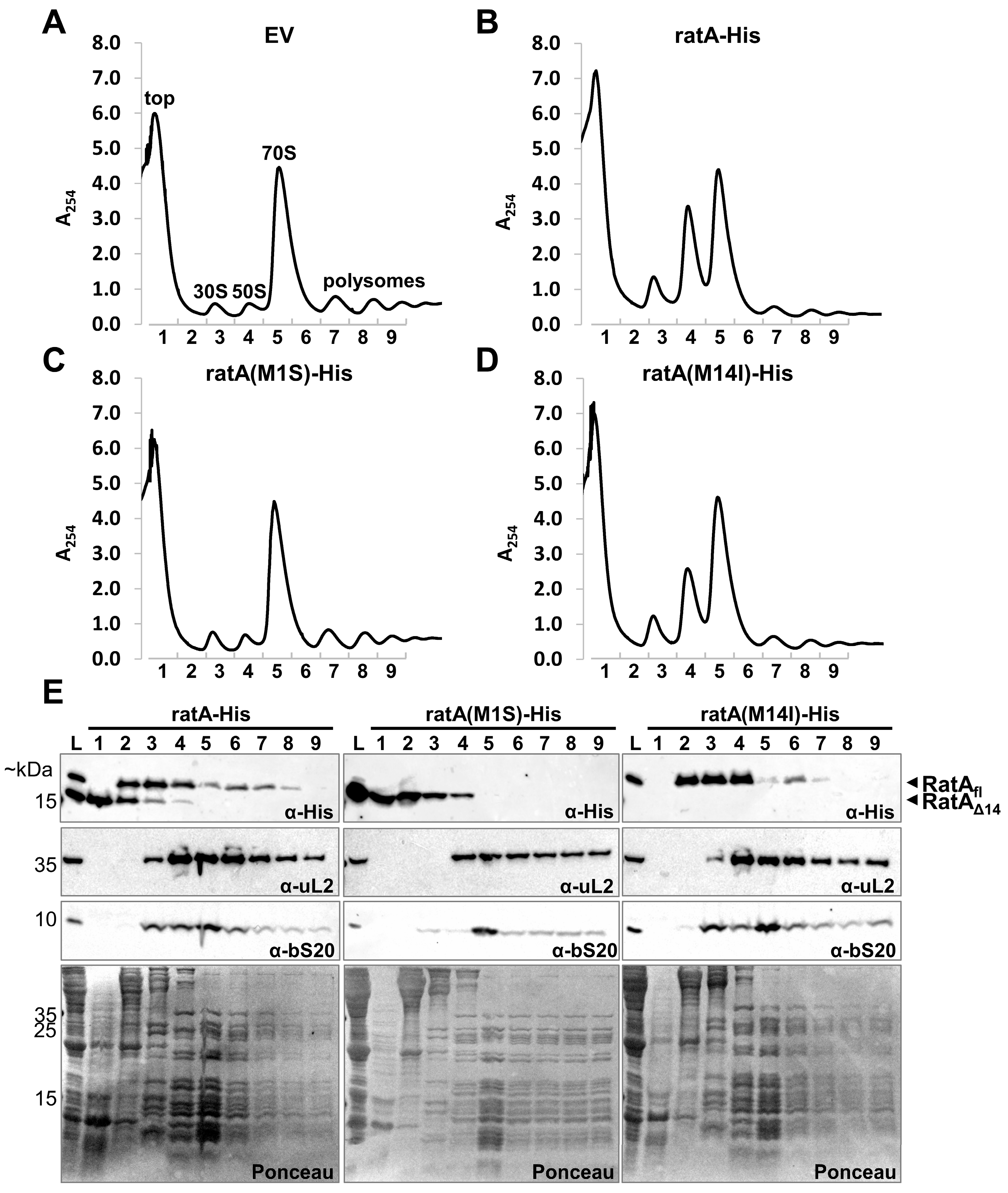
2.3. RatAfl Does Not Exclusively Target the 50S Subunit
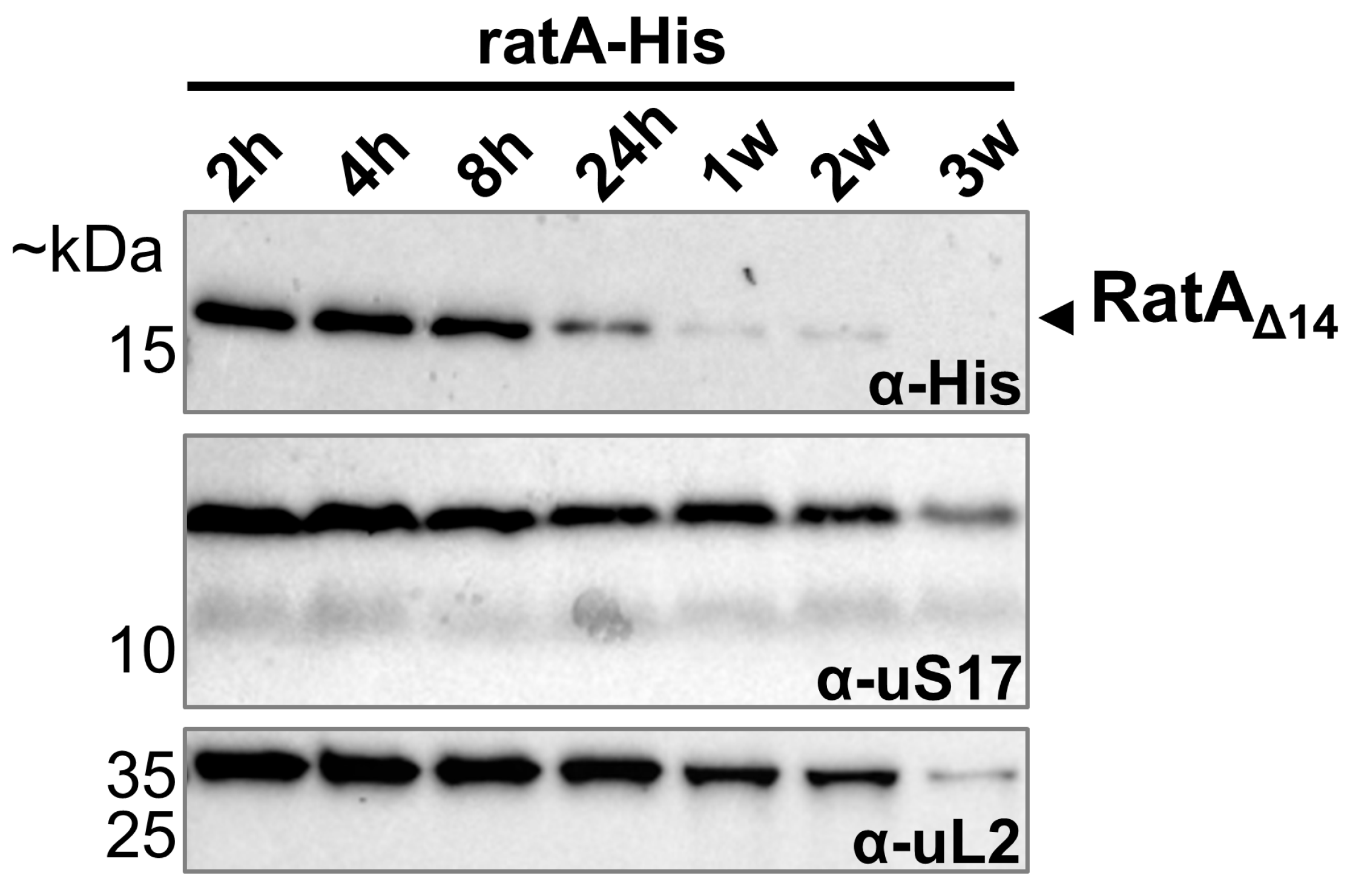
2.4. Only ratAΔ14 Is Detected in Unstressed Cells
2.5. The ratA σ70 Promoter Is Partially Located Inside the Annotated ORF
2.6. Only RatAΔ14 Is Detected over Different Growth Phases Specifically Under Aerobic Conditions
3. Discussion
3.1. The Toxic Effect of RatA Is an Artifact of Overexpressing an Incorrectly Annotated Gene
3.2. False Annotations of the Initiating Start Codon Could Be More Widespread
4. Materials and Methods
4.1. Bacterial Strains and Plasmids Construction
4.2. Bacterial Culturing and Growth Conditions
4.3. Colony Size Determination
4.4. Western Blot Analysis
4.5. Sucrose Gradient Analysis
4.6. Northern Blot and Primer Extension Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NCBI | National Center for Biotechnology Information |
| RatA | Ribosome association toxin A |
| TA | Toxin–antitoxin |
| UPEC | Uropathogenic E. coli |
| ORF | Open reading frame |
| TSS | Transcription start site |
| UniProtKB | UniProt Knowledgebase |
| gDNA | Genomic DNA |
References
- Vila, J.; Sáez-López, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia coli: An Old Friend with New Tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef] [PubMed]
- Blattner, F.R.; Plunkett, G.; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The Complete Genome Sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Farrell, C.M.; Feldgarden, M.; Fine, A.M.; Funk, K.; et al. Database Resources of the National Center for Biotechnology Information in 2023. Nucleic Acids Res. 2023, 51, D29–D38. [Google Scholar] [CrossRef] [PubMed]
- Karp, P.D.; Paley, S.; Caspi, R.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Moore, L.R.; Subhraveti, P.; Gama-Castro, S.; Tierrafria, V.H.; et al. The EcoCyc Database (2023). EcoSal Plus 2023, 11, eesp-0002-2023. [Google Scholar] [CrossRef]
- Brown, J.M.; Shaw, K.J. A Novel Family of Escherichia coli Toxin-Antitoxin Gene Pairs. J. Bacteriol. 2003, 185, 6600–6608. [Google Scholar] [CrossRef]
- Singh, G.; Yadav, M.; Ghosh, C.; Rathore, J.S. Bacterial Toxin-Antitoxin Modules: Classification, Functions, and Association with Persistence. Curr. Res. Microb. Sci. 2021, 2, 100047. [Google Scholar] [CrossRef]
- Zhang, Y.; Inouye, M. RatA (YfjG), an Escherichia coli Toxin, Inhibits 70S Ribosome Association to Block Translation Initiation. Mol. Microbiol. 2011, 79, 1418–1429. [Google Scholar] [CrossRef]
- Norton, J.P.; Mulvey, M.A. Toxin-Antitoxin Systems Are Important for Niche-Specific Colonization and Stress Resistance of Uropathogenic Escherichia coli. PLoS Pathog. 2012, 8, e1002954. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef]
- Fino, C.; Vestergaard, M.; Ingmer, H.; Pierrel, F.; Gerdes, K.; Harms, A. PasT of Escherichia coli Sustains Antibiotic Tolerance and Aerobic Respiration as a Bacterial Homolog of Mitochondrial Coq10. Microbiologyopen 2020, 9, e1064. [Google Scholar] [CrossRef]
- Vesper, O.; Amitai, S.; Belitsky, M.; Byrgazov, K.; Kaberdina, A.C.; Engelberg-Kulka, H.; Moll, I. Selective Translation of Leaderless mRNAs by Specialized Ribosomes Generated by MazF in Escherichia coli. Cell 2011, 147, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Temmel, H.; Müller, C.; Sauert, M.; Vesper, O.; Reiss, A.; Popow, J.; Martinez, J.; Moll, I. The RNA Ligase RtcB Reverses MazF-Induced Ribosome Heterogeneity in Escherichia coli. Nucleic Acids Res. 2017, 45, 4708–4721. [Google Scholar] [CrossRef] [PubMed]
- Sauert, M.; Wolfinger, M.T.; Vesper, O.; Müller, C.; Byrgazov, K.; Moll, I. The MazF-Regulon: A Toolbox for the Post-Transcriptional Stress Response in Escherichia coli. Nucleic Acids Res. 2016, 44, 6660–6675. [Google Scholar] [CrossRef]
- Thomason, M.K.; Bischler, T.; Eisenbart, S.K.; Förstner, K.U.; Zhang, A.; Herbig, A.; Nieselt, K.; Sharma, C.M.; Storz, G. Global Transcriptional Start Site Mapping Using Differential RNA Sequencing Reveals Novel Antisense RNAs in Escherichia coli. J. Bacteriol. 2015, 197, 18–28. [Google Scholar] [CrossRef]
- Reier, K.; Lahtvee, P.-J.; Liiv, A.; Remme, J. A Conundrum of R-Protein Stability: Unbalanced Stoichiometry of r-Proteins during Stationary Phase in Escherichia coli. mBio 2022, 13, e01873-22. [Google Scholar] [CrossRef]
- Battesti, A.; Majdalani, N.; Gottesman, S. The RpoS-Mediated General Stress Response in Escherichia coli. Annu. Rev. Microbiol. 2011, 65, 189–213. [Google Scholar]
- Fasnacht, M.; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front. Mol. Biosci. 2021, 8, 392. [Google Scholar] [CrossRef]
- Li, X.; Mei, H.; Chen, F.; Tang, Q.; Yu, Z.; Cao, X.; Andongma, B.T.; Chou, S.-H.; He, J. Transcriptome Landscape of Mycobacterium Smegmatis. Front. Microbiol. 2017, 8, 2505. [Google Scholar] [CrossRef]
- Mendoza-Vargas, A.; Olvera, L.; Olvera, M.; Grande, R.; Vega-Alvarado, L.; Taboada, B.; Jimenez-Jacinto, V.; Salgado, H.; Juárez, K.; Contreras-Moreira, B.; et al. Genome-Wide Identification of Transcription Start Sites, Promoters and Transcription Factor Binding Sites in E. Coli. PLoS ONE 2009, 4, e7526. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-Step Inactivation of Chromosomal Genes in Escherichia coli K-12 Using PCR Products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight Regulation, Modulation, and High-Level Expression by Vectors Containing the Arabinose PBAD Promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed]
- Lutz, R.; Bujard, H. Independent and Tight Regulation of Transcriptional Units in Escherichia coli Via the LacR/O, the TetR/O and AraC/I1-I2 Regulatory Elements. Nucleic Acids Res. 1997, 25, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Solovyev, S.V.; Salamov, A.; Li, R.W. Automatic Annotation of Microbial Genomes and Metagenomic Acts. Gen Metagenomics and Its Applications in Agriculture, Biomedicine and Environmental Studies; Nova Science Publishers: Hauppauge, NY, USA, 2011. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fasnacht, M.; Schratt, D.; Moll, I. Lessons from RatA: Why the Basics in Molecular Biology Are Still Crucial! Int. J. Mol. Sci. 2025, 26, 3100. https://doi.org/10.3390/ijms26073100
Fasnacht M, Schratt D, Moll I. Lessons from RatA: Why the Basics in Molecular Biology Are Still Crucial! International Journal of Molecular Sciences. 2025; 26(7):3100. https://doi.org/10.3390/ijms26073100
Chicago/Turabian StyleFasnacht, Michel, Denise Schratt, and Isabella Moll. 2025. "Lessons from RatA: Why the Basics in Molecular Biology Are Still Crucial!" International Journal of Molecular Sciences 26, no. 7: 3100. https://doi.org/10.3390/ijms26073100
APA StyleFasnacht, M., Schratt, D., & Moll, I. (2025). Lessons from RatA: Why the Basics in Molecular Biology Are Still Crucial! International Journal of Molecular Sciences, 26(7), 3100. https://doi.org/10.3390/ijms26073100