
Potential Use of Selected Natural Compounds with Anti-Biofilm Activity


Abstract

1. Introduction
2. Quercetin
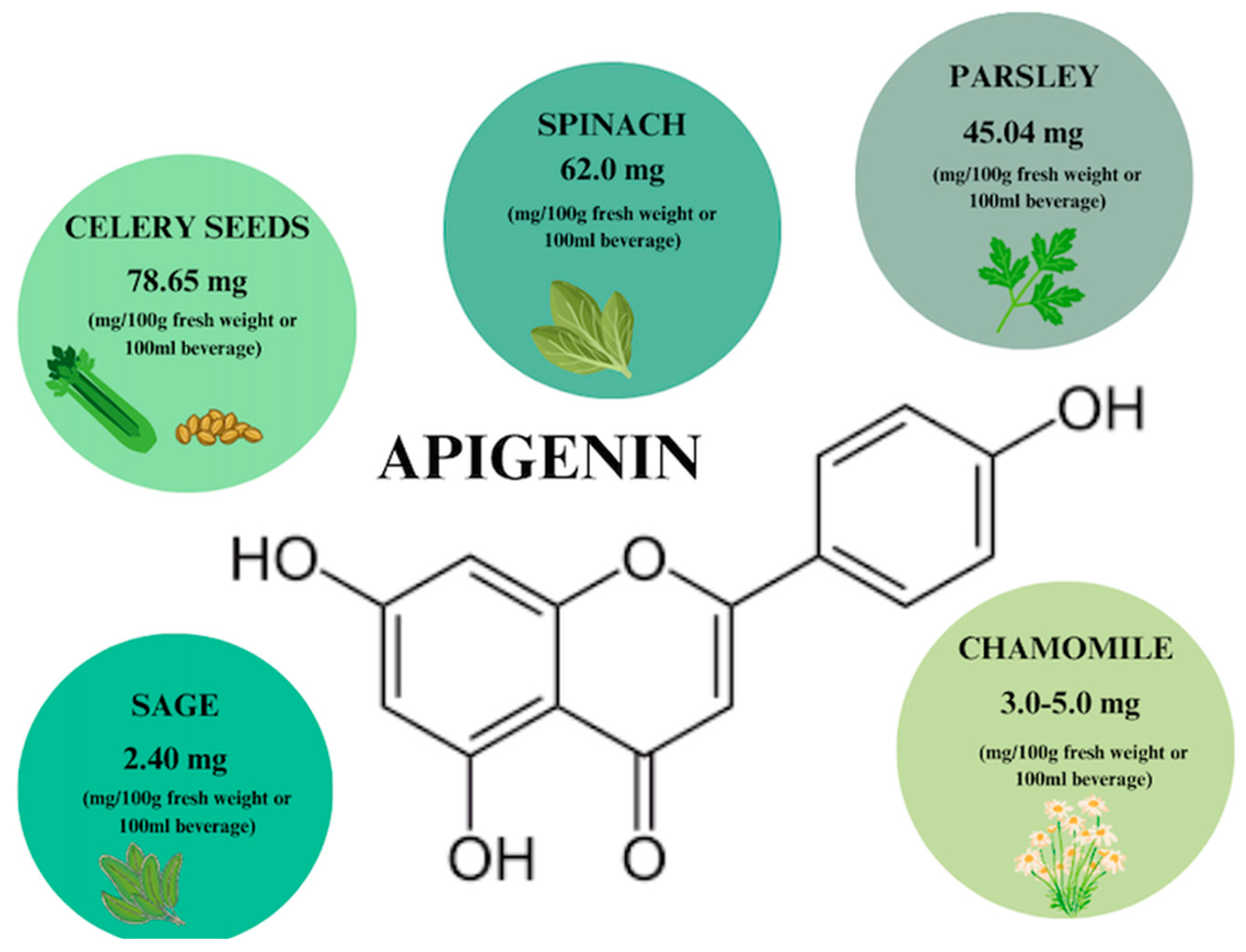
3. Apigenin
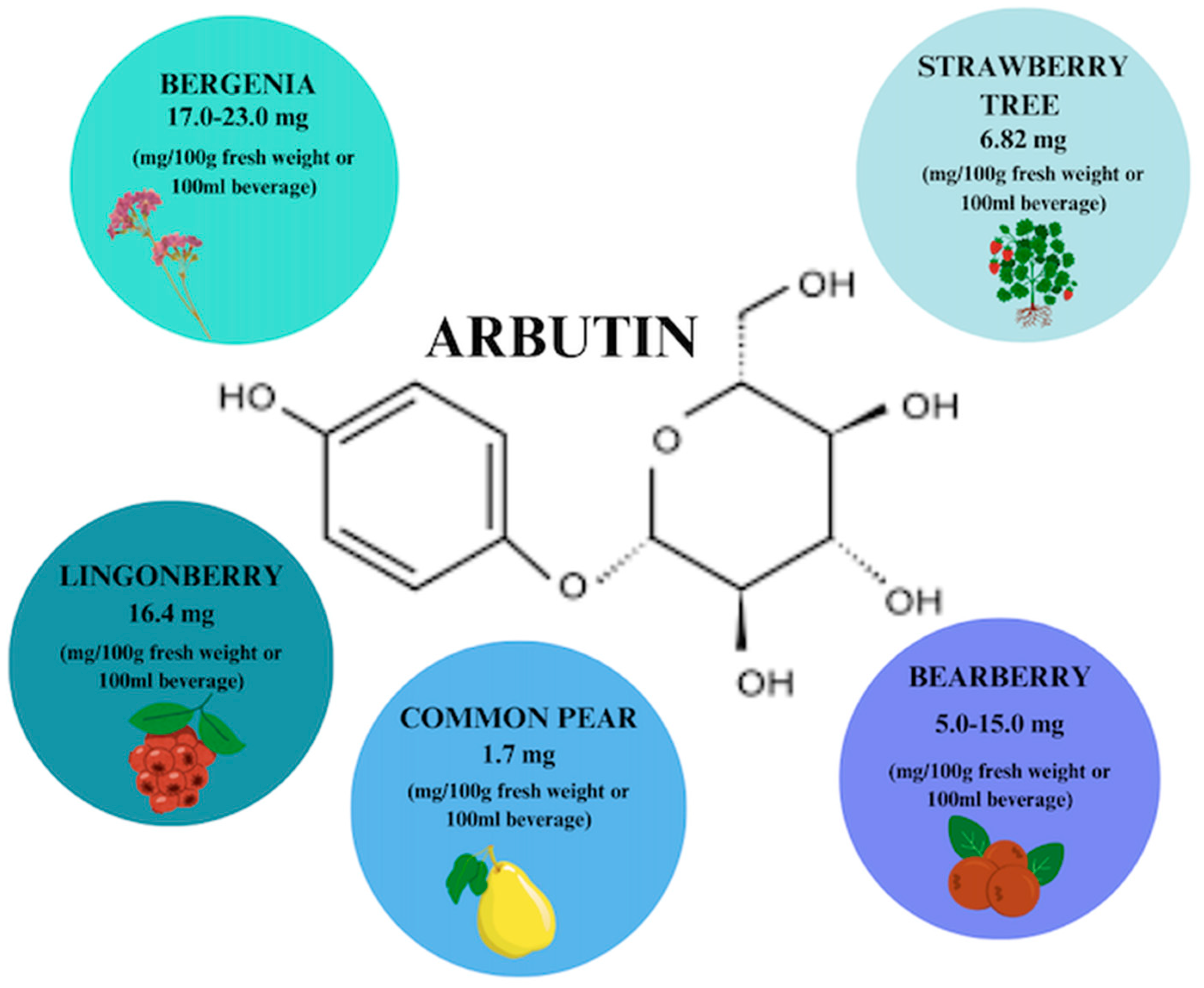
4. Arbutin
5. Proanthocyanidin
6. Vitamin C
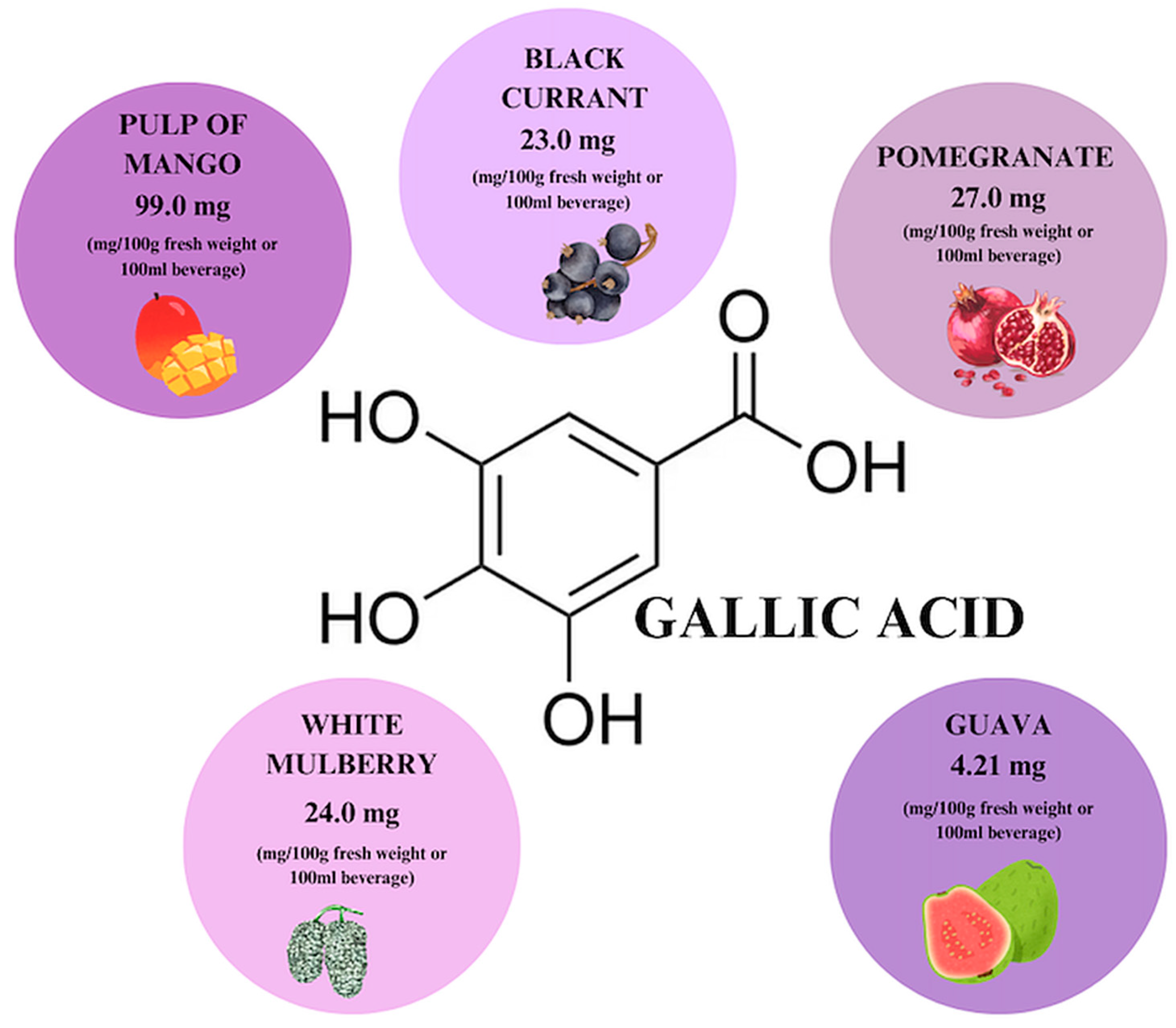
7. Gallic Acid
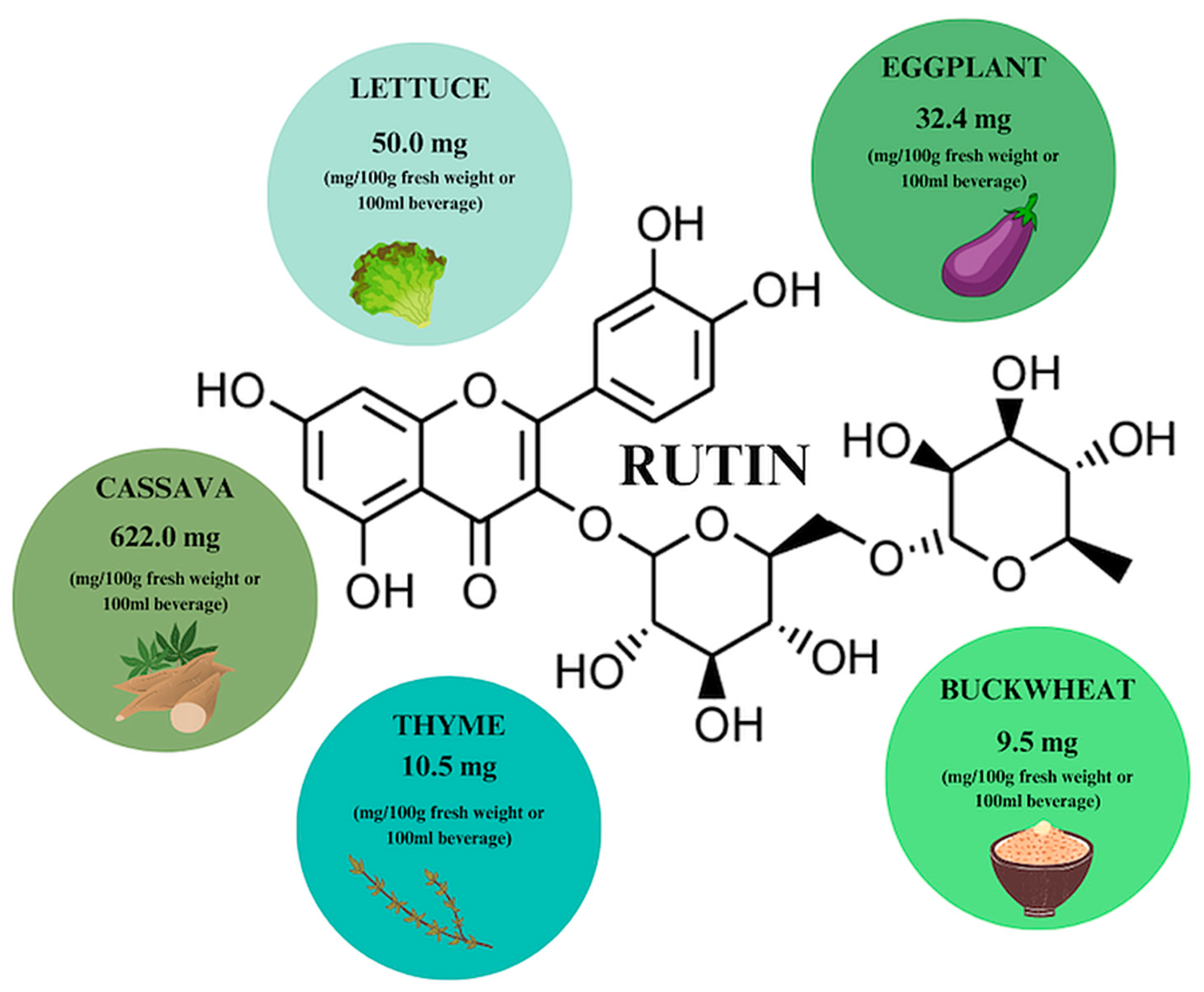
8. Rutin
9. Discussion
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asma, S.T.; Imre, K.; Morar, A.; Imre, M.; Acaroz, U.; Shah, S.R.A.; Hussain, S.Z.; Arslan-Acaroz, D.; Istanbullugil, F.R.; Madani, K.; et al. Natural Strategies as Potential Weapons against Bacterial Biofilms. Life 2022, 12, 1618. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Gupta, K.; Mandal, M. Microbial biofilm: Formation, architecture, antibiotic resistance, and control strategies. Braz. J. Microbiol. 2021, 52, 1701–1718. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Hansen, M.F.; Burmølle, M.; Heyndrickx, M.; Flint, S.; Lu, W.; Chen, W.; Zhang, H. Trans-kingdom interactions in mixed biofilm communities. FEMS Microbiol. Rev. 2022, 46, fuac024. [Google Scholar] [CrossRef]
- Rahman, K.A.; Sudarsan, R.; Eberl, H.J. A Mixed-Culture Biofilm Model with Cross-Diffusion. Bull. Math. Biol. 2015, 77, 2086–2124. [Google Scholar] [CrossRef]
- Ruhal, R.; Kataria, R. Biofilm patterns in gram-positive and gram-negative bacteria. Microbiol. Res. 2021, 251, 126829. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic treatment of biofilm infections. Acta Pathol. Microbiol. Immunol. Scand. 2017, 125, 304–319. [Google Scholar] [CrossRef] [PubMed]
- Lowery, N.V.; McNally, L.; Ratcliff, W.C.; Brown, S.P. Division of Labor, Bet Hedging, and the Evolution of Mixed Biofilm Investment Strategies. mBio 2017, 8, e00672-17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.L.; Hou, H.M.; Zhang, G.L.; Wang, Y.F.; Hao, H.S. AHLs Regulate Biofilm Formation and Swimming Motility of Hafnia alvei H4. Front. Microbiol. 2019, 10, 1330. [Google Scholar] [CrossRef] [PubMed]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural products as inspiration for the development of bacterial anti-biofilm agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Mukherjee, S.; Mukherjee, R.; Haldar, J. Small molecular adjuvants repurpose antibiotics towards Gram-negative bacterial infections and multispecies bacterial biofilms. Chem. Sci. 2024, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Corona, A.; De Santis, V.; Agarossi, A.; Prete, A.; Cattaneo, D.; Tomasini, G.; Bonetti, G.; Patroni, A.; Latronico, N. Antibiotic Therapy Strategies for Treating Gram-Negative Severe Infections in the Critically Ill: A Narrative Review. Antibiotics 2023, 12, 1262. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed Ahmed, M.A.E.G.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed]
- Grégoire, N.; Aranzana-Climent, V.; Magréault, S.; Marchand, S.; Couet, W. Clinical Pharmacokinetics and Pharmacodynamics of Colistin. Clin. Pharmacokinet. 2017, 56, 1441–1460. [Google Scholar] [CrossRef] [PubMed]
- Magar, R.T.; Sohng, J.K. A Review on Structure, Modifications and Structure-Activity Relation of Quercetin and Its Derivatives. J. Microbiol. Biotechnol. 2020, 30, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Tobar-Delgado, E.; Mejía-España, D.; Osorio-Mora, O.; Serna-Cock, L. Rutin: Family Farming Products’ Extraction Sources, Industrial Applications and Current Trends in Biological Activity Protection. Molecules 2023, 28, 5864. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Arbutin as a Skin Depigmenting Agent with Antimelanogenic and Antioxidant Properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef]
- Nemzer, B.V.; Al-Taher, F.; Yashin, A.; Revelsky, I.; Yashin, Y. Cranberry: Chemical Composition, Antioxidant Activity and Impact on Human Health: Overview. Molecules 2022, 27, 1503. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.H.; Seo, J.M.; Kim, N.H.; Arasu, M.V.; Kim, S.; Yoon, M.K.; Kim, S.-J. Variation of quercetin glycoside derivatives in three onion (Allium cepa L.) varieties. Saudi J. Biol. Sci. 2017, 24, 1387–1391. [Google Scholar] [CrossRef]
- Musini, A.; Singh, H.N.; Vulise, J.; Pammi, S.S.S.; Giri, A. Quercetin’s anti-biofilm effectiveness against drug-resistant Staphylococcus aureus and its validation by in silico modeling. Res. Microbiol. 2024, 175, 104091. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Yi, H.; Peng, H.; Wu, X.; Xu, X.; Kuang, T.; Zhang, J.; Du, L.; Fan, G. The Therapeutic Effects and Mechanisms of Quercetin on Metabolic Diseases: Pharmacological Data and Clinical Evidence. Oxid. Med. Cell. Longev. 2021, 2021, 6678662. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Badhwar, R.; Mangla, B.; Neupane, Y.R.; Khanna, K.; Popli, H. Quercetin loaded silver nanoparticles in hydrogel matrices for diabetic wound healing. Nanotechnology 2021, 32, 505102. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Bhattacharya, D. Antimicrobial Activity of Quercetin: An Approach to Its Mechanistic Principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef]
- Naeem, A.; Yu, C.; Zang, Z.; Zhu, W.; Deng, X.; Guan, Y. Synthesis and Evaluation of Rutin–Hydroxypropyl β-Cyclodextrin Inclusion Complexes Embedded in Xanthan Gum-Based (HPMC-g-AMPS) Hydrogels for Oral Controlled Drug Delivery. Antioxidants 2023, 12, 552. [Google Scholar] [CrossRef] [PubMed]
- DeRango-Adem, E.F.; Blay, J. Does Oral Apigenin Have Real Potential for a Therapeutic Effect in the Context of Human Gastrointestinal and Other Cancers? Front. Pharmacol. 2021, 12, 681477. [Google Scholar] [CrossRef] [PubMed]
- Ou, K.; Gu, L. Absorption and metabolism of proanthocyanidins. J. Funct. Foods 2014, 7, 43–53. [Google Scholar] [CrossRef]
- National Institutes of Health. Fact Sheet for Consumers. Vitamin C—Health Professional. Available online: https://ods.od.nih.gov/factsheets/VitaminC-HealthProfessional/#disc (accessed on 16 June 2024).
- Yang, K.; Zhang, L.; Liao, P.; Xiao, Z.; Zhang, F.; Sindaye, D.; Xin, Z.; Tan, C.; Deng, J.; Yin, Y.; et al. Impact of Gallic Acid on Gut Health: Focus on the Gut Microbiome, Immune Response, and Mechanisms of Action. Front. Immunol. 2020, 11, 580208. [Google Scholar] [CrossRef] [PubMed]
- Frutos, M.J.; Rincón-Frutos, L.; Valero-Cases, E. Rutin. In Nonvitamin and Nonmineral Nutritional Supplements; Elsevier: Amsterdam, The Netherlands, 2019; pp. 111–117. [Google Scholar]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Gobin, M.; Proust, R.; Lack, S.; Duciel, L.; Des Courtils, C.; Pauthe, E.; Gand, A.; Seyer, D. A Combination of the Natural Molecules Gallic Acid and Carvacrol Eradicates P. aeruginosa and S. aureus Mature Biofilms. Int. J. Mol. Sci. 2022, 23, 7118. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Basist, P.; Alhalmi, A.; Khan, R.; Noman, O.M.; Alahdab, A. Synthesis of Quercetin-Loaded Silver Nanoparticles and Assessing Their Anti-Bacterial Potential. Micromachines 2023, 14, 2154. [Google Scholar] [CrossRef]
- Dizeci, N.; Karaca, B.; Onar, O.; Cihan, A.C.; Akata, I.; Yildirim, O. The Remarkable Anti-biofilm Activity of the Sweet Tooth Mushroom, Hydnum repandum (Agaricomycetes), Displaying Synergetic Interactions with Antibiotics. Int. J. Med. Mushrooms 2021, 23, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.J.; Li, C.; Dai, W.; Lou, Z.; Sun, X.; Wang, H.; Khan, A.A.; Wan, C. The Anti-Biofilm Activity and Mechanism of Apigenin-7-O-Glucoside Against Staphylococcus aureus and Escherichia coli. Infect. Drug Resist. 2023, 16, 2129–2140. [Google Scholar] [CrossRef] [PubMed]
- Cela-López, J.M.; Camacho Roldán, C.J.; Gómez-Lizarraga, G.; Martínez, V. Effects of Itxasol© Components on Gene Expression in Bacteria Related to Infections of the Urinary Tract and to the Inflammation Process. Int. J. Mol. Sci. 2021, 22, 12655. [Google Scholar] [CrossRef] [PubMed]
- Weawsiangsang, S.; Rattanachak, N.; Ross, S.; Ross, G.M.; Baldock, R.A.; Jongjitvimol, T.; Jongjitwimol, J. Hydroquinine Enhances the Efficacy of Contact Lens Solutions for Inhibiting Pseudomonas aeruginosa Adhesion and Biofilm Formation. Antibiotics 2024, 13, 56. [Google Scholar] [CrossRef] [PubMed]
- Genovese, C.; D’Angeli, F.; Bellia, F.; Distefano, A.; Spampinato, M.; Attanasio, F.; Nicolosi, D.; Di Salvatore, V.; Tempera, G.; Lo Furno, D.; et al. In Vitro Antibacterial, Anti-Adhesive and Anti-Biofilm Activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson Root Extract against Methicillin-Resistant Staphylococcus aureus Strains. Antibiotics 2021, 10, 428. [Google Scholar]
- Maisuria, V.B.; Okshevsky, M.; Déziel, E.; Tufenkji, N. Proanthocyanidin Interferes with Intrinsic Antibiotic Resistance Mechanisms of Gram-Negative Bacteria. Adv. Sci. 2022, 9, e2202641. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, W.M.; Refaie, M.M.M.; Yousef, R.K.M.; Abd El Fatah, A.S.; Mousa, Y.M.; Rashwan, R. Assessment of Antibacterial and Anti-biofilm Effects of Vitamin C Against Pseudomonas aeruginosa Clinical Isolates. Front. Microbiol. 2022, 13, 847449. [Google Scholar] [CrossRef]
- Hassuna, N.A.; Rabie, E.M.; Mahd, W.K.M.; Refaie, M.M.M.; Yousef, R.K.M.; Abdelraheem, W.M. Antibacterial effect of vitamin C against uropathogenic E. coli in vitro and in vivo. BMC Microbiol. 2023, 23, 112. [Google Scholar] [CrossRef] [PubMed]
- Abdel Bar, F.M.; Alossaimi, M.A.; Elekhnawy, E.; Alzeer, M.A.A.; Abo Kamer, A.; Moglad, E.; ElNaggar, M.H. Anti-Quorum Sensing and Anti-Biofilm Activity of Pelargonium × hortorum Root Extract against Pseudomonas aeruginosa: Combinatorial Effect of Catechin and Gallic Acid. Molecules 2022, 27, 7841. [Google Scholar] [CrossRef]
- Al-Shabib, N.; Husain, F.; Ahmad, I.; Khan, M.; Khan, R.A.; Khan, J.M. Rutin inhibits mono and multi-species biofilm formation by foodborne drug-resistant Escherichia coli and Staphylococcus aureus. Food Control 2017, 79, 325–332. [Google Scholar] [CrossRef]
- Hu, W.S.; Nam, D.M.; Choi, J.Y.; Kim, J.S.; Koo, O.K. Anti-attachment, anti-biofilm, and antioxidant properties of Brassicaceae extracts on Escherichia coli O157:H7. Food Sci. Biotechnol. 2019, 28, 1881–1890. [Google Scholar] [CrossRef]
- Qaralleh, H. Chemical Composition and Quorum Sensing Inhibitory Effect of Nepeta curviflora Methanolic Extract against ESBL Pseudomonas aeruginosa. J. Pharmacopunct. 2023, 26, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Waheed, A.; Zameer, S.; Ashrafi, K.; Ali, A.; Sultana, N.; Aqil, M.; Sultana, Y.; Iqbal, Z. Insights into Pharmacological Potential of Apigenin through Various Pathways on a Nanoplatform in Multitude of Diseases. Curr. Pharm. Des. 2023, 29, 1326–1340. [Google Scholar] [CrossRef]
- Cannataro, R.; Fazio, A.; La Torre, C.; Caroleo, M.C.; Cione, E. Polyphenols in the Mediterranean Diet: From Dietary Sources to microRNA Modulation. Antioxidants 2021, 10, 328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, D.; Huang, Y.; Gao, Y.; Qian, S. Biopharmaceutics classification and intestinal absorption study of apigenin. Int. J. Pharm. 2012, 436, 311–317. [Google Scholar] [CrossRef]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. Biomed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: Food Sources, Bioavailability, Metabolism, and Bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, F.; Guo, Z.; Lei, J.; Zhou, B. Recent advancement in bioeffect, metabolism, stability, and delivery systems of apigenin, a natural flavonoid compound: Challenges and perspectives. Front. Nutr. 2023, 10, 1221227. [Google Scholar] [CrossRef] [PubMed]
- Nahar, L.; Al-Groshi, A.; Kumar, A.; Sarker, S.D. Arbutin: Occurrence in Plants, and Its Potential as an Anticancer Agent. Molecules 2022, 27, 8786. [Google Scholar] [CrossRef] [PubMed]
- Repert, S.; Matthes, S.; Rozhon, W. Quantification of Arbutin in Cosmetics, Drugs and Food Supplements by Hydrophilic-Interaction Chromatography. Molecules 2022, 27, 5673. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.X.; Wang, S.; Wei, L.; Cui, Y.Y.; Chen, Y.H. Proanthocyanidins: Components, Pharmacokinetics and Biomedical Properties. Am. J. Chin. Med. 2020, 48, 813–869. [Google Scholar] [CrossRef]
- Nam, J.W.; Phansalkar, R.S.; Lankin, D.C.; Bisson, J.; McAlpine, J.B.; Leme, A.A.; Vidal, C.M.P.; Ramirez, B.; Niemitz, M.; Bedran-Russo, A.; et al. Subtle Chemical Shifts Explain the NMR Fingerprints of Oligomeric Proanthocyanidins with High Dentin Biomodification Potency. J. Org. Chem. 2015, 80, 7495–7507. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Pang, Y.; Dixon, R.A. The Mysteries of Proanthocyanidin Transport and Polymerization. Plant Physiol. 2010, 153, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Urpi-Sarda, M.; Sánchez-Patán, F.; Llorach, R.; Garrido, I.; Gómez-Cordovés, C.; Andres-Lacueva, C.; Bartolomé, B. Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites. Food Funct. 2010, 1, 233. [Google Scholar] [CrossRef]
- Spranger, I.; Sun, B.; Mateus, A.M.; de Freitas, V.; Ricardo-da-Silva, J.M. Chemical characterization and antioxidant activities of oligomeric and polymeric procyanidin fractions from grape seeds. Food Chem. 2008, 108, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, X.; Hu, X.; Chen, F.; Ma, C. Health benefits of proanthocyanidins linking with gastrointestinal modulation: An updated review. Food Chem. 2023, 404, 134596. [Google Scholar] [CrossRef]
- Andersone, A.; Janceva, S.; Lauberte, L.; Ramata-Stunda, A.; Nikolajeva, V.; Zaharova, N.; Rieksts, G.; Telysheva, G. Anti-Inflammatory, Anti-Bacterial, and Anti-Fungal Activity of Oligomeric Proanthocyanidins and Extracts Obtained from Lignocellulosic Agricultural Waste. Molecules 2023, 28, 863. [Google Scholar] [CrossRef]
- Ulrey, R.K.; Barksdale, S.M.; Zhou, W.; van Hoek, M.L. Cranberry proanthocyanidins have anti-biofilm properties against Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2014, 14, 499. [Google Scholar]
- Fenech, M.; Amaya, I.; Valpuesta, V.; Botella, M.A. Vitamin C Content in Fruits: Biosynthesis and Regulation. Front. Plant Sci. 2019, 9, 2006. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on Dietary Reference Values for Vitamin C. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/3418 (accessed on 21 July 2024).
- Lykkesfeldt, J.; Tveden-Nyborg, P. The Pharmacokinetics of Vitamin C. Nutrients 2019, 11, 2412. [Google Scholar] [CrossRef]
- Xu, C.; Dong, N.; Chen, K.; Yang, X.; Zeng, P.; Hou, C.; Chan, E.W.C.; Yao, X.; Chen, S. Bactericidal, anti-biofilm, and anti-virulence activity of vitamin C against carbapenem-resistant hypervirulent Klebsiella pneumoniae. iScience 2022, 25, 103894. [Google Scholar] [CrossRef]
- Kalinowska, M.; Gołębiewska, E.; Świderski, G.; Męczyńska-Wielgosz, S.; Lewandowska, H.; Pietryczuk, A.; Cudowski, A.; Astel, A.; Świsłocka, R.; Samsonowicz, M.; et al. Plant-Derived and Dietary Hydroxybenzoic Acids—A Comprehensive Study of Structural, Anti-/Pro-Oxidant, Lipophilic, Antimicrobial, and Cytotoxic Activity in MDA-MB-231 and MCF-7 Cell Lines. Nutrients 2021, 13, 3107. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M.; Lorenzo, A.; Nabavi, S.; Talas, Z.; Nabavi, S. Polyphenols: Well Beyond the Antioxidant Capacity: Gallic Acid and Related Compounds as Neuroprotective Agents: You are What You Eat! Curr. Pharm. Biotechnol. 2014, 15, 362–372. [Google Scholar] [CrossRef]
- Tahmasebi, H.; Dehbashi, S.; Nasaj, M.; Arabestani, M.R. Molecular epidemiology and collaboration of siderophore-based iron acquisition with surface adhesion in hypervirulent Pseudomonas aeruginosa isolates from wound infections. Sci. Rep. 2022, 12, 7791. [Google Scholar] [CrossRef]
- Park, S.G.; Li, M.X.; Cho, W.K.; Joung, Y.K.; Huh, K.M. Thermosensitive gallic acid-conjugated hexanoyl glycol chitosan as a novel wound healing biomaterial. Carbohydr. Polym. 2021, 260, 117808. [Google Scholar] [CrossRef]
- Alshahrani, S.M. Development and optimization of oral nanoemulsion of rutin for enhancing its dissolution rate, permeability, and oral bioavailability. Pharm. Dev. Technol. 2022, 27, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, J.H.; Kim, Y.G.; Tan, Y.; Lee, J. Hydroquinones Inhibit Biofilm Formation and Virulence Factor Production in Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 10683. [Google Scholar] [CrossRef] [PubMed]
- Vostalova, J.; Vidlar, A.; Simanek, V.; Galandakova, A.; Kosina, P.; Vacek, J.; Vrbkova, J.; Zimmermann, B.F.; Ulrichova, J.; Student, V. Are High Proanthocyanidins Key to Cranberry Efficacy in the Prevention of Recurrent Urinary Tract Infection? Phytother. Res. 2015, 29, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Jeyanthi, V.; Velusamy, P.; Kumar, G.V.; Kiruba, K. Effect of naturally isolated hydroquinone in disturbing the cell membrane integrity of Pseudomonas aeruginosa MTCC 741 and Staphylococcus aureus MTCC 740. Heliyon 2021, 7, e07021. [Google Scholar] [CrossRef]
- Takahashi, S.; Hamasuna, R.; Yasuda, M.; Arakawa, S.; Tanaka, K.; Ishikawa, K.; Kiyota, H.; Hayami, H.; Yamamoto, S.; Kubo, T.; et al. A Randomized Clinical Trial to Evaluate the Preventive Effect of Cranberry Juice (UR65) for Patients with Recurrent Urinary Tract Infection. J. Infect. Chemother. 2013, 19, 112–117. [Google Scholar] [CrossRef]
- Skov, M.; Pressler, T.; Lykkesfeldt, J.; Poulsen, H.E.; Jensen, P.Ø.; Johansen, H.K.; Qvist, T.; Kræmer, D.; Høiby, N.; Ciofu, O. The effect of short-term, high-dose oral N-acetylcysteine treatment on oxidative stress markers in cystic fibrosis patients with chronic P. aeruginosa infection—A pilot study. J. Cyst. Fibros. 2015, 14, 211–218. [Google Scholar] [CrossRef]
- Yingcharoenthana, S.; Ampornaramveth, R.; Subbalekha, K.; Sinpitaksakul, P.; Kamolratanakul, P. A split-mouth randomized clinical trial to evaluate the effect of local and systemic administration of vitamin C on extraction wound healing. J. Oral Sci. 2021, 63, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.Y.; Yuan, M.; Cui, Z.Q.; Wu, Z.M.; Yu, Z.J.; Song, K.; Tang, B.; Fu, B.-D. Rutin inhibits quorum sensing, biofilm formation and virulence genes in avian pathogenic Escherichia coli. Microb. Pathog. 2018, 119, 54–59. [Google Scholar] [CrossRef]
- Kerdar, T.; Rabienejad, N.; Alikhani, Y.; Moradkhani, S.; Dastan, D. Clinical, in vitro and phytochemical, studies of Scrophularia striata mouthwash on chronic periodontitis disease. J. Ethnopharmacol. 2019, 239, 111872. [Google Scholar] [CrossRef] [PubMed]
- Katske, F.; A Shoskes, D.; Sender, M.; Poliakin, R.; Gagliano, K.; Rajfer, J. Treatment of Interstitial Cystitis with a Quercetin Supplement. Tech. Urol. 2001, 7, 44–46. [Google Scholar] [PubMed]
- Heinz, S.A.; Henson, D.A.; Austin, M.D.; Jin, F.; Nieman, D.C. Quercetin supplementation and upper respiratory tract infection: A randomized community clinical trial. Pharmacol. Res. 2010, 62, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lee, U.; Cho, H.R.; Lee, K.B.; Shin, Y.J.; Bae, M.J.; Park, K.Y. Effects of Pear Extracts on Microbiome and Immunocytokines to Alleviate Air Pollution-Related Respiratory Hypersensitivity. J. Med. Food. 2023, 26, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Aschauer, S.; Gouya, G.; Klickovic, U.; Storka, A.; Weisshaar, S.; Vollbracht, C.; Krick, B.; Weiss, G.; Wolzt, M. Effect of systemic high dose vitamin C therapy on forearm blood flow reactivity during endotoxemia in healthy human subjects. Vascul. Pharmacol. 2014, 61, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Garaiova, I.; Paduchová, Z.; Nagyová, Z.; Wang, D.; Michael, D.R.; Plummer, S.F.; Marchesi, J.; Ďuračková, Z.; Muchová, J. Probiotics with vitamin C for the prevention of upper respiratory tract symptoms in children aged 3–10 years: Randomised controlled trial. Benef. Microbes 2021, 12, 431–440. [Google Scholar] [CrossRef]
- Askari, S.F.; Jahromi, B.N.; Dehghanian, A.; Zarei, A.; Tansaz, M.; Badr, P.; Azadi, A.; Mohagheghzadeh, A. Effect of a novel herbal vaginal suppository containing myrtle and oak gall in the treatment of vaginitis: A randomized clinical trial. Daru 2020, 28, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Anaya-Loyola, M.A.; García-Marín, G.; García-Gutiérrez, D.G.; Castaño-Tostado, E.; Reynoso-Camacho, R.; López-Ramos, J.E.; Enciso-Moreno, J.A.; Pérez-Ramírez, I.F. A mango (Mangifera indica L.) juice by-product reduces gastrointestinal and upper respiratory tract infection symptoms in children. Food Res. Int. 2020, 136, 109492. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytosubstance | Bioavailability [%] | References | |
|---|---|---|---|
| QCT (Quercetin glucoside) | Oral Intake | Skin Penetration | |
| 6.7 (3–17) | 17 | [23] | |
| AP | 30 | - | [28] |
| ARB (Hydroquinone) | - | 45 | [18] |
| PAC ((-)epicatechine) | 5–10 | 35 | [29] |
| VC | 70–90 | 95 | [30] |
| GA | 70 | 11 | [31] |
| RT | 20 | 60 | [32,33] |
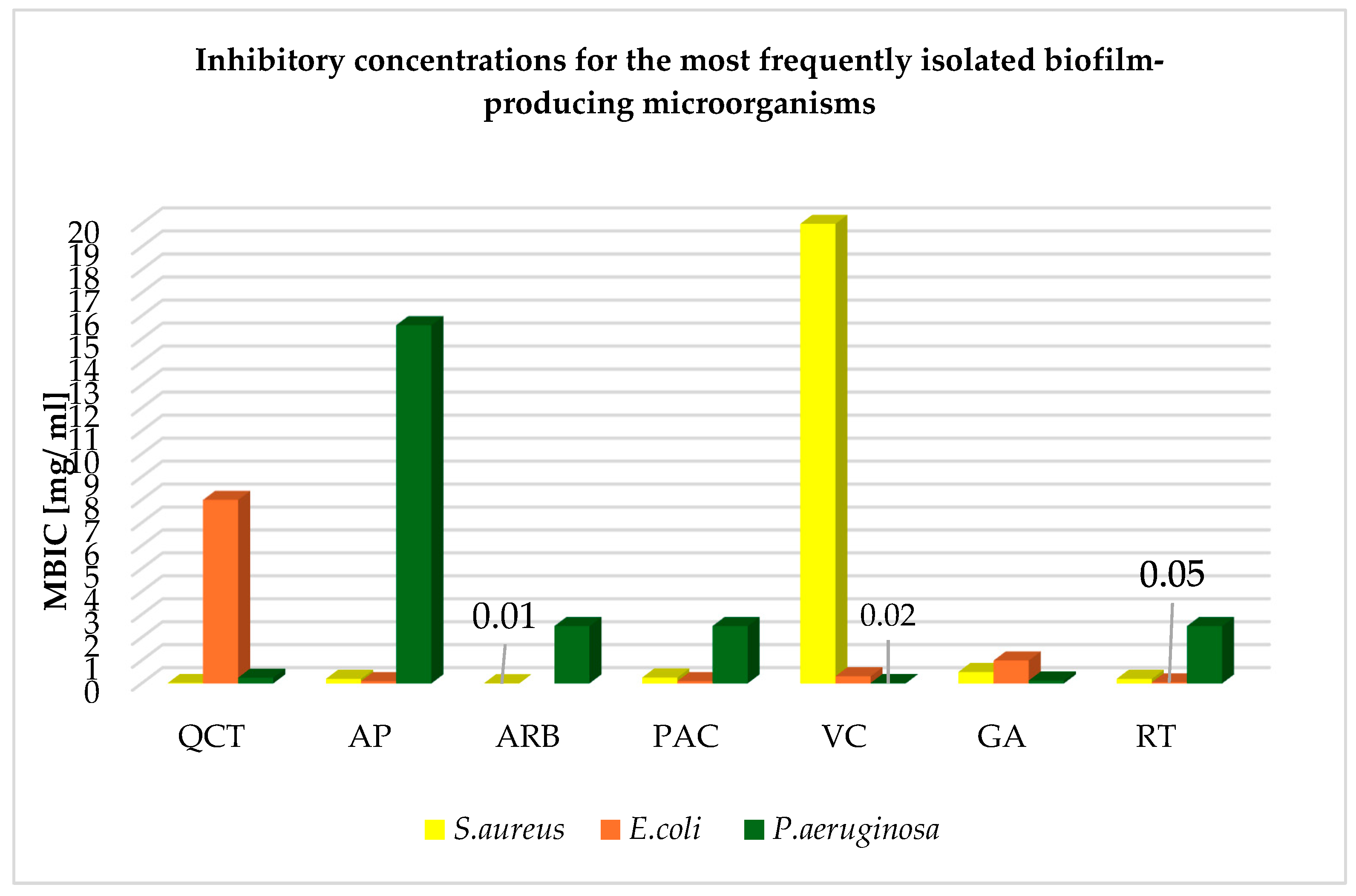
| Phytosubstance | Bacteria Species (Specificity) | Form | Number of Strains | Inhibitory Concentration [mg/mL] | References | |
|---|---|---|---|---|---|---|
| MIC | MBIC | |||||
| Quercetin | S. aureus (MRSA) | In silico model | ND | 0.025 | 0.02 | [24] |
| S. aureus (mixed biofilm) | EtOH solution | ND | 0.064 | 0.25 | [34] | |
| E. coli (ATCC) | EtOH solution | ND | 1.0 | 8.0 | [35] | |
| P. aeruginosa (mixed biofilm) | EtOH solution | ND | 0.128 | 0.25 | [34] | |
| Apigenin | S. aureus (MRSA) | EtOH solution Hydnum repandum extract | ND | 3.12 | 9.36 | [36] |
| Apigenin-7-O-Glucoside | ND | 0.28 | 0.20 | [37] | ||
| E. coli (ATCC) | EtOH solution Hydnum repandum extract | ND | 1.56 | 25 | [36] | |
| Apigenin-7-O-Glucoside | ND | 0.14 | 0.10 | [37] | ||
| P. aeruginosa (ATCC) | EtOH solution Hydnum repandum extract | ND | 3.12 | 15.60 | [36] | |
| Arbutin | S. aureus (MSSA) | DMSO solution | 6538 | 0.4 | 0.01 | [38] |
| E. coli (ESBL) | Itxasol© | ND | 0.256 | - | [38] | |
| P. aeruginosa (clinical strains) | Solution | ND | 1.25 | 2.50 | [39] | |
| Proanthocyanidins | S. aureus (MRSA) | Root extract Krameria lappacea | 10 | 0.064 | 0.256 | [40] |
| E. coli (tetracycline-resistant) | Cranberry (Vaccinium macrocarpon) extract | 21 | 0.1 | 0.1 | [41] | |
| P. aeruginosa (tetracycline-resistant) | Cranberry (Vaccinium macrocarpon) extract | 21 | 0.05 | 0.1 | [41] | |
| Vitamin C | S. aureus (ATCC) | H2O solution | ND | 0.15 | 20 | [42] |
| E. coli (MDR) | H2O solution | 85 | 1.25 | 0.312 | [43] | |
| P. aeruginosa (ATCC) | H2O solution | 50 | 0.625 | 0.156 | [42] | |
| Gallic acid | S. aureus (mixed biofilm) | EtOH solution | ND | 2.5 | 0.5 | [34] |
| E. coli (ATCC) | H2O solution | ND | 1.71 | 1.0 | [37] | |
| P. aeruginosa (ATCC) | Root extract Pelargonium hortorum | 10 | 0.512 | 0.128 | [44] | |
| P. aeruginosa (mixed biofilm) | EtOH solution | ND | 2.5 | 5 | [34] | |
| Rutin | S. aureus (ATCC) | Solution | 22 | 1.0 | 0.2 | [45] |
| E. coli O157:H7 (ATCC) | MetOH extract | ND | 0.03 | 0.05 | [46] | |
| P. aeruginosa (ESBL, clinical isolate) | MetOH extract Nepeta curviflora | ND | 10 | 2.5 | [47] | |
| Phytosubstance | Disease | Study Design | Dose (mg per Day) | Duration of Treatment (Weeks) | Patients (Age Mean) | Result | References |
|---|---|---|---|---|---|---|---|
| QCT | UTI complicated by Escherichia coli | CT | 500 | 4 | 20 (53) | Improved general condition and symptom score; no side effects | [82] |
| URTI | Randomized community CT | 500 or 1000 | 12 | 1002 (51) | Increased efficiency. Reduced URTI severity; reduced number of sick days | [83] | |
| ARB | UTI complicated by E. coli | RCT | 420 | One-time intake | 4 (ND) | Accumulation of hydroquinone in bacterial cells | [76] |
| Respiratory hypersensitivity | Randomized double-blind clinical studies | 5.01 | 4 | 20 (ND) | Growth of physiological bacterial microflora stimulation; activity of pro-inflammatory cytokines reduction | [84] | |
| PAC | UTI complicated by E. coli | RCT | 40 | 24 | 118 (50) | Reduced UTI recurrences | [77] |
| UTI complicated by E. coli | RCT | 500 | 24 | 182 (ND) | Reduced recurrences and the first episode of UTI occurrence | [75] | |
| VC | Acute inflammation induced by E. coli LPS | Balanced, placebo-controlled cross-over study | 320 or 480 per kg | One-time intake | 36 (ND) | Abolished effects of LPS secreted by E. coli (restored response to Ach, dilated blood vessels) | [85] |
| CF complicated by Pseudomonas aeruginosa | RCT | 2400 NACs which increase VC content | 4 | 21(ND) | Improved lung function | [78] | |
| URTI | RCT | 50 together with probiotic | 24 | 171 (6) | Reduced severity of symptoms | [86] | |
| Wound healing after dental extraction | RCT | 600 mg one-time intake | 3 | 30 (ND) | Improved soft tissue healing | [79] | |
| GA | Vaginitis | RCT | 0.276 | 1 | 120 (36) | Therapeutic effect on mixed vaginitis; reduced pain; eliminated symptoms | [87] |
| URTI and digestive system | RCT | 7.8 | 8 | ND (7) | Reduced symptoms’ frequency; improved blood test results | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fydrych, D.; Jeziurska, J.; Wełna, J.; Kwiecińska-Piróg, J. Potential Use of Selected Natural Compounds with Anti-Biofilm Activity. Int. J. Mol. Sci. 2025, 26, 607. https://doi.org/10.3390/ijms26020607
Fydrych D, Jeziurska J, Wełna J, Kwiecińska-Piróg J. Potential Use of Selected Natural Compounds with Anti-Biofilm Activity. International Journal of Molecular Sciences. 2025; 26(2):607. https://doi.org/10.3390/ijms26020607
Chicago/Turabian StyleFydrych, Dagmara, Jagoda Jeziurska, Jana Wełna, and Joanna Kwiecińska-Piróg. 2025. "Potential Use of Selected Natural Compounds with Anti-Biofilm Activity" International Journal of Molecular Sciences 26, no. 2: 607. https://doi.org/10.3390/ijms26020607
APA StyleFydrych, D., Jeziurska, J., Wełna, J., & Kwiecińska-Piróg, J. (2025). Potential Use of Selected Natural Compounds with Anti-Biofilm Activity. International Journal of Molecular Sciences, 26(2), 607. https://doi.org/10.3390/ijms26020607