

The Role of Long Non-Coding RNA in Rheumatoid Arthritis

Abstract
1. Introduction
2. Long Non-Coding RNA in the Pathogenesis of Rheumatoid Arthritis
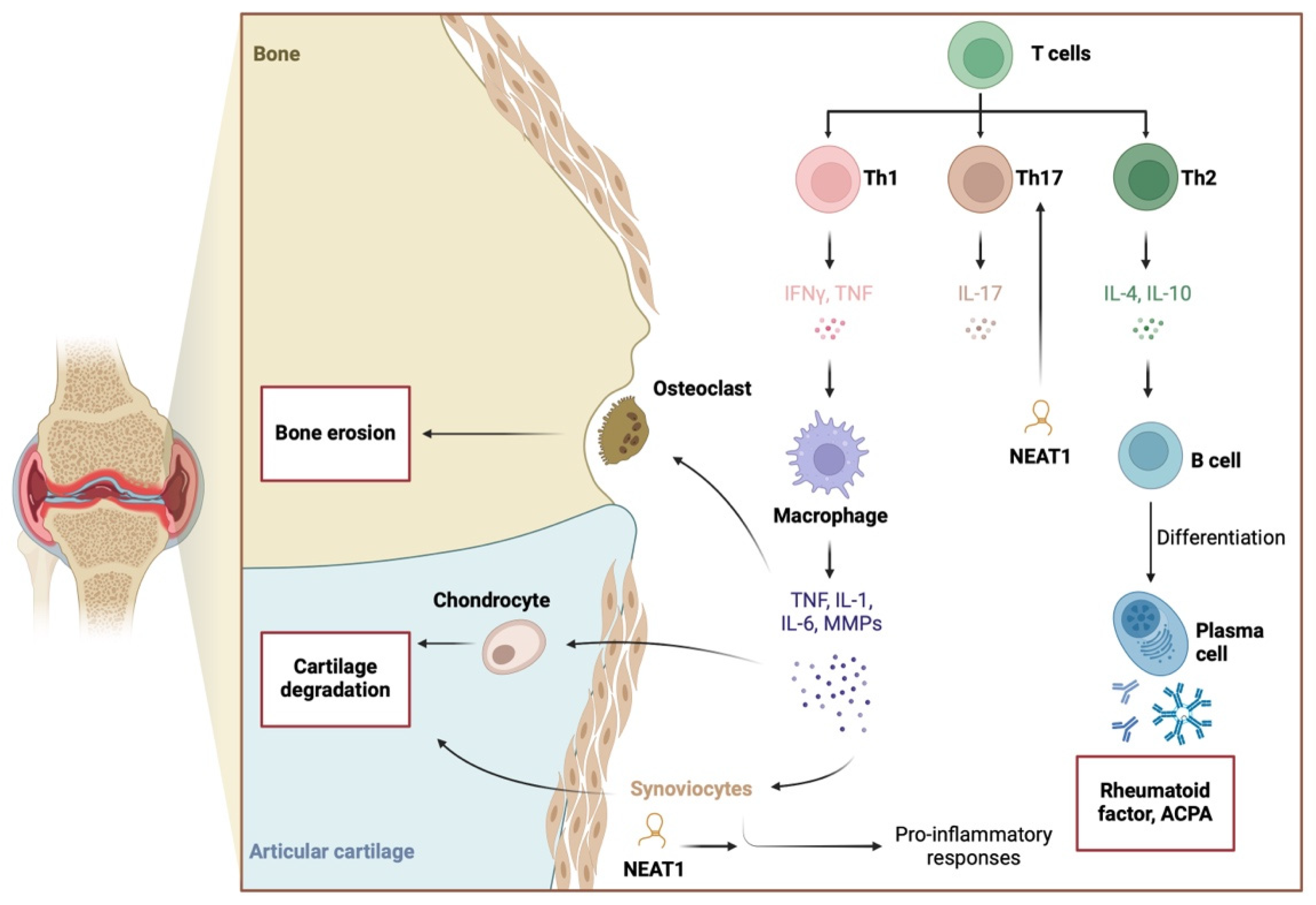
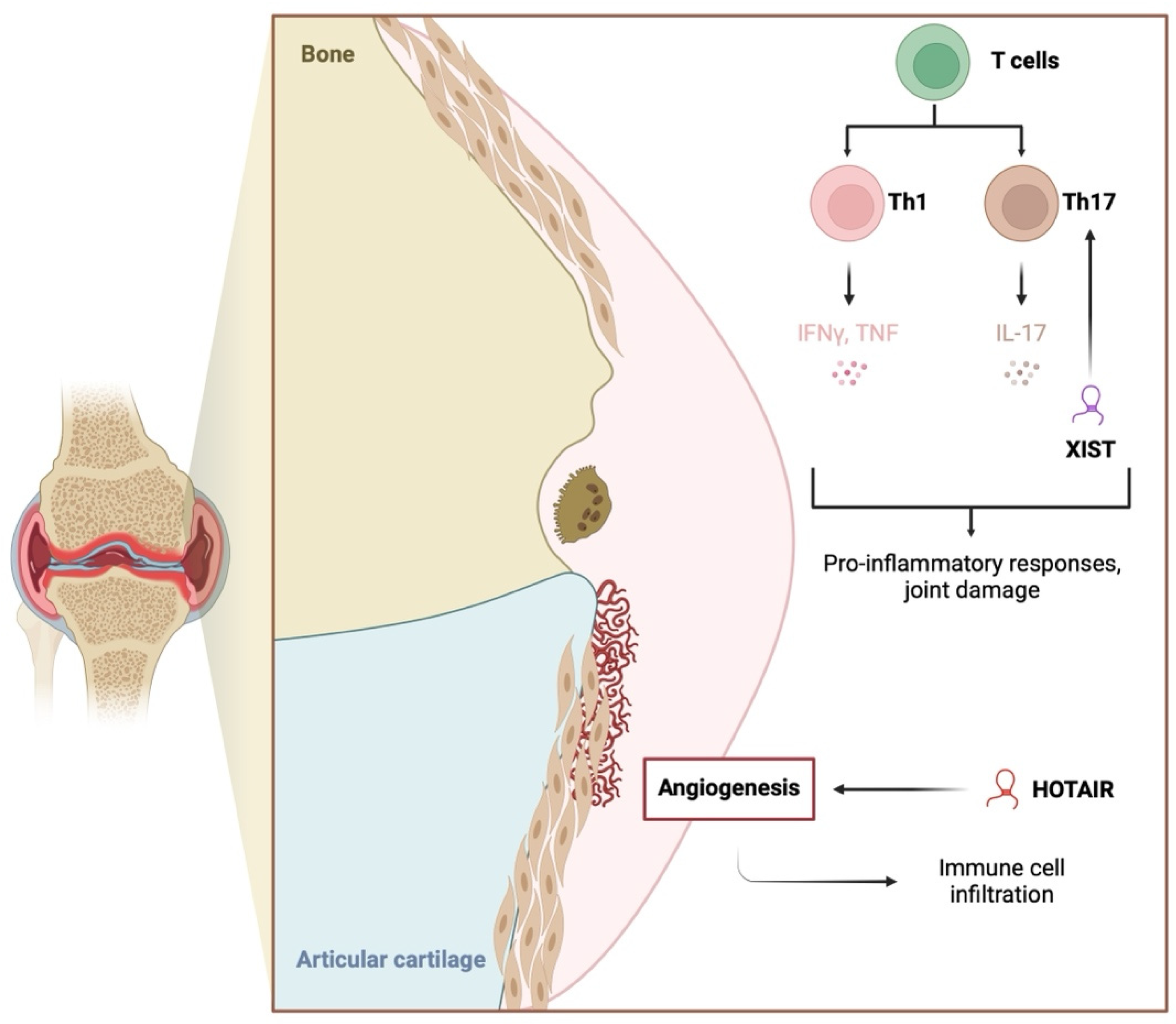
2.1. Inflammation
2.2. RA-FLS Functionality
3. Long Non-Coding RNAs as Diagnostic Biomarkers in Rheumatoid Arthritis
4. Targeting Long Non-Coding RNA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Matteo, A.; Bathon, J.M.; Emery, P. Rheumatoid arthritis. Lancet 2023, 402, 2019–2033. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, B.; Mo, L.; Chen, H.; Zhao, Y.; Tan, T.; Chen, L.; Li, Y.; Yao, P.; Tang, Y. Associations between biological ageing and the risk of, genetic susceptibility to, and life expectancy associated with rheumatoid arthritis: A secondary analysis of two observational studies. Lancet Healthy Longev. 2024, 5, e45–e55. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, J.; Hrkac, S.; Tecer, J.; Golob, M.; Ljilja Posavec, A.; Kolar Mitrovic, H.; Grgurevic, L. Pathogenesis of Extraarticular Manifestations in Rheumatoid Arthritis-A Comprehensive Review. Biomedicines 2023, 11, 1262. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Wang, Y.Y.; Li, Z.Y.; He, P.J.; Liu, S.; Li, Q.S. The prevalence and risk factors of rheumatoid arthritis-associated interstitial lung disease: A systematic review and meta-analysis. Ann. Med. 2024, 56, 2332406. [Google Scholar] [CrossRef]
- Krajewska-Wlodarczyk, M.; Szelag, M.; Batko, B.; Zuber, Z.; Orleanski, M.; Podwojcic, K.; Sowinski, J.; Jopek, J.; Swiderek, M.; Maluchnik, M.; et al. Rheumatoid arthritis epidemiology: A nationwide study in Poland. Rheumatol. Int. 2024, 44, 1155–1163. [Google Scholar] [CrossRef]
- Almutairi, K.; Nossent, J.; Preen, D.; Keen, H.; Inderjeeth, C. The global prevalence of rheumatoid arthritis: A meta-analysis based on a systematic review. Rheumatol. Int. 2021, 41, 863–877. [Google Scholar] [CrossRef]
- Chiu, Y.M.; Lu, Y.P.; Lan, J.L.; Chen, D.Y.; Wang, J.D. Lifetime Risks, Life Expectancy, and Health Care Expenditures for Rheumatoid Arthritis: A Nationwide Cohort Followed Up From 2003 to 2016. Arthritis Rheumatol. 2021, 73, 750–758. [Google Scholar] [CrossRef]
- Frazzei, G.; Musters, A.; de Vries, N.; Tas, S.W.; van Vollenhoven, R.F. Prevention of rheumatoid arthritis: A systematic literature review of preventive strategies in at-risk individuals. Autoimmun. Rev. 2023, 22, 103217. [Google Scholar] [CrossRef]
- Smolen, J.S.; Landewe, R.B.M.; Bergstra, S.A.; Kerschbaumer, A.; Sepriano, A.; Aletaha, D.; Caporali, R.; Edwards, C.J.; Hyrich, K.L.; Pope, J.E.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2022 update. Ann. Rheum. Dis. 2023, 82, 3–18. [Google Scholar] [CrossRef]
- Paradowska-Gorycka, A.; Wajda, A.; Romanowska-Prochnicka, K.; Walczuk, E.; Kuca-Warnawin, E.; Kmiolek, T.; Stypinska, B.; Rzeszotarska, E.; Majewski, D.; Jagodzinski, P.P.; et al. Th17/Treg-Related Transcriptional Factor Expression and Cytokine Profile in Patients With Rheumatoid Arthritis. Front. Immunol. 2020, 11, 572858. [Google Scholar] [CrossRef]
- Ye, Z.; Chen, L.; Fang, Y.; Zhao, L. Blood MALT1, Th1, and Th17 cells are dysregulated, inter-correlated, and correlated with disease activity in rheumatoid arthritis patients; meanwhile, MALT1 decline during therapy relates to treatment outcome. J. Clin. Lab. Anal. 2022, 36, e24112. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Fu, D.; Fang, X.; Xie, Y.; Zheng, X.; Fan, W.; Hu, F.; Li, Z. Casein Kinase II exacerbates rheumatoid arthritis via promoting Th1 and Th17 cell inflammatory responses. Expert Opin. Ther. Targets 2021, 25, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Masoumi, M.; Alesaeidi, S.; Khorramdelazad, H.; Behzadi, M.; Baharlou, R.; Alizadeh-Fanalou, S.; Karami, J. Role of T Cells in the Pathogenesis of Rheumatoid Arthritis: Focus on Immunometabolism Dysfunctions. Inflammation 2023, 46, 88–102. [Google Scholar] [CrossRef] [PubMed]
- The Nobel Prize in Physiology or Medicine. Available online: https://www.nobelprize.org/prizes/medicine/ (accessed on 20 October 2024).
- Calin, G.A.; Hube, F.; Ladomery, M.R.; Delihas, N.; Ferracin, M.; Poliseno, L.; Agnelli, L.; Alahari, S.K.; Yu, A.M.; Zhong, X.B. The 2024 Nobel Prize in Physiology or Medicine: microRNA Takes Center Stage. Noncoding RNA 2024, 10, 62. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, R.; Zhou, W.; Cai, X.; Ye, Z. LncRNA NEAT1 regulates the proliferation and production of the inflammatory cytokines in rheumatoid arthritis fibroblast-like synoviocytes by targeting miR-204-5p. Hum. Cell 2021, 34, 372–382. [Google Scholar] [CrossRef]
- Wang, J.; Yan, S.; Yang, J.; Lu, H.; Xu, D.; Wang, Z. Non-coding RNAs in Rheumatoid Arthritis: From Bench to Bedside. Front. Immunol. 2019, 10, 3129. [Google Scholar] [CrossRef]
- Dolcino, M.; Pelosi, A.; Fiore, P.F.; Patuzzo, G.; Tinazzi, E.; Lunardi, C.; Puccetti, A. Long Non-Coding RNAs Play a Role in the Pathogenesis of Psoriatic Arthritis by Regulating MicroRNAs and Genes Involved in Inflammation and Metabolic Syndrome. Front. Immunol. 2018, 9, 1533. [Google Scholar] [CrossRef]
- Zhou, Y.; Gu, L.N.; Zhang, J.; Pan, J.; Zhang, J.M.; Zhao, D.Y.; Liu, F. LncRNA-AK149641 regulates the secretion of tumor necrosis factor-α in P815 mast cells by targeting the nuclear factor-kappa B signaling pathway. Sci. Rep. 2020, 10, 16655. [Google Scholar] [CrossRef]
- Tang, J.; Yi, S.; Liu, Y. Long non-coding RNA PVT1 can regulate the proliferation and inflammatory responses of rheumatoid arthritis fibroblast-like synoviocytes by targeting microRNA-145-5p. Hum. Cell 2020, 33, 1081–1090. [Google Scholar] [CrossRef]
- Ye, Y.; Gao, X.; Yang, N. LncRNA ZFAS1 promotes cell migration and invasion of fibroblast-like synoviocytes by suppression of miR-27a in rheumatoid arthritis. Hum. Cell 2018, 31, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Guo, X.H.; Mo, B.Y.; Wang, M.L.; Luo, X.Q.; Chen, Y.X.; Liu, F.; Olsen, N.; Pan, Y.F.; Zheng, S.G. LncRNA PICSAR promotes cell proliferation, migration and invasion of fibroblast-like synoviocytes by sponging miRNA-4701-5p in rheumatoid arthritis. EBioMedicine 2019, 50, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Mo, B.Y.; Guo, X.H.; Yang, M.R.; Liu, F.; Bi, X.; Liu, Y.; Fang, L.K.; Luo, X.Q.; Wang, J.; Bellanti, J.A.; et al. Long Non-Coding RNA GAPLINC Promotes Tumor-Like Biologic Behaviors of Fibroblast-Like Synoviocytes as MicroRNA Sponging in Rheumatoid Arthritis Patients. Front. Immunol. 2018, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Chini, A.; Guha, P.; Malladi, V.S.; Guo, Z.; Mandal, S.S. Novel long non-coding RNAs associated with inflammation and macrophage activation in human. Sci. Rep. 2023, 13, 4036. [Google Scholar] [CrossRef]
- Guo, T.; Xing, Y.; Chen, Z.; Zhu, H.; Yang, L.; Xiao, Y.; Xu, J. Long Non-Coding RNA NEAT1 Knockdown Alleviates Rheumatoid Arthritis by Reducing IL-18 through p300/CBP Repression. Inflammation 2022, 45, 100–115. [Google Scholar] [CrossRef]
- Yang, J.; Wang, S.; Liu, L.; Wang, J.; Shao, Y. Long non-coding RNA NEAT1 and its targets (microRNA-21 and microRNA-125a) in rheumatoid arthritis: Altered expression and potential to monitor disease activity and treatment outcome. J. Clin. Lab. Anal. 2021, 35, e24076. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, X.; Hu, Q.; Chen, X.; Chen, Y.; Huang, L. Galectin-3 activates TLR4/NF-kappaB signaling to promote lung adenocarcinoma cell proliferation through activating lncRNA-NEAT1 expression. BMC Cancer 2018, 18, 580. [Google Scholar] [CrossRef]
- Wang, Z.; Li, K.; Huang, W. Long non-coding RNA NEAT1-centric gene regulation. Cell. Mol. Life Sci. 2020, 77, 3769–3779. [Google Scholar] [CrossRef]
- Vasilev, G.; Manolova, I.; Ivanova, M.; Stanilov, I.; Miteva, L.; Stanilova, S. The role of IL-18 in addition to Th17 cytokines in rheumatoid arthritis development and treatment in women. Sci. Rep. 2021, 11, 15391. [Google Scholar] [CrossRef]
- Landy, E.; Carol, H.; Ring, A.; Canna, S. Biological and clinical roles of IL-18 in inflammatory diseases. Nat. Rev. Rheumatol. 2024, 20, 33–47. [Google Scholar] [CrossRef]
- Shui, X.; Chen, S.; Lin, J.; Kong, J.; Zhou, C.; Wu, J. Knockdown of lncRNA NEAT1 inhibits Th17/CD4(+) T cell differentiation through reducing the STAT3 protein level. J. Cell. Physiol. 2019, 234, 22477–22484. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Su, R.; Guo, Q.; Su, R.; Gao, C.; Li, X.; Wang, C. Differential immunological profiles in seronegative versus seropositive rheumatoid arthritis: Th17/Treg dysregulation and IL-4. Front. Immunol. 2024, 15, 1447213. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, J.; Zhang, K.; Ma, B.; Li, X.; Wei, R.; Nian, H. LncRNA Neat1 targets NonO and miR-128-3p to promote antigen-specific Th17 cell responses and autoimmune inflammation. Cell Death Dis. 2023, 14, 610. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Y.; Tan, W.; Gao, H.; Xiao, S.; Gao, J.; Zhu, Z. Silencing of long non-coding RNA NEAT1 improves Treg/Th17 imbalance in preeclampsia via the miR-485-5p/AIM2 axis. Bioengineered 2021, 12, 8768–8777. [Google Scholar] [CrossRef]
- Safari, F.; Farajnia, S.; Arya, M.; Zarredar, H.; Nasrolahi, A. CRISPR and personalized Treg therapy: New insights into the treatment of rheumatoid arthritis. Immunopharmacol. Immunotoxicol. 2018, 40, 201–211. [Google Scholar] [CrossRef]
- Karamali, N.; Mahmoudi, Z.; Roghani, S.A.; Assar, S.; Pournazari, M.; Soufivand, P.; Karaji, A.G.; Rezaiemanesh, A. Overexpression of Synoviolin and miR-125a-5p, miR-19b-3p in peripheral blood of rheumatoid arthritis patients after treatment with conventional DMARDs and methylprednisolone. Clin. Rheumatol. 2024, 43, 147–157. [Google Scholar] [CrossRef]
- Yagishita, N.; Aratani, S.; Leach, C.; Amano, T.; Yamano, Y.; Nakatani, K.; Nishioka, K.; Nakajima, T. RING-finger type E3 ubiquitin ligase inhibitors as novel candidates for the treatment of rheumatoid arthritis. Int. J. Mol. Med. 2012, 30, 1281–1286. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sato, Y.; Kobayashi, T.; Ito, E.; Soma, T.; Kimura, A.; Miyamoto, K.; Kobayashi, S.; Harato, K.; Matsumoto, M.; et al. Synoviolin is not a pathogenic factor for auto-inflammatory diseases. Biochem. Biophys. Res. Commun. 2021, 558, 183–188. [Google Scholar] [CrossRef]
- Toh, M.L.; Marotte, H.; Blond, J.L.; Jhumka, U.; Eljaafari, A.; Mougin, B.; Miossec, P. Overexpression of synoviolin in peripheral blood and synoviocytes from rheumatoid arthritis patients and continued elevation in nonresponders to infliximab treatment. Arthritis Rheum. 2006, 54, 2109–2118. [Google Scholar] [CrossRef]
- Li, W.; Cao, T.; Luo, C.; Cai, J.; Zhou, X.; Xiao, X.; Liu, S. Crosstalk between ER stress, NLRP3 inflammasome, and inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 6129–6140. [Google Scholar] [CrossRef]
- Zhang, P.; Cao, L.; Zhou, R.; Yang, X.; Wu, M. The lncRNA Neat1 promotes activation of inflammasomes in macrophages. Nat. Commun. 2019, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, J.; Xu, Q.; Wang, X.; Zhong, W.; Wu, F.; Lin, X.; Zhang, Q.; Ye, Q. Long non-coding RNA NEAT1 exacerbates NLRP3-mediated pyroptosis in allergic rhinitis through regulating the PTBP1/FOXP1 cascade. Int. Immunopharmacol. 2024, 137, 112337. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, Y.; Guo, L.; Yuan, S.; Sun, J.; Zhao, K.; Wang, J.; An, R. Exosomal Lnc NEAT1 from endothelial cells promote bone regeneration by regulating macrophage polarization via DDX3X/NLRP3 axis. J. Nanobiotechnol. 2023, 21, 98. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, C.; Gao, H.; Li, C.; Li, D.; Liu, P.; Huang, M.; Shen, X.; Liu, L. Therapeutic effect of Cryptotanshinone on experimental rheumatoid arthritis through downregulating p300 mediated-STAT3 acetylation. Biochem. Pharmacol. 2017, 138, 119–129. [Google Scholar] [CrossRef]
- Krosel, M.; Gabathuler, M.; Moser, L.; Maciukiewicz, M.; Zullig, T.; Seifritz, T.; Tomsic, M.; Distler, O.; Ospelt, C.; Klein, K. The histone acetyl transferases CBP and p300 regulate stress response pathways in synovial fibroblasts at transcriptional and functional levels. Sci. Rep. 2023, 13, 17112. [Google Scholar] [CrossRef]
- Seong, A.R.; Yoo, J.Y.; Choi, K.; Lee, M.H.; Lee, Y.H.; Lee, J.; Jun, W.; Kim, S.; Yoon, H.G. Delphinidin, a specific inhibitor of histone acetyltransferase, suppresses inflammatory signaling via prevention of NF-kappaB acetylation in fibroblast-like synoviocyte MH7A cells. Biochem. Biophys. Res. Commun. 2011, 410, 581–586. [Google Scholar] [CrossRef]
- Song, J.; Kim, D.; Han, J.; Kim, Y.; Lee, M.; Jin, E.J. PBMC and exosome-derived Hotair is a critical regulator and potent marker for rheumatoid arthritis. Clin. Exp. Med. 2015, 15, 121–126. [Google Scholar] [CrossRef]
- Shaker, O.G.; Mahmoud, R.H.; Abdelaleem, O.O.; Ahmed, T.I.; Fouad, N.A.; Hussein, H.A.; Nassr, M.H.; Zaki, O.M.; Abdelghaffar, N.K.; Hefzy, E.M. Expression Profile of Long Noncoding RNAs, lnc-Cox2, and HOTAIR in Rheumatoid Arthritis Patients. J. Interferon Cytokine Res. 2019, 39, 174–180. [Google Scholar] [CrossRef]
- Medhat, E.; Ayeldeen, G.; Hosni Ahmed, H.; Shaker, O.; Gheita, T.; Salama Ashour, S. HOTAIR and THRIL Long Non Coding RNAs and Their Target Genes in Rheumatoid Arthritis patients. Rep. Biochem. Mol. Biol. 2022, 10, 614–621. [Google Scholar] [CrossRef]
- Qiu, H.; Liu, M.; Shi, X.; Ma, M.; Zhang, J.; Liu, H. LncRNA HOTAIR inhibits the progression of fibroblast-like synoviocytes by sponging miRNA-106b-5p in rheumatoid arthritis. Autoimmunity 2022, 55, 567–576. [Google Scholar] [CrossRef]
- Elhai, M.; Micheroli, R.; Houtman, M.; Mirrahimi, M.; Moser, L.; Pauli, C.; Burki, K.; Laimbacher, A.; Kania, G.; Klein, K.; et al. The long non-coding RNA HOTAIR contributes to joint-specific gene expression in rheumatoid arthritis. Nat. Commun. 2023, 14, 8172. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, Y.; Huang, D.; Sun, Y. LncRNA HOTAIR regulates the PI3K/AKT pathway via the miR-126-3p/PIK3R2 axis to participate in synovial angiogenesis in rheumatoid arthritis. Immun. Inflamm. Dis. 2023, 11, e1064. [Google Scholar] [CrossRef] [PubMed]
- Chi, K.; Geng, X.; Liu, C.; Zhang, Y.; Cui, J.; Cai, G.; Chen, X.; Wang, F.; Hong, Q. LncRNA-HOTAIR promotes endothelial cell pyroptosis by regulating the miR-22/NLRP3 axis in hyperuricaemia. J. Cell. Mol. Med. 2021, 25, 8504–8521. [Google Scholar] [CrossRef]
- You, H.; Li, H.; Gou, W. lncRNA HOTAIR promotes ROS generation and NLRP3 inflammasome activation by inhibiting Nrf2 in diabetic retinopathy. Medicine 2023, 102, e35155. [Google Scholar] [CrossRef]
- Obaid, M.; Udden, S.M.N.; Deb, P.; Shihabeddin, N.; Zaki, M.H.; Mandal, S.S. LncRNA HOTAIR regulates lipopolysaccharide-induced cytokine expression and inflammatory response in macrophages. Sci. Rep. 2018, 8, 15670. [Google Scholar] [CrossRef]
- Kielbowski, K.; Stanska, W.; Bakinowska, E.; Rusinski, M.; Pawlik, A. The Role of Alarmins in the Pathogenesis of Rheumatoid Arthritis, Osteoarthritis, and Psoriasis. Curr. Issues Mol. Biol. 2024, 46, 3640–3675. [Google Scholar] [CrossRef]
- Shin, J.J.; Park, J.; Shin, H.S.; Arab, I.; Suk, K.; Lee, W.H. Roles of lncRNAs in NF-kappaB-Mediated Macrophage Inflammation and Their Implications in the Pathogenesis of Human Diseases. Int. J. Mol. Sci. 2024, 25, 2670. [Google Scholar] [CrossRef]
- Pang, J.L.; Wang, J.W.; Hu, P.Y.; Jiang, J.S.; Yu, C. HOTAIR alleviates ox-LDL-induced inflammatory response in Raw264.7 cells via inhibiting NF-kappaB pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6991–6998. [Google Scholar] [CrossRef]
- Wang, Y.; Yi, K.; Liu, X.; Tan, Y.; Jin, W.; Li, Y.; Zhou, J.; Wang, H.; Kang, C. HOTAIR Up-Regulation Activates NF-kappaB to Induce Immunoescape in Gliomas. Front. Immunol. 2021, 12, 785463. [Google Scholar] [CrossRef]
- Chen, R.; Chen, B.; Li, D.; Wang, Q.; Zhu, Y.; Li, M.; Wang, Y.; Fang, S.; Guo, L. HOTAIR contributes to chemoresistance by activating NF-kappaB signaling in small-cell lung cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 2997–3004. [Google Scholar]
- Wang, J.; Chen, J.; Jing, G.; Dong, D. LncRNA HOTAIR Promotes Proliferation of Malignant Melanoma Cells through NF-varkappaB Pathway. Iran. J. Public Health 2020, 49, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Song, J.; Feng, X.; Yang, H.; Zhong, W. LncRNA XIST is involved in rheumatoid arthritis fibroblast-like synoviocytes by sponging miR-126-3p via the NF-kappaB pathway. Autoimmunity 2021, 54, 326–335. [Google Scholar] [CrossRef]
- Wei, Y.; Dai, L.; Deng, Y.; Ye, Z. LncRNA XIST promotes adjuvant-induced arthritis by increasing the expression of YY1 via miR-34a-5p. Arch. Rheumatol. 2023, 38, 82–94. [Google Scholar] [CrossRef]
- Lin, J.; He, Y.; Chen, J.; Zeng, Z.; Yang, B.; Ou, Q. A critical role of transcription factor YY1 in rheumatoid arthritis by regulation of interleukin-6. J. Autoimmun. 2017, 77, 67–75. [Google Scholar] [CrossRef]
- Lin, J.; Tang, J.; Lin, J.; He, Y.; Yu, Z.; Jiang, R.; Yang, B.; Ou, Q. YY1 regulation by miR-124-3p promotes Th17 cell pathogenicity through interaction with T-bet in rheumatoid arthritis. JCI Insight 2021, 6, e149985. [Google Scholar] [CrossRef]
- Mu, N.; Gu, J.; Huang, T.; Zhang, C.; Shu, Z.; Li, M.; Hao, Q.; Li, W.; Zhang, W.; Zhao, J.; et al. A novel NF-kappaB/YY1/microRNA-10a regulatory circuit in fibroblast-like synoviocytes regulates inflammation in rheumatoid arthritis. Sci. Rep. 2016, 6, 20059. [Google Scholar] [CrossRef]
- Sun, L.; Hu, L.; Chen, P.; Li, Y.; Tu, J.; Chen, J. Long Non-Coding RNA AL928768.3 Promotes Rheumatoid Arthritis Fibroblast-Like Synoviocytes Proliferation, Invasion and Inflammation, While Inhibits Apoptosis Via Activating Lymphotoxin Beta Mediated NF-kappaB Signaling Pathway. Inflammation 2024, 47, 543–556. [Google Scholar] [CrossRef]
- Li, K.; Liu, W.; Zhao, X.; Lin, W.; Zhou, W.; Zhang, Q. LncRNA SNHG3 discriminates rheumatoid arthritis from healthy individuals and regulates inflammatory response and oxidative stress via modulating miR-128-3p. Mod. Rheumatol. 2024, 34, 1153–1161. [Google Scholar] [CrossRef]
- Elazazy, O.; Midan, H.M.; Shahin, R.K.; Elesawy, A.E.; Elballal, M.S.; Sallam, A.M.; Elbadry, A.M.M.; Elrebehy, M.A.; Bhnsawy, A.; Doghish, A.S. Long non-coding RNAs and rheumatoid arthritis: Pathogenesis and clinical implications. Pathol. Res. Pract. 2023, 246, 154512. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, M.; Hu, J. Non-coding RNAs involved in fibroblast-like synoviocyte functioning in arthritis rheumatoid: From pathogenesis to therapy. Cytokine 2024, 173, 156418. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Xu, S.; Xiao, Y.; Qiu, Q.; Shi, M.; Wang, J.; Liang, L.; Zhan, Z.; Yang, X.; Olsen, N.; et al. Long noncoding RNA LERFS negatively regulates rheumatoid synovial aggression and proliferation. J. Clin. Investig. 2018, 128, 4510–4524. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Shen, C.; Shen, T.; Wang, J.; Zhang, X.; Zhang, Q.; Sun, R.; Dai, L.; Xu, H. LncRNA. Ann. Transl. Med. 2021, 9, 1368. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Ouyang, Q.; Feng, Y.; Lu, X.; Han, Y.; Ren, H.; Huang, Q.; Zhao, J.; Xiao, C.; Yang, M. LncNFYB promotes the proliferation of rheumatoid arthritis fibroblast-like synoviocytes via LncNFYB/ANXA2/ERK1/2 axis. J. Biol. Chem. 2024, 300, 105591. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, F.F.; Xie, J. LncRNA linc00152/NF-κB feedback loop promotes fibroblast-like synovial cells inflammation in rheumatoid arthritis via regulating miR-103a/TAK1 axis and YY1 expression. Immun. Inflamm. Dis. 2021, 9, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Guo, P.; Chen, M.; Chen, D.; Cheng, Y.; He, L. FOXM1/LINC00152 feedback loop regulates proliferation and apoptosis in rheumatoid arthritis fibroblast-like synoviocytes via Wnt/β-catenin signaling pathway. BioSci. Rep. 2020, 40, BSR20191900. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tian, Q.; Tian, Y.; Zheng, Z. MicroRNA-122-3p plays as the target of long non-coding RNA LINC00665 in repressing the progress of arthritis. Bioengineered 2022, 13, 13328–13340. [Google Scholar] [CrossRef]
- Fu, Q.; Song, M.J.; Fang, J. LncRNA OSER1-AS1 regulates the inflammation and apoptosis of rheumatoid arthritis fibroblast like synoviocytes via regulating miR-1298-5p/E2F1 axis. Bioengineered 2022, 13, 4951–4963. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, L.; Yuan, X.; Xu, N.; Zhao, S.; Yang, L.; Zhang, N. LncRNA NEAT1 Targets Fibroblast-Like Synoviocytes in Rheumatoid Arthritis via the miR-410-3p/YY1 Axis. Front. Immunol. 2020, 11, 1975. [Google Scholar] [CrossRef]
- Xu, S.; Liu, D.; Kuang, Y.; Li, R.; Wang, J.; Shi, M.; Zou, Y.; Qiu, Q.; Liang, L.; Xiao, Y.; et al. Long Noncoding RNA HAFML Promotes Migration and Invasion of Rheumatoid Fibroblast-like Synoviocytes. J. Immunol. 2023, 210, 135–147. [Google Scholar] [CrossRef]
- Lao, M.X.; Xu, H.S. Involvement of long non-coding RNAs in the pathogenesis of rheumatoid arthritis. Chin. Med. J. 2020, 133, 941–950. [Google Scholar] [CrossRef]
- Wang, G.; Tang, L.; Zhang, X.; Li, Y. LncRNA DILC participates in rheumatoid arthritis by inducing apoptosis of fibroblast-like synoviocytes and down-regulating IL-6. BioSci. Rep. 2019, 39, BSR20182374. [Google Scholar] [CrossRef] [PubMed]
- Piao, X.; Zhou, J.; Hu, J. Role of RP11-83J16.1, a novel long non-coding RNA, in rheumatoid arthritis. Am. J. Transl. Res. 2020, 12, 1397–1414. [Google Scholar] [PubMed]
- Zou, Y.; Xu, H. Involvement of long noncoding RNAs in the pathogenesis of autoimmune diseases. J. Transl. Autoimmun. 2020, 3, 100044. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zheng, N.; Luo, D.; Lin, D.; Que, W.; Wang, H.; Huang, Q.; Yang, J.; Ye, J.; Chen, X. Long Non-Coding RNA NR-133666 Promotes the Proliferation and Migration of Fibroblast-Like Synoviocytes Through Regulating the miR-133c/MAPK1 Axis. Front. Pharmacol. 2022, 13, 887330. [Google Scholar] [CrossRef]
- Pan, F.; Zhu, L.; Lv, H.; Pei, C. Quercetin promotes the apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis by upregulating lncRNA MALAT1. Int. J. Mol. Med. 2016, 38, 1507–1514. [Google Scholar] [CrossRef]
- Zheng, J.; Zeng, P.; Zhang, H.; Zhou, Y.; Liao, J.; Zhu, W.; Jia, N.; Lin, L. Long noncoding RNA ZFAS1 silencing alleviates rheumatoid arthritis via blocking miR-296-5p-mediated down-regulation of MMP-15. Int. Immunopharmacol. 2021, 90, 107061. [Google Scholar] [CrossRef]
- Zhang, C.W.; Wu, X.; Liu, D.; Zhou, W.; Tan, W.; Fang, Y.X.; Zhang, Y.; Liu, Y.Q.; Li, G.Q. Long non-coding RNA PVT1 knockdown suppresses fibroblast-like synoviocyte inflammation and induces apoptosis in rheumatoid arthritis through demethylation of sirt6. J. Biol. Eng. 2019, 13, 60. [Google Scholar] [CrossRef]
- Garcia-Carbonell, R.; Divakaruni, A.S.; Lodi, A.; Vicente-Suarez, I.; Saha, A.; Cheroutre, H.; Boss, G.R.; Tiziani, S.; Murphy, A.N.; Guma, M. Critical Role of Glucose Metabolism in Rheumatoid Arthritis Fibroblast-like Synoviocytes. Arthritis Rheumatol. 2016, 68, 1614–1626. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, N.; Guo, X.Y.; Li, H.J.; Guo, Y.; Lu, L. Influences of the lncRNA TUG1-miRNA-34a-5p network on fibroblast-like synoviocytes (FLSs) dysfunction in rheumatoid arthritis through targeting the lactate dehydrogenase A (LDHA). J. Clin. Lab. Anal. 2021, 35, e23969. [Google Scholar] [CrossRef]
- Peng, H.; Ren, S.; Liu, Y.; Zhou, H.; Tang, X.; Yang, J.; Tian, J.; Xu, P.; Xu, H.; Wang, S. Elevated Expression of the Long Noncoding RNA IFNG-AS1 in the Peripheral Blood from Patients with Rheumatoid Arthritis. J. Immunol. Res. 2020, 2020, 6401978. [Google Scholar] [CrossRef]
- Moharamoghli, M.; Hassan-Zadeh, V.; Dolatshahi, E.; Alizadeh, Z.; Farazmand, A. The expression of GAS5, THRIL, and RMRP lncRNAs is increased in T cells of patients with rheumatoid arthritis. Clin. Rheumatol. 2019, 38, 3073–3080. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Gao, H.; Tang, J.; Jiang, R.; Xiao, L.; Sheng, H.; Lin, J. A novel diagnostic model based on lncRNA PTPRE expression, neutrophil count and red blood cell distribution width for diagnosis of seronegative rheumatoid arthritis. Clin. Exp. Med. 2024, 24, 86. [Google Scholar] [CrossRef] [PubMed]
- Selim, A.M.; Elsabagh, Y.A.; El-Sawalhi, M.M.; Ismail, N.A.; Senousy, M.A. Serum lncRNA ITGB2-AS1 and ICAM-1 as novel biomarkers for rheumatoid arthritis and osteoarthritis diagnosis. BMC Med. Genom. 2024, 17, 247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, W.; Wen, Q.W.; Wang, T.H.; Qin, W.; Huang, H.; Mo, Y.J.; Wu, X.D.; Cen, H. Associations of genetic polymorphisms within. Autoimmunity 2020, 53, 408–414. [Google Scholar] [CrossRef]
- Wu, H.; Chen, Q.; Wang, S.; Yang, C.; Xu, L.; Xiao, H.; Xie, T.; Pan, Q. Serum exosomes lncRNAs: TCONS_I2_00013502 and ENST00000363624 are new diagnostic markers for rheumatoid arthritis. Front. Immunol. 2024, 15, 1419683. [Google Scholar] [CrossRef]
- Zhao, X.; Lin, W.; Zhou, W. Clinical significance of long non-coding RNA NORAD in rheumatoid arthritis. Adv. Rheumatol. 2024, 64, 9. [Google Scholar] [CrossRef]
- Gong, X.; Fan, X.; Zhang, Z.; Cai, Q.; Guo, M.; Gao, C.; Xiao, L.; Yue, T. Circulating lnc-ITSN1-2 expression presents a high value in diagnosis of rheumatoid arthritis and correlates with disease activity. Int. J. Clin. Exp. Pathol. 2017, 10, 10451–10458. [Google Scholar]
- Li, H.M.; Wang, L.J.; Wang, Y.P.; Li, X.M.; Pan, H.F. Differences in the expression of long noncoding RNAs in peripheral blood mononuclear cells indicate potential biomarkers for rheumatoid arthritis. Int. Immunopharmacol. 2024, 134, 112218. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, S.; Yu, L.; Qu, B.; Xu, L.; Liu, L.; Sun, H.; Li, C.; Shi, Y.; Liu, H. Long noncoding RNA profiling revealed differentially expressed lncRNAs associated with disease activity in PBMCs from patients with rheumatoid arthritis. PLoS ONE 2017, 12, e0186795. [Google Scholar] [CrossRef]
- Luo, Q.; Xu, C.; Li, X.; Zeng, L.; Ye, J.; Guo, Y.; Huang, Z.; Li, J. Comprehensive analysis of long non-coding RNA and mRNA expression profiles in rheumatoid arthritis. Exp. Ther. Med. 2017, 14, 5965–5973. [Google Scholar] [CrossRef]
- Arslan, D.; Inan, B.; Kilinc, M.; Bekircan-Kurt, C.E.; Erdem-Ozdamar, S.; Tan, E. Nusinersen for adults with spinal muscular atrophy. Neurol. Sci. 2023, 44, 2393–2400. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.J. Inotersen: First Global Approval. Drugs 2018, 78, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Collotta, D.; Bertocchi, I.; Chiapello, E.; Collino, M. Antisense oligonucleotides: A novel Frontier in pharmacological strategy. Front. Pharmacol. 2023, 14, 1304342. [Google Scholar] [CrossRef] [PubMed]
- Adewunmi, O.; Shen, Y.; Zhang, X.H.; Rosen, J.M. Targeted Inhibition of lncRNA Malat1 Alters the Tumor Immune Microenvironment in Preclinical Syngeneic Mouse Models of Triple-Negative Breast Cancer. Cancer Immunol. Res. 2023, 11, 1462–1479. [Google Scholar] [CrossRef]
- Liao, Z.; Ni, Z.; Cao, J.; Liao, J.; Zhu, H.; Zhong, X.; Cao, G.; Huang, L.; Li, X.; Jiang, G.; et al. LncRNA MALAT1-Targeting Antisense Oligonucleotide Ameliorates the AngII-Induced Vascular Smooth Muscle Cell Proliferation and Migration Through Nrf2/GPX4 Antioxidant Pathway. J. Cardiovasc. Pharmacol. 2024, 84, 515–527. [Google Scholar] [CrossRef]
- Pan, Y.; Xin, W.; Wei, W.; Tatenhorst, L.; Graf, I.; Popa-Wagner, A.; Gerner, S.T.; Huber, S.E.; Kilic, E.; Hermann, D.M.; et al. Knockdown of NEAT1 prevents post-stroke lipid droplet agglomeration in microglia by regulating autophagy. Cell. Mol. Life Sci. 2024, 81, 30. [Google Scholar] [CrossRef]
- Ozes, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic targeting using tumor specific peptides inhibits long non-coding RNA HOTAIR activity in ovarian and breast cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef]
- Mou, X.; Liew, S.W.; Kwok, C.K. Identification and targeting of G-quadruplex structures in MALAT1 long non-coding RNA. Nucleic Acids Res. 2022, 50, 397–410. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| LncRNA | Impact on RA-FLS | References |
|---|---|---|
| LncNFYB | Promote proliferation | [74] |
| LINC00152 | Promote proliferation | [75,76] |
| LINC00665 | Promote proliferation | [77] |
| HOTAIR | Promote migration and invasion | [52] |
| PVT1 | Promote proliferation and suppress apoptosis | [88] |
| NEATT | Promote proliferation, migration and invasionSuppress apoptosis | [70,78,79,81] |
| RP11-83J16.1 | Promote proliferation, migration, and inflammation | [78,82] |
| NR-133666 | Promote proliferation and migration | [85] |
| HAFML | Promote migration and invasion | [80] |
| GAPLINC | Promote proliferation, migration and invasion | [24] |
| PICSAR | Promote migration and invasion | [24] |
| ZFAS1 | Promote migration and invasion | [87] |
| H19 | Promotion migration and invasion | [87] |
| SNHG1 | Suppress apoptosis | [78,80] |
| HMS | Suppress apoptosis | [78,80] |
| MALAT1 | Promote apoptosis | [86] |
| LERFS | Suppress migration, invasion and proliferation | [72,81] |
| OSER1-AS1 | Suppress proliferationPromote apoptosis | [78,81] |
| GAS5 | Suppress proliferationPromote apoptosis | [73,84] |
| DILC | Promote apoptosis | [82] |
| THRIL | Suppress proliferation, migration and invasion | [73] |
| lncRNA | Therapeutic Potential | Diagnostic Potential | References |
|---|---|---|---|
| NEAT1 | Reducing the expression of NEAT1 could be clinically beneficial | - | [17,26,32,35,70,78,79,80,81] |
| HOTAIR | Further studies are required due to conflicting results | - | [48,51,52] |
| XIST | Reducing the expression of XIST could be clinically beneficial | - | [63] |
| lnc-AL928768.3 | Reducing the expression of lnc-AL928768.3 could be clinically beneficial | AUC 0.752 | [68] |
| LncNFYB | Reducing the expression of LncNFYB could be clinically beneficial | - | [74] |
| LERFS | Increasing the expression of LERFS could be clinically beneficial | - | [72,81] |
| OSER1-AS1 | Increasing the expression of OSER1-AS1 could be clinically beneficial | - | [78] |
| DILC | Increasing the expression of DILC could be clinically beneficial | - | [82] |
| RP11-83J16.1 | Reducing the expression of RP11-83J16.1 could be clinically beneficial | - | [83] |
| MALAT1 | Reducing the expression of MALAT1 could be clinically beneficial | - | [86] |
| ITGB2-AS1 | - | AUC 0.772 | [94] |
| lnc-ITSN1-2 | - | AUC 0.898 | [98] |
| LINC00152 | - | AUC 0.618 | [99] |
| Lnc-ADM-1 | - | AUC 0.590 | |
| Lnc-FTH1-7 | - | AUC 0.655 | |
| SNHG3 | Increasing the expression of SNHG3 could be clinically beneficial | AUC 0.915 | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiełbowski, K.; Ćmil, M.; Biniek, W.J.; Bakinowska, E.; Pawlik, A. The Role of Long Non-Coding RNA in Rheumatoid Arthritis. Int. J. Mol. Sci. 2025, 26, 560. https://doi.org/10.3390/ijms26020560
Kiełbowski K, Ćmil M, Biniek WJ, Bakinowska E, Pawlik A. The Role of Long Non-Coding RNA in Rheumatoid Arthritis. International Journal of Molecular Sciences. 2025; 26(2):560. https://doi.org/10.3390/ijms26020560
Chicago/Turabian StyleKiełbowski, Kajetan, Maciej Ćmil, Wojciech Jerzy Biniek, Estera Bakinowska, and Andrzej Pawlik. 2025. "The Role of Long Non-Coding RNA in Rheumatoid Arthritis" International Journal of Molecular Sciences 26, no. 2: 560. https://doi.org/10.3390/ijms26020560
APA StyleKiełbowski, K., Ćmil, M., Biniek, W. J., Bakinowska, E., & Pawlik, A. (2025). The Role of Long Non-Coding RNA in Rheumatoid Arthritis. International Journal of Molecular Sciences, 26(2), 560. https://doi.org/10.3390/ijms26020560