
Antimicrobial Photodynamic Activity of the Zn(II) Phthalocyanine RLP068/Cl Versus Antimicrobial-Resistant Priority Pathogens


 and
and
Abstract
1. Introduction
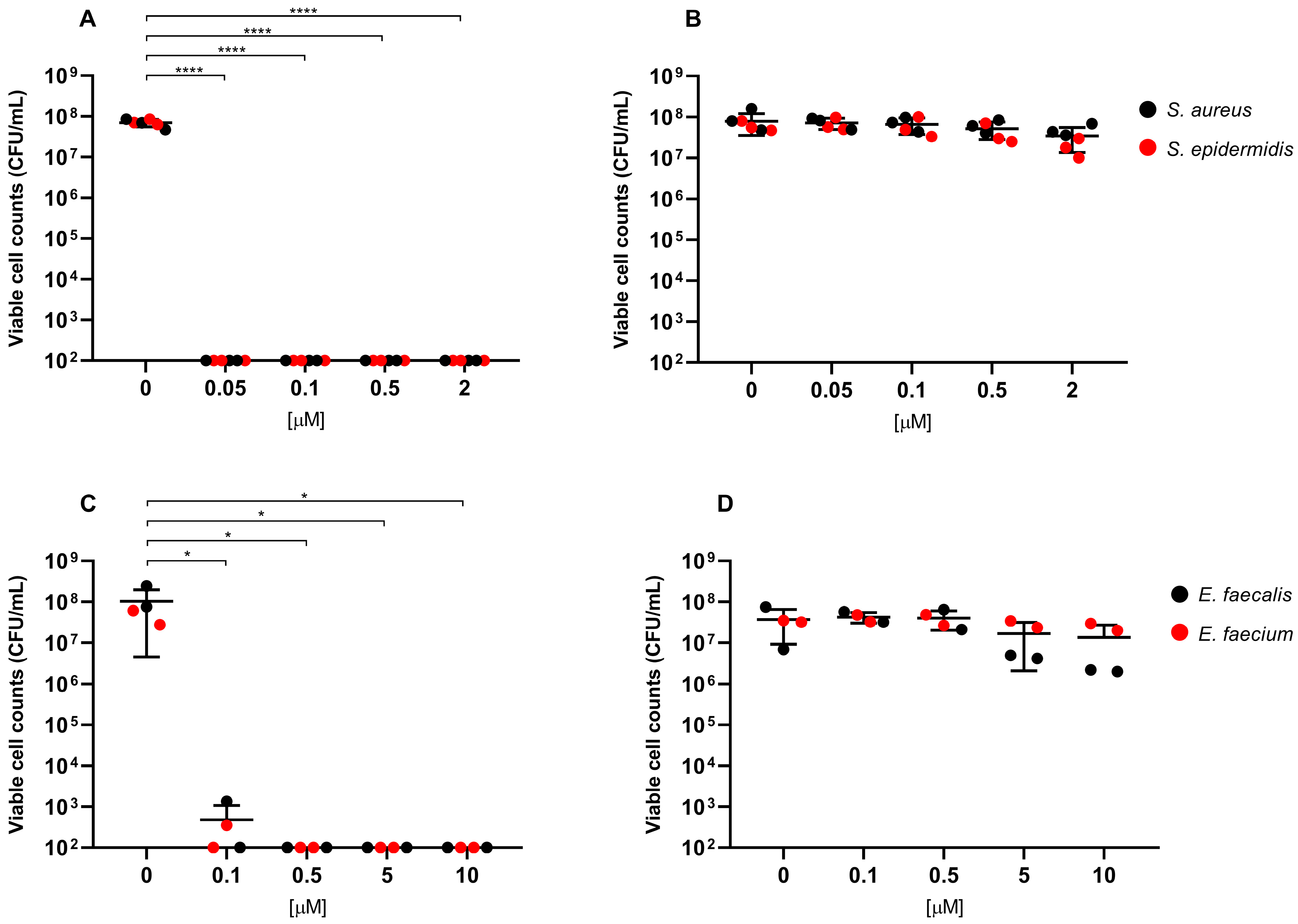
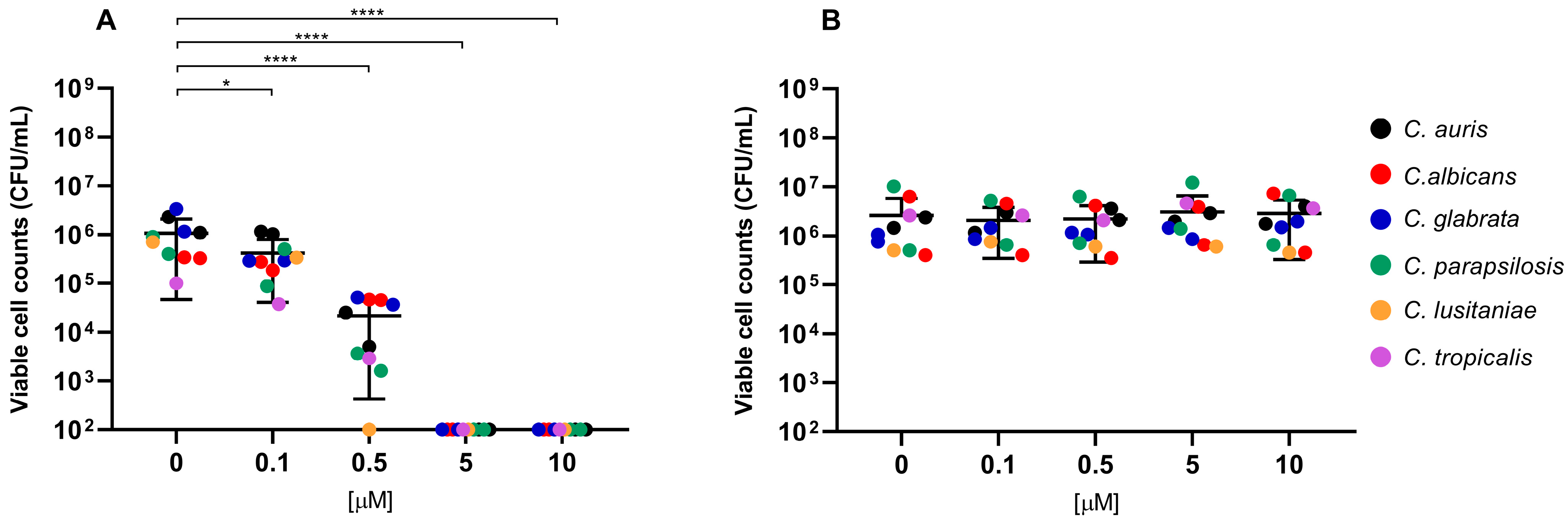
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Photosensitizer and Light Source
4.3. Phototoxicity Assay
4.4. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ho, C.S.; Wong, C.T.H.; Aung, T.T.; Lakshminarayanan, R.; Mehta, J.S.; Rauz, S.; McNally, A.; Kintses, B.; Peacock, S.J.; de la Fuente-Nunez, C.; et al. Antimicrobial resistance: A concise update. Lancet Microbe 2025, 6, 100947. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.T.; Gifford, H.; Rhodes, J. Emerging Antifungal Resistance in Fungal Pathogens. Curr. Clin. Microbiol. Rep. 2024, 11, 43–50. [Google Scholar] [CrossRef]
- Sharma, M.; Chakrabarti, A. Candidiasis and Other Emerging Yeasts. Curr. Fungal Infect. Rep. 2023, 17, 15–24. [Google Scholar] [CrossRef]
- Cristina, M.L.; Spagnolo, A.M.; Sartini, M.; Carbone, A.; Oliva, M.; Schinca, E.; Boni, S.; Pontali, E. An Overview on Candida auris in Healthcare Settings. J. Fungi 2023, 9, 913. [Google Scholar] [CrossRef]
- D’Andrea, M.M.; Fraziano, M.; Thaller, M.C.; Rossolini, G.M. The Urgent Need for Novel Antimicrobial Agents and Strategies to Fight Antibiotic Resistance. Antibiotics 2019, 8, 254. [Google Scholar] [CrossRef]
- Mathur, A.; Parihar, A.S.; Modi, S.; Kalra, A. Photodynamic Therapy for ESKAPE Pathogens: An Emerging Approach to Combat Antimicrobial Resistance (AMR). Microb. Pathog. 2023, 183, 106307. [Google Scholar] [CrossRef] [PubMed]
- Songca, S.P.; Adjei, Y. Applications of Antimicrobial Photodynamic Therapy against Bacterial Biofilms. Int. J. Mol. Sci. 2022, 23, 3209. [Google Scholar] [CrossRef]
- Allamyradov, Y.; ben Yosef, J.; Annamuradov, B.; Ateyeh, M.; Street, C.; Whipple, H.; Er, A.O. Photodynamic Therapy Review: Past, Present, Future, Opportunities and Challenges. Photochem 2024, 4, 434–461. [Google Scholar] [CrossRef]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial photodynamic therapy—What we know and what we don’t. Crit. Rev. Microbiol. 2018, 44, 571–589. [Google Scholar] [CrossRef] [PubMed]
- Leanse, L.G.; Harrington, O.D.; Fang, Y.; Ahmed, I.; Goh, X.S.; Dai, T. Evaluating the Potential for Resistance Development to Antimicrobial Blue Light (at 405 Nm) in Gram-Negative Bacteria: In Vitro and in Vivo Studies. Front. Microbiol. 2018, 9, 2403. [Google Scholar] [CrossRef]
- Surur, A.K.; de Oliveira, A.B.; De Annunzio, S.R.; Ferrisse, T.M.; Fontana, C.R. Bacterial resistance to antimicrobial photodynamic therapy: A critical update. J. Photochem. Photobiol. 2024, 255, 112905. [Google Scholar] [CrossRef]
- Giuliani, F.; Martinelli, M.; Cocchi, A.; Arbia, D.; Fantetti, L.; Roncucci, G. In vitro resistance selection studies of RLP068/Cl, a new Zn(II) phthalocyanine suitable for antimicrobial photodynamic therapy. Antimicrob. Agents Chemother. 2010, 54, 637–642. [Google Scholar] [CrossRef]
- Soncin, M.C.; Fabbris, A.; Busetti, D.; Dei, D.; Nistri, G. Roncucci, and G. Jori. Approaches to selectivity in the Zn(II)-phthalocyanine-photosensitized inactivation of wild-type and antibiotic-resistant Staphylococcus aureus. Photochem. Photobiol. Sci. 2002, 1, 815–819. [Google Scholar] [CrossRef]
- Vassena, C.; Fenu, S.; Giuliani, F.; Fantetti, L.; Roncucci, G.; Simonutti, G.; Romanò, C.L.; De Francesco, R.; Drago, L. Photodynamic Antibacterial and Antibiofilm Activity of RLP068/Cl against Staphylococcus Aureus and Pseudomonas Aeruginosa Forming Biofilms on Prosthetic Material. Int. J. Antimicrob. Agents 2014, 44, 47–55. [Google Scholar] [CrossRef]
- Simonetti, O.; Cirioni, O.; Orlando, F.; Alongi, C.; Lucarini, G.; Silvestri, C.; Zizzi, A.; Fantetti, L.; Roncucci, G.; Giacometti, A.; et al. Effectiveness of Antimicrobial Photodynamic Therapy with a Single Treatment of RLP068/Cl in an Experimental Model of Staphylococcus aureus Wound Infection. Br. J. Dermatol. 2011, 164, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Monami, M.; Scatena, A.; Schlecht, M.; Lobmann, R.; Landi, L.; Ricci, L.; Mannucci, E. Antimicrobial Photodynamic Therapy in Infected Diabetic Foot Ulcers: A Multicenter Preliminary Experience. J. Am. Podiatr. Med. Assoc. 2020, 110, 5. [Google Scholar] [CrossRef] [PubMed]
- Mosti, G.; Picerni, P.; Licau, M.; Mattaliano, V. Photodynamic Therapy in Infected Venous and Mixed Leg Ulcers: A Pilot Experience. J. Wound Care 2018, 27, 816–821. [Google Scholar] [CrossRef]
- Vecchio, D.; Dai, T.; Huang, L.; Fantetti, L.; Roncucci, G.; Hamblin, M.R. Antimicrobial photodynamic therapy with RLP068 kills methicillin-resistant Staphylococcus aureus and improves wound healing in a mouse model of infected skin abrasion PDT with RLP068/Cl in Infected Mouse Skin Abrasion. J. Biophotonics 2012, 6, 733–742. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Yan, E.; Kwek, G.; Qing, N.S.; Lingesh, S.; Xing, B. Antimicrobial Photodynamic Therapy for the Remote Eradication of Bacteria. ChemPlusChem 2023, 88, e202300009. [Google Scholar] [CrossRef]
- Ribeiro, M.; Gomes, I.B.; Saavedra, M.J.; Simões, M. Photodynamic Therapy and Combinatory Treatments for the Control of Biofilm-Associated Infections. Lett. Appl. Microbiol. 2022, 75, 548–564. [Google Scholar] [CrossRef]
- Oyama, J.; Ramos-Milaré, Á.C.F.H.; Lera-Nonose, D.S.S.L.; Nesi-Reis, V.; Demarchi, I.G.; Aristides, S.M.A.; Teixeira, J.J.V.; Silveira, T.G.V.; Lonardoni, M.V.C. Photodynamic Therapy in Wound Healing in Vivo: A Systematic Review. Photodiagnosis Photodyn. Ther. 2020, 30, 101682. [Google Scholar] [CrossRef]
- De Mello, M.M.; De Barros, P.P.; De Cassia Bernardes, R.; Alves, S.R.; Ramanzini, N.P.; Figueiredo-Godoi, L.M.A.; Prado, A.C.C.; Jorge, A.O.C.; Junqueira, J.C. Antimicrobial Photodynamic Therapy against Clinical Isolates of Carbapenem-Susceptible and Carbapenem-Resistant Acinetobacter baumannii. Lasers Med. Sci. 2019, 34, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Edwards, L.; Turner, D.; Champion, C.; Khandelwal, M.; Zingler, K.; Stone, C.; Rajapaksha, R.D.; Yang, J.; Ranasinghe, M.I.; Kornienko, A.; et al. Photoactivated 2,3-Distyrylindoles Kill Multi-Drug Resistant Bacteria. Bioorganic Med. Chem. Lett. 2018, 28, 1879–1886. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, Y.; Wang, L.; Han, L.; Lei, J.; Ishaq, H.M.; Xu, J. Mechanistic Aspects of the Photodynamic Inactivation of Vancomycin-Resistant Enterococci Mediated by 5-Aminolevulinic Acid and 5-Aminolevulinic Acid Methyl Ester. Curr. Microbiol. 2014, 70, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Songsantiphap, C.; Vanichanan, J.; Chatsuwan, T.; Asawanonda, P.; Boontaveeyuwat, E. Methylene Blue–Mediated Antimicrobial Photodynamic Therapy against Clinical Isolates of Extensively Drug Resistant Gram-Negative Bacteria Causing Nosocomial Infections in Thailand, an in Vitro Study. Front. Cell. Infect. Microbiol. 2022, 12, 929242. [Google Scholar] [CrossRef] [PubMed]
- Mantareva, V.; Kussovski, V.; Angelov, I.; Borisova, E.; Avramov, L.; Schnurpfeil, G.; Wöhrle, D. Photodynamic Activity of Water-Soluble Phthalocyanine Zinc(II) Complexes against Pathogenic Microorganisms. Bioorganic Med. Chem. 2007, 15, 4829–4835. [Google Scholar] [CrossRef]
- Mantareva, V.; Gol, C.; Kussovski, V.; Durmuş, M.; Angelov, I. Impact of Water-Soluble Zwitterionic Zn(II) Phthalocyanines against Pathogenic Bacteria. Z. Naturforschung C 2019, 74, 183–191. [Google Scholar] [CrossRef]
- Miretti, M.; Clementi, R.; Tempesti, T.C.; Baumgartner, M.T. Photodynamic Inactivation of Multiresistant Bacteria (KPC) Using Zinc(II)Phthalocyanines. Bioorganic Med. Chem. Lett. 2017, 27, 4341–4344. [Google Scholar] [CrossRef]
- Espitia-Almeida, F.; Valle-Molinares, R.; Quiroz, E.N.; Pacheco-Londoño, L.C.; Galán-Freyle, N.J. Photodynamic Antimicrobial Activity of a Novel 5,10,15,20-Tetrakis (4-Ethylphenyl) Porphyrin against Clinically Important Bacteria. Pharmaceuticals 2023, 16, 1059. [Google Scholar] [CrossRef]
- Hamblin, M.R.; Hasan, T. Photodynamic Therapy: A New Antimicrobial Approach to Infectious Disease? Photochem. Photobiol. Sci. 2004, 3, 436–450. [Google Scholar] [CrossRef]
- Sperandio, F.; Huang, Y.-Y.; Hamblin, M. Antimicrobial Photodynamic Therapy to Kill Gram-Negative Bacteria. Recent Pat. Anti-Infect. Drug Discov. 2013, 8, 108–120. [Google Scholar] [CrossRef]
- Huang, L.; Xuan, Y.; Koide, Y.; Zhiyentayev, T.; Tanaka, M.; Hamblin, M.R. Type I and Type II Mechanisms of Antimicrobial Photodynamic Therapy: An in Vitro Study on Gram-negative and Gram-positive Bacteria. Lasers Surg. Med. 2012, 44, 490–499. [Google Scholar] [CrossRef]
- Elagin, V.; Budruev, I.; Antonyan, A.; Bureev, P.; Ignatova, N.; Streltsova, O.; Kamensky, V. Enhancement of the Efficacy of Photodynamic Therapy against Uropathogenic Gram-Negative Bacteria Species. Photonics 2023, 10, 310. [Google Scholar] [CrossRef]
- Yanten, N.; Vilches, S.; Palavecino, C.E. Photodynamic Therapy for the Treatment of Pseudomonas aeruginosa Infections: A Scoping Review. Photodiagnosis Photodyn. Ther. 2023, 44, 103803. [Google Scholar] [CrossRef]
- Rezaie, P.; Pourhajibagher, M.; Chiniforush, N.; Hosseini, N.; Bahador, A. The Effect of Quorum-Sensing and Efflux Pumps Interactions in Pseudomonas aeruginosa Against Photooxidative Stress. J. Lasers Med. Sci. 2018, 9, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, I.V.; Da Silva Oliveira, D.D.; Calefi, G.G.; Silva, N.B.S.; Martins, C.H.G.; De Oliveira Rezende Júnior, C.; Tsubone, T.M. Photosensitizer Associated with Efflux Pump Inhibitors as a Strategy for Photodynamic Therapy against Bacterial Resistance. Eur. J. Med. Chem. 2024, 284, 117197. [Google Scholar] [CrossRef]
- Batishchev, O.V.; Kalutskii, M.A.; Varlamova, E.A.; Konstantinova, A.N.; Makrinsky, K.I.; Ermakov, Y.A.; Meshkov, I.N.; Sokolov, V.S.; Gorbunova, Y.G. Antimicrobial Activity of Photosensitizers: Arrangement in Bacterial Membrane Matters. Front. Mol. Biosci. 2023, 10, 1192794. [Google Scholar] [CrossRef] [PubMed]
- Souza, S.O.; Raposo, B.L.; Sarmento-Neto, J.F.; Rebouças, J.S.; Macêdo, D.P.C.; Figueiredo, R.C.B.Q.; Santos, B.S.; Freitas, A.Z.; Filho, P.E.C.; Ribeiro, M.S.; et al. Photoinactivation of Yeast and Biofilm Communities of Candida albicans Mediated by ZnTnHex-2-PyP4+ Porphyrin. J. Fungi 2022, 8, 556. [Google Scholar] [CrossRef]
- Mendoza-Reyes, D.F.; Gómez-Gaviria, M.; Mora-Montes, H.M. Candida Lusitaniae: Biology, Pathogenicity, Virulence Factors, Diagnosis, and Treatment. Infect. Drug Resist. 2022, 15, 5121–5135. [Google Scholar] [CrossRef]
- Karner, L.; Drechsler, S.; Metzger, M.; Hacobian, A.; Schädl, B.; Slezak, P.; Grillari, J.; Dungel, P. Antimicrobial Photodynamic Therapy Fighting Polymicrobial Infections—A Journey from in Vitro to in Vivo. Photochem. Photobiol. Sci. 2020, 19, 1332–1343. [Google Scholar] [CrossRef] [PubMed]
- Fabris, C.M.; Soncin, E.; Mazzon, P.; Calzavara-Pinton, F.; Lia, C.; Giacomo, D.; Dei, S.; Tampucci, G. Roncucci, and G. Jori. A novel tetracationic phthalocyanine as a potential skin phototherapeutic agent. Exp. Dermatol. 2005, 14, 675–683. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Strain ID | Species | Resistance Phenotype (Genes) a | MBC (μM) ≥ 3 log b,c | MBC (μM) ≥ 5 log b,c |
|---|---|---|---|---|
| FI-01 | Klebsiella pneumoniae | CEFR, COLR, AGR, FLQR, CARBAR (blaNDM) (blaNDM) (blaNDM) (blaNDM) | 5 | 10 |
| FI-02 | Klebsiella pneumoniae | CEFR, AGR, CARBAR (blaNDM) | 5 | 10 |
| FI-03 | Klebsiella pneumoniae | CEFR, AGR, FLQR, CARBAR (blaNDM) | 5 | 10 |
| FI-04 | Klebsiella pneumoniae | CEFR, AGR, FLQR, CARBAR (blaNDM) | 10 | 20 |
| FI-05 | Klebsiella pneumoniae | CEFR, COLR, FLQR, CARBAR (mcr-1.2; blaKPC-3) | 5 | 10 |
| FI-06 | Klebsiella pneumoniae | CEFR, COLR, FLQR (blaVIM-1) | 10 | 20 |
| FI-07 | Klebsiella pneumoniae | CEFR, COLR, AGR, FLQR, CARBAR (blaKPC-2) | 10 | 20 |
| FI-08 | Klebsiella pneumoniae | CEFR, COLR, FLQR, CARBAR (blaKPC-3) | 10 | 20 |
| FI-09 | Klebsiella pneumoniae | CEFR, COLR, FLQR, CARBAR (blaKPC-3) | 10 | 20 |
| FI-10 | Klebsiella pneumoniae | CEFR, COLR, AGR, FLQR, CARBAR (blaKPC-3) | 5 | 20 |
| FI-11 | Klebsiella pneumoniae | CEFR, FLQR, CARBAR (blaKPC-2) | 5 | 10 |
| FI-12 | Klebsiella pneumoniae | CEFR, AGR, FLQR, CARBAR (blaOXA-48-like) | 10 | 20 |
| FI-14 | Klebsiella pneumoniae | CEFR, COLR, FLQR, CARBAR (blaKPC-3) | 20 | 40 |
| FI-15 | Klebsiella pneumoniae | CEFR, FLQR (blaKPC-31) | 10 | 20 |
| FI-20 | Klebsiella pneumoniae | CEFR, FLQR, CARBAR (blaKPC-3) | 10 | 20 |
| FI-13 | Escherichia coli | CEFR, FLQR (blaOXA-48-like) | 5 | 5 |
| FI-16 | Escherichia coli | CEFR, AGR, FLQR, CARBAR (blaKPC, blaVIM) | 5 | 10 |
| FI-17 | Serratia marcescens | CEFR, COLR, AGR, FLQR, CARBAR (blaNDM) | 10 | 10 |
| FI-18 | Citrobacter freundii | CEFR, FLQR, CARBAR (blaVIM) | 10 | 20 |
| FI-21 | Citrobacter freundii | FLQR, CARBAR (blaVIM) | 10 | 20 |
| FI-22 | Citrobacter freundii | CEFR, FLQR, CARBAR (blaKPC, blaVIM) | 10 | 20 |
| FI-24 | Providencia stuartii | CEFR, AGR, FLQR, CARBAR (blaKPC) | 10 | 20 |
| FI-32 | Enterobacter ludwigii | CARBAR (blaNMC-A) | 10 | 20 |
| FI-33 | Enterobacter ludwigii | CARBAR (blaIMI-2) | 10 | 20 |
| FI-23 | Pseudomonas aeruginosa | FLQR, CARBAR (blaIMP-13) | 20 | >40 |
| FI-25 | Pseudomonas aeruginosa | FLQR, CARBAR (blaVIM-1) | 20 | >40 |
| FI-26 | Pseudomonas aeruginosa | AGR, FLQR, CARBAR (blaVIM-2) | 10 | 20 |
| FI-27 | Pseudomonas aeruginosa | FLQR, CARBAR (blaVIM-1) | 10 | 20 |
| FI-28 | Pseudomonas aeruginosa | AGR, FLQR, CARBAR (blaVIM-2) | 10 | 20 |
| FI-29 | Pseudomonas aeruginosa | AGR, FLQR, CARBAR (blaGES-5) | 5 | 10 |
| FI-34 | Pseudomonas aeruginosa | CEFR, AGR, FLQR, CARBAR (blaFIM) | 10 | 40 |
| FI-30 | Acinetobacter baumannii | AGR, FLQR, CARBAR (blaOXA-23) | 5 | 10 |
| FI-31 | Acinetobacter baumannii | FLQR, CARBAR (blaNDM) | 5 | 5 |
| FI-35 | Acinetobacter baumannii | AGR, FLQR, CARBAR | 5 | 10 |
| FI-36 | Acinetobacter baumannii | AGR, FLQR, CARBAR | 5 | 10 |
| FI-37 | Acinetobacter baumannii | AGR, FLQR, CARBAR | 5 | 5 |
| FI-38 | Acinetobacter baumannii | AGR, FLQR, CARBAR | 5 | 5 |
| FI-39 | Acinetobacter baumannii | AGR, FLQR, CARBAR | 5 | 10 |
| FI-40 | Acinetobacter baumannii | AGR, FLQR, CARBAR (blaOXA-23) | 5 | 10 |
| Strain ID | Species | Resistance Phenotype (Genes) a | MBC (μM) ≥ 3 log b,c | MBC (μM) ≥ 5 log b,c |
|---|---|---|---|---|
| FI-43 | Staphylococcus aureus | ERYR, OXAR, LINR (cfr, poxtA) | 0.05 | 0.05 |
| FI-45 | Staphylococcus aureus | ERYR, OXAR | 0.05 | 0.05 |
| FI-50 | Staphylococcus aureus | OXAR, DAPTOR | 0.05 | 0.05 |
| FI-44 | Staphylococcus epidermidis | ERYR, OXAR, LINR (cfr) | 0.05 | 0.05 |
| FI-46 | Staphylococcus epidermidis | ERYR, OXAR, DAPTOR | 0.05 | 0.05 |
| FI-47 | Staphylococcus epidermidis | OXAR, LINR (cfr) | 0.05 | 0.05 |
| FI-41 | Enterococcus faecalis | ERYR, OXAR, LINR (optrA) | 0.1 | 0.1 |
| FI-42 | Enterococcus faecalis | ERYR, OXAR, LINR (optrA) | 0.1 | 0.1 |
| FI-48 | Enterococcus faecium | GLYR (vanA) | 0.1 | 0.1 |
| FI-49 | Enterococcus faecium | GLYR (vanB) | 0.1 | 0.1 |
| Strain ID | Species | Resistance Phenotype a | MBC (μM) ≥ 3 log b,c |
|---|---|---|---|
| FI-51 | Candida auris | None (Reference type strain) | 5 |
| FI-52 | Candida auris | None (Reference type strain) | 5 |
| FI-53 | Candida albicans | None | 5 |
| FI-54 | Candida albicans | AZLR, ECINR | 5 |
| FI-55 | Candida glabrata | None | 5 |
| FI-56 | Candida glabrata | None | 5 |
| FI-57 | Candida parapsilosis | None | 5 |
| FI-58 | Candida parapsilosis | ECINR | 5 |
| FI-59 | Candida lusitaniae | None | 0.5 |
| FI-60 | Candida tropicalis | None | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baccani, I.; Cuffari, S.; Giuliani, F.; Rossolini, G.M.; Pollini, S. Antimicrobial Photodynamic Activity of the Zn(II) Phthalocyanine RLP068/Cl Versus Antimicrobial-Resistant Priority Pathogens. Int. J. Mol. Sci. 2025, 26, 7545. https://doi.org/10.3390/ijms26157545
Baccani I, Cuffari S, Giuliani F, Rossolini GM, Pollini S. Antimicrobial Photodynamic Activity of the Zn(II) Phthalocyanine RLP068/Cl Versus Antimicrobial-Resistant Priority Pathogens. International Journal of Molecular Sciences. 2025; 26(15):7545. https://doi.org/10.3390/ijms26157545
Chicago/Turabian StyleBaccani, Ilaria, Sara Cuffari, Francesco Giuliani, Gian Maria Rossolini, and Simona Pollini. 2025. "Antimicrobial Photodynamic Activity of the Zn(II) Phthalocyanine RLP068/Cl Versus Antimicrobial-Resistant Priority Pathogens" International Journal of Molecular Sciences 26, no. 15: 7545. https://doi.org/10.3390/ijms26157545
APA StyleBaccani, I., Cuffari, S., Giuliani, F., Rossolini, G. M., & Pollini, S. (2025). Antimicrobial Photodynamic Activity of the Zn(II) Phthalocyanine RLP068/Cl Versus Antimicrobial-Resistant Priority Pathogens. International Journal of Molecular Sciences, 26(15), 7545. https://doi.org/10.3390/ijms26157545