

Systematic Analysis of Dof Gene Family in Prunus persica Unveils Candidate Regulators for Enhancing Cold Tolerance
Abstract
1. Introduction
2. Results
2.1. Effects of ALA Treatment on Floral Organs and Physiological and Biochemical Parameters in Peach Blossoms
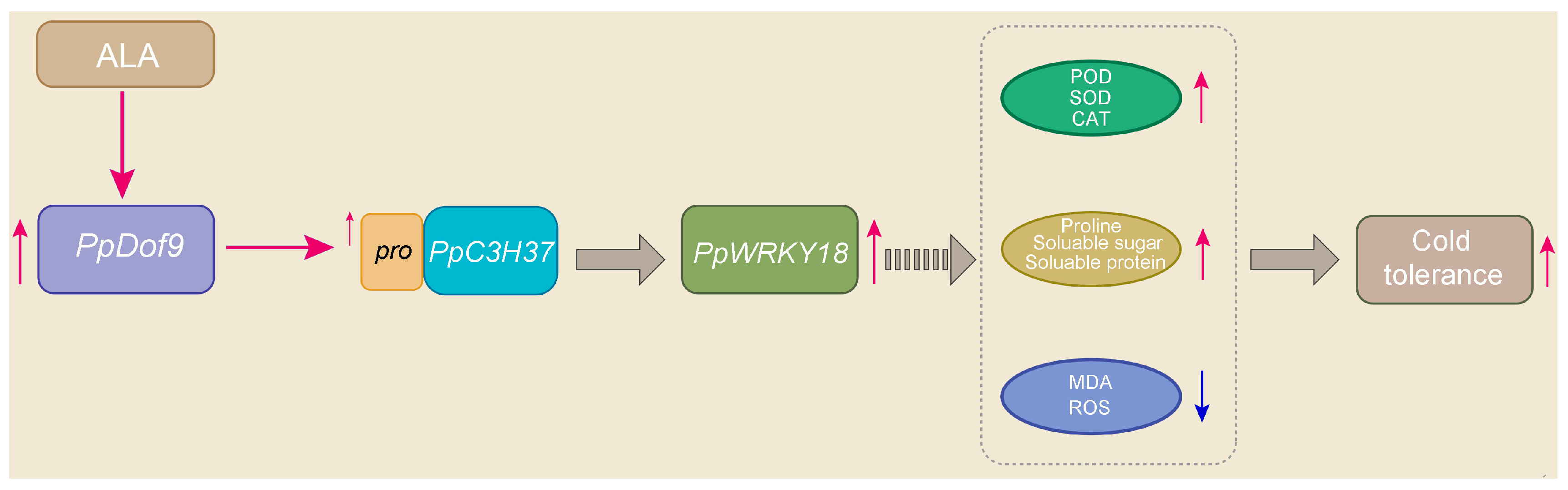
2.2. Transcription Factor PpDof9 Interacted with the PpC3H37 Promoter
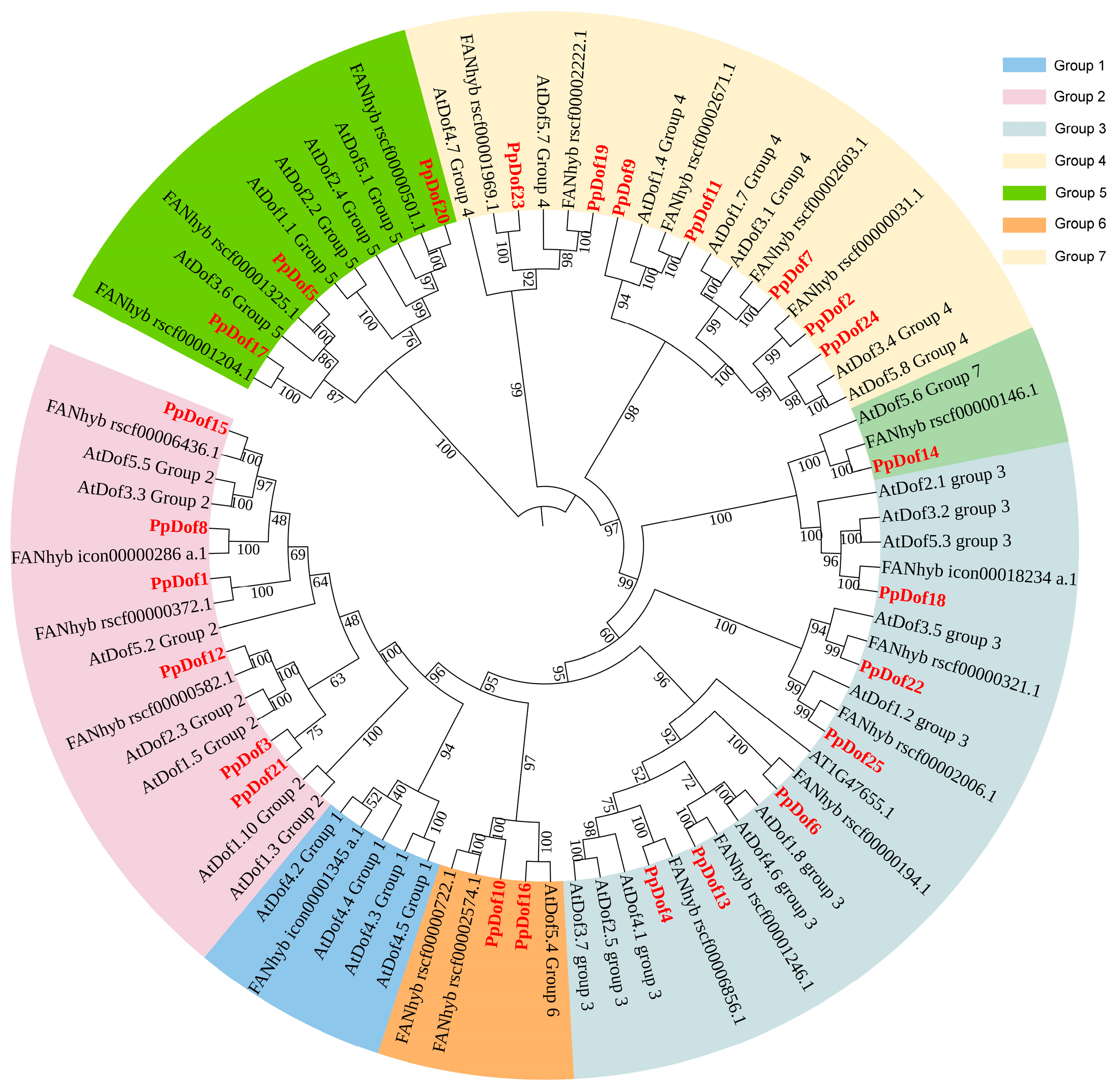
2.3. Identification of PpDof Gene Family Members
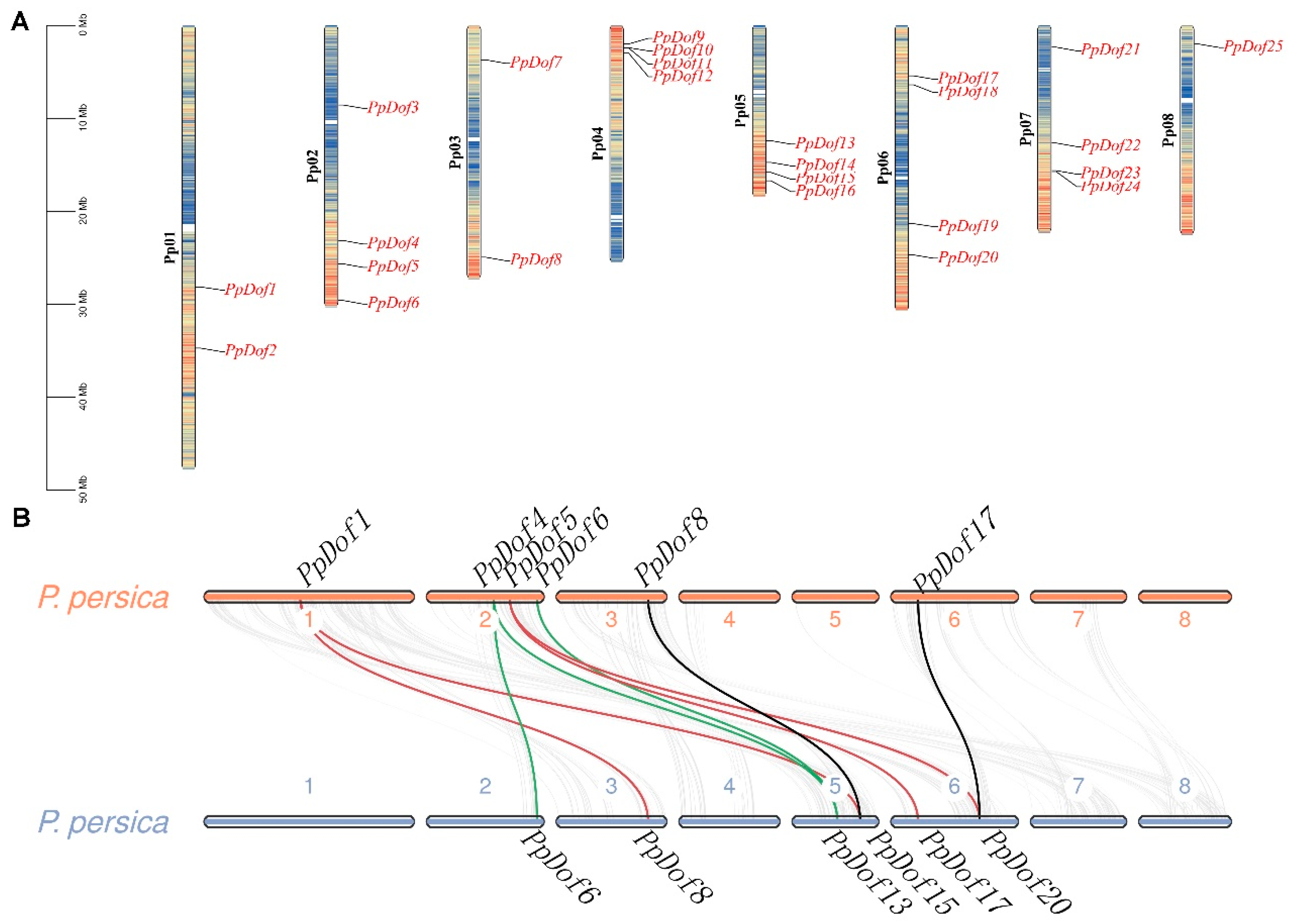
2.4. Chromosomal Localization and Synteny Analysis of the PpDof Gene Family in Peaches
2.5. Analysis of the Conserved Motifs of the PpDof Gene Family
2.6. Analysis of Cis-Regulatory Elements in the Promoters of PpDof Family Members
2.7. ALA Regulated PpDof-Related Genes Under Cold Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemical Treatments
4.2. Physiological and Biochemical Parameter Determination in Peach Ovaries
4.3. Yeast One-Hybrid Assay
4.4. Identification of Dof Genes in Peaches, Construction of a Phylogenetic Tree and Collinearity Analysis
4.5. Calculation of Basic Information for Gene Family Members
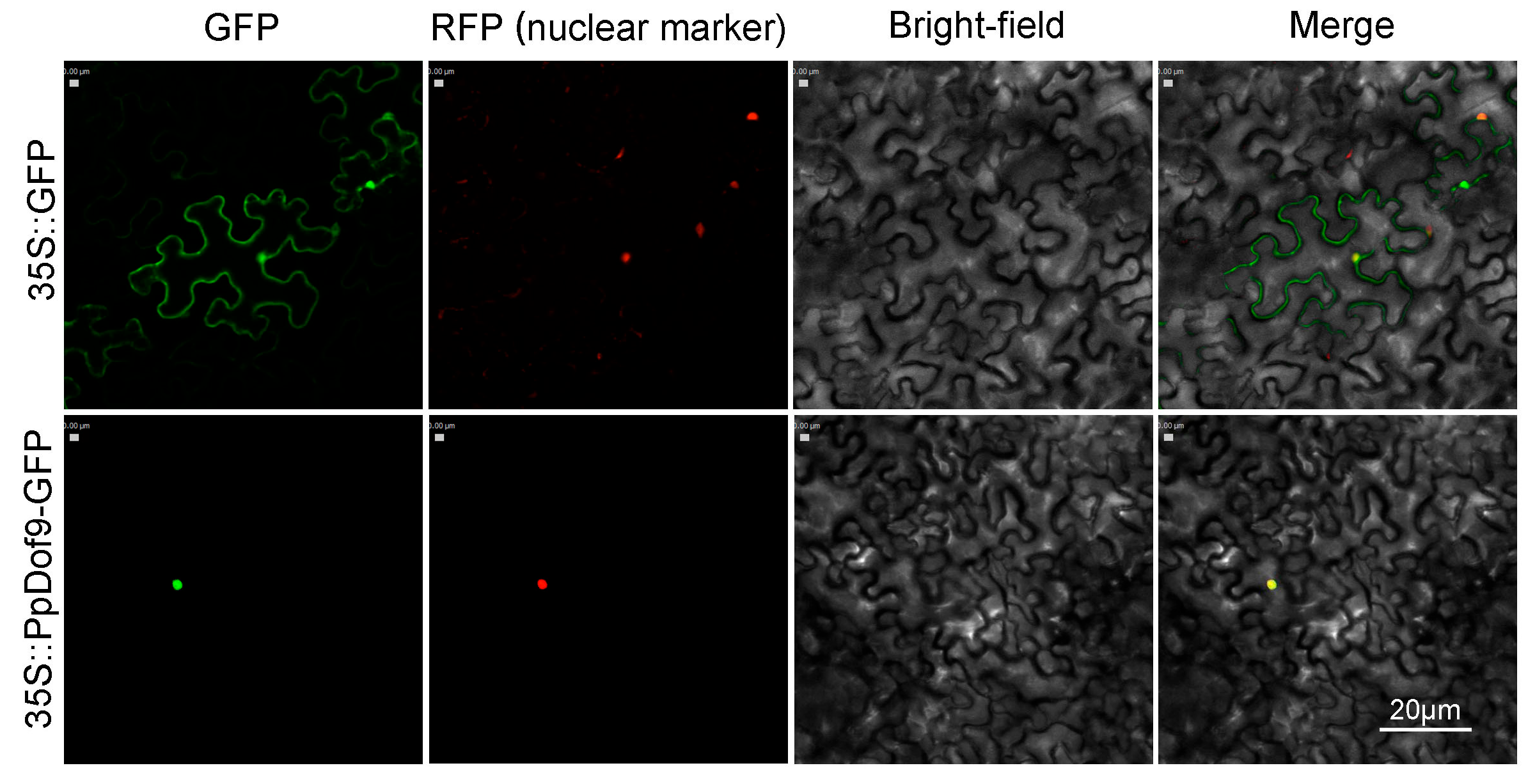
4.6. Subcellular Localization of PpDof9
4.7. RNA Extraction, Reverse Transcription, and RT-qPCR Analysis
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.W.; Meng, X.Q.; Jia, H.J.; Yu, M.-L.; Ma, R.-J.; Wang, L.-R.; Cao, K.; Shen, Z.-J.; Niu, L.; Tian, J.-B.; et al. Peach genetic resources: Diversity, population structure and linkage disequilibrium. BMC Genet. 2013, 14, 84. [Google Scholar] [CrossRef]
- Shen, Z.J.; Ma, R.J.; Cai, Z.X.; Yu, M.; Zhang, Z. Diversity, population structure, and evolution of local peach cultivars in China identified by simple sequence repeats. Genet. Mol. Res. 2015, 14, 101–117. [Google Scholar] [CrossRef]
- Abdel-Sattar, M.; Al-Obeed, R.S.; Aboukarima, A.M.; Eshra, D.H. Development of an artificial neural network as a tool for predicting the chemical attributes of fresh peach fruits. PLoS ONE 2021, 16, e0251185. [Google Scholar] [CrossRef]
- Atagul, O.; Calle, A.; Demirel, G.; Lawton, J.M.; Bridges, W.C.; Gasic, K. Estimating heat requirement for flowering in peach germplasm. Agronomy 2022, 12, 1002. [Google Scholar] [CrossRef]
- Penso, G.A.; Citadin, I.; Scariotto, S.; Santos, C.E.M.D.; Junior, A.W.; Bruckner, C.H.; Rodrigo, J. Development of peach flower buds under low winter chilling conditions. Agronomy 2020, 10, 428. [Google Scholar] [CrossRef]
- Wang, H.P.; Liu, Z.C.; Li, J.M.; Luo, S.L.; Zhang, J.; Xie, J.M. Hydrogen sulfide interacts with 5-aminolevulinic acid to enhance the antioxidant capacity of pepper (Capsicum annuum L.) seedlings under chilling stress. Agronomy 2022, 12, 572. [Google Scholar] [CrossRef]
- Han, D.G.; Zhang, Z.Y.; Ding, H.B.; Wang, Y.F.; Liu, W.; Li, H.X.; Yang, G.H. Molecular cloning and functional analysis of MbWRKY3 involved in improved drought tolerance in transformed tobacco. J. Plant Interact. 2018, 13, 329–337. [Google Scholar] [CrossRef]
- Liu, W.D.; Wang, T.H.; Wang, Y.; Liang, X.Q.; Han, J.L.; Han, D.G. MbMYBC1, a M. baccata MYB transcription factor, contribute to cold and drought stress tolerance in transgenic Arabidopsis. Front. Plant Sci. 2023, 14, 1141446. [Google Scholar] [CrossRef]
- Li, W.H.; Li, P.; Chen, H.Y.; Zhong, J.L.; Liang, X.Q.; Wei, Y.F.; Zhang, L.H.; Wang, H.B.; Han, D.G. Overexpression of a Fragaria vesca 1R-MYB Transcription Factor Gene (FvMYB114) Increases Salt and Cold Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2023, 24, 5261. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Wei, Y.F.; Zhang, L.H.; Wang, Y.; Song, P.H.; Li, X.G.; Han, D.G. FvMYB44, a strawberry R2R3-MYB transcription factor, improved salt and cold stress tolerance in transgenic Arabidopsis. Agronomy 2023, 13, 1051. [Google Scholar] [CrossRef]
- Umemura, Y.; Ishiduka, T.; Yamamoto, R.; Esaka, M. The Dof domain, a zinc finger DNA-binding domain conserved only in higher plants, truly functions as a Cys2/Cys2 Zn finger domain. Plant J. 2004, 37, 741–749. [Google Scholar] [CrossRef]
- Marzábal, P.; Gas, E.; Fontanet, P.; Vicente-Carbajosa, J.; Torrent, M.; Ludevid, M.D. The maize Dof protein PBF activates transcription of gamma-zein during maize seed development. Plant Mol. Biol. 2008, 67, 441–454. [Google Scholar] [CrossRef]
- Tokunaga, S.; Sanda, S.; Uraguchi, Y.; Nakagawa, S.; Sawayama, S. Overexpression of the DOF-Type Transcription Factor Enhances Lipid Synthesis in Chlorella vulgaris. Appl. Biochem. Biotechnol. 2019, 189, 116–128. [Google Scholar] [CrossRef]
- Song, X.W.; Tang, S.J.; Liu, H.; Meng, Y.; Luo, H.F.; Wang, B.; Hou, X.L.; Yan, B.; Yang, C.; Guo, Z.H.; et al. Inheritance of acquired adaptive cold tolerance in rice through DNA methylation. Cell 2025, 188, 1–12. [Google Scholar] [CrossRef]
- Wang, T.; Yue, J.J.; Wang, X.J.; Xu, L.; Li, L.-B.; Gu, X.-P. Genome-wide identification and characterization of the Dof gene family in moso bamboo (Phyllostachys heterocycla var. Pubescens). Genes Genom. 2016, 38, 733–745. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, L.J. ALA upregulates MdPTPA expression to increase the PP2A activity and promote stomatal opening in apple leaves. Plant Sci. 2022, 325, 111490. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lou, Y.R.; Wang, L.J. MdDGK3-like as a Negative Regulator Participates in ALA-Induced PP2AC to Promote Stomatal Opening in Apple leaves. Hortic. Plant J. 2023, 9, 898–908. [Google Scholar] [CrossRef]
- Sun, Y.P.; Zhang, Z.; Wang, L.J. Promotion of 5-aminolevulinic acid treatment on leaf photosynthesis is related with increase of antioxidant enzyme activity in watermelon seedlings grown under shade condition. Photosynthetica 2009, 47, 347–354. [Google Scholar] [CrossRef]
- Sun, Y.P.; Liu, J.; Cao, R.X.; Huang, Y.J.; Hall, A.M.; Guo, C.B.; Wang, L.J. Effects of 5-aminolevulinic acid treatment on photosynthesis of strawberry. Photosynthetica 2017, 55, 276–284. [Google Scholar] [CrossRef]
- Zheng, J.; An, Y.Y.; Wang, L.J. 24-Epibrassinolide enhances 5-ALA-induced anthocyanin and flavonol accumulation in calli of ‘Fuji’ apple flesh. Plant Cell Tissue Organ Cult. 2018, 134, 319–330. [Google Scholar] [CrossRef]
- Liang, R.L.; Wang, X.Q.; Zhang, J.; Gan, X.; Wang, L.J. Effects of exogenous ALA on leaf photosynthesis, photosynthate transport, and sugar accumulation in Prunus persica L. Forests 2023, 14, 723. [Google Scholar] [CrossRef]
- Cai, C.Y.; He, S.S.; An, Y.Y.; Wang, L.J. Exogenous 5-aminolevulinic acid improves strawberry tolerance to osmotic stress and its possible mechanisms. Physiol. Plant. 2022, 168, 948–962. [Google Scholar] [CrossRef] [PubMed]
- An, Y.Y.; Qi, L.; Wang, L.J. ALA pretreatment improves wa terlogging tolerance of fig plants. PLoS ONE 2016, 11, e147202. [Google Scholar] [CrossRef]
- He, S.S.; Yang, H.; Cao, R.Q.; Tang, Q.; An, Y.Y.; Wang, L.J. 5-aminolevulinic acid-induced salt tolerance in strawberry (cv. ‘Benihoppe’): Possible role of nitric oxide on interception of salt ions in roots. Sci. Hortic. 2022, 304, 111294. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hoseiny, H.M.; Elsheery, N.I.; Kalaji, H.M.; de Los Santos-Villalobos, S.; Wróbel, J.; Hassan, I.F.; Gaballah, M.S.; Abdelrhman, L.A.; Mira, A.M.; et al. 5-aminolevulinic acid and 24-epibrassinolide improve the drought stress resilience and productivity of banana plants. Plants 2022, 11, 743. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Zhang, J.T.; Liu, L.B.; Zhang, L.Z.; Gan, X.; Zhong, Y.; Wang, L.J. ALA up-regulated PpWRKY18 to enhance freezing tolerance of nectarine pistils. Hortic. Plant J. 2024, in press. [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.X.; Li, L.J.; Zhou, J.; He, M.Q.; Lyu, D.G.; Zhao, D.Y.; Qin, S.J. Integrated transcriptomics and metabolomics analyses reveal key genes and essential metabolic pathways for the acquisition of cold tolerance during dormancy in apple. Environ. Exp. Bot. 2023, 213, 105413. [Google Scholar] [CrossRef]
- Wang, L.J.; Zhang, J.T.; Zhong, Y.; Zhang, L.Z.; Yang, H.; Liu, L.B.; Zhou, J.Y.; Iqbal, M.M.; Gan, X. Regulation of 5-Aminolevunilic acid and its application in agroforestry. Forests 2023, 14, 1857. [Google Scholar] [CrossRef]
- Wen, C.L.; Cheng, Q.; Zhao, L.; Mao, A.; Yang, J.; Yu, S.; Weng, Y.; Xu, Y. Identification and characterisation of Dof transcription factors in the cucumber genome. Sci. Rep. 2016, 6, 23072. [Google Scholar] [CrossRef]
- Lee, B.D.; Kim, M.R.; Kang, M.Y.; Cha, J.Y.; Han, S.H.; Nawkar, G.M.; Sakuraba, Y.; Lee, S.Y.; Imaizumi, T.; McClung, C.R.; et al. The F-box protein FKF1 inhibits dimerization of COP1 in the control of photoperiodic flowering. Nat. Commun. 2017, 8, 2259. [Google Scholar] [CrossRef]
- Corrales, A.R.; Nebauer, S.G.; Carrillo, L.; Fernández-Nohales, P.; Marqués, J.; Renau-Morata, B.; Granell, A.; Pollmann, S.; Vicente-Carbajosa, J.; Molina, R.V.; et al. Characterization of tomato Cycling Dof Factors reveals conserved and new functions in the control of flowering time and abiotic stress responses. J. Exp. Bot. 2014, 65, 995–1012. [Google Scholar] [CrossRef]
- Lijavetzky, D.; Carbonero, P.; Vicente-Carbajosa, J. Genome-wide comparative phylogenetic analysis of the rice and Arabidopsis Dof gene families. BMC Evol. Biol. 2003, 3, 17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zeng, B.; Zhao, H.; Zhang, M.; Xie, S.; Lai, J. Genome-wide transcription factor gene prediction and their expressional tissue-specificities in maize. J. Integr. Plant Biol. 2012, 54, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, S. Dof domain proteins: Plant-specific transcription factors associated with diverse phenomena unique to plants. Plant Cell Physiol. 2004, 45, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- De Grassi, A.; Lanave, C.; Saccone, C. Genome duplication and gene-family evolution: The case of three OXPHOS gene families. Gene 2008, 421, 1–6. [Google Scholar] [CrossRef]
- Kolathur, K.K. Role of promoters in regulating alternative splicing. Gene 2021, 782, 145523. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Johnson, L.S.; Eddy, S.R.; Portugaly, E. Hidden Markov model speed heuristic and iterative HMM search procedure. BMC Bioinform. 2010, 11, 431. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, S. The Dof family of plant transcription factors. Trends Plant Sci. 2002, 7, 555–560. [Google Scholar] [CrossRef]
- Tang, H.; Krishnakumar, V.; Zeng, X.; Xu, Z.; Taranto, A.; Lomas, J.S.; Zhang, Y.; Huang, Y.; Wang, Y.; Yim, W.C.; et al. JCVI: A versatile toolkit for comparative genomics analysis. iMeta 2024, 3, e211. [Google Scholar] [CrossRef]
- Zhao, M.; Li, N.H.; Chen, S.M.; Wu, J.J.; He, S.F.; Zhao, Y.X.; Wang, X.R.; Chen, X.Y.; Zhang, C.Z.; Fang, X.; et al. GmWAK1, Novel Wall-Associated Protein Kinase, Positively Regulates Response of Soybean to Phytophthora sojae Infection. Int. J. Mol. Sci. 2023, 24, 798. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence ID | Number of Amino Acid | Molecular Weight | Theoretical pI | Instability Index | Aliphatic Index | Grand Average of Hydropathicity | Subcellular Localization Prediction |
|---|---|---|---|---|---|---|---|
| PpDof6 | 314 | 34,776.11 | 7.76 | 49.88 | 45.35 | −0.904 | nucl |
| PpDof5 | 359 | 38,910.01 | 8.74 | 57.24 | 51.89 | −0.736 | nucl |
| PpDof3 | 202 | 22,168.98 | 7.61 | 35.43 | 65.89 | −0.444 | cyto |
| PpDof4 | 278 | 30,370.41 | 8.15 | 40.33 | 51.69 | −0.799 | nucl |
| PpDof25 | 317 | 35,045.02 | 4.67 | 51.54 | 57.22 | −0.704 | nucl |
| PpDof18 | 318 | 35,194.72 | 6.78 | 58.19 | 49.4 | −0.845 | nucl |
| PpDof19 | 443 | 46,689.04 | 8.72 | 56.54 | 48.96 | −0.763 | nucl |
| PpDof17 | 362 | 39,227.18 | 8.77 | 51.13 | 57.13 | −0.788 | nucl |
| PpDof20 | 378 | 39,718.75 | 9.36 | 57.83 | 46.24 | −0.665 | nucl |
| PpDof12 | 162 | 18,190.76 | 9.32 | 51.75 | 52.35 | −0.688 | chlo |
| PpDof9 | 278 | 30,732 | 8.76 | 65.06 | 55.83 | −0.721 | nucl |
| PpDof10 | 292 | 31,543.29 | 9.01 | 64.41 | 44.18 | −0.942 | nucl |
| PpDof11 | 376 | 40,988.84 | 8.09 | 54.3 | 49.34 | −0.824 | nucl |
| PpDof15 | 466 | 50,778.37 | 5.35 | 61.72 | 54.68 | −0.855 | nucl |
| PpDof13 | 317 | 34,615.2 | 9 | 50.73 | 56.94 | −0.773 | nucl |
| PpDof16 | 326 | 34,843.02 | 6.59 | 61.69 | 50.03 | −0.706 | nucl |
| PpDof14 | 327 | 36,439.52 | 6.55 | 41.01 | 47.71 | −0.815 | nucl |
| PpDof23 | 339 | 36,002.86 | 8.98 | 58.97 | 56.43 | −0.537 | nucl |
| PpDof24 | 245 | 25,122.85 | 8.51 | 35.98 | 47.43 | −0.448 | nucl |
| PpDof22 | 310 | 33,657.01 | 6.26 | 44.96 | 57.29 | −0.653 | nucl |
| PpDof21 | 474 | 51,461.92 | 7.71 | 55.94 | 66.01 | −0.524 | nucl |
| PpDof8 | 509 | 55,278.07 | 6.05 | 55.43 | 51.38 | −0.871 | nucl |
| PpDof7 | 226 | 24,245.53 | 8.74 | 51.36 | 45.75 | −0.857 | nucl |
| PpDof2 | 283 | 30,140.23 | 5.67 | 46.64 | 44.42 | −0.575 | nucl |
| PpDof1 | 515 | 55,136.67 | 5.88 | 52.7 | 54.25 | −0.707 | nucl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Wang, X.; Yan, J.; Cai, Z.; Zhang, B.; Xu, J.; Ma, R.; Yu, M.; Shen, Z. Systematic Analysis of Dof Gene Family in Prunus persica Unveils Candidate Regulators for Enhancing Cold Tolerance. Int. J. Mol. Sci. 2025, 26, 7509. https://doi.org/10.3390/ijms26157509
Chen Z, Wang X, Yan J, Cai Z, Zhang B, Xu J, Ma R, Yu M, Shen Z. Systematic Analysis of Dof Gene Family in Prunus persica Unveils Candidate Regulators for Enhancing Cold Tolerance. International Journal of Molecular Sciences. 2025; 26(15):7509. https://doi.org/10.3390/ijms26157509
Chicago/Turabian StyleChen, Zheng, Xiaojun Wang, Juan Yan, Zhixiang Cai, Binbin Zhang, Jianlan Xu, Ruijuan Ma, Mingliang Yu, and Zhijun Shen. 2025. "Systematic Analysis of Dof Gene Family in Prunus persica Unveils Candidate Regulators for Enhancing Cold Tolerance" International Journal of Molecular Sciences 26, no. 15: 7509. https://doi.org/10.3390/ijms26157509
APA StyleChen, Z., Wang, X., Yan, J., Cai, Z., Zhang, B., Xu, J., Ma, R., Yu, M., & Shen, Z. (2025). Systematic Analysis of Dof Gene Family in Prunus persica Unveils Candidate Regulators for Enhancing Cold Tolerance. International Journal of Molecular Sciences, 26(15), 7509. https://doi.org/10.3390/ijms26157509