
Isorhamnetin: Reviewing Recent Developments in Anticancer Mechanisms and Nanoformulation-Driven Delivery


Abstract
1. Introduction
Methods
2. Isorhamnetin
2.1. Chemical Structure of Isorhamnetin and Its Significance in Biomedical Applications
2.2. Isorhamnetin: Sources and Its Nutritional Significance
3. Mechanism of Action of Isorhamnetin in Cancers
3.1. Effect of Isorhamnetin on Cell Cycle Regulations
3.1.1. Impact of Isorhamnetin on Cyclins and CDKs
3.1.2. Arrest at G1/S and G2/M Phases
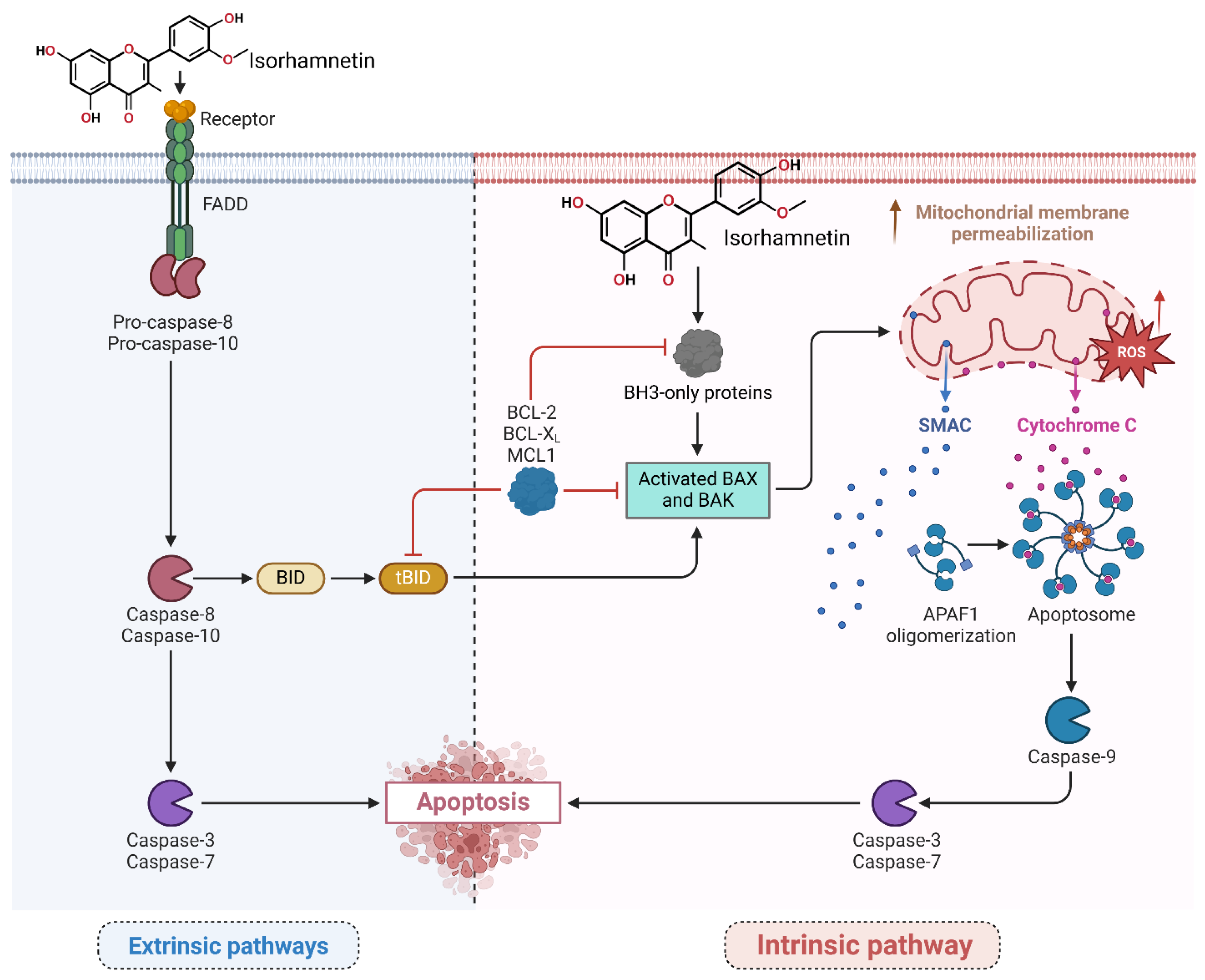
3.2. Apoptosis Induction Pathways
Activation of Intrinsic and Extrinsic Pathways by Isorhamnetin
3.3. Suppression of Angiogenesis and Metastasis

3.3.1. Stage 1: Inhibition of Cancer Cell Invasion and Migration at the Primary Tumor Site
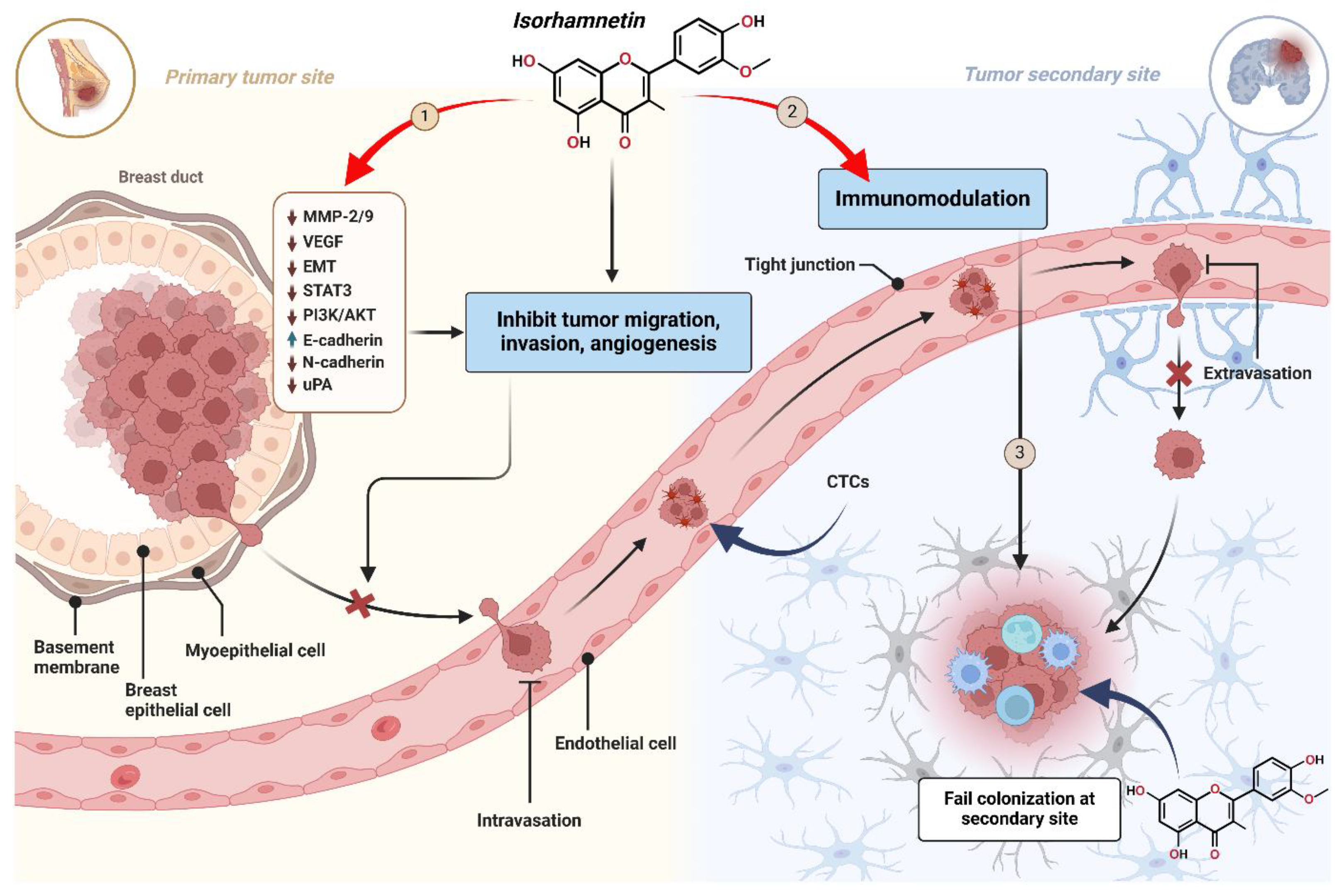
3.3.2. Stage 2: Enhancement of Immune-Mediated Clearance of Circulating Tumor Cells (CTCs) by Isorhamnetin
3.3.3. Stage 3: Prevention of Colonization and Survival at Secondary Tumor Sites by Isorhamnetin
3.4. Antioxidant and Anti-Inflammatory Effects of Isorhamnetin
3.4.1. Reduction in Endogenous ROS and RNS Levels by Isorhamnetin
3.4.2. Kappa-Light-Chain-Enhancer of Activated B Cells (NF-κB) and Cyclooxygenase-2 (COX-2) Inhibition by Isorhamnetin
3.5. Activation of the p53 Pathway by Isorhamnetin
3.6. Activation of MAPK Pathway by Isorhamnetin
3.7. Modulation of Tumor Microenvironment and Immune Response by Isorhamnetin
3.7.1. Impact of Isorhamnetin on CAFs
3.7.2. Modulation of Immune Response by Isorhamnetin
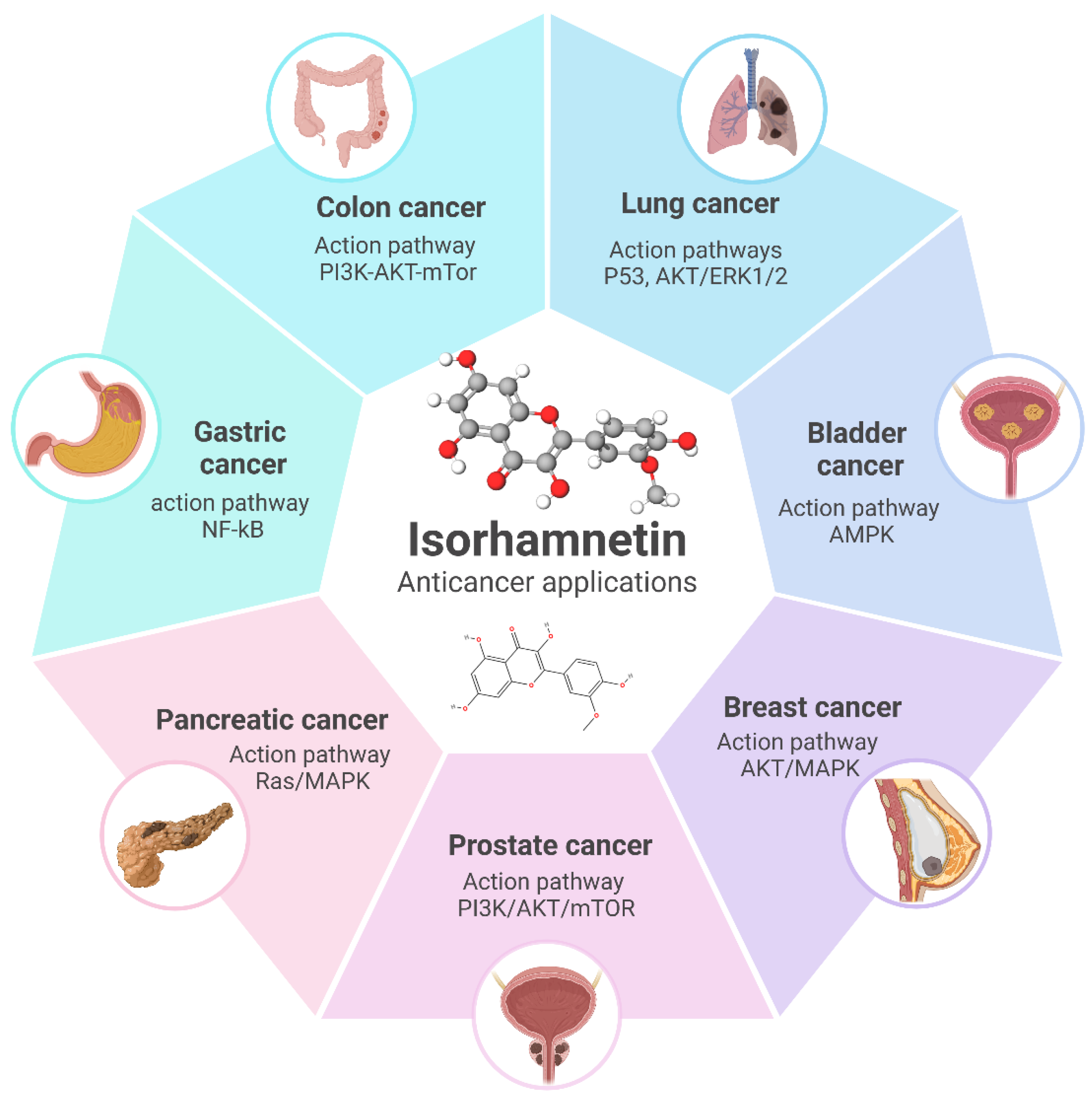
4. Cancer Type-Specific Effects of Isorhamnetin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Targeted Pathway | Isorhamnetin Concentration | Main Findings | Ref. |
|---|---|---|---|---|
| Breast cancer (in vitro) | Akt/mTOR and MEK/ERK signaling pathways | IC50: ~10 µM | ↑ Apoptosis, ↑ Bax, ↑ cleaved caspase-3 ↓ Proliferation, ↓ Bcl-2, ↓ Akt, ↓, mTOR, ↓ MEK1/2, and ↓ ERK1/2 signaling | [288] |
| Canine mammary tumors (in vitro and in vivo) | EGFR-STAT3-PD-L1 signaling pathway | 10 µM, 20 µM, and 40 µM | ↑ Caspase-3, ↑ Apoptosis ↓ EGFR, ↓ STAT3, ↓ PD-L1, ↓ Migration, ↓ Invasion, ↓ Ki-67 | [118] |
| Gastric cancer (in vitro and in vivo) | PI3K/Akt signaling pathway | IC50: ~50 µM | ↑ Mitochondrial apoptosis, ↑ Caspase-3, ↑ Apoptosis. ↓ Proliferation, ↓ Invasion, ↓ Metastasis, ↓ SRC, ↓ AKT1, ↓ EGFR, ↓ PI3K/Akt, | [289] |
| Lung cancer (in vitro) | PI3K-Akt signaling pathway | - | ↑ Apoptosis, ↑ G1 Arrest ↓ Migration, ↓ Invasion, ↓ p-PI3K, ↓ p-AKT, ↓ PI3K/Akt pathway | [290] |
| Prostate Cancer (in vitro) | PI3K/Akt/mTOR signaling pathway | 5 µM, 10 µM, and 20 µM | ↑ Apoptosis (intrinsic) ↓ Proliferation, ↓ Migration, ↓ Invasion, ↓ PI3K/Akt/mTOR, ↑ E-cadherin, ↓ Vimentin, ↓ N-cadherin, ↓ MMP-2/9 | [85] |
| Hepatocellular carcinoma (in vivo) | Akt, MAPKs, and Nrf2 signaling pathways; PPAR-γ activation | 100 mg/kg body weight (in vivo dose) | ↓ Pro-inflammatory cytokines, ↓ Nrf2, ↓ Akt, ↓ MAPK, ↑ PPAR-γ, ↑ Autophagy, ↑ Apoptosis, ↑ G1 Arrest | [291] |
| Melanoma (in vitro) | - | IC50: 8.26 μg/ml | ↑ Apoptosis, ↑ Sub-G0/G1 Arrest, ↓ S Phase, ↓ G2/M Phase, ↓ BCL-2, ↑ Bax, ↑ Caspase-3/9, ↑ DNA Fragmentation | [292] |
| Lung cancer (in vitro) | NF-κB signaling pathway and IL-13-mediated apoptotic mechanisms | 20 µM | ↑ Radiosensitivity, ↓ NF-κB, ↑ Apoptosis, ↑ Mitochondrial dysfunction, ↑ IL-13 | [293] |
| Melanoma (in vitro and in vivo) | PI3K/Akt and NF-κB pathways, with involvement of PFKFB4 | 10–100 μmol/L | ↓ Proliferation, ↓ Migration, ↓ Colony formation, ↑ Bax, ↑ Caspase-3, ↓ BCL-2, ↓ PI3K/Akt, ↓ NF-κB, ↓ PFKFB4, ↑ Apoptosis | [294] |
| Oral cancer (in vitro and in vivo) | Glycolysis signaling pathway, explicitly targeting HK2 | 0.1–30 μM | ↓ Proliferation, ↓ Glycolysis, ↓ HK2, ↓ Ki-67, ↓ Tumor growth, ↔ PFK, ↔ PKM2 | [246] |
| Stomach adenocarcinoma (in vitro) | MAPK/mTOR signaling pathway | 20 μM, 30 μM, 40 μM, and 60 μM | ↓ Proliferation, ↓ Migration, ↓ Colony formation, ↑ Apoptosis, ↑ G2/M Arrest, ↓ MAPK14, ↓ MAPK/mTOR, EMT modulation | [215] |
| Colorectal adenocarcinoma (in vitro and in vivo) | Apoptosis (Caspase-9 and Bcl-2) | - | ↑ Apoptosis, ↑ ROS, ↑ G0/G1 Arrest, ↓ Tumor growth, ↑ Caspase-9, ↑ Hdac11, ↑ Bai1, ↓ Bcl-2 | [295] |
| Ovarian cancer (in vitro and in vivo) | ESR1-mediated signaling pathways. | 5 μM, 10 μM, 15 μM, and 20 μM | ↓ Proliferation, ↓ Migration, ↓ Invasion, ↓ Ki-67, ↓ MMP-2, ↓ MMP-9, ↓ Tumor volume/weight, Targeting ESR1 | [296] |
| Gastric cancer (in vitro and in vivo) | Mitochondria-dependent apoptosis pathway | 20 µM | ↑ Caspase-3, ↑ Cytochrome c, ↓ Mitochondrial membrane potential, ↑ ROS, ↑ Mitochondrial dysfunction, ↓ Migration, ↓ Proliferation, ↓ Tumor size (time & dose dependent) | [297] |
| Bladder cancer (in vitro and in vivo) | PPARγ/PTEN/AKT signaling pathway | 10 μM, 50 μM, and 100 μM (in vitro); 5 mg/kg (in vivo) | ↓ Proliferation, ↓ Tumorigenicity, ↓ G0/G1 → S transition, ↑ PPARγ/PTEN, ↓ AKT, ↓ CA9, ↑ Apoptosis, ↓ Tumor growth, ↓ Ki67 | [155] |
| Colorectal cancer (in vitro) | ROS-mediated apoptosis and anti-inflammatory pathways | 5–150 μM | ↓ Mitochondrial, ↓ Metabolic, ↓ Lysosomal activity, ↑ ROS, ↓ IL-8, ↓ Proliferation, ↑ Apoptosis, ↑ Cell cycle disruption (≥100 μM) | [298] |
| Lung cancer (in vitro) | Akt/ERK-mediated epithelial-to-mesenchymal transition (EMT) | 2.5, 5, and 10 μM | ↓ Proliferation, ↓ Adhesion, ↓ Invasion, ↓ Migration, ↓ MMP-2/9, ↑ E-cadherin, ↓ N-cadherin, ↓ Vimentin, ↓ Snail, ↓ Akt/ERK, EMT reversal, ↓ Metastasis | [96] |
| Breast cancer (in vitro) | p38 MAPK and STAT3 signaling pathway | - | ↓ Adhesion, ↓ Migration, ↓ Invasion, ↓ MMP-2/9, ↓ p38 MAPK, ↓ STAT3, ↔ ERK1/2, ↔ JNK, ↔ uPA | [98] |
| Breast cancer (in vitro and in vivo) | AMPK/mTOR/p70S6K signaling, ROS generation, G2/M cell cycle arrest, apoptosis pathway | 10, 20, 30, 50 μM | ↑ Apoptosis, ↑ G2/M Arrest, ↓ CDK1/Cyclin B1, ↑ ROS (×6.78 times), ↑ DNA damage, ↑ AMPK, ↓ mTOR/p70S6K, ↓ Proliferation | [62] |
| Endometrial cancer (in vitro and in vivo) | Mitochondrial dysfunction, cell death receptor pathway, endoplasmic reticulum (ER) stress pathway, UPR response, MMP2/9 expression | 0 μM, 20 μM, 40 μM, and 60 μM | ↑ Apoptosis (mitochondrial & death receptor), ↑ ER stress pathway, ↓ MMP-2/9, ↓ Metastasis, ↓ Tumor growth | [117] |
| Breast cancer (in vitro) | Akt/mTOR and MEK/ERK signaling pathways and cell cycle inhibition | 100, 33.3, 11.1, 3.7, 1.2, 0.4 and 0 µM | ↓ Proliferation, ↑ Apoptosis, ↓ Akt/mTOR, ↓ MEK/ERK, ↑ Akt & MEK activation (EGF reversal) | [288] |
| Colorectal cancer (in vitro) | HIF-1α, ROS, Nrf2, glucose transporter 1, lactate dehydrogenase A, pyruvate dehydrogenase kinase 1, heme oxygenase-1, COX-2 | 3, 10, 30, 69 µM | ↓ HIF-1α (CoCl2, hypoxia, H2O2-induced), ↓ Hypoxia genes, ↓ ROS, ↓ Migration, ↓ Invasion, ↑ Nrf2, ↑ Antioxidant proteins | [299] |
| Gastric cancer (in vitro) | PI3K–AKT–mTOR signaling pathway | 20, 40, 80, 160, and 320 µM/L | ↓ Autophagy (under hypoxia), ↓ Proliferation, ↓ Mitochondrial membrane potential, ↑ Mitochondrial apoptosis, ↓ PI3K/Akt/mTOR, ↑ Apoptosis (vs. 3-MA) | [246] |
| Colon cancer (in vitro) | Apoptosis, cell cycle regulation, mitochondrial | 50 µg/mL and 100 µg/mL | ↑ G2/M Arrest, ↑ Bax/Bcl-2 ratio, ↑ Apoptosis (mitochondrial), ↑ ROS, ↑ Caspase-dependent cell death | [274] |
| Colon cancer (In vitro and in vivo) | Apoptosis, Hsp70 inhibition | - | ↑ Apoptosis, ↓ Hsp70, ↑ Apaf1, ↑ Caspase-3/9, ↓ Tumor growth (colon cancer model) | [267] |
| Gastric cancer (In vitro and in silico) | MAPK/mTOR signaling pathway | 20 µM and 30 µM | ↓ Proliferation, ↓ Migration, ↑ Apoptosis, ↑ MAPK/mTOR activation (apoptosis induction) | [215] |
5. Synergistic and Adjuvant Roles of Isorhamnetin for Biomedical Applications
| Cancer | Combination | Main Findings in the Combination Treatment | Study Target | Ref. |
|---|---|---|---|---|
| No | Isorhamnetin + escitalopram |
| Antidepression (in vivo) | [54] |
| Yes | Isorhamnetin + carboplatin + cisplatin |
| Lung cancer (in vitro) | [306] |
| No | Isorhamnetin + cisplatin |
| Kidney protection (in vitro, in vivo) | [38] |
| Yes | Isorhamnetin + radiotherapy |
| Lung cancer radiosensitization (in vitro) | [293] |
| No | Quercetin + Isorhamnetin + Quercetin-3-glucuronide |
| anti-inflammatory effects (in vitro) | [327] |
| Yes | Isorhamnetin + Isorhamnetin-3-glucuronide + Quercetin |
| Breast cancer cytotoxic effects (in vitro) | [268] |
| Yes | Isorhamnetin + Doxorubicin |
| Breast cancer (in vitro, in vivo) | [62] |
| No | Combination + sildenafil (in vivo) |
| Pulmonary arterial hypertension (PAH) | [247] |
6. Advances in Delivery Systems for Isorhamnetin for Anticancer Applications
6.1. Delivery Methods
6.2. Advantages of Isorhamnetin-Based Nanoformulation in Anticancer Applications
6.3. Challenges
7. Future Perspectives
7.1. Role of Emerging Technologies in Isorhamnetin Research
7.2. Potential for Integration into Precision Oncology
7.3. Limitations and Future Research Directions
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASR | Age-standardized rate |
| CDK | Cyclin-dependent kinase |
| EMT | Epithelial–mesenchymal transition |
| MMP | Matrix metalloproteinase |
| OSCC | Oral squamous cell carcinoma |
| VEGF | Vascular endothelial growth factor |
| ROS | Reactive oxygen species |
| RNS | Reactive nitrogen species |
| SOD | Superoxide dismutase |
| MAPK | Mitogen-activated protein kinase |
| AMPK/mTOR | AMP-activated protein kinase/Mammalian Target of Rapamycin |
| pRb | Retinoblastoma Protein |
| MCL | Myeloid Cell Leukemia |
| Fas | First apoptosis signal receptor |
| TRAIL | Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand |
| FADD | Fas-Associated protein with Death Domain |
| BID | BH3 Interacting-domain death agonist |
| APAF | Apoptotic Protease Activating Factor |
| SMAC | Second Mitochondria-derived Activator of Caspases |
| ACSL | Acyl-CoA Synthetase Long-Chain Family Member |
| PTGS | Prostaglandin-Endoperoxide Synthase |
| DSS | Dextran Sulfate Sodium |
| VRAP | VEGF Receptor-Associated Protein |
| ERK | Extracellular Signal-Regulated Kinase |
| JNK | c-Jun N-terminal Kinase |
| uPA | Urokinase-type Plasminogen Activator |
| NSCLC | Non-Small-Cell Lung Cancer |
| COLA1 | Collagen Type I Alpha 1 |
| α-SMA | Alpha-Smooth Muscle Actin |
| HIF | Hypoxia-Inducible Factor |
| CTCs | Circulating Tumor Cells |
| ECM | Extracellular Matrix |
| TME | Tumor microenvironment |
| NK | Natural killer |
| ICD | Immunogenic cell death |
| DAMP | damage-associated molecular pattern |
| CAF | Cancer-associated fibroblast |
| TAM | Tumor-associated macrophage |
| PFN | Perforin |
| IFN | Interferon |
| TNF | Tumor necrosis factor |
| HO | Heme oxygenase |
| PAH | Pulmonary arterial hypertension |
| NPs | Nanoparticles |
| EPR | Enhanced permeability and retention |
| GPCR | G-protein coupled receptor |
| RTK | Receptor tyrosine kinase |
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer Statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, H.; Tan, L.; Siu, K.T.H.; Guan, X.-Y. Exploring Treatment Options in Cancer: Tumor Treatment Strategies. Signal Transduct. Target. Ther. 2024, 9, 175. [Google Scholar] [CrossRef]
- Kamrani, A.; Hosseinzadeh, R.; Shomali, N.; Heris, J.A.; Shahabi, P.; Mohammadinasab, R.; Sadeghvand, S.; Ghahremanzadeh, K.; Sadeghi, M.; Akbari, M. New Immunotherapeutic Approaches for Cancer Treatment. Pathol.—Res. Pract. 2023, 248, 154632. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Cianca, S.I.; Romero-Castillo, C.; Gálvez-Romero, J.L.; Juárez, Z.N.; Hernández, L.R. Antioxidant and Anti-Inflammatory Compounds from Edible Plants with Anti-Cancer Activity and Their Potential Use as Drugs. Molecules 2023, 28, 1488. [Google Scholar] [CrossRef] [PubMed]
- Mucha, P.; Skoczyńska, A.; Małecka, M.; Hikisz, P.; Budzisz, E. Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes. Molecules 2021, 26, 4886. [Google Scholar] [CrossRef]
- Ojo, O.A.; Ojo, A.B.; Barnabas, M.; Iyobhebhe, M.; Elebiyo, T.C.; Evbuomwan, I.O.; Michael, T.; Ajiboye, B.O.; Oyinloye, B.E.; Oloyede, O.I. Phytochemical Properties and Pharmacological Activities of the Genus Pennisetum: A Review. Sci. Afr. 2022, 16, e01132. [Google Scholar] [CrossRef]
- Pavithra, R.; Khan, M.R.; Khan, M.S. Recent Advancements in Natural Compounds for Cancer Therapy and Prevention. Phytochem. Rev. 2024, 23, 1835–1859. [Google Scholar] [CrossRef]
- Ali Abdalla, Y.O.; Subramaniam, B.; Nyamathulla, S.; Shamsuddin, N.; Arshad, N.M.; Mun, K.S.; Awang, K.; Nagoor, N.H. Natural Products for Cancer Therapy: A Review of Their Mechanism of Actions and Toxicity in the Past Decade. J. Trop. Med. 2022, 2022, 5794350. [Google Scholar] [CrossRef]
- Rana, J.N.; Mumtaz, S. Prunin: An Emerging Anticancer Flavonoid. Int. J. Mol. Sci. 2025, 26, 2678. [Google Scholar] [CrossRef]
- Han, Z.; Li, C.; Liu, G. Recent Advances in the Extraction, Purification and Analytical Techniques for Flavonoids from Plants: Taking Hawthorn as an Example. J. Food Compos. Anal. 2025, 141, 107372. [Google Scholar] [CrossRef]
- Li, W.; Wu, J.; Zhang, J.; Wang, J.; Xiang, D.; Luo, S.; Li, J.; Liu, X. Puerarin-Loaded PEG-PE Micelles with Enhanced Anti-Apoptotic Effect and Better Pharmacokinetic Profile. Drug Deliv. 2018, 25, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Huang, J.; Li, H.; Zhao, D.; Liu, Z.; Zhu, L.; Zhang, Z.; Peng, W. Quercetin: A Promising Therapy for Diabetic Encephalopathy through Inhibition of Hippocampal Ferroptosis. Phytomedicine 2024, 126, 154887. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, R.; Mukhopadhyay, C. Chapter 14—Green Synthesis of Bioactive Flavonoids as Cardioprotective and Anticancer Drug Agents. In Advances in Green and Sustainable Chemistry, 2nd ed.; Banik, B.K., Ed.; Elsevier: Amsterdam, The Netherlands, 2024; Volume 1, pp. 305–343. ISBN 978-0-443-16166-7. [Google Scholar]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Wang, Y.; Xu, J.; Zheng, Y.; Zhou, W.; Wang, Y.; Luo, C. Precisely Tailoring Molecular Structure of Doxorubicin Prodrugs to Enable Stable Nanoassembly, Rapid Activation, and Potent Antitumor Effect. Pharmaceutics 2024, 16, 1582. [Google Scholar] [CrossRef]
- Cristina Marcarini, J.; Ferreira Tsuboy, M.S.; Cabral Luiz, R.; Regina Ribeiro, L.; Beatriz Hoffmann-Campo, C.; Ségio Mantovani, M. Investigation of Cytotoxic, Apoptosis-Inducing, Genotoxic and Protective Effects of the Flavonoid Rutin in HTC Hepatic Cells. Exp. Toxicol. Pathol. 2011, 63, 459–465. [Google Scholar] [CrossRef]
- Gong, G.; Guan, Y.-Y.; Zhang, Z.-L.; Rahman, K.; Wang, S.-J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A Review of Pharmacological Effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef]
- Shi, H.; He, J.; Li, X.; Han, J.; Wu, R.; Wang, D.; Yang, F.; Sun, E. Isorhamnetin, the Active Constituent of a Chinese Herb Hippophae rhamnoides L., Is a Potent Suppressor of Dendritic-Cell Maturation and Trafficking. Int. Immunopharmacol. 2018, 55, 216–222. [Google Scholar] [CrossRef]
- Biswas, P.; Kaium, M.A.; Islam Tareq, M.M.; Tauhida, S.J.; Hossain, M.R.; Siam, L.S.; Parvez, A.; Bibi, S.; Hasan, M.H.; Rahman, M.M.; et al. The Experimental Significance of Isorhamnetin as an Effective Therapeutic Option for Cancer: A Comprehensive Analysis. Biomed. Pharmacother. 2024, 176, 116860. [Google Scholar] [CrossRef]
- Khalil, M.N.A.; Afifi, S.M.; Eltanany, B.M.; Pont, L.; Benavente, F.; El-Sonbaty, S.M.; Sedeek, M.S. Assessment of the Effect of Drying on Brassica Greens via a Multiplex Approach Based on LC-QTOF-MS/MS, Molecular Networking, and Chemometrics along with Their Antioxidant and Anticancer Activities. Food Res. Int. 2024, 180, 114053. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) Isorhamnetin. PubChem Compound Summary for CID 5281654, Isorhamnetin. 2025. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Isorhamnetin (accessed on 10 July 2025).
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.-K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and Bioavailability of Quercetin Glycosides in Humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef]
- Park, Y.-K.; Lee, C.Y. Identification of Isorhamnetin 4‘-Glucoside in Onions. J. Agric. Food Chem. 1996, 44, 34–36. [Google Scholar] [CrossRef]
- Mubashar Sabir, S.; Khan, M.F.; Rocha, J.B.T.; Boligon, A.A.; Athayde, M.L. Phenolic Profile, Antioxidant Activities and Genotoxic Evaluations of Calendula officinalis. J. Food Biochem. 2015, 39, 316–324. [Google Scholar] [CrossRef]
- Kalai, F.Z.; Boulaaba, M.; Ferdousi, F.; Isoda, H. Effects of Isorhamnetin on Diabetes and Its Associated Complications: A Review of In Vitro and In Vivo Studies and a Post Hoc Transcriptome Analysis of Involved Molecular Pathways. Int. J. Mol. Sci. 2022, 23, 704. [Google Scholar] [CrossRef]
- Abudalo, R.; Gammoh, O.; Altaber, S.; Bseiso, Y.; Qnais, E.; Wedyan, M.; Oqal, M.; Alqudah, A. Mitigation of Cisplatin-Induced Cardiotoxicity by Isorhamnetin: Mechanistic Insights into Oxidative Stress, Inflammation, and Apoptosis Modulation. Toxicol. Rep. 2024, 12, 564–573. [Google Scholar] [CrossRef]
- Hu, D.; Wang, H.-J.; Yu, L.-H.; Guan, Z.-R.; Jiang, Y.-P.; Hu, J.-H.; Yan, Y.-X.; Zhou, Z.-H.; Lou, J.-S. The Role of Ginkgo Folium on Antitumor: Bioactive Constituents and the Potential Mechanism. J. Ethnopharmacol. 2024, 321, 117202. [Google Scholar] [CrossRef]
- Martiniakova, M.; Penzes, N.; Biro, R.; Sarocka, A.; Kovacova, V.; Mondockova, V.; Ciernikova, S.; Omelka, R. Sea Buckthorn and Its Flavonoids Isorhamnetin, Quercetin, and Kaempferol Favorably Influence Bone and Breast Tissue Health. Front. Pharmacol. 2024, 15, 1462823. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chi, G.; Shen, B.; Tian, Y.; Feng, H. Isorhamnetin Ameliorates LPS-Induced Inflammatory Response through Downregulation of NF-ΚB Signaling. Inflammation 2016, 39, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, N.; Fan, F.; Geng, Z.; Zhao, K.; Wang, J.; Zhang, Y.; Tang, C.; Wang, X.; Meng, X. Tibetan Medicine Duoxuekang Capsule Ameliorates High-Altitude Polycythemia Accompanied by Brain Injury. Front. Pharmacol. 2021, 12, 680636. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, H.; Shen, C.; Dong, X.; Li, J.; Liu, J. Effects of Isorhamnetin on Liver Injury in Heat Stroke-Affected Rats under Dry-Heat Environments via Oxidative Stress and Inflammatory Response. Sci. Rep. 2024, 14, 7476. [Google Scholar] [CrossRef]
- Xu, H.; Yu, S.; Lin, C.; Dong, D.; Xiao, J.; Ye, Y.; Wang, M. Roles of Flavonoids in Ischemic Heart Disease: Cardioprotective Effects and Mechanisms against Myocardial Ischemia and Reperfusion Injury. Phytomedicine 2024, 126, 155409. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, M.; Farzaei, M.H.; Kiani, S.; Khodarahmi, R. Immunomodulatory; Anti-Inflammatory/Antioxidant Effects of Polyphenols: A Comparative Review on the Parental Compounds and Their Metabolites. Food Rev. Int. 2021, 37, 759–811. [Google Scholar] [CrossRef]
- González-Arceo, M.; Gomez-Lopez, I.; Carr-Ugarte, H.; Eseberri, I.; González, M.; Cano, M.P.; Portillo, M.P.; Gómez-Zorita, S. Anti-Obesity Effects of Isorhamnetin and Isorhamnetin Conjugates. Int. J. Mol. Sci. 2023, 24, 299. [Google Scholar] [CrossRef]
- Dayem, A.A.; Choi, H.Y.; Kim, Y.B.; Cho, S.-G. Antiviral Effect of Methylated Flavonol Isorhamnetin against Influenza. PLoS ONE 2015, 10, e0121610. [Google Scholar] [CrossRef]
- Yang, B.; Li, X.-P.; Ni, Y.-F.; Du, H.-Y.; Wang, R.; Li, M.-J.; Wang, W.-C.; Li, M.-M.; Wang, X.-H.; Li, L.; et al. Protective Effect of Isorhamnetin on Lipopolysaccharide-Induced Acute Lung Injury in Mice. Inflammation 2016, 39, 129–137. [Google Scholar] [CrossRef]
- Wang, L.; Xie, Y.; Xiao, B.; He, X.; Ying, G.; Zha, H.; Yang, C.; Jin, X.; Li, G.; Ping, L.; et al. Isorhamnetin Alleviates Cisplatin-Induced Acute Kidney Injury via Enhancing Fatty Acid Oxidation. Free Radic. Biol. Med. 2024, 212, 22–33. [Google Scholar] [CrossRef]
- Lei, J.; Yang, J.; Bao, C.; Lu, F.; Wu, Q.; Wu, Z.; Lv, H.; Zhou, Y.; Liu, Y.; Zhu, N.; et al. Isorhamnetin: What Is the in Vitro Evidence for Its Antitumor Potential and Beyond? Front. Pharmacol. 2024, 15, 1309178. [Google Scholar] [CrossRef]
- Wang, H.; Chen, L.; Yang, B.; Du, J.; Chen, L.; Li, Y.; Guo, F. Structures, Sources, Identification/Quantification Methods, Health Benefits, Bioaccessibility, and Products of Isorhamnetin Glycosides as Phytonutrients. Nutrients 2023, 15, 1947. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Akobirshoeva, A.; Zilfikarov, I.N.; Vennos, C. Isorhamnetin and Quercetin Derivatives as Anti-Acetylcholinesterase Principles of Marigold (Calendula officinalis) Flowers and Preparations. Int. J. Mol. Sci. 2017, 18, 1685. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I. New Isorhamnetin Glycosides and Other Phenolic Compounds from Calendula officinalis. Chem. Nat. Compd. 2013, 49, 833–840. [Google Scholar] [CrossRef]
- Bhagwat, S.; Haytowitz, D.B. USDA Database for the Flavonoid Content of Selected Foods, Release 3.3 (March 2018); US Department of Agriculture: Beltsville, MD, USA, 2022. [CrossRef]
- Arts, I.C.W.; Hollman, P.C.H. Polyphenols and Disease Risk in Epidemiologic Studies2, 3. Am. J. Clin. Nutr. 2005, 81, 317S–325S. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xi, Y.; Jiang, W. Protective Roles of Flavonoids and Flavonoid-Rich Plant Extracts against Urolithiasis: A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 2125–2135. [Google Scholar] [CrossRef] [PubMed]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Törrönen, A.R. Identification and Quantification of Phenolic Compounds in Berries of Fragaria and Rubus Species (Family Rosaceae). J. Agric. Food Chem. 2004, 52, 6178–6187. [Google Scholar] [CrossRef]
- Kim, B.-G.; Sung, S.H.; Jung, N.R.; Chong, Y.; Ahn, J.-H. Biological Synthesis of Isorhamnetin 3-O-Glucoside Using Engineered Glucosyltransferase. J. Mol. Catal. B Enzym. 2010, 63, 194–199. [Google Scholar] [CrossRef]
- Schieber, A.; Keller, P.; Streker, P.; Klaiber, I.; Carle, R. Detection of Isorhamnetin Glycosides in Extracts of Apples (Malus domestica Cv. “Brettacher”) by HPLC-PDA and HPLC-APCI-MS/MS. Phytochem. Anal. 2002, 13, 87–94. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M. Phenolic Compounds and Chromatographic Profiles of Pear Skins (Pyrus spp.). J. Agric. Food Chem. 2008, 56, 9094–9101. [Google Scholar] [CrossRef]
- Lee, J.; Mitchell, A.E. Quercetin and Isorhamnetin Glycosides in Onion (Allium cepa L.): Varietal Comparison, Physical Distribution, Coproduct Evaluation, and Long-Term Storage Stability. J. Agric. Food Chem. 2011, 59, 857–863. [Google Scholar] [CrossRef]
- Singh, J.; Jayaprakasha, G.K.; Patil, B.S. Extraction, Identification, and Potential Health Benefits of Spinach Flavonoids: A Review. In Advances in Plant Phenolics: From Chemistry to Human Health; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2018; Volume 1286, pp. 107–136. ISBN 9780841232969. [Google Scholar]
- Xu, S.L.; Choi, R.C.Y.; Zhu, K.Y.; Leung, K.-W.; Guo, A.J.Y.; Bi, D.; Xu, H.; Lau, D.T.W.; Dong, T.T.X.; Tsim, K.W.K. Isorhamnetin, A Flavonol Aglycone from Ginkgo biloba L., Induces Neuronal Differentiation of Cultured PC12 Cells: Potentiating the Effect of Nerve Growth Factor. Evid.-Based Complement. Altern. Med. 2012, 2012, 278273. [Google Scholar] [CrossRef]
- Mohamed Sharif, K.O.; Tufekci, E.F.; Ustaoglu, B.; Altunoglu, Y.C.; Zengin, G.; Llorent-Martínez, E.J.; Guney, K.; Baloglu, M.C. Anticancer and Biological Properties of Leaf and Flower Extracts of Echinacea purpurea (L.) Moench. Food Biosci. 2021, 41, 101005. [Google Scholar] [CrossRef]
- Gammoh, O.; Qnais, E.Y.; Athamneh, R.Y.; Al-Jaidi, B.; Al-Tawalbeh, D.; Altaber, S.; Alqudah, A.; Aljabali, A.A.A.; Tambuwala, M.M. Unraveling the Potential of Isorhamnetin as an Adjuvant in Depression Treatment with Escitalopram. Curr. Issues Mol. Biol. 2023, 45, 7668–7679. [Google Scholar] [CrossRef]
- Guo, W.; Wang, W.; Lei, F.; Zheng, R.; Zhao, X.; Gu, Y.; Yang, M.; Tong, Y.; Wang, Y. Identifying the Main Components and Mechanisms of Action of Artemisia annua L. in the Treatment of Endometrial Cancer Using Network Pharmacology. ACS Omega 2024, 9, 8055–8066. [Google Scholar] [CrossRef]
- Park, C.; Cha, H.-J.; Choi, E.O.; Lee, H.; Hwang-Bo, H.; Ji, S.Y.; Kim, M.Y.; Kim, S.Y.; Hong, S.H.; Cheong, J.; et al. Isorhamnetin Induces Cell Cycle Arrest and Apoptosis Via Reactive Oxygen Species-Mediated AMP-Activated Protein Kinase Signaling Pathway Activation in Human Bladder Cancer Cells. Cancers 2019, 11, 1494. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Song, Y.; Wang, J.; Wang, T.; Yang, L.; Shi, Y.; Song, B.; Yu, Z. Isorhamnetin Inhibits Hypertrophic Scar Formation through TGF-β1/Smad and TGF-β1/CREB3L1 Signaling Pathways. Heliyon 2024, 10, e33802. [Google Scholar] [CrossRef] [PubMed]
- Diehl, F.F.; Sapp, K.M.; Vander Heiden, M.G. The Bidirectional Relationship between Metabolism and Cell Cycle Control. Trends Cell Biol. 2024, 34, 136–149. [Google Scholar] [CrossRef]
- Stallaert, W.; Taylor, S.R.; Kedziora, K.M.; Taylor, C.D.; Sobon, H.K.; Young, C.L.; Limas, J.C.; Varblow Holloway, J.; Johnson, M.S.; Cook, J.G.; et al. The Molecular Architecture of Cell Cycle Arrest. Mol. Syst. Biol. 2022, 18, e11087. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.J.; Kabeer, A.; Abbas, Z.; Siddiqui, H.A.; Calina, D.; Sharifi-Rad, J.; Cho, W.C. Interplay of Oxidative Stress, Cellular Communication and Signaling Pathways in Cancer. Cell Commun. Signal. 2024, 22, 7. [Google Scholar] [CrossRef]
- Lu, R.; Liu, J.; Thakur, K.; Cao, H.; Mejuto, J.C.; simal Gandara, J.; Zhang, J.-G. Protopanaxadiol Triggers G0/G1 Cell Cycle Arrest and Apoptosis in Human Cervical Cancer HeLa Cells through the PPER Pathway. Food Biosci. 2024, 62, 105388. [Google Scholar] [CrossRef]
- Yang, T.; Xiao, Y.; Liu, S.; Luo, F.; Tang, D.; Yu, Y.; Xie, Y. Isorhamnetin Induces Cell Cycle Arrest and Apoptosis by Triggering DNA Damage and Regulating the AMPK/MTOR/P70S6K Signaling Pathway in Doxorubicin-Resistant Breast Cancer. Phytomedicine 2023, 114, 154780. [Google Scholar] [CrossRef]
- Ganbold, M.; Louphrasitthiphol, P.; Miyamoto, T.; Miyazaki, Y.; Oda, T.; Tominaga, K.; Isoda, H. Isorhamnetin Exerts Anti-Proliferative Effect on Cancer-Associated Fibroblasts by Inducing Cell Cycle Arrest. Biomed. Pharmacother. 2025, 185, 117954. [Google Scholar] [CrossRef]
- Wang, J.-L.; Quan, Q.; Ji, R.; Guo, X.-Y.; Zhang, J.-M.; Li, X.; Liu, Y.-G. Isorhamnetin Suppresses PANC-1 Pancreatic Cancer Cell Proliferation through S Phase Arrest. Biomed. Pharmacother. 2018, 108, 925–933. [Google Scholar] [CrossRef]
- Deshpande, A.; Sicinski, P.; Hinds, P.W. Cyclins and Cdks in Development and Cancer: A Perspective. Oncogene 2005, 24, 2909–2915. [Google Scholar] [CrossRef]
- Chen, Q.; Song, S.; Wang, Z.; Shen, Y.; Xie, L.; Li, J.; Jiang, L.; Zhao, H.; Feng, X.; Zhou, Y.; et al. Isorhamnetin Induces the Paraptotic Cell Death through ROS and the ERK/MAPK Pathway in OSCC Cells. Oral Dis. 2021, 27, 240–250. [Google Scholar] [CrossRef]
- Huang, M.-F.; Wang, Y.-X.; Chou, Y.-T.; Lee, D.-F. Therapeutic Strategies for RB1-Deficient Cancers: Intersecting Gene Regulation and Targeted Therapy. Cancers 2024, 16, 1558. [Google Scholar] [CrossRef]
- Chang, C.-H.; Liu, F.; Militi, S.; Hester, S.; Nibhani, R.; Deng, S.; Dunford, J.; Rendek, A.; Soonawalla, Z.; Fischer, R.; et al. The PRb/RBL2-E2F1/4-GCN5 Axis Regulates Cancer Stem Cell Formation and G0 Phase Entry/Exit by Paracrine Mechanisms. Nat. Commun. 2024, 15, 3580. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Shi, J.; Zhu, J. Cellular and Molecular Events in Colorectal Cancer: Biological Mechanisms, Cell Death Pathways, Drug Resistance and Signalling Network Interactions. Discov. Oncol. 2024, 15, 294. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Pullaguri, N.; Rath, S.N.; Bajaj, A.; Sahu, V.; Ealla, K.K.R. Dysregulation of Calcium Homeostasis in Cancer and Its Role in Chemoresistance. Cancer Drug Resist. 2024, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Guo, S.; Guo, J.; Du, Q.; Wu, C.; Wu, Y.; Zhang, Y. Cell Death Pathways: Molecular Mechanisms and Therapeutic Targets for Cancer. MedComm 2024, 5, e693. [Google Scholar] [CrossRef]
- Rassu, G.; Vlčková, H.K.; Giunchedi, P.; Dias, P.; Cossu, M.; Pourová, J.; Harčárová, P.; Lomozová, Z.; Nováková, L.; Gavini, E.; et al. A Water-Soluble Preparation for Intravenous Administration of Isorhamnetin and Its Pharmacokinetics in Rats. Chem. Biol. Interact. 2024, 396, 111064. [Google Scholar] [CrossRef]
- Mustafa, M.; Ahmad, R.; Tantry, I.Q.; Ahmad, W.; Siddiqui, S.; Alam, M.; Abbas, K.; Moinuddin; Hassan, M.I.; Habib, S.; et al. Apoptosis: A Comprehensive Overview of Signaling Pathways, Morphological Changes, and Physiological Significance and Therapeutic Implications. Cells 2024, 13, 1838. [Google Scholar] [CrossRef]
- Yuan, W.; Ji, G.; Shi, X.; Sun, Z.; Liu, C.; Yu, Y.; Li, W.; Wang, X.; Hu, H. The Male Reproductive Toxicity after 5-Fluorouracil Exposure: DNA Damage, Oxidative Stress, and Mitochondrial Dysfunction in Vitro and in Vivo. Ecotoxicol. Environ. Saf. 2024, 278, 116465. [Google Scholar] [CrossRef]
- Rana, J.N.; Mumtaz, S.; Han, I.; Choi, E.H. Harnessing the Synergy of Nanosecond High-Power Microwave Pulses and Cisplatin to Increase the Induction of Apoptosis in Cancer Cells through the Activation of ATR/ATM and Intrinsic Pathways. Free Radic. Biol. Med. 2024, 225, 221–235. [Google Scholar] [CrossRef]
- Rana, J.N.; Mumtaz, S.; Han, I.; Choi, E.H. Formation of Reactive Species via High Power Microwave Induced DNA Damage and Promoted Intrinsic Pathway-Mediated Apoptosis in Lung Cancer Cells: An in Vitro Investigation. Fundam. Res. 2024, 4, 1542–1556. [Google Scholar] [CrossRef] [PubMed]
- Tragkola, V.; Anestopoulos, I.; Kyriakou, S.; Amery, T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Naturally-Derived Phenethyl Isothiocyanate Modulates Apoptotic Induction through Regulation of the Intrinsic Cascade and Resulting Apoptosome Formation in Human Malignant Melanoma Cells. Toxicol. Mech. Methods 2024, 34, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cheng, J.; Yao, Q.; Duan, J.; Chen, H.; Zhang, Z.; Yang, L.; Hua, R.; Li, Q. Isorhamnetin Improves Oocyte Maturation by Activating the Pi3k/Akt Signaling Pathway. Mol. Nutr. Food Res. 2024, 68, 2300904. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-H.; Lee, S.-E.; Yoon, J.-W.; Park, C.-O.; Park, H.-J.; Kim, S.-H.; Lee, D.-G.; Pyeon, D.-B.; Kim, E.-Y.; Park, S.-P. Isorhamnetin Improves in Vitro Maturation of Oxidative Stress-Exposed Porcine Oocytes and Subsequent Embryo Development. Zygote 2023, 31, 14–24. [Google Scholar] [CrossRef]
- Li, X.; Wang, T.; Zhou, Q.; Li, F.; Liu, T.; Zhang, K.; Wen, A.; Feng, L.; Shu, X.; Tian, S.; et al. Isorhamnetin Alleviates Mitochondrial Injury in Severe Acute Pancreatitis via Modulation of KDM5B/HtrA2 Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 3784. [Google Scholar] [CrossRef]
- Liu, M.; Lu, J.; Chen, Y.; Zhang, D.; Huang, W.; Shi, M.; Zhang, Y.; Wu, T.; Chen, Z.; Wu, L.; et al. Investigation of the Underlying Mechanism of Huangqi-Dangshen for Myasthenia Gravis Treatment via Molecular Docking and Network Pharmacology. Evid.-Based Complement. Altern. Med. 2023, 2023, 5301024. [Google Scholar] [CrossRef]
- An, X.; Yu, W.; Liu, J.; Tang, D.; Yang, L.; Chen, X. Oxidative Cell Death in Cancer: Mechanisms and Therapeutic Opportunities. Cell Death Dis. 2024, 15, 556. [Google Scholar] [CrossRef]
- Sun, B.; Sun, G.-B.; Xiao, J.; Chen, R.-C.; Wang, X.; Wu, Y.; Cao, L.; Yang, Z.-H.; Sun, X.-B. Isorhamnetin Inhibits H2O2-Induced Activation of the Intrinsic Apoptotic Pathway in H9c2 Cardiomyocytes through Scavenging Reactive Oxygen Species and ERK Inactivation. J. Cell. Biochem. 2012, 113, 473–485. [Google Scholar] [CrossRef]
- AL-Ishaq, R.K.; Liskova, A.; Kubatka, P.; Büsselberg, D. Enzymatic Metabolism of Flavonoids by Gut Microbiota and Its Impact on Gastrointestinal Cancer. Cancers 2021, 13, 3934. [Google Scholar] [CrossRef]
- Cai, F.; Zhang, Y.; Li, J.; Huang, S.; Gao, R. Isorhamnetin Inhibited the Proliferation and Metastasis of Androgen-Independent Prostate Cancer Cells by Targeting the Mitochondrion-Dependent Intrinsic Apoptotic and PI3K/Akt/MTOR Pathway. Biosci. Rep. 2020, 40, BSR20192826. [Google Scholar] [CrossRef]
- Jiang, H.; Yamashita, Y.; Nakamura, A.; Croft, K.; Ashida, H. Quercetin and Its Metabolite Isorhamnetin Promote Glucose Uptake through Different Signalling Pathways in Myotubes. Sci. Rep. 2019, 9, 2690. [Google Scholar] [CrossRef]
- Rodríguez, L.; Badimon, L.; Méndez, D.; Padró, T.; Vilahur, G.; Peña, E.; Carrasco, B.; Vogel, H.; Palomo, I.; Fuentes, E. Antiplatelet Activity of Isorhamnetin via Mitochondrial Regulation. Antioxidants 2021, 10, 666. [Google Scholar] [CrossRef]
- Matboli, M.; Saad, M.; Hasanin, A.H.; Saleh, L.A.; Baher, W.; Bekhet, M.M.; Eissa, S. New Insight into the Role of Isorhamnetin as a Regulator of Insulin Signaling Pathway in Type 2 Diabetes Mellitus Rat Model: Molecular and Computational Approach. Biomed. Pharmacother. 2021, 135, 111176. [Google Scholar] [CrossRef]
- Ru, Y.; Luo, Y.; Liu, D.; Huang, Q.; Zhou, X.; Linghu, M.; Luo, X.; Lv, Z.; Wu, Y.; Zhang, H.; et al. Isorhamnetin Alleviates Ferroptosis-Mediated Colitis by Activating the NRF2/HO-1 Pathway and Chelating Iron. Int. Immunopharmacol. 2024, 135, 112318. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Shin, B.Y.; Han, J.Y.; Kim, M.G.; Wi, J.E.; Kim, Y.W.; Cho, I.J.; Kim, S.C.; Shin, S.M.; Ki, S.H. Isorhamnetin Protects against Oxidative Stress by Activating Nrf2 and Inducing the Expression of Its Target Genes. Toxicol. Appl. Pharmacol. 2014, 274, 293–301. [Google Scholar] [CrossRef]
- Li, R.; Song, X.; Guo, Y.; Song, P.; Duan, D.; Chen, Z.-S. Natural Products: A Promising Therapeutics for Targeting Tumor Angiogenesis. Front. Oncol. 2021, 11, 772915. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Lee, D.-E.; Lee, K.W.; Son, J.E.; Seo, S.K.; Li, J.; Jung, S.K.; Heo, Y.-S.; Mottamal, M.; Bode, A.M.; et al. Isorhamnetin Suppresses Skin Cancer through Direct Inhibition of MEK1 and PI3-K. Cancer Prev. Res. 2011, 4, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Masarkar, N.; Pal, M.; Roy, M.; Yadav, A.K.; Pandya, B.; Lokhande, S.; Kanwar, J.R.; Ray, S.K.; Mukherjee, S. In-Silico Screening of Bioactive Compounds of Moringa Oleifera as Potential Inhibitors Targeting HIF-1α/VEGF/GLUT-1 Pathway against Breast Cancer. J. Complement. Integr. Med. 2024, 22, 149–164. [Google Scholar] [CrossRef]
- Laack, E.; Scheffler, A.; Burkholder, I.; Boeters, I.; Andritzky, B.; Schuch, G.; Görn, M.; Vohwinkel, G.; Edler, L.; Fiedler, W.; et al. Pretreatment Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinase-9 (MMP-9) Serum Levels in Patients with Metastatic Non-Small Cell Lung Cancer (NSCLC). Lung Cancer 2005, 50, 51–58. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, L.; Zhang, H.; Li, Y.; Lai, S. Effects of Isorhamnetin on Protein Expression of VEGF, MMP-2 and Endostatin in Lewis Lung Cancer Mouse. Int. J. Clin. Exp. Med. 2017, 10, 11488–11495. [Google Scholar]
- Luo, W.; Liu, Q.; Jiang, N.; Li, M.; Shi, L. Isorhamnetin Inhibited Migration and Invasion via Suppression of Akt/ERK-Mediated Epithelial-to-Mesenchymal Transition (EMT) in A549 Human Non-Small-Cell Lung Cancer Cells. Biosci. Rep. 2019, 39, BSR20190159. [Google Scholar] [CrossRef] [PubMed]
- Belal, A.; Elanany, M.A.; Raafat, M.; Hamza, H.T.; Mehany, A.B. Calendula officinalis Phytochemicals for the Treatment of Wounds Through Matrix Metalloproteinases-8 and 9 (MMP-8 and MMP-9): In Silico Approach. Nat. Prod. Commun. 2022, 17, 1934578X221098848. [Google Scholar] [CrossRef]
- Li, C.; Yang, D.; Zhao, Y.; Qiu, Y.; Cao, X.; Yu, Y.; Guo, H.; Gu, X.; Yin, X. Inhibitory Effects of Isorhamnetin on the Invasion of Human Breast Carcinoma Cells by Downregulating the Expression and Activity of Matrix Metalloproteinase-2/9. Nutr. Cancer 2015, 67, 1191–1200. [Google Scholar] [CrossRef]
- Hui, Q.; Yang, N.; Xiong, C.; Zhou, S.; Zhou, X.; Jin, Q.; Xu, X. Isorhamnetin Suppresses the Epithelial-Mesenchymal Transition of the Retinal Pigment Epithelium Both in Vivo and in Vitro through Nrf2-Dependent AKT/GSK-3β Pathway. Exp. Eye Res. 2024, 240, 109823. [Google Scholar] [CrossRef]
- Gao, M.; Ge, Z.; Deng, R.; Bao, B.; Yao, W.; Cao, Y.; Shan, M.; Cheng, F.; Yan, H.; Chen, P.; et al. Evaluation of VEGF Mediated Pro-Angiogenic and Hemostatic Effects and Chemical Marker Investigation for Typhae Pollen and Its Processed Product. J. Ethnopharmacol. 2021, 268, 113591. [Google Scholar] [CrossRef]
- Tan, Z.; Liu, J.; Hou, M.; Zhou, J.; Chen, Y.; Chen, X.; Leng, Y. Isorhamnetin Inhibits Cholangiocarcinoma Proliferation and Metastasis via PI3K/AKT Signaling Pathway. Discov. Oncol. 2025, 16, 469. [Google Scholar] [CrossRef]
- Zhang, C.; Qin, C.; Dewanjee, S.; Bhattacharya, H.; Chakraborty, P.; Jha, N.K.; Gangopadhyay, M.; Jha, S.K.; Liu, Q. Tumor-Derived Small Extracellular Vesicles in Cancer Invasion and Metastasis: Molecular Mechanisms, and Clinical Significance. Mol. Cancer 2024, 23, 18. [Google Scholar] [CrossRef]
- Shi, X.; Wang, X.; Yao, W.; Shi, D.; Shao, X.; Lu, Z.; Chai, Y.; Song, J.; Tang, W.; Wang, X. Mechanism Insights and Therapeutic Intervention of Tumor Metastasis: Latest Developments and Perspectives. Signal Transduct. Target. Ther. 2024, 9, 192. [Google Scholar] [CrossRef]
- Owczarek, K.; Caban, M.; Sosnowska, D.; Kajszczak, D.; Lewandowska, U. The Anti-Metastatic Potential of Aronia Leaf Extracts on Colon Cancer Cells. Nutrients 2024, 16, 4110. [Google Scholar] [CrossRef]
- Choi, J.-Y.; Jeong, M.; Lee, K.; Kim, J.-O.; Lee, W.H.; Park, I.; Kwon, H.C.; Choi, J.-H. Sedum Middendorffianum Maxim Induces Apoptosis and Inhibits the Invasion of Human Ovarian Cancer Cells via Oxidative Stress Regulation. Antioxidants 2023, 12, 1386. [Google Scholar] [CrossRef]
- Tossetta, G.; Fantone, S.; Togni, L.; Santarelli, A.; Olivieri, F.; Marzioni, D.; Rippo, M.R. Modulation of NRF2/KEAP1 Signaling by Phytotherapeutics in Periodontitis. Antioxidants 2024, 13, 1270. [Google Scholar] [CrossRef]
- Incorvaia, L.; Badalamenti, G.; Rini, G.; Arcara, C.; Fricano, S.; Sferrazza, C.; di Trapani, D.; Gebbia, N.; Leto, G. MMP-2, MMP-9 and Activin A Blood Levels in Patients with Breast Cancer or Prostate Cancer Metastatic to the Bone. Anticancer Res. 2007, 27, 1519–1525. [Google Scholar]
- Yao, Z.; Yuan, T.; Wang, H.; Yao, S.; Zhao, Y.; Liu, Y.; Jin, S.; Chu, J.; Xu, Y.; Zhou, W.; et al. MMP-2 Together with MMP-9 Overexpression Correlated with Lymph Node Metastasis and Poor Prognosis in Early Gastric Carcinoma. Tumor Biol. 2017, 39, 1010428317700411. [Google Scholar] [CrossRef] [PubMed]
- Kenny, H.A.; Lengyel, E. MMP-2 Functions as an Early Response Protein in Ovarian Cancer Metastasis. Cell Cycle 2009, 8, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Dwivedi, S.K.D.; Bhattacharya, R.; Mukherjee, P.; Rao, G. VEGF Signaling: Role in Angiogenesis and Beyond. Biochim. Biophys. Acta—Rev. Cancer 2024, 1879, 189079. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. VEGF as a Key Mediator of Angiogenesis in Cancer. Oncology 2005, 69, 4–10. [Google Scholar] [CrossRef]
- Bhattacharya, R.; Brown, J.S.; Gatenby, R.A.; Ibrahim-Hashim, A. A Gene for All Seasons: The Evolutionary Consequences of HIF-1 in Carcinogenesis, Tumor Growth and Metastasis. Semin. Cancer Biol. 2024, 102–103, 17–24. [Google Scholar] [CrossRef]
- Vazhappilly, C.G.; Alsawaf, S.; Mathew, S.; Nasar, N.A.; Hussain, M.I.; Cherkaoui, N.M.; Ayyub, M.; Alsaid, S.Y.; Thomas, J.G.; Cyril, A.C.; et al. Pharmacodynamics and Safety in Relation to Dose and Response of Plant Flavonoids in Treatment of Cancers. Inflammopharmacology 2024, 33, 11–47. [Google Scholar] [CrossRef]
- Demir, K.; Turgut, R.; Şentürk, S.; Işıklar, H.; Günalan, E. The Therapeutic Effects of Bioactive Compounds on Colorectal Cancer via PI3K/Akt/MTOR Signaling Pathway: A Critical Review. Food Sci. Nutr. 2024, 12, 9951–9973. [Google Scholar] [CrossRef]
- Rashidi, M.; Matour, E.; Nasab, H.B.; Cheraghzadeh, M.; Shakerian, E. Isorhamnetin Exerts Antifibrotic Effects by Attenuating Platelet-Derived Growth Factor-BB-Induced HSC-T6 Cells Activation via Suppressing PI3K-AKT Signaling Pathway. Iran. Biomed. J. 2023, 27, 199. [Google Scholar] [CrossRef]
- Zhai, T.; Zhang, X.; Hei, Z.; Jin, L.; Han, C.; Ko, A.T.; Yu, X.; Wang, J. Isorhamnetin Inhibits Human Gallbladder Cancer Cell Proliferation and Metastasis via PI3K/AKT Signaling Pathway Inactivation. Front. Pharmacol. 2021, 12, 628621. [Google Scholar] [CrossRef]
- Ye, L.; Ma, R.-H.; Zhang, X.-X.; Thakur, K.; Zhang, J.-G.; Khan, M.R.; Busquets, R.; Wei, Z.-J. Isorhamnetin Induces Apoptosis and Suppresses Metastasis of Human Endometrial Carcinoma Ishikawa Cells via Endoplasmic Reticulum Stress Promotion and Matrix Metalloproteinase-2/9 Inhibition In Vitro and In Vivo. Foods 2022, 11, 3415. [Google Scholar] [CrossRef] [PubMed]
- Mei, C.; Zhang, X.; Zhi, Y.; Liang, Z.; Xu, H.; Liu, Z.; Liu, Y.; Lyu, Y.; Wang, H. Isorhamnetin Regulates Programmed Death Ligand-1 Expression by Suppressing the EGFR–STAT3 Signaling Pathway in Canine Mammary Tumors. Int. J. Mol. Sci. 2024, 25, 670. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, H.; Shi, J.; Wang, Z.; Liang, Y.; Yu, J.; Wang, H.; Song, Z.; Tang, Z.; Zhang, D.; et al. Isorhamnetin Alleviates Renal Fibrosis by Inducing Endogenous Hydrogen Sulfide and Regulating Thiol-Based Redox State in Obstructed Kidneys. Biomolecules 2024, 14, 1233. [Google Scholar] [CrossRef]
- Yin, Q.; Wang, L.; Yu, H.; Chen, D.; Zhu, W.; Sun, C. Pharmacological Effects of Polyphenol Phytochemicals on the JAK-STAT Signaling Pathway. Front. Pharmacol. 2021, 12, 716672. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Mintie, C.A.; Ahmad, N. Grape Chemopreventive Agents Against Angiogenesis and Metastasis BT—Natural Products for Cancer Chemoprevention: Single Compounds and Combinations; Pezzuto, J.M., Vang, O., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 375–400. ISBN 978-3-030-39855-2. [Google Scholar]
- Sarwar, M.S.; Zhang, H.J.; Tsang, S.W. Perspectives of Plant Natural Products in Inhibition of Cancer Invasion and Metastasis by Regulating Multiple Signaling Pathways. Curr. Med. Chem. 2018, 25, 5057–5087. [Google Scholar] [CrossRef]
- Datta, A.; Deng, S.; Gopal, V.; Yap, K.C.; Halim, C.E.; Lye, M.L.; Ong, M.S.; Tan, T.Z.; Sethi, G.; Hooi, S.C.; et al. Cytoskeletal Dynamics in Epithelial-Mesenchymal Transition: Insights into Therapeutic Targets for Cancer Metastasis. Cancers 2021, 13, 1882. [Google Scholar] [CrossRef]
- Liang, D.; Liu, L.; Zhao, Y.; Luo, Z.; He, Y.; Li, Y.; Tang, S.; Tang, J.; Chen, N. Targeting Extracellular Matrix through Phytochemicals: A Promising Approach of Multi-Step Actions on the Treatment and Prevention of Cancer. Front. Pharmacol. 2023, 14, 1186712. [Google Scholar] [CrossRef]
- Kashyap, D.; Sharma, A.; Sak, K.; Tuli, H.S.; Buttar, H.S.; Bishayee, A. Fisetin: A Bioactive Phytochemical with Potential for Cancer Prevention and Pharmacotherapy. Life Sci. 2018, 194, 75–87. [Google Scholar] [CrossRef]
- Talambedu, U.; Sushil, K.; Arvind, K.; Mahesh, K.; DA, M.; Syed, F.; Peyush, G.; HP, P.; Veena, P. Molecular Docking Studies of Anti-Cancerous Candidates in Hippophae rhamnoides and Hippophae salicifolia. J. Biomed. Res. 2014, 28, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Oyeneyin, O.E.; Ipinloju, N.; da Costa, R.A.; Adigun, T.B.; Issahaku, A.R.; Wilhelm, A.; Alfa-Ibrahim, A.A.; Macaulay, A.M.; Muhammad, S.A. In Silico Evaluation of Bioactive Compounds of Citrullus Lanatus as Potential Noncovalent KRAS Inhibitors in the Treatment of Human Cancer. Chem. Afr. 2024, 7, 2553–2567. [Google Scholar] [CrossRef]
- Thipe, V.C.; Hall, N.; Pandurangi, A.; Ajayi, S.; Emeh, P.; Gauttam, I.; Ghamgui, R.; Hameedat, F.; Khelil, S.; Ly, N.K.; et al. Nano-Ayurvedic Medicine Approaches Using Ginkgo Biloba-Phytochemicals Functionalized Gold Nanoparticles Against Breast Cancer. Nanotechnol. Sci. Appl. 2024, 17, 189–210. [Google Scholar] [CrossRef]
- Gu, X.; Wei, S.; Lv, X. Circulating Tumor Cells: From New Biological Insights to Clinical Practice. Signal Transduct. Target. Ther. 2024, 9, 226. [Google Scholar] [CrossRef]
- Rajput, S.; Kumar Sharma, P.; Malviya, R. Fluid Mechanics in Circulating Tumour Cells: Role in Metastasis and Treatment Strategies. Med. Drug Discov. 2023, 18, 100158. [Google Scholar] [CrossRef]
- Di Russo, S.; Liberati, F.R.; Riva, A.; Di Fonzo, F.; Macone, A.; Giardina, G.; Arese, M.; Rinaldo, S.; Cutruzzolà, F.; Paone, A. Beyond the Barrier: The Immune-Inspired Pathways of Tumor Extravasation. Cell Commun. Signal. 2024, 22, 104. [Google Scholar] [CrossRef]
- Dompé, C.; Chojnowska, A.; Ramlau, R.; Nowicki, M.; Alix-Panabières, C.; Budna-Tukan, J. Unveiling the Dynamics of Circulating Tumor Cells in Colorectal Cancer: From Biology to Clinical Applications. Front. Cell Dev. Biol. 2024, 12, 1498032. [Google Scholar] [CrossRef]
- Menyailo, M.E.; Bokova, U.A.; Ivanyuk, E.E.; Khozyainova, A.A.; Denisov, E.V. Metastasis Prevention: Focus on Metastatic Circulating Tumor Cells. Mol. Diagn. Ther. 2021, 25, 549–562. [Google Scholar] [CrossRef]
- Paterlini-Brechot, P.; Benali, N.L. Circulating Tumor Cells (CTC) Detection: Clinical Impact and Future Directions. Cancer Lett. 2007, 253, 180–204. [Google Scholar] [CrossRef]
- Leone, K.; Poggiana, C.; Zamarchi, R. The Interplay between Circulating Tumor Cells and the Immune System: From Immune Escape to Cancer Immunotherapy. Diagnostics 2018, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Wang, D.; Ma, X.; Chen, W.; Guo, S.; Guan, H. Effects of Total Flavonoids of Sea Buckthorn (Hippophae rhamnoides L.) on Cytotoxicity of NK92-MI Cells. Int. J. Immunopathol. Pharmacol. 2017, 30, 353–361. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Iordache, F.; Stanca, L.; Cimpeanu, C.; Furnaris, F.; Geicu, O.I.; Bilteanu, L.; Serban, A.I. Comprehensive and Critical View on the Anti-Inflammatory and Immunomodulatory Role of Natural Phenolic Antioxidants. Eur. J. Med. Chem. 2024, 265, 116075. [Google Scholar] [CrossRef]
- Han, L.; Fu, Q.; Deng, C.; Luo, L.; Xiang, T.; Zhao, H. Immunomodulatory Potential of Flavonoids for the Treatment of Autoimmune Diseases and Tumour. Scand. J. Immunol. 2022, 95, e13106. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Q.; Liang Cheng, M.; Ma, L.; Zhi Meng, Q.; Duan, L.; Chen, Y.; Wu Tan, J.; Chen, M.; Ting Liang, T.; et al. Effect of the Miaoyao Fanggan Sachet-Derived Isorhamnetin on TLR2/4 and NKp46 Expression in Mice. J. Ethnopharmacol. 2012, 144, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.; Mijares, M.R.; De Sanctis, J.B. Effects of Flavonoids and Its Derivatives on Immune Cell Responses. Recent. Pat. Inflamm. Allergy Drug Discov. 2019, 13, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, H.; Hua, J.; Luo, S. Cytotoxic Activity of Phenolic Compounds in Bairui Granules Obtained from the Chinese Medicinal Plant Thesium Chinense. Front. Chem. 2024, 12, 1506792. [Google Scholar] [CrossRef]
- Shakoor, H.; Feehan, J.; Apostolopoulos, V.; Platat, C.; Al Dhaheri, A.S.; Ali, H.I.; Ismail, L.C.; Bosevski, M.; Stojanovska, L. Immunomodulatory Effects of Dietary Polyphenols. Nutrients 2021, 13, 728. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, X.; Shao, R.; Zhao, R.; Yanamandra, A.K.; Xing, Z.; Ding, M.; Wang, J.; Liu, T.; Zheng, Q.; et al. Bioactive Constituents of Verbena Officinalis Alleviate Inflammation and Enhance Killing Efficiency of Natural Killer Cells. Int. J. Mol. Sci. 2023, 24, 7144. [Google Scholar] [CrossRef]
- Ren, X.; Han, L.; Li, Y.; Zhao, H.; Zhang, Z.; Zhuang, Y.; Zhong, M.; Wang, Q.; Ma, W.; Wang, Y. Isorhamnetin Attenuates TNF-α-Induced Inflammation, Proliferation, and Migration in Human Bronchial Epithelial Cells via MAPK and NF-ΚB Pathways. Anat. Rec. 2021, 304, 901–913. [Google Scholar] [CrossRef]
- Liu, J.; Ren, L.; Wang, H.; Li, Z. Isoquercitrin Induces Endoplasmic Reticulum Stress and Immunogenic Cell Death in Gastric Cancer Cells. Biochem. Genet. 2023, 61, 1128–1142. [Google Scholar] [CrossRef]
- Fakhri, S.; Moradi, S.Z.; Yarmohammadi, A.; Narimani, F.; Wallace, C.E.; Bishayee, A. Modulation of TLR/NF-ΚB/NLRP Signaling by Bioactive Phytocompounds: A Promising Strategy to Augment Cancer Chemotherapy and Immunotherapy. Front. Oncol. 2022, 12, 834072. [Google Scholar] [CrossRef]
- Liu, H.; Han, J.; Lv, Y.; Zhao, Z.; Zheng, S.; Sun, Y.; Sun, T. Isorhamnetin and Anti-PD-L1 Antibody Dual-Functional Mesoporous Silica Nanoparticles Improve Tumor Immune Microenvironment and Inhibit YY1-Mediated Tumor Progression. J. Nanobiotechnol. 2023, 21, 208. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, K.; Yuan, W.; Sun, K. Radix Bupleuri-Radix Paeoniae Alba Inhibits the Development of Hepatocellular Carcinoma through Activation of the PTEN/PD-L1 Axis within the Immune Microenvironment. Nutr. Cancer 2024, 76, 63–79. [Google Scholar] [CrossRef]
- Masuelli, L.; Benvenuto, M.; Focaccetti, C.; Ciuffa, S.; Fazi, S.; Bei, A.; Miele, M.T.; Piredda, L.; Manzari, V.; Modesti, A.; et al. Targeting the Tumor Immune Microenvironment with “Nutraceuticals”: From Bench to Clinical Trials. Pharmacol. Ther. 2021, 219, 107700. [Google Scholar] [CrossRef]
- Ganbold, M.; Louphrasitthiphol, P.; Miyazaki, Y.; Oda, T.; Tominaga, K.; Isoda, H. Abstract C068: Isorhamnetin Restricts Cancer-Associated Fibroblasts (CAFs) Phenotypic Plasticity, Poising Them towards Tumor-Restraining MyCAFs: Potential Use of Isorhamnetin as Neoadjuvant in Pancreatic Ductal Adenocarcinoma (PDAC). Cancer Res. 2022, 82, C068. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, Q.; Zhou, X.; Wang, X.; Li, H.; Zhang, W.; Yuan, H.; Sun, C. Flavonoids Regulate Tumor-Associated Macrophages —From Structure-Activity Relationship to Clinical Potential (Review). Pharmacol. Res. 2022, 184, 106419. [Google Scholar] [CrossRef]
- Hu, Q.; Li, Z.; Li, Y.; Deng, X.; Chen, Y.; Ma, X.; Zeng, J.; Zhao, Y. Natural Products Targeting Signaling Pathways Associated with Regulated Cell Death in Gastric Cancer: Recent Advances and Perspectives. Phyther. Res. 2023, 37, 2661–2692. [Google Scholar] [CrossRef]
- Zulkefli, N.; Che Zahari, C.N.; Sayuti, N.H.; Kamarudin, A.A.; Saad, N.; Hamezah, H.S.; Bunawan, H.; Baharum, S.N.; Mediani, A.; Ahmed, Q.U.; et al. Flavonoids as Potential Wound-Healing Molecules: Emphasis on Pathways Perspective. Int. J. Mol. Sci. 2023, 24, 4607. [Google Scholar] [CrossRef]
- Aonuma, K.; Ferdousi, F.; Xu, D.; Tominaga, K.; Isoda, H. Effects of Isorhamnetin in Human Amniotic Epithelial Stem Cells in Vitro and Its Cardioprotective Effects in Vivo. Front. Cell Dev. Biol. 2020, 8, 578197. [Google Scholar] [CrossRef]
- Zhang, P.; Sun, Y.; Shi, L.; Sun, D.; Wang, L.; Feng, D.; Ding, C. Effect of Isorhamnetin on Carbonic Anhydrase IX Expression and Tumorigenesis of Bladder Cancer by Activating PPARγ/PTEN/AKT Pathway. Tissue Cell 2023, 82, 102048. [Google Scholar] [CrossRef]
- Manu, K.A.; Shanmugam, M.K.; Ramachandran, L.; Li, F.; Siveen, K.S.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Arfuso, F.; Kumar, A.P.; et al. Isorhamnetin Augments the Anti-Tumor Effect of Capeciatbine through the Negative Regulation of NF-ΚB Signaling Cascade in Gastric Cancer. Cancer Lett. 2015, 363, 28–36. [Google Scholar] [CrossRef]
- Seo, K.; Yang, J.H.; Kim, S.C.; Ku, S.K.; Ki, S.H.; Shin, S.M. The Antioxidant Effects of Isorhamnetin Contribute to Inhibit COX-2 Expression in Response to Inflammation: A Potential Role of HO-1. Inflammation 2014, 37, 712–722. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.; Zhang, Z.; Tong, L.; Yu, S.; Liu, Y.; Yang, F. Flavonoid Compounds in Hippophae rhamnoides L. Protect Endothelial Cells from Oxidative Damage Through the PI3K/AKT-ENOS Pathway. Chem. Biodivers. 2024, 21, e202400300. [Google Scholar] [CrossRef]
- Mumtaz, S.; Rana, J.N. Impact of Nonthermal Plasma on Human Metapneumovirus (HMPV): Mechanisms of Viral Inactivation and Replication Inhibition. Contrib. Plasma Phys. 2025, 65, e70044. [Google Scholar] [CrossRef]
- Huang, R.; Zhou, P.-K. DNA Damage Repair: Historical Perspectives, Mechanistic Pathways and Clinical Translation for Targeted Cancer Therapy. Signal Transduct. Target. Ther. 2021, 6, 254. [Google Scholar] [CrossRef]
- Pfaff, A.; Chernatynskaya, A.; Vineyard, H.; Ercal, N. Thiol Antioxidants Protect Human Lens Epithelial (HLE B-3) Cells against Tert-Butyl Hydroperoxide-Induced Oxidative Damage and Cytotoxicity. Biochem. Biophys. Rep. 2022, 29, 101213. [Google Scholar] [CrossRef]
- Song, H.-K.; Kim, H.J.; Kim, S.C.; Kim, T. Therapeutic Effects of Sigesbeckia Pubescens Makino Against Atopic Dermatitis-Like Skin Inflammation Through the JAK2/STAT Signaling Pathway. Int. J. Mol. Sci. 2025, 26, 4191. [Google Scholar] [CrossRef]
- Pal, C. Small Molecules Targeting Mitochondria: A Mechanistic Approach to Combating Doxorubicin-Induced Cardiotoxicity. Cardiovasc. Toxicol. 2024. [Google Scholar] [CrossRef]
- Mihaylova, R.; Gevrenova, R.; Petrova, A.; Savov, Y.; Zheleva-Dimitrova, D.; Balabanova, V.; Momekov, G.; Simeonova, R. Mitigating Effects of Tanacetum balsamita L. on Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD). Plants 2024, 13, 2086. [Google Scholar] [CrossRef]
- Sattari, M.; Amri, J.; Shahaboddin, M.E.; Sattari, M.; Tabatabaei-Malazy, O.; Azmon, M.; Meshkani, R.; Panahi, G. The Protective Effects of Fisetin in Metabolic Disorders: A Focus on Oxidative Stress and Associated Events. J. Diabetes Metab. Disord. 2024, 23, 1753–1771. [Google Scholar] [CrossRef]
- Mao, Y.; Zha, Y.; Zang, Y.; Gao, Y.; Sun, J.; Liu, Y.; Wang, Z.; Wei, Z.; Wang, M.; Yang, Y. Isorhamnetin Improves Diabetes-Induced Erectile Dysfunction in Rats through Activation of the PI3K/AKT/ENOS Signaling Pathway. Biomed. Pharmacother. 2024, 177, 116987. [Google Scholar] [CrossRef]
- Papas, M.; Arroyo, L.; Bassols, A.; Catalán, J.; Bonilla-Correal, S.; Gacem, S.; Yeste, M.; Miró, J. Activities of Antioxidant Seminal Plasma Enzymes (SOD, CAT, GPX and GSR) Are Higher in Jackasses than in Stallions and Are Correlated with Sperm Motility in Jackasses. Theriogenology 2019, 140, 180–187. [Google Scholar] [CrossRef]
- Cengiz, M.; Gür, B.; Gür, F.; Şahintürk, V.; Bayrakdar, A.; Şahin, I.K.; Başkoy, S.A.; Bilici, N.; Onur, S.; Kaya, Y.; et al. The Protective Effects of Selenium and Boron on Cyclophosphamide-Induced Hepatic Oxidative Stress, Inflammation, and Apoptosis in Rats. Heliyon 2024, 10, e38713. [Google Scholar] [CrossRef]
- Chen, F.Y.; Ma, P.; Bo, L.; Lv, J.Y.; Zhou, W.; Zhou, R. Isorhamnetin Alleviates Symptoms and Inhibits Oxidative Stress Levels in Rats with Pulmonary Arterial Hypertension. Iran. J. Basic Med. Sci. 2024, 27, 1616–1623. [Google Scholar] [CrossRef]
- Xu, Y.; Li, J.; Lin, Z.; Liang, W.; Qin, L.; Ding, J.; Chen, S.; Zhou, L. Isorhamnetin Alleviates Airway Inflammation by Regulating the Nrf2/Keap1 Pathway in a Mouse Model of COPD. Front. Pharmacol. 2022, 13, 860362. [Google Scholar] [CrossRef]
- Rashidi, M.; Cheraghzadeh, M.; Shakerian, E.; Matour, E.; Beheshti Nasab, H.; Salehipour Bavarsad, S. Investigating the Inhibitory Effect of Isorhamnetin on the Expression of NOXs Genes as Well as the Level of ROS in Hepatic Stellate Cells and Its Role in Preventing the Progression of Liver Fibrogenesis. J. Isfahan Med. Sch. 2024, 42, 113–120. [Google Scholar] [CrossRef]
- Xu, X.; Guo, Y.; Chen, M.; Li, N.; Sun, Y.; Ren, S.; Xiao, J.; Wang, D.; Liu, X.; Pan, Y. Hypoglycemic Activities of Flowers of Xanthoceras Sorbifolia and Identification of Anti-Oxidant Components by off-Line UPLC-QTOF-MS/MS-Free Radical Scavenging Detection. Chin. Herb. Med. 2024, 16, 151–161. [Google Scholar] [CrossRef]
- Chen, Y.; He, W.; Cao, H.; Wang, Z.; Liu, J.; Wang, B.; Wang, C. Research Progress of Sea Buckthorn (Hippophae rhamnoides L.) in Prevention and Treatment of Cardiovascular Disease. Front. Cardiovasc. Med. 2024, 11, 1477636. [Google Scholar] [CrossRef]
- Aly, S.H.; Mahmoud, A.M.A.; Abdel Mageed, S.S.; Khaleel, E.F.; Badi, R.M.; Elkaeed, E.B.; Rasheed, R.A.; El Hassab, M.A.; Eldehna, W.M. Exploring the Phytochemicals, Antioxidant Properties, and Hepatoprotective Potential of Moricandia Sinaica Leaves against Paracetamol-Induced Toxicity: Biological Evaluations and in Silico Insights. PLoS ONE 2024, 19, e0307901. [Google Scholar] [CrossRef]
- Wang, Z.; Mao, Y.; Zang, Y.; Zha, Y.; Sun, J.; Wei, Z.; He, S.; Zhang, X.; Wang, M.; Yang, Y. Transcriptomic Analysis Reveals the Mechanism of Isorhamnetin in the Treatment of Diabetes Mellitus Erectile Dysfunction. Free Radic. Biol. Med. 2024, 224, 366–381. [Google Scholar] [CrossRef]
- Jian, J.; Yu-Qing, L.; Rang-Yue, H.; Xia, Z.; Ke-Huan, X.; Ying, Y.; Li, W.; Rui-zhi, T. Isorhamnetin Ameliorates Cisplatin-Induced Acute Kidney Injury in Mice by Activating SLPI-Mediated Anti-Inflammatory Effect in Macrophage. Immunopharmacol. Immunotoxicol. 2024, 46, 319–329. [Google Scholar] [CrossRef]
- Helen, H.; Gunawan, M.C.; Halim, P.; Dinata, M.R.; Ahmed, A.; Dalimunthe, A.; Marianne, M.; Ribeiro, R.I.M.D.A.; Hasibuan, P.A.Z.; Nurkolis, F.; et al. Flavonoids as Modulators of MiRNA Expression in Pancreatic Cancer: Pathways, Mechanisms, And Therapeutic Potential. Biomed. Pharmacother. 2024, 179, 117347. [Google Scholar] [CrossRef]
- De Cecco, F.; Franceschelli, S.; Panella, V.; Maggi, M.A.; Bisti, S.; Bravo Nuevo, A.; D’Ardes, D.; Cipollone, F.; Speranza, L. Biological Response of Treatment with Saffron Petal Extract on Cytokine-Induced Oxidative Stress and Inflammation in the Caco-2/Human Leukemia Monocytic Co-Culture Model. Antioxidants 2024, 13, 1257. [Google Scholar] [CrossRef]
- Dąbek, J.; Kułach, A.; Gąsior, Z. Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells (NF-ΚB): A New Potential Therapeutic Target in Atherosclerosis? Pharmacol. Rep. 2010, 62, 778–783. [Google Scholar] [CrossRef]
- Pérez-Pérez, D.; Fuentes-Pananá, E.M.; Flores-Hermenegildo, J.M.; Romero-Ramirez, H.; Santos-Argumedo, L.; Kilimann, M.W.; Rodríguez-Alba, J.C.; Lopez-Herrera, G. Lipopolysaccharide-Responsive Beige-like Anchor Is Involved in Regulating NF-ΚB Activation in B Cells. Front. Immunol. 2024, 15, 1409434. [Google Scholar] [CrossRef] [PubMed]
- Abdel Bar, F.M.; Alonazi, R.; Elekhnawy, E.; Samra, R.M.; Alqarni, M.H.; Badreldin, H.; Magdy, G. HPLC-PDA and in Vivo Anti-Inflammatory Potential of Isorhamnetin-3-O-β-D-Glucoside from Zygophyllum simplex L. J. Ethnopharmacol. 2025, 338, 119089. [Google Scholar] [CrossRef] [PubMed]
- Gungor, H.; Ekici, M.; Ates, M.B. Lipid-Lowering, Anti-Inflammatory, and Hepatoprotective Effects of Isorhamnetin on Acetaminophen-Induced Hepatotoxicity in Mice. Drug Chem. Toxicol. 2023, 46, 566–574. [Google Scholar] [CrossRef]
- Hogan, I.A.; Kuo, Y.-C.; Abubakar, A.N.; Lawal, B.; Agboola, A.R.; Lukman, H.Y.; Onikanni, S.A.; Olawale, F.; Fadaka, A.O.; Ibrahim, Y.O.; et al. Attenuation of Hyperglycemia-Associated Dyslipidemic, Oxidative, Cognitive, and Inflammatory Crises via Modulation of Neuronal ChEs/NF-ΚB/COX-2/NOx, and Hepatorenal Functional Deficits by the Tridax Procumbens Extract. Biomed. Pharmacother. 2023, 158, 114114. [Google Scholar] [CrossRef]
- Alqudah, A.; Qnais, E.Y.; Wedyan, M.A.; Altaber, S.; Bseiso, Y.; Oqal, M.; AbuDalo, R.; Alrosan, K.; Alrosan, A.Z.; Bani Melhim, S.; et al. Isorhamnetin Reduces Glucose Level, Inflammation, and Oxidative Stress in High-Fat Diet/Streptozotocin Diabetic Mice Model. Molecules 2023, 28, 502. [Google Scholar] [CrossRef]
- Saikia, L.; Talukdar, N.C.; Dutta, P.P. Exploring the Therapeutic Role of Flavonoids Through AMPK Activation in Metabolic Syndrome: A Narrative Review. Phyther. Res. 2025, 39, 1403–1421. [Google Scholar] [CrossRef]
- Rana, J.N.; Mumtaz, S.; Choi, E.H.; Han, I. ROS Production in Response to High-Power Microwave Pulses Induces P53 Activation and DNA Damage in Brain Cells: Radiosensitivity and Biological Dosimetry Evaluation. Front. Cell Dev. Biol. 2023, 11, 1067861. [Google Scholar] [CrossRef]
- Efe, G.; Rustgi, A.K.; Prives, C. P53 at the Crossroads of Tumor Immunity. Nat. Cancer 2024, 5, 983–995. [Google Scholar] [CrossRef]
- Peuget, S.; Zhou, X.; Selivanova, G. Translating P53-Based Therapies for Cancer into the Clinic. Nat. Rev. Cancer 2024, 24, 192–215. [Google Scholar] [CrossRef]
- Fischer, M.; Sammons, M.A. Determinants of P53 DNA Binding, Gene Regulation, and Cell Fate Decisions. Cell Death Differ. 2024, 31, 836–843. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J.; Yang, W.; Sheng, H.; Jia, B.; Cheng, P.; Xu, S.; Hong, X.; Jiang, C.; Yang, Y.; et al. Multiple Roles of P53 in Cancer Development: Regulation of Tumor Microenvironment, M6A Modification and Diverse Cell Death Mechanisms. J. Adv. Res. 2024. [Google Scholar] [CrossRef]
- Liu, Y.; Su, Z.; Tavana, O.; Gu, W. Understanding the Complexity of P53 in a New Era of Tumor Suppression. Cancer Cell 2024, 42, 946–967. [Google Scholar] [CrossRef]
- Indeglia, A.; Murphy, M.E. Elucidating the Chain of Command: Our Current Understanding of Critical Target Genes for P53-Mediated Tumor Suppression. Crit. Rev. Biochem. Mol. Biol. 2024, 59, 128–138. [Google Scholar] [CrossRef]
- Nishiyama, Y.; Morita, A.; Tatsuta, S.; Kanamaru, M.; Sakaue, M.; Ueda, K.; Shono, M.; Fujita, R.; Wang, B.; Hosoi, Y.; et al. Isorhamnetin Promotes 53BP1 Recruitment through the Enhancement of ATM Phosphorylation and Protects Mice from Radiation Gastrointestinal Syndrome. Genes 2021, 12, 1514. [Google Scholar] [CrossRef]
- Vaddavalli, P.L.; Schumacher, B. The P53 Network: Cellular and Systemic DNA Damage Responses in Cancer and Aging. Trends Genet. 2022, 38, 598–612. [Google Scholar] [CrossRef]
- Li, C.; Chen, H.; Chen, X.; Wang, P.; Shi, Y.; Xie, X.; Chen, Y.; Cai, X. Identification of Inflammatory Response-Related Molecular Mechanisms Based on the ATM/ATR/P53 Pathway in Tumor Cells. Comput. Biol. Med. 2024, 180, 108776. [Google Scholar] [CrossRef]
- Uchida, Y.; Ferdousi, F.; Zheng, Y.-W.; Oda, T.; Isoda, H. Global Gene Expression Profiling Reveals Isorhamnetin Induces Hepatic-Lineage Specific Differentiation in Human Amniotic Epithelial Cells. Front. Cell Dev. Biol. 2020, 8, 578036. [Google Scholar] [CrossRef]
- Mohamed, E.M.; Hetta, M.H.; Rateb, M.E.; Selim, M.A.; AboulMagd, A.M.; Badria, F.A.; Abdelmohsen, U.R.; Alhadrami, H.A.; Hassan, H.M. Bioassay-Guided Isolation, Metabolic Profiling, and Docking Studies of Hyaluronidase Inhibitors from Ravenala Madagascariensis. Molecules 2020, 25, 1714. [Google Scholar] [CrossRef]
- Chau, T.P.; Devanesan, S.; Ayub, R.; Perumal, K. Identification and Characterization of Major Bioactive Compounds from Andrographis Paniculata (Burm. f.) Extracts Showed Multi-Biomedical Applications. Environ. Res. 2024, 242, 117763. [Google Scholar] [CrossRef]
- Li, J.; Sun, L.; Wang, F.; Yin, S.; Li, S.; Zhang, J.; Wu, D. Pro-Differentiative, Pro-Adhesive and Pro-Migratory Activities of Isorhamnetin in MC3T3-E1 Osteoblasts via Activation of ERK-Dependent BMP2-Smad Signaling. Cell Biochem. Biophys. 2024, 82, 3607–3617. [Google Scholar] [CrossRef] [PubMed]
- Shi, A.; Liu, L.; Li, S.; Qi, B. Natural Products Targeting the MAPK-Signaling Pathway in Cancer: Overview. J. Cancer Res. Clin. Oncol. 2024, 150, 6. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.-J. Pathological Roles of MAPK Signaling Pathways in Human Diseases. Biochim. Biophys. Acta—Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S. Targeting MAPK Signaling: A Promising Approach for Treating Inflammatory Lung Disease. Pathol.—Res. Pract. 2024, 254, 155122. [Google Scholar] [CrossRef]
- Edvinsson, L.; Krause, D.N. Switching Off Vascular MAPK Signaling: A Novel Strategy to Prevent Delayed Cerebral Ischemia Following Subarachnoid Hemorrhage. Transl. Stroke Res. 2024, 16, 952–961. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.T. MAPK Signal Pathways in the Regulation of Cell Proliferation in Mammalian Cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. P38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef]
- Sahu, R.; Rawal, R.K. Modulation of the C-JNK/P38-MAPK Signaling Pathway: Investigating the Therapeutic Potential of Natural Products in Hypertension. Phytomed. Plus 2024, 4, 100564. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Z.; Wang, S.; Wang, X.; Mao, J. Targeting MAPK-ERK/JNK Pathway: A Potential Intervention Mechanism of Myocardial Fibrosis in Heart Failure. Biomed. Pharmacother. 2024, 173, 116413. [Google Scholar] [CrossRef] [PubMed]
- Baier, A.; Szyszka, R. Compounds from Natural Sources as Protein Kinase Inhibitors. Biomolecules 2020, 10, 1546. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A. Targeting the RAS Upstream and Downstream Signaling Pathway for Cancer Treatment. Eur. J. Pharmacol. 2024, 979, 176727. [Google Scholar] [CrossRef]
- Mozzarelli, A.M.; Simanshu, D.K.; Castel, P. Functional and Structural Insights into RAS Effector Proteins. Mol. Cell 2024, 84, 2807–2821. [Google Scholar] [CrossRef]
- Kondoh, K.; Torii, S.; Nishida, E. Control of MAP Kinase Signaling to the Nucleus. Chromosoma 2005, 114, 86–91. [Google Scholar] [CrossRef]
- Mebratu, Y.; Tesfaigzi, Y. How ERK1/2 Activation Controls Cell Proliferation and Cell Death: Is Subcellular Localization the Answer? Cell Cycle 2009, 8, 1168–1175. [Google Scholar] [CrossRef]
- Plotnikov, A.; Flores, K.; Maik-Rachline, G.; Zehorai, E.; Kapri-Pardes, E.; Berti, D.A.; Hanoch, T.; Besser, M.J.; Seger, R. The Nuclear Translocation of ERK1/2 as an Anticancer Target. Nat. Commun. 2015, 6, 6685. [Google Scholar] [CrossRef]
- Shi, X.; Yu, Q.; Wang, K.; Fu, Y.; Zhang, S.; Liao, Z.; Li, Y.; Cai, T. Active Ingredients Isorhamnetin of Croci Srigma Inhibit Stomach Adenocarcinomas Progression by MAPK/MTOR Signaling Pathway. Sci. Rep. 2023, 13, 12607. [Google Scholar] [CrossRef]
- Gao, F.; Zhou, C.; Qiu, W.; Wu, H.; Li, J.; Peng, J.; Qiu, M.; Liang, C.; Gao, J.; Luo, S. Total Flavonoids from Semen Cuscutae Target MMP9 and Promote Invasion of EVT Cells via Notch/AKT/MAPK Signaling Pathways. Sci. Rep. 2018, 8, 17342. [Google Scholar] [CrossRef]
- Qiu, S.; Sun, G.; Zhang, Y.; Li, X.; Wang, R. Involvement of the NF-ΚB Signaling Pathway in the Renoprotective Effects of Isorhamnetin in a Type 2 Diabetic Rat Model. Biomed. Rep. 2016, 4, 628–634. [Google Scholar] [CrossRef]
- Gao, L.; Yao, R.; Liu, Y.; Wang, Z.; Huang, Z.; Du, B.; Zhang, D.; Wu, L.; Xiao, L.; Zhang, Y. Isorhamnetin Protects against Cardiac Hypertrophy through Blocking PI3K–AKT Pathway. Mol. Cell. Biochem. 2017, 429, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, T.; Chen, K.; Xia, Y.; Dai, W.; Xu, S.; Xu, L.; Wang, F.; Wu, L.; Li, J.; et al. Isorhamnetin: A Hepatoprotective Flavonoid Inhibits Apoptosis and Autophagy via P38/PPAR-α Pathway in Mice. Biomed. Pharmacother. 2018, 103, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Zarneshan, S.N.; Fakhri, S.; Farzaei, M.H.; Khan, H.; Saso, L. Astaxanthin Targets PI3K/Akt Signaling Pathway toward Potential Therapeutic Applications. Food Chem. Toxicol. 2020, 145, 111714. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, N.; Bawari, S.; Burcher, J.T.; Sinha, D.; Tewari, D.; Bishayee, A. Targeting Cell Signaling Pathways in Lung Cancer by Bioactive Phytocompounds. Cancers 2023, 15, 3980. [Google Scholar] [CrossRef]
- Brillo, V.; Chieregato, L.; Leanza, L.; Muccioli, S.; Costa, R. Mitochondrial Dynamics, ROS, and Cell Signaling: A Blended Overview. Life 2021, 11, 332. [Google Scholar] [CrossRef]
- Aggarwal, V.; Kashyap, D.; Sak, K.; Tuli, H.S.; Jain, A.; Chaudhary, A.; Garg, V.K.; Sethi, G.; Yerer, M.B. Molecular Mechanisms of Action of Tocotrienols in Cancer: Recent Trends and Advancements. Int. J. Mol. Sci. 2019, 20, 656. [Google Scholar] [CrossRef]
- Liu, N.; Feng, J.; Lu, X.; Yao, Z.; Liu, Q.; Lv, Y.; Han, Y.; Deng, J.; Zhou, Y. Isorhamnetin Inhibits Liver Fibrosis by Reducing Autophagy and Inhibiting Extracellular Matrix Formation via the TGF-Β1/Smad3 and TGF-Β1/P38 MAPK Pathways. Mediat. Inflamm. 2019, 2019, 6175091. [Google Scholar] [CrossRef]
- Hwang, S.-L.; Shih, P.-H.; Yen, G.-C. Neuroprotective Effects of Citrus Flavonoids. J. Agric. Food Chem. 2012, 60, 877–885. [Google Scholar] [CrossRef]
- Jayashankar, B.; Mishra, K.P.; Kumar, M.S.Y.; Udayasankar, K.; Misra, K.; Ganju, L.; Singh, S.B. A Supercritical CO2 Extract from Seabuckthorn Leaves Inhibits Pro-Inflammatory Mediators via Inhibition of Mitogen Activated Protein Kinase P38 and Transcription Factor Nuclear Factor-ΚB. Int. Immunopharmacol. 2012, 13, 461–467. [Google Scholar] [CrossRef]
- Tuntipopipat, S.; Muangnoi, C.; Chingsuwanrote, P.; Parengam, M.; Chantravisut, P.; Charoenkiatkul, S.; Svasti, S. Anti-Inflammatory Activities of Red Curry Paste Extract on Lipopolysaccharide-Activated Murine Macrophage Cell Line. Nutrition 2011, 27, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, X.; Liu, T.; Guan, M.; Feng, X.; Dong, W.; Chu, X.; Liu, J.; Tian, X.; Ci, X.; et al. Kaempferol Regulates MAPKs and NF-ΚB Signaling Pathways to Attenuate LPS-Induced Acute Lung Injury in Mice. Int. Immunopharmacol. 2012, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Kim, S.C.; Shin, B.Y.; Jin, S.H.; Jo, M.J.; Jegal, K.H.; Kim, Y.W.; Lee, J.R.; Ku, S.K.; Cho, I.J.; et al. O-Methylated Flavonol Isorhamnetin Prevents Acute Inflammation through Blocking of NF-ΚB Activation. Food Chem. Toxicol. 2013, 59, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Kim Young, S.; Jin, C.; Kim Hong, C.; Yoo Hyun, Y.; Choi Hyun, S.; Kim, G.; Yoon Min, H.; Park Tae, H.; Choi Hyun, Y. Isorhamnetin Alleviates Lipopolysaccharide-Induced Inflammatory Responses in BV2 Microglia by Inactivating NF-ΚB, Blocking the TLR4 Pathway and Reducing ROS Generation. Int. J. Mol. Med. 2019, 43, 682–692. [Google Scholar] [CrossRef]
- Yang, J.H.; Kim, S.C.; Kim, K.M.; Jang, C.H.; Cho, S.S.; Kim, S.J.; Ku, S.K.; Cho, I.J.; Ki, S.H. Isorhamnetin Attenuates Liver Fibrosis by Inhibiting TGF-β/Smad Signaling and Relieving Oxidative Stress. Eur. J. Pharmacol. 2016, 783, 92–102. [Google Scholar] [CrossRef]
- Jin, J.Y.; Choi, E.Y.; Park, H.R.; Choi, J.I.; Choi, I.S.; Kim, S.J. Isorhamnetin Inhibits Revotella Intermedia Lipopolysaccharide-Induced Production of Interleukin-6 in Murine Macrophages via Anti-Inflammatory Heme Oxygenase-1 Induction and Inhibition of Nuclear Factor-ΚB and Signal Transducer and Activator of Transcripti. J. Periodontal Res. 2013, 48, 687–695. [Google Scholar] [CrossRef]
- Jnawali, H.N.; Jeon, D.; Jeong, M.-C.; Lee, E.; Jin, B.; Ryoo, S.; Yoo, J.; Jung, I.D.; Lee, S.J.; Park, Y.-M.; et al. Antituberculosis Activity of a Naturally Occurring Flavonoid, Isorhamnetin. J. Nat. Prod. 2016, 79, 961–969. [Google Scholar] [CrossRef]
- Jazvinšćak Jembrek, M.; Oršolić, N.; Mandić, L.; Sadžak, A.; Šegota, S. Anti-Oxidative, Anti-Inflammatory and Anti-Apoptotic Effects of Flavonols: Targeting Nrf2, NF-ΚB and P53 Pathways in Neurodegeneration. Antioxidants 2021, 10, 1628. [Google Scholar] [CrossRef]
- Asgharian, P.; Tazekand, A.P.; Hosseini, K.; Forouhandeh, H.; Ghasemnejad, T.; Ranjbar, M.; Hasan, M.; Kumar, M.; Beirami, S.M.; Tarhriz, V.; et al. Potential Mechanisms of Quercetin in Cancer Prevention: Focus on Cellular and Molecular Targets. Cancer Cell Int. 2022, 22, 257. [Google Scholar] [CrossRef]
- Xiong, F.; Zhang, Y.; Li, T.; Tang, Y.; Song, S.-Y.; Zhou, Q.; Wang, Y. A Detailed Overview of Quercetin: Implications for Cell Death and Liver Fibrosis Mechanisms. Front. Pharmacol. 2024, 15, 1389179. [Google Scholar] [CrossRef]
- de Visser, K.E.; Joyce, J.A. The Evolving Tumor Microenvironment: From Cancer Initiation to Metastatic Outgrowth. Cancer Cell 2023, 41, 374–403. [Google Scholar] [CrossRef] [PubMed]
- El Alaa, R.S.A.; Al-Mannai, W.; Darwish, N.; Al-Mansoori, L. Adipose-Derived Stromal Cells and Cancer-Associated Fibroblasts: Interactions and Implications in Tumor Progression. Int. J. Mol. Sci. 2024, 25, 11558. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Xu, J. Cancer-Associated Fibroblasts: A Versatile Mediator in Tumor Progression, Metastasis, and Targeted Therapy. Cancer Metastasis Rev. 2024, 43, 1095–1116. [Google Scholar] [CrossRef] [PubMed]
- Nedaeinia, R.; Najafgholian, S.; Salehi, R.; Goli, M.; Ranjbar, M.; Nickho, H.; Haghjooy Javanmard, S.A.; Ferns, G.; Manian, M. The Role of Cancer-Associated Fibroblasts and Exosomal MiRNAs-Mediated Intercellular Communication in the Tumor Microenvironment and the Biology of Carcinogenesis: A Systematic Review. Cell Death Discov. 2024, 10, 380. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Gu, W.; Su, H.; Wang, X.; Wang, X.; Zhang, J.; Xu, M.; Sheng, W. Unlocking the Crucial Role of Cancer-Associated Fibroblasts in Tumor Metastasis: Mechanisms and Therapeutic Prospects. J. Adv. Res. 2024, 71, 399–413. [Google Scholar] [CrossRef]
- Lv, K.; He, T. Cancer-Associated Fibroblasts: Heterogeneity, Tumorigenicity and Therapeutic Targets. Mol. Biomed. 2024, 5, 70. [Google Scholar] [CrossRef]
- Ma, Y.; Qi, Y.; Zhou, Z.; Yan, Y.; Chang, J.; Zhu, X.; Han, J.; Wu, H.; Tao, Y.; Fan, F. Shenqi Fuzheng Injection Modulates Tumor Fatty Acid Metabolism to Downregulate MDSCs Infiltration, Enhancing PD-L1 Antibody Inhibition of Intracranial Growth in Melanoma. Phytomedicine 2024, 122, 155171. [Google Scholar] [CrossRef]
- Li, W.; Chen, Z.; Yan, M.; He, P.; Chen, Z.; Dai, H. The Protective Role of Isorhamnetin on Human Brain Microvascular Endothelial Cells from Cytotoxicity Induced by Methylglyoxal and Oxygen–Glucose Deprivation. J. Neurochem. 2016, 136, 651–659. [Google Scholar] [CrossRef]
- Ahn, H.; Lee, G.-S. Isorhamnetin and Hyperoside Derived from Water Dropwort Inhibits Inflammasome Activation. Phytomedicine 2017, 24, 77–86. [Google Scholar] [CrossRef]
- Li, C.; Li, J.; Li, Y.; Li, L.; Luo, Y.; Li, J.; Zhang, Y.; Wang, Y.; Liu, X.; Zhou, X.; et al. Isorhamnetin Promotes MKN-45 Gastric Cancer Cell Apoptosis by Inhibiting PI3K-Mediated Adaptive Autophagy in a Hypoxic Environment. J. Agric. Food Chem. 2021, 69, 8130–8143. [Google Scholar] [CrossRef]
- Chang, Z.; Wang, J.; Jing, Z.; Ma, P.; Xu, Q.; Na, J.; Tian, J.; Ma, X.; Zhou, W.; Zhou, R. Protective Effects of Isorhamnetin on Pulmonary Arterial Hypertension: In Vivo and in Vitro Studies. Phyther. Res. 2020, 34, 2730–2744. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.-T.; Yang, T.-L.; Gong, L.; Wu, P. Isorhamnetin Protects against Hypoxia/Reoxygenation-Induced Injure by Attenuating Apoptosis and Oxidative Stress in H9c2 Cardiomyocytes. Gene 2018, 666, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Chi, G.; Zhong, W.; Liu, Y.; Lu, G.; Lü, H.; Wang, D.; Sun, F. Isorhamnetin Protects Mice from Lipopolysaccharide-Induced Acute Lung Injury via the Inhibition of Inflammatory Responses. Inflamm. Res. 2016, 65, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ma, L.; Wei, Y.; Cui, Y.; Li, X.; Wei, Y.; Zhang, S.; Zhang, L.; Zhou, H.; Wang, G.; et al. Isorhamnetin Alleviates Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting MTOR Signaling Pathway. Immunopharmacol. Immunotoxicol. 2022, 44, 387–399. [Google Scholar] [CrossRef]
- Liu, G.; Jiang, C.; Li, D.; Yao, L.; Lin, Y.; Wang, B.; Qiu, J.; Wang, W.; Wang, W. Isorhamnetin Alleviates Esophageal Mucosal Injury in a Chronic Model of Reflux Esophagitis. Eur. J. Pharmacol. 2019, 864, 172720. [Google Scholar] [CrossRef]
- Shahrezaei, A.; Sohani, M.; Sohouli, M.; Taherkhani, S.; Nasirinezhad, F. The Involvement and Significance of M2 Macrophages in Neuropathic Pain Following Spinal Cord Injury: A Systematic Review. J. Physiol. Sci. 2024, 74, 45. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, K.; Wang, X.; Zhao, Y.; Shi, J.; Liu, Z. Roles of IL-4, IL-13, and Their Receptors in Lung Cancer. J. Interf. Cytok. Res. 2024, 44, 399–407. [Google Scholar] [CrossRef]
- Guo, J.; Yan, W.; Duan, H.; Wang, D.; Zhou, Y.; Feng, D.; Zheng, Y.; Zhou, S.; Liu, G.; Qin, X. Therapeutic Effects of Natural Products on Liver Cancer and Their Potential Mechanisms. Nutrients 2024, 16, 1642. [Google Scholar] [CrossRef]
- Jaramillo, S.; Lopez, S.; Varela, L.M.; Rodriguez-Arcos, R.; Jimenez, A.; Abia, R.; Guillen, R.; Muriana, F.J.G. The Flavonol Isorhamnetin Exhibits Cytotoxic Effects on Human Colon Cancer Cells. J. Agric. Food Chem. 2010, 58, 10869–10875. [Google Scholar] [CrossRef]
- Yuan, J.; Ofengeim, D. A Guide to Cell Death Pathways. Nat. Rev. Mol. Cell Biol. 2024, 25, 379–395. [Google Scholar] [CrossRef]
- Meier, P.; Legrand, A.J.; Adam, D.; Silke, J. Immunogenic Cell Death in Cancer: Targeting Necroptosis to Induce Antitumour Immunity. Nat. Rev. Cancer 2024, 24, 299–315. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sharma, U.; Dangi, N.; Gupta, M.K.; Agrawal, A.; Mali, S.N.; Banik, B.K. A Comprehensive Review on Immunoregulatory Effects of Phytochemicals. Curr. Drug Discov. Technol. 2024, 22, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Jayashankar, B.; Singh, D.; Tanwar, H.; Mishra, K.P.; Murthy, S.; Chanda, S.; Mishra, J.; Tulswani, R.; Misra, K.; Singh, S.B.; et al. Augmentation of Humoral and Cellular Immunity in Response to Tetanus and Diphtheria Toxoids by Supercritical Carbon Dioxide Extracts of Hippophae rhamnoides L. Leaves. Int. Immunopharmacol. 2017, 44, 123–136. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Regulation of Adaptive Immunity by the Innate Immune System. Science 2010, 327, 291–295. [Google Scholar] [CrossRef]
- Zhan, Y.; Ta, W.; Tang, W.; Hua, R.; Wang, J.; Wang, C.; Lu, W. Potential Antiviral Activity of Isorhamnetin against SARS-CoV-2 Spike Pseudotyped Virus in Vitro. Drug Dev. Res. 2021, 82, 1124–1130. [Google Scholar] [CrossRef]
- Teng, B.; Lu, Y.-H.; Wang, Z.-T.; Tao, X.-Y.; Wei, D.-Z. In Vitro Anti-Tumor Activity of Isorhamnetin Isolated from Hippophae rhamnoides L. against BEL-7402 Cells. Pharmacol. Res. 2006, 54, 186–194. [Google Scholar] [CrossRef]
- Skalski, B.; Lis, B.; Pecio, Ł.; Kontek, B.; Olas, B.; Żuchowski, J.; Stochmal, A. Isorhamnetin and Its New Derivatives Isolated from Sea Buckthorn Berries Prevent H2O2/Fe—Induced Oxidative Stress and Changes in Hemostasis. Food Chem. Toxicol. 2019, 125, 614–620. [Google Scholar] [CrossRef]
- ul Islam, B.; Suhail, M.; Khan, M.S.; Ahmad, A.; Zughaibi, T.A.; Husain, F.M.; Rehman, M.T.; Tabrez, S. Flavonoids and PI3K/Akt/MTOR Signaling Cascade: A Potential Crosstalk in Anticancer Treatment. Curr. Med. Chem. 2021, 28, 8083–8097. [Google Scholar] [CrossRef]
- Taher, R.F.; Al-Karmalawy, A.A.; Abd El Maksoud, A.I.; Khalil, H.; Hassan, A.; El-Khrisy, E.-D.A.; El-Kashak, W. Two New Flavonoids and Anticancer Activity of Hymenosporum Flavum: In Vitro and Molecular Docking Studies. J. Herbmed Pharmacol. 2021, 10, 443–458. [Google Scholar] [CrossRef]
- Mohammed, H.A.; Khan, R.A.; Abdel-Hafez, A.A.; Abdel-Aziz, M.; Ahmed, E.; Enany, S.; Mahgoub, S.; Al-Rugaie, O.; Alsharidah, M.; Aly, M.S.A.; et al. Phytochemical Profiling, In Vitro and In Silico Anti-Microbial and Anti-Cancer Activity Evaluations and Staph GyraseB and h-TOP-IIβ Receptor-Docking Studies of Major Constituents of Zygophyllum coccineum L. Aqueous-Ethanolic Extract and Its Subsequent Fra. Molecules 2021, 26, 577. [Google Scholar] [CrossRef]
- Luan, Y.; Luan, Y.; Zhao, Y.; Xiong, F.; Li, Y.; Liu, L.; Cao, Y.; Dai, F. Isorhamnetin in Tsoong Blocks Hsp70 Expression to Promote Apoptosis of Colon Cancer Cells. Saudi J. Biol. Sci. 2019, 26, 1011–1022. [Google Scholar] [CrossRef]
- Wu, Q.; Kroon, P.A.; Shao, H.; Needs, P.W.; Yang, X. Differential Effects of Quercetin and Two of Its Derivatives, Isorhamnetin and Isorhamnetin-3-Glucuronide, in Inhibiting the Proliferation of Human Breast-Cancer MCF-7 Cells. J. Agric. Food Chem. 2018, 66, 7181–7189. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, X.; Chen, C.; Cai, S.; Hu, J. Isorhamnetin Suppresses Colon Cancer Cell Growth through the PI3K-Akt-mTOR Pathway. Ann. Transl. Med. 2022, 10, 1216. [Google Scholar] [CrossRef] [PubMed]
- Hasibuan, P.A.Z.; Simanjuntak, Y.; Hey-Hawkins, E.; Lubis, M.F.; Rohani, A.S.; Park, M.N.; Kim, B.; Syahputra, R.A. Unlocking the Potential of Flavonoids: Natural Solutions in the Fight against Colon Cancer. Biomed. Pharmacother. 2024, 176, 116827. [Google Scholar] [CrossRef] [PubMed]
- Shan, H.U.; Yongchuan, D. Mechanism of Isorhamnetin on Breast Cancer Cells. J. China Pharm. Univ. 2013, 44, 563–567. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Weng, J.-H.; Zeng, S. Simultaneous Determination of Quercetin, Kaempferol and Isorhamnetin Accumulated Human Beast Cancer Cells, by High-Performance Liquid Chromatography. J. Pharm. Biomed. Anal. 2005, 39, 328–333. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Y.; Jiang, X.; Zhang, H.; Gao, Z.; Li, Y.; Fu, R.; Li, L.; Li, J.; Cui, H.; et al. ROS-Mediated Activation and Mitochondrial Translocation of CaMKII Contributes to Drp1-Dependent Mitochondrial Fission and Apoptosis in Triple-Negative Breast Cancer Cells by Isorhamnetin and Chloroquine. J. Exp. Clin. Cancer Res. 2019, 38, 225. [Google Scholar] [CrossRef]
- Antunes-Ricardo, M.; Hernández-Reyes, A.; Uscanga-Palomeque, A.C.; Rodríguez-Padilla, C.; Martínez-Torres, A.C.; Gutiérrez-Uribe, J.A. Isorhamnetin Glycoside Isolated from Opuntia Ficus-Indica (L.) MilI Induces Apoptosis in Human Colon Cancer Cells through Mitochondrial Damage. Chem. Biol. Interact. 2019, 310, 108734. [Google Scholar] [CrossRef]
- Nimmala, S.S.R.; Thupakula, S.; Padiya, R. Targeting Insulin Resistance: Myricetin and Isorhamnetin from Hardwickia Binata, and Luteolin from Hedysarum Alpinum Enhance Glucose Uptake and AMPK Signaling in HepG2 Cells. Nat. Prod. Res. 2024, 1–8. [Google Scholar] [CrossRef]
- Liu, X.; Li, S.; Mei, W.; Liu, X.; Zhou, R. Isorhamnetin Downregulates MMP2 and MMP9 to Inhibit Development of Rheumatoid Arthritis through SRC/ERK/CREB Pathway. Chin. J. Integr. Med. 2024, 30, 299–310. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, X.; Jiang, D.; Tong, J.; Guo, D.; Lin, L. Isorhamnetin plays an analgesic role by regulating NF-κB pathway. J. Third Mil. Med. Univ. 2021, 43, 1366–1371. [Google Scholar]
- Lan, T.; Wang, J.; Zeng, R.; Gao, C.; Liu, X.; Luo, L.; Liang, Y.; Guo, Z.; Wang, W.; Hong, M. Therapeutic Targets and Molecular Mechanisms of Huangqin Decoction in Liver Cancer: A Network Pharmacology and Molecular Docking Approach. J. Herb. Med. 2024, 43, 100822. [Google Scholar] [CrossRef]
- Zhou, Z.; Nan, Y.; Li, X.; Ma, P.; Du, Y.; Chen, G.; Ning, N.; Huang, S.; Gu, Q.; Li, W.; et al. Hawthorn with “Homology of Medicine and Food”: A Review of Anticancer Effects and Mechanisms. Front. Pharmacol. 2024, 15, 1384189. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ai, Z.; Hu, Y.; Ren, G.; Zhang, J.; Tang, P.; Zou, H.; Li, X.; Wang, Y.; Nan, B.; et al. Ginseng Glucosyl Oleanolate Inhibit Cervical Cancer Cell Proliferation and Angiogenesis via PI3K/AKT/HIF-1α Pathway. NPJ Sci. Food 2024, 8, 105. [Google Scholar] [CrossRef]
- Ramachandran, L.; Manu, K.A.; Shanmugam, M.K.; Li, F.; Siveen, K.S.; Vali, S.; Kapoor, S.; Abbasi, T.; Surana, R.; Smoot, D.T.; et al. Isorhamnetin Inhibits Proliferation and Invasion and Induces Apoptosis through the Modulation of Peroxisome Proliferator-Activated Receptor γ Activation Pathway in Gastric Cancer. J. Biol. Chem. 2012, 287, 38028–38040. [Google Scholar] [CrossRef]
- Fang, W.; Du, J.; Nie, M.; Wang, X. Recent Advances in Flavonoid Compounds for the Treatment of Prostate Cancer. Mol. Biol. Rep. 2024, 51, 653. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.; Yamthe, L.R.; Tali, B.T.; Adetunji, C.O.; Rahavian, A.; Mudau, F.N.; Martorell, M.; Setzer, W.N.; Rodrigues, C.F.; et al. Phytochemicals in Prostate Cancer: From Bioactive Molecules to Upcoming Therapeutic Agents. Nutrients 2019, 11, 1483. [Google Scholar] [CrossRef]
- Yang, Z.; Zheng, Y.; Tursumamat, N.; Zhu, M. Synthesis of 3′-O-Alkyl Homologues and a Biotin Probe of Isorhamnetin and Evaluation of Cytotoxic Efficacy on Cancer Cells. Chem. Biodivers. 2021, 18, e2100301. [Google Scholar] [CrossRef]
- Matboli, M.; Ibrahim, D.; Hasanin, A.H.; Hassan, M.K.; Habib, E.K.; Bekhet, M.M.; Afifi, A.M.; Eissa, S. Epigenetic Modulation of Autophagy Genes Linked to Diabetic Nephropathy by Administration of Isorhamnetin in Type 2 Diabetes Mellitus Rats. Epigenomics 2021, 13, 187–202. [Google Scholar] [CrossRef]
- Shimoda, H.; Takeda, S.; Shimizu, N.; Hirano, M.; Hitoe, S. Suppressive Effect of Triterpenoids and a Flavonol Glycoside in Seaberry Extract on Carbacol-Induced Contraction of Bladder Smooth Muscle and TGF-β-Induced Contraction of Collagen Gel Containing Bladder Smooth Muscle Cells. J. Funct. Foods 2017, 31, 152–159. [Google Scholar] [CrossRef]
- Liu, S.; Li, L.; Ren, D. Anti-Cancer Potential of Phytochemicals: The Regulation of the Epithelial-Mesenchymal Transition. Molecules 2023, 28, 5069. [Google Scholar] [CrossRef]
- Hu, S.; Huang, L.; Meng, L.; Sun, H.; Zhang, W.; Xu, Y. Isorhamnetin Inhibits Cell Proliferation and Induces Apoptosis in Breast Cancer via Akt and Mitogen-Activated Protein Kinase Kinase Signaling Pathways. Mol. Med. Rep. 2015, 12, 6745–6751. [Google Scholar] [CrossRef]
- Lv, L.; Du, J.; Wang, D.; Yan, Z. A Comprehensive Study to Investigate the Tumor-Suppressive Role of Radix Bupleuri on Gastric Cancer with Network Pharmacology and Molecular Docking. Drug Des. Dev. Ther. 2024, 18, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xian, Z.; Wu, F.; Cao, H.; Wang, L.; Tang, Q.; Du, Y.; Zheng, X. Traditional Chinese Medicine Combined with Chemotherapy in the Treatment of Advanced Non-Small Cell Lung Cancer: Key Drug Screening and Mechanism Analysis. Naunyn. Schmiedebergs. Arch. Pharmacol. 2024, 398, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Das, A.K.; Bhattacharya, S.; Gachhui, R.; Sil, P.C. Isorhamnetin Exerts Anti-Tumor Activity in DEN + CCl4-Induced HCC Mice. Med. Oncol. 2023, 40, 188. [Google Scholar] [CrossRef] [PubMed]
- Kalaivani, D.; Arun, V. Antimelanogenic and Anticancer Activity of Isorhamnetin Isolated from Acalypha Indica Linn. on A375 Cell Line. Curr. Trends Biotechnol. Pharm. 2022, 16, 445–455. [Google Scholar] [CrossRef]
- Du, Y.; Jia, C.; Liu, Y.; Li, Y.; Wang, J.; Sun, K. Isorhamnetin Enhances the Radiosensitivity of A549 Cells Through Interleukin-13 and the NF-ΚB Signaling Pathway. Front. Pharmacol. 2021, 11, 610772. [Google Scholar] [CrossRef]
- Duan, R.; Liang, X.; Chai, B.; Zhou, Y.; Du, H.; Suo, Y.; Chen, Z.; Li, Q.; Huang, X. Isorhamnetin Induces Melanoma Cell Apoptosis via the PI3K/Akt and NF-ΚB Pathways. BioMed Res. Int. 2020, 2020, 1057943. [Google Scholar] [CrossRef]
- Antunes-Ricardo, M.; Guardado-Félix, D.; Rocha-Pizaña, M.R.; Garza-Martínez, J.; Acevedo-Pacheco, L.; Gutiérrez-Uribe, J.A.; Villela-Castrejón, J.; López-Pacheco, F.; Serna-Saldívar, S.O. Opuntia Ficus-Indica Extract and Isorhamnetin-3-O-Glucosyl-Rhamnoside Diminish Tumor Growth of Colon Cancer Cells Xenografted in Immune-Suppressed Mice through the Activation of Apoptosis Intrinsic Pathway. Plant Foods Hum. Nutr. 2021, 76, 434–441. [Google Scholar] [CrossRef]
- Wang, M.; Xu, Z.; Cai, Q.; Deng, Y.; Shi, W.; Zhou, H.; Wang, D.; Li, J. Isorhamnetin Inhibits Progression of Ovarian Cancer by Targeting ESR1. Ann. Transl. Med. 2022, 10, 1216. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, B.; Pu, N.; Ran, X.; Lian, T.; Cai, Y.; Xing, W.; Sun, K. Isorhamnetin Suppresses Human Gastric Cancer Cell Proliferation through Mitochondria-Dependent Apoptosis. Molecules 2022, 27, 5191. [Google Scholar] [CrossRef] [PubMed]
- Greifová, H.; Tokárová, K.; Jambor, T.; Štefunková, N.; Speváková, I.; Dupák, R.; Balytska, O.; Bažány, D.; Capcarová, M.; Lukáč, N. Isorhamnetin Influences the Viability, Superoxide Production and Interleukin-8 Biosynthesis of Human Colorectal Adenocarcinoma HT-29 Cells In Vitro. Life 2023, 13, 1921. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Seo, K.; Ki, S.H.; Shin, S.M. Isorhamnetin Inhibits Reactive Oxygen Species-Dependent Hypoxia Inducible Factor (HIF)- 1α Accumulation. Biol. Pharm. Bull. 2016, 39, 1830–1838. [Google Scholar] [CrossRef]
- Mirazimi, S.M.A.; Dashti, F.; Tobeiha, M.; Shahini, A.; Jafari, R.; Khoddami, M.; Sheida, A.H.; EsnaAshari, P.; Aflatoonian, A.H.; Elikaii, F.; et al. Application of Quercetin in the Treatment of Gastrointestinal Cancers. Front. Pharmacol. 2022, 13, 860209. [Google Scholar] [CrossRef]
- Yan, M.; Wu, S.; Wang, Y.; Liang, M.; Wang, M.; Hu, W.; Yu, G.; Mao, Z.; Huang, F.; Zhou, J. Recent Progress of Supramolecular Chemotherapy Based on Host–Guest Interactions. Adv. Mater. 2024, 36, 2304249. [Google Scholar] [CrossRef]
- Khajuria, R.; Sarwar, A. Review of Reinforcement Learning Applications in Segmentation, Chemotherapy, and Radiotherapy of Cancer. Micron 2024, 178, 103583. [Google Scholar] [CrossRef]
- Mallikarjun, K.S.; Eldaya, R.W.; Miller-Thomas, M.M.; Orlowski, H.L.; Parsons, M.S. Good Gone Bad: Complications of Chemotherapy, Immunotherapy, and Radiotherapy on the CNS. Curr. Probl. Diagn. Radiol. 2024, 53, 133–149. [Google Scholar] [CrossRef]
- Fromm, J.; Klein, A.; Kirilova, M.; Lindner, L.H.; Nachbichler, S.; Holzapfel, B.M.; Goller, S.S.; Knösel, T.; Dürr, H.R. The Effect of Chemo- and Radiotherapy on Tumor Necrosis in Soft Tissue Sarcoma– Does It Influence Prognosis? BMC Cancer 2024, 24, 303. [Google Scholar] [CrossRef]
- Meyer, C.T.; Wooten, D.J.; Paudel, B.B.; Bauer, J.; Hardeman, K.N.; Westover, D.; Lovly, C.M.; Harris, L.A.; Tyson, D.R.; Quaranta, V. Quantifying Drug Combination Synergy along Potency and Efficacy Axes. Cell Syst. 2019, 8, 97–108.e16. [Google Scholar] [CrossRef]
- Zhang, B.Y.; Wang, Y.M.; Gong, H.; Zhao, H.; Lv, X.Y.; Yuan, G.H.; Han, S.R. Isorhamnetin Flavonoid Synergistically Enhances the Anticancer Activity and Apoptosis Induction by Cisplatin and Carboplatin in Non-Small Cell Lung Carcinoma (NSCLC). Int. J. Clin. Exp. Pathol. 2015, 8, 25. [Google Scholar] [PubMed]
- Yang, B.; Zhang, F.; Yuan, W.; Du, L.; Jiang, X. Preparation of Isorhamnetin Nanoparticles and Their Targeting Efficiency to Nasopharynx Cancer. J. Nanosci. Nanotechnol. 2021, 21, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Shanmugasundaram, P.; Komala, M.; Bhargavi, B.; Padmavathy, J. Nanoparticle Formulation of Bioflavonoids for Enhanced Anti-Cancer Activity. Int. J. Appl. Pharm. 2020, 12, 29–35. [Google Scholar] [CrossRef]
- García-Valderrama, E.J.; Mamidi, N.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; Del Angel-Sanchez, K.; Elías-Zúñiga, A. Engineering and Evaluation of Forcespun Gelatin Nanofibers as an Isorhamnetin Glycosides Delivery System. Pharmaceutics 2022, 14, 1116. [Google Scholar] [CrossRef]
- Aboulthana, W.M.K.; Refaat, E.; Khaled, S.E.; Ibrahim, N.E.-S.; Youssef, A.M. Metabolite Profiling and Biological Activity Assessment of Casuarina Equisetifolia Bark after Incorporating Gold Nanoparticles. Asian Pac. J. Cancer Prev. 2022, 23, 3457–3471. [Google Scholar] [CrossRef]
- Alhamdi, H.W.; Mokhtar, F.A.; Ridouane, F.L.; Shati, A.A.; Elbehairi, S.E.I.; Fahmy, L.I.; Alfaifi, M.Y.; Sedky, N.K.; Fahmy, H.A. Computational Metal-Flavonoids Complexes Presentation of Greenly Synthesized Silver Nanoparticles Combined Flavonoids from Lens culinaris L. as Anticancer Agents Using BcL-2 and IspC Proteins. Artif. Cells Nanomed. Biotechnol. 2024, 52, 529–550. [Google Scholar] [CrossRef]
- Mathur, J.; Goswami, P. Positive Impact of Green Synthesized Silica Nanoparticles in Plant Growth Promotion and Physiological Responses of Eruca Sativa Mill. J. Soil Sci. Plant Nutr. 2024, 24, 2263–2275. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, X.; Liu, Y.; Ma, Y.; Wang, X.; Wang, X.; Xu, L. Chemiluminescence of Off-Line and on-Line Gold Nanoparticle-Catalyzed Luminol System in the Presence of Flavonoid. Luminescence 2017, 32, 666–673. [Google Scholar] [CrossRef]
- Abdul-Razek, N.; Khalil, R.G.; Abdel-Latif, M.; Kamel, M.M.; Alhazza, I.M.; Awad, E.M.; Ebaid, H.; Abuelsaad, A.S.A. Investigating the Tumor-Suppressive, Antioxidant Effects and Molecular Binding Affinity of Quercetin-Loaded Selenium Nanoparticles in Breast Cancer Cells. Bionanoscience 2024, 15, 135. [Google Scholar] [CrossRef]
- Han, H.S.; Jung, J.S.; Jeong, Y.-I.; Choi, K.C. Biological Synthesis of Copper Nanoparticles Using Edible Plant Allium Monanthum: Characterization of Antibacterial, Antioxidant, and Anti-Inflammatory Properties Using In Silico Molecular Docking Analysis. Materials 2023, 16, 6669. [Google Scholar] [CrossRef]
- Shree, D.; Patra, C.N.; Sahoo, B.M. Fabrication and Applications of Polymeric Nanoparticles for Herbal Drug Delivery and Targeting. Curr. Tradit. Med. 2023, 9, 23–33. [Google Scholar] [CrossRef]
- Hu, J.; Teng, B.; Xu, Z.; Wan, Y.; Jin, G. A Porous Form Coomassie Brilliant Blue G250-Isorhamnetin Fluorescent Composite Coated with Acrylic Resin for Tumor Cell Imaging. Front. Chem. 2023, 11, 1260533. [Google Scholar] [CrossRef]
- Tritean, N.; Dimitriu, L.; Dima, Ș.-O.; Stoica, R.; Trică, B.; Ghiurea, M.; Moraru, I.; Cimpean, A.; Oancea, F.; Constantinescu-Aruxandei, D. Cytocompatibility, Antimicrobial and Antioxidant Activity of a Mucoadhesive Biopolymeric Hydrogel Embedding Selenium Nanoparticles Phytosynthesized by Sea Buckthorn Leaf Extract. Pharmaceuticals 2024, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Zou, F.; Zhao, H.; Ma, A.; Song, D.; Zhang, X.; Zhao, X. Preparation of an Isorhamnetin Phospholipid Complex for Improving Solubility and Anti-Hyperuricemia Activity. Pharm. Dev. Technol. 2022, 27, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jiang, D. Synthesis, Bioactivity Evaluation, and Cell Imaging of Isorhamine Zwitterionic Polymers. Russ. J. Bioorg. Chem. 2023, 49, 1309–1318. [Google Scholar] [CrossRef]
- Yayintas, O.T.; Demir, N.; Canbolat, F.; Ayna, T.K.; Pehlivan, M. Characterization, Biological Activity, and Anticancer Effect of Green-Synthesized Gold Nanoparticles Using Nasturtium officinale L. BMC Complement. Med. Ther. 2024, 24, 346. [Google Scholar] [CrossRef]
- Macêdo, H.L.; de Oliveira, L.L.; de Oliveira, D.N.; Lima, K.F.; Cavalcanti, I.M.; Campos, L.A. Nanostructures for Delivery of Flavonoids with Antibacterial Potential against Klebsiella Pneumoniae. Antibiotics 2024, 13, 844. [Google Scholar] [CrossRef]
- Wang, T.; Li, Q.; Bi, K. Bioactive Flavonoids in Medicinal Plants: Structure, Activity and Biological Fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Hämäläinen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-Inflammatory Effects of Flavonoids: Genistein, Kaempferol, Quercetin, and Daidzein Inhibit STAT-1 and NF-ΚB Activations, Whereas Flavone, Isorhamnetin, Naringenin, and Pelargonidin Inhibit Only NF-ΚB Activation along with Their Inhibitory Effect on I. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar] [CrossRef]
- Zuo, A.; Yanying, Y.; Li, J.; Binbin, X.; Xiongying, Y.; Yan, Q.; Shuwen, C. Study on the Relation of Structure and Antioxidant Activity of Isorhamnetin, Quercetin, Phloretin, Silybin and Phloretin Isonicotinyl Hydrazone. Free Radic. Antioxid. 2011, 1, 39–47. [Google Scholar] [CrossRef]
- La, X.; Zhang, Z.; Dong, C.; Li, H.; He, X.; Kang, Y.; Wu, C.; Li, Z. Isorhamnetin in Quinoa Whole-Grain Flavonoids Intervenes in Non-Alcoholic Fatty Liver Disease by Modulating Bile Acid Metabolism through Regulation of FXR Expression. Foods 2024, 13, 3076. [Google Scholar] [CrossRef]
- Boesch-Saadatmandi, C.; Loboda, A.; Wagner, A.E.; Stachurska, A.; Jozkowicz, A.; Dulak, J.; Döring, F.; Wolffram, S.; Rimbach, G. Effect of Quercetin and Its Metabolites Isorhamnetin and Quercetin-3-Glucuronide on Inflammatory Gene Expression: Role of MiR-155. J. Nutr. Biochem. 2011, 22, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S. Anti-Inflammatory Action of Isorhamnetin. Inflammation 2014, 37, 1200–1201. [Google Scholar] [CrossRef] [PubMed]
- Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; Martínez-Vitela, C.; Serna-Saldívar, S.O. Topical Anti-Inflammatory Effects of Isorhamnetin Glycosides Isolated from Opuntia Ficus-Indica. BioMed Res. Int. 2015, 2015, 847320. [Google Scholar] [CrossRef] [PubMed]
- Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; López-Pacheco, F.; Alvarez, M.M.; Serna-Saldívar, S.O. In Vivo Anti-Inflammatory Effects of Isorhamnetin Glycosides Isolated from Opuntia Ficus-Indica (L.) Mill Cladodes. Ind. Crops Prod. 2015, 76, 803–808. [Google Scholar] [CrossRef]
- Bouzaid, H.; Espírito Santo, L.; Ferreira, D.M.; Machado, S.; Costa, A.S.G.; Dias, M.I.; Calhelha, R.C.; Barros, L.; Chater, O.; Rodi, Y.K.; et al. Detailed Phytochemical Composition, Cyto-/Hepatotoxicity, and Antioxidant/Anti-Inflammatory Profile of Moroccan Spices: A Study on Coriander, Caraway, and Mystical Cumin. Molecules 2024, 29, 3485. [Google Scholar] [CrossRef]
- Păcularu-Burada, B.; Cîrîc, A.-I.; Begea, M. Anti-Aging Effects of Flavonoids from Plant Extracts. Foods 2024, 13, 2441. [Google Scholar] [CrossRef]
- Belkhir, S.; Debbache-Benaida, N.; Medjahed, Z.; Atia, A.; Ayouni, K.; Boudjouan, F.; Saidene, N.; Benamrouche, N.; Dumas, F.; Atmani-Kilani, D. Interaction of Clematis Flammula Extracts with Membrane Models: Characterization of Anti-Inflammatory and Antibacterial Activities. Plant Biosyst.—Int. J. Deal. All Asp. Plant Biol. 2024, 158, 130–141. [Google Scholar] [CrossRef]
- Chauhan, A.K.; Kim, J.; Lee, Y.; Balasubramanian, P.K.; Kim, Y. Isorhamnetin Has Potential for the Treatment of Escherichia Coli-Induced Sepsis. Molecules 2019, 24, 3984. [Google Scholar] [CrossRef]
- Hahn, D.; Shin, S.H.; Bae, J.-S. Natural Antioxidant and Anti-Inflammatory Compounds in Foodstuff or Medicinal Herbs Inducing Heme Oxygenase-1 Expression. Antioxidants 2020, 9, 1191. [Google Scholar] [CrossRef]
- Guo, B.; Yu, Y.; Wang, M.; Li, R.; He, X.; Tang, S.; Liu, Q.; Mao, Y. Targeting the JAK2/STAT3 Signaling Pathway with Natural Plants and Phytochemical Ingredients: A Novel Therapeutic Method for Combatting Cardiovascular Diseases. Biomed. Pharmacother. 2024, 172, 116313. [Google Scholar] [CrossRef] [PubMed]
- Dutra, R.C.; Campos, M.M.; Santos, A.R.S.; Calixto, J.B. Medicinal Plants in Brazil: Pharmacological Studies, Drug Discovery, Challenges and Perspectives. Pharmacol. Res. 2016, 112, 4–29. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.L.; Tai, C.-J.; Chattopadhyay, K.; Hu, X.-Y.; Heinrich, M. Editorial: Clinical Phytopharmacology. Front. Pharmacol. 2024, 14, 1353483. [Google Scholar] [CrossRef] [PubMed]
- Parveen, A.; Parveen, B.; Parveen, R.; Ahmad, S. Challenges and Guidelines for Clinical Trial of Herbal Drugs. J. Pharm. Bioallied Sci. 2015, 7, 329–333. [Google Scholar] [CrossRef]
- Singla, R.K.; Wang, X.; Gundamaraju, R.; Joon, S.; Tsagkaris, C.; Behzad, S.; Khan, J.; Gautam, R.; Goyal, R.; Rakmai, J.; et al. Natural Products Derived from Medicinal Plants and Microbes Might Act as a Game-Changer in Breast Cancer: A Comprehensive Review of Preclinical and Clinical Studies. Crit. Rev. Food Sci. Nutr. 2023, 63, 11880–11924. [Google Scholar] [CrossRef]
- Qu, N.; Song, K.; Ji, Y.; Liu, M.; Chen, L.; Lee, R.J.; Teng, L. Albumin Nanoparticle-Based Drug Delivery Systems. Int. J. Nanomed. 2024, 19, 6945–6980. [Google Scholar] [CrossRef]
- Ajith, S.; Almomani, F.; Elhissi, A.; Husseini, G.A. Nanoparticle-Based Materials in Anticancer Drug Delivery: Current and Future Prospects. Heliyon 2023, 9, e21227. [Google Scholar] [CrossRef]
- Shivani Singh, G.; Narwal, S.; Chopra, B.; Dhingra, A.K. Quercetin-Based Nanoformulation: A Potential Approach for Cancer Treatment. Anti-Cancer Agents Med. Chem. (Formerly Curr. Med. Chem.-Anti-Cancer Agents) 2023, 23, 1983–2007. [Google Scholar] [CrossRef]
- Akanchise, T.; Angelova, A. Ginkgo Biloba and Long COVID: In Vivo and In Vitro Models for the Evaluation of Nanotherapeutic Efficacy. Pharmaceutics 2023, 15, 1562. [Google Scholar] [CrossRef]
- Wang, H.; Cui, Y.; Fu, Q.; Deng, B.; Li, G.; Yang, J.; Wu, T.; Xie, Y. A Phospholipid Complex to Improve the Oral Bioavailability of Flavonoids. Drug Dev. Ind. Pharm. 2015, 41, 1693–1703. [Google Scholar] [CrossRef]
- Rathod, S.; Arya, S.; Kanike, S.; Shah, S.A.; Bahadur, P.; Tiwari, S. Advances on Nanoformulation Approaches for Delivering Plant-Derived Antioxidants: A Case of Quercetin. Int. J. Pharm. 2022, 625, 122093. [Google Scholar] [CrossRef]
- Dajas, F.; Abin-Carriquiry, J.A.; Arredondo, F.; Blasina, F.; Echeverry, C.; Martínez, M.; Rivera, F.; Vaamonde, L. Quercetin in Brain Diseases: Potential and Limits. Neurochem. Int. 2015, 89, 140–148. [Google Scholar] [CrossRef]
- Fusi, F.; Spiga, O.; Trezza, A.; Sgaragli, G.; Saponara, S. The Surge of Flavonoids as Novel, Fine Regulators of Cardiovascular Cav Channels. Eur. J. Pharmacol. 2017, 796, 158–174. [Google Scholar] [CrossRef]
- Malik, J.; Mandal, S.C.; Choudhary, S.; Parihar, S.; Rahamathulla, M. Herbal Medicines for Management of Alzheimer’s Disease BT—Role of Herbal Medicines: Management of Lifestyle Diseases; Dhara, A.K., Mandal, S.C., Eds.; Springer Nature Singapore: Singapore, 2023; pp. 231–250. ISBN 978-981-99-7703-1. [Google Scholar]
- Yuan, D.; Guo, Y.; Pu, F.; Yang, C.; Xiao, X.; Du, H.; He, J.; Lu, S. Opportunities and Challenges in Enhancing the Bioavailability and Bioactivity of Dietary Flavonoids: A Novel Delivery System Perspective. Food Chem. 2024, 430, 137115. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement Strategies for the Oral Bioavailability of Poorly Water-Soluble Flavonoids: An Overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef] [PubMed]
- Zverev, Y.F.; Rykunova, A.Y. Modern Nanocarriers as a Factor in Increasing the Bioavailability and Pharmacological Activity of Flavonoids. Appl. Biochem. Microbiol. 2022, 58, 1002–1020. [Google Scholar] [CrossRef] [PubMed]
- Mohite, P.; Puri, A.; Bharati, D.; Singh, S. Polyphenol-Encapsulated Nanoparticles for the Treatment of Chronic Metabolic Diseases. In Role of Flavonoids in Chronic Metabolic Diseases; From Bench to Clinic; Mishra, N., Ashique, S., Gowda, B.H.J., Farid, A., Garg, A., Eds.; Wiley-Scrivener: Hoboken, NJ, USA, 2024; pp. 375–416. ISBN 9781394238071. [Google Scholar]
- Chen, H.; Wang, G.; Li, X.; Wang, J.; Wang, X.; Wang, Y.; Liu, Z.; Liu, J.; Ding, Y.; Guo, J.; et al. Multi-Functional D-Alpha-Tocopheryl Polyethylene Glycol Succinate Surface Modified Nanocrystals Improve the Stability and Oral Bioavailability of Pueraria Flavonoids. J. Drug Deliv. Sci. Technol. 2024, 95, 105623. [Google Scholar] [CrossRef]
- Ciceu, A.; Fenyvesi, F.; Hermenean, A.; Ardelean, S.; Dumitra, S.; Puticiu, M. Advancements in Plant-Based Therapeutics for Hepatic Fibrosis: Molecular Mechanisms and Nanoparticulate Drug Delivery Systems. Int. J. Mol. Sci. 2024, 25, 9346. [Google Scholar] [CrossRef]
- Li, N.; Wang, M.; Lyu, Z.; Shan, K.; Chen, Z.; Chen, B.; Chen, Y.; Hu, X.; Dou, B.; Zhang, J.; et al. Medicinal Plant-Based Drug Delivery System for Inflammatory Bowel Disease. Front. Pharmacol. 2023, 14, 1158945. [Google Scholar] [CrossRef]
- Sezgin-Bayindir, Z.; Losada-Barreiro, S.; Bravo-Díaz, C.; Sova, M.; Kristl, J.; Saso, L. Nanotechnology-Based Drug Delivery to Improve the Therapeutic Benefits of NRF2 Modulators in Cancer Therapy. Antioxidants 2021, 10, 685. [Google Scholar] [CrossRef]
- Sharma, A.; Wairkar, S. Flavonoids for Treating Pulmonary Fibrosis: Present Status and Future Prospects. Phyther. Res. 2024, 38, 4406–4423. [Google Scholar] [CrossRef]
- Majid, I.; Majid, D.; Makroo, H.A.; Dar, B.N. Enhancing the Bioavailability and Gut Health Benefits of Quercetin from Sprouted Onions: A Comprehensive Review in the Context of Food-Derived Bioactives. Food Chem. Adv. 2024, 4, 100725. [Google Scholar] [CrossRef]
- Attar, E.S.; Chaudhari, V.H.; Deokar, C.G.; Dyawanapelly, S.; Devarajan, P.V. Nano Drug Delivery Strategies for an Oral Bioenhanced Quercetin Formulation. Eur. J. Drug Metab. Pharmacokinet. 2023, 48, 495–514. [Google Scholar] [CrossRef]
- Shree Harini, K.; Ezhilarasan, D. Flavonoids-Based Nanomedicines for the Treatment of Liver Fibrosis: A Recent Progress. J. Drug Deliv. Sci. Technol. 2024, 93, 105467. [Google Scholar] [CrossRef]
- Gervasi, T.; Calderaro, A.; Barreca, D.; Tellone, E.; Trombetta, D.; Ficarra, S.; Smeriglio, A.; Mandalari, G.; Gattuso, G. Biotechnological Applications and Health-Promoting Properties of Flavonols: An Updated View. Int. J. Mol. Sci. 2022, 23, 1710. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Anand, A.; Halagali, P.; Inamdar, A.; Pathak, R.; Taghizadeh-Hesary, F.; Ashique, S. Advancement of Nanoengineered Flavonoids for Chronic Metabolic Diseases. In Role of Flavonoids in Chronic Metabolic Diseases: From Bench to Clinic; Mishra, N., Ashique, S., Gowda, B.H.J., Farid, A., Garg, A., Eds.; Scrivener Publishing LLC: Austin, TX, USA, 2024; pp. 459–510. ISBN 9781394238071. [Google Scholar]
- Patel, D.; Sethi, N.; Patel, P.; Shah, S.; Patel, K. Exploring the Potential of P-Glycoprotein Inhibitors in the Targeted Delivery of Anti-Cancer Drugs: A Comprehensive Review. Eur. J. Pharm. Biopharm. 2024, 198, 114267. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.; Ajazuddin; Patel, R.J.; Saraf, S.; Saraf, S. Recent Expansion of Pharmaceutical Nanotechnologies and Targeting Strategies in the Field of Phytopharmaceuticals for the Delivery of Herbal Extracts and Bioactives. J. Control. Release 2016, 241, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, B.; Wan, Y.; Sun, Y.; Wang, L.; Sun, J.; Li, C. Drug Delivery Based Pharmacological Enhancement and Current Insights of Quercetin with Therapeutic Potential against Oral Diseases. Biomed. Pharmacother. 2020, 128, 110372. [Google Scholar] [CrossRef]
- Vlachopoulos, A.; Karlioti, G.; Balla, E.; Daniilidis, V.; Kalamas, T.; Stefanidou, M.; Bikiaris, N.D.; Christodoulou, E.; Koumentakou, I.; Karavas, E.; et al. Poly(Lactic Acid)-Based Microparticles for Drug Delivery Applications: An Overview of Recent Advances. Pharmaceutics 2022, 14, 359. [Google Scholar] [CrossRef]
- Samborska, K.; Boostani, S.; Geranpour, M.; Hosseini, H.; Dima, C.; Khoshnoudi-Nia, S.; Rostamabadi, H.; Falsafi, S.R.; Shaddel, R.; Akbari-Alavijeh, S.; et al. Green Biopolymers from By-Products as Wall Materials for Spray Drying Microencapsulation of Phytochemicals. Trends Food Sci. Technol. 2021, 108, 297–325. [Google Scholar] [CrossRef]
- Khursheed, R.; Singh, S.K.; Wadhwa, S.; Kapoor, B.; Gulati, M.; Kumar, R.; Ramanunny, A.K.; Awasthi, A.; Dua, K. Treatment Strategies against Diabetes: Success so Far and Challenges Ahead. Eur. J. Pharmacol. 2019, 862, 172625. [Google Scholar] [CrossRef]
- Birnbaum, D.T.; Brannon-Peppas, L. Microparticle Drug Delivery Systems BT—Drug Delivery Systems in Cancer Therapy; Brown, D.M., Ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 117–135. ISBN 978-1-59259-427-6. [Google Scholar]
- Kohane, D.S. Microparticles and Nanoparticles for Drug Delivery. Biotechnol. Bioeng. 2007, 96, 203–209. [Google Scholar] [CrossRef]
- Kállai-Szabó, N.; Farkas, D.; Lengyel, M.; Basa, B.; Fleck, C.; Antal, I. Microparticles and Multi-Unit Systems for Advanced Drug Delivery. Eur. J. Pharm. Sci. 2024, 194, 106704. [Google Scholar] [CrossRef] [PubMed]
- Di Marzio, L.; Ventura, C.A.; Cosco, D.; Paolino, D.; Di Stefano, A.; Stancanelli, R.; Tommasini, S.; Cannavà, C.; Celia, C.; Fresta, M. Nanotherapeutics for Anti-Inflammatory Delivery. J. Drug Deliv. Sci. Technol. 2016, 32, 174–191. [Google Scholar] [CrossRef]
- Alizadeh-Sani, M.; Mohammadian, E.; Rhim, J.-W.; Jafari, S.M. PH-Sensitive (Halochromic) Smart Packaging Films Based on Natural Food Colorants for the Monitoring of Food Quality and Safety. Trends Food Sci. Technol. 2020, 105, 93–144. [Google Scholar] [CrossRef]
- Chanaj-Kaczmarek, J.; Paczkowska, M.; Osmałek, T.; Kaproń, B.; Plech, T.; Szymanowska, D.; Karaźniewicz-Łada, M.; Kobus-Cisowska, J.; Cielecka-Piontek, J. Hydrogel Delivery System Containing Calendulae Flos Lyophilized Extract with Chitosan as a Supporting Strategy for Wound Healing Applications. Pharmaceutics 2020, 12, 634. [Google Scholar] [CrossRef]
- Kar, K. Functionalization of Food Polyphenols for Nanodeliveries. In Polyphenols; Rudrapal, M., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2023; pp. 134–154. ISBN 9781394188864. [Google Scholar]
- Jia, S.; Huang, S.; Jimo, R.; AXi, Y.; Lu, Y.; Kong, Z.; Ma, J.; Li, H.; Luo, X.; Qu, Y.; et al. In-Situ Forming Carboxymethyl Chitosan Hydrogel Containing Paeonia Suffruticosa Andr. Leaf Extract for Mixed Infectious Vaginitis Treatment by Reshaping the Micro-Biota. Carbohydr. Polym. 2024, 339, 122255. [Google Scholar] [CrossRef]
- Frenț, O.-D.; Stefan, L.; Morgovan, C.M.; Duteanu, N.; Dejeu, I.L.; Marian, E.; Vicaș, L.; Manole, F. A Systematic Review: Quercetin—Secondary Metabolite of the Flavonol Class, with Multiple Health Benefits and Low Bioavailability. Int. J. Mol. Sci. 2024, 25, 12091. [Google Scholar] [CrossRef]
- Xiong, X.; Liu, Z.; Che, X.; Zhang, X.; Li, X.; Gao, W. Chemical Composition, Pharmacological Activity and Development Strategies of Rubus Chingii: A Review. Chin. Herb. Med. 2024, 16, 313–326. [Google Scholar] [CrossRef]
- Mancera-Andrade, E.I.; Parsaeimehr, A.; Ruiz-Ruiz, F.; Rorrer, G.L.; González-Valdez, J.; Iqbal, H.M.N.; Parra-Saldivar, R. Isorhamnetin Encapsulation into Biogenic Silica from Cyclotella sp. Using a Microfluidic Device for Drug Delivery Applications. Biocatal. Agric. Biotechnol. 2019, 19, 101175. [Google Scholar] [CrossRef]
- Duque-Soto, C.; Leyva-Jiménez, F.J.; Quirantes-Piné, R.; López-Bascón, M.A.; Lozano-Sánchez, J.; Borrás-Linares, I. Evaluation of Olive Leaf Phenolic Compounds’ Gastrointestinal Stability Based on Co-Administration and Microencapsulation with Non-Digestible Carbohydrates. Nutrients 2024, 16, 93. [Google Scholar] [CrossRef]
- Gargouri, W.; Elleuche, M.; Fernández-Muiño, M.A.; Sancho, M.T.; Osés, S.M. Microencapsulated Propolis Powder: A Promising Ingredient of Chewing Gum. Powder Technol. 2024, 440, 119777. [Google Scholar] [CrossRef]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Sreedhar, R.; Giridharan, V.V.; Watanabe, K. Molecular Targets of Quercetin with Anti-Inflammatory Properties in Atopic Dermatitis. Drug Discov. Today 2016, 21, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-K. Natural Products in Cosmetics. Nat. Prod. Bioprospect. 2022, 12, 40. [Google Scholar] [CrossRef]
- Galasso, C.; Ruocco, N.; Mutalipassi, M.; Barra, L.; Costa, V.; Giommi, C.; Dinoi, A.; Genovese, M.; Pica, D.; Romano, C.; et al. Marine Polysaccharides, Proteins, Lipids, and Silica for Drug Delivery Systems: A Review. Int. J. Biol. Macromol. 2023, 253, 127145. [Google Scholar] [CrossRef] [PubMed]
- Moazed, K.T. Chemicals and Small Molecules for Iris Color Modification BT—Quest for Eye Color Modification; Moazed, K.T., Ed.; Springer Nature Switzerland: Cham, Switzerland, 2024; pp. 95–126. ISBN 978-3-031-64322-4. [Google Scholar]
- Wu, J.; Wang, X.; Zhao, Y.; Hou, Y.; Gong, P. Overview of CFTR Activators and Their Recent Studies for Dry Eye Disease: A Review. RSC Med. Chem. 2023, 14, 2459–2472. [Google Scholar] [CrossRef]
- Sharma, A.R.; Lee, Y.-H.; Bat-Ulzii, A.; Chatterjee, S.; Bhattacharya, M.; Chakraborty, C.; Lee, S.-S. Bioactivity, Molecular Mechanism, and Targeted Delivery of Flavonoids for Bone Loss. Nutrients 2023, 15, 919. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; Tan, W.; Ma, Z.; Yu, T.; Zhao, T. Preparation and Evaluation of Thesium Chinense Extract Loaded Polyvinyl Alcohol/Sodium Alginate Hydrogel for Wound Healing. Polym. Bull. 2024, 81, 6225–6247. [Google Scholar] [CrossRef]
- Subramanian, S.; Duraipandian, C.; Alsayari, A.; Ramachawolran, G.; Wong, L.S.; Sekar, M.; Gan, S.H.; Subramaniyan, V.; Seethalakshmi, S.; Jeyabalan, S.; et al. Wound Healing Properties of a New Formulated Flavonoid-Rich Fraction from Dodonaea Viscosa Jacq. Leaves Extract. Front. Pharmacol. 2023, 14, 1096905. [Google Scholar] [CrossRef]
- Chin, C.-Y.; Jalil, J.; Ng, P.Y.; Ng, S.-F. Development and Formulation of Moringa Oleifera Standardised Leaf Extract Film Dressing for Wound Healing Application. J. Ethnopharmacol. 2018, 212, 188–199. [Google Scholar] [CrossRef]
- Dewanjee, S.; Chakraborty, P.; Bhattacharya, H.; Singh, S.K.; Dua, K.; Dey, A.; Jha, N.K. Recent Advances in Flavonoid-Based Nanocarriers as an Emerging Drug Delivery Approach for Cancer Chemotherapy. Drug Discov. Today 2023, 28, 103409. [Google Scholar] [CrossRef]
- Kaushal, N.; Singh, M.; Singh Sangwan, R. Flavonoids: Food Associations, Therapeutic Mechanisms, Metabolism and Nanoformulations. Food Res. Int. 2022, 157, 111442. [Google Scholar] [CrossRef] [PubMed]
- Alshweiat, A.; Jaber, M.; Abuawad, A.; Athamneh, T.; Oqal, M. Recent Insights into Nanoformulation Delivery Systems of Flavonoids against Glioblastoma. J. Drug Deliv. Sci. Technol. 2024, 91, 105271. [Google Scholar] [CrossRef]
- Chandekar, L.; Katgeri, R.; Takke, A. The Potential Clinical Uses and Nanoformulation Strategies of Kaempferol, a Dietary Flavonoid. Rev. Bras. Farmacogn. 2022, 32, 693–707. [Google Scholar] [CrossRef]
- Alshehri, M.A.; Panneerselvam, C. Development of Quercetin Loaded Biosynthesized Chitosan Grafted Iron Oxide Nanoformulation and Their Antioxidant, Antibacterial, and Anti-Cancer Properties. J. Drug Deliv. Sci. Technol. 2024, 101, 106247. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, Y.; Li, Y.; Rui, Y.; Zhang, P. Enhancing the Efficacy of Active Pharmaceutical Ingredients in Medicinal Plants through Nanoformulations: A Promising Field. Nanomaterials 2024, 14, 1598. [Google Scholar] [CrossRef]
- Mod Razif, M.R.; Chan, S.Y.; Chew, Y.-L.; Hassan, M.; Ahmad Hisham, S.; Abdul Rahman, S.; Mai, C.-W.; Teo, M.Y.; Kee, P.E.; Khoo, K.S.; et al. Recent Developments in Luteolin-Loaded Nanoformulations for Enhanced Anti-Carcinogenic Activities: Insights from In Vitro and In Vivo Studies. Sci 2024, 6, 68. [Google Scholar] [CrossRef]
- Kumar, G.; Jain, P.; Virmani, T.; Sharma, A.; Akhtar, M.S.; Aldosari, S.A.; Khan, M.F.; Duarte, S.O.D.; Fonte, P. Enhancing Therapy with Nano-Based Delivery Systems: Exploring the Bioactive Properties and Effects of Apigenin. Ther. Deliv. 2024, 15, 717–735. [Google Scholar] [CrossRef]
- Shinde, V.R.; Revi, N.; Murugappan, S.; Singh, S.P.; Rengan, A.K. Enhanced Permeability and Retention Effect: A Key Facilitator for Solid Tumor Targeting by Nanoparticles. Photodiagnosis Photodyn. Ther. 2022, 39, 102915. [Google Scholar] [CrossRef]
- Kalyane, D.; Raval, N.; Maheshwari, R.; Tambe, V.; Kalia, K.; Tekade, R.K. Employment of Enhanced Permeability and Retention Effect (EPR): Nanoparticle-Based Precision Tools for Targeting of Therapeutic and Diagnostic Agent in Cancer. Mater. Sci. Eng. C 2019, 98, 1252–1276. [Google Scholar] [CrossRef]
- Lahooti, B.; Akwii, R.G.; Zahra, F.T.; Sajib, M.S.; Lamprou, M.; Alobaida, A.; Lionakis, M.S.; Mattheolabakis, G.; Mikelis, C.M. Targeting Endothelial Permeability in the EPR Effect. J. Control. Release 2023, 361, 212–235. [Google Scholar] [CrossRef]
- Shekhar, S.; Chauhan, M.; Yadav, B.; Dutt, R.; Hu, L.; Muthu, M.S.; Singh, R.P. Enhanced Permeability and Retention Effect-Focused Tumor-Targeted Nanomedicines: Latest Trends, Obstacles and Future Perspective. Nanomedicine 2022, 17, 1213–1216. [Google Scholar] [CrossRef]
- Pandey, P.; Lakhanpal, S.; Mahmood, D.; Kang, H.N.; Kim, B.; Kang, S.; Choi, J.; Moon, S.; Pandey, S.; Ballal, S.; et al. Bergenin, a Bioactive Flavonoid: Advancements in the Prospects of Anticancer Mechanism, Pharmacokinetics and Nanoformulations. Front. Pharmacol. 2025, 15, 1481587. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Aatif, M. Potential Health Benefits of Polyphenols and Their Nanoformulations in Humans BT—Breaking Boundaries: Pioneering Sustainable Solutions Through Materials and Technology; Alam, M.W., Ed.; Springer Nature Singapore: Singapore, 2025; pp. 367–388. ISBN 978-981-97-9827-8. [Google Scholar]
- Silva-Pinto, P.A.; de Pontes, J.T.C.; Aguilar-Morón, B.; Canales, C.S.C.; Pavan, F.R.; Roque-Borda, C.A. Phytochemical Insights into Flavonoids in Cancer: Mechanisms, Therapeutic Potential, and the Case of Quercetin. Heliyon 2025, 11, e42682. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Sowbhagya, R.; Ansari, M.A.; Alzohairy, M.A.; Alomary, M.N.; Almalik, A.I.; Ahmad, W.; Tripathi, T.; Elderdery, A.Y. Polyphenols and Their Nanoformulations: Protective Effects against Human Diseases. Life 2022, 12, 1639. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Khadija, J.F.; Islam, M.R.; Shohag, S.; Mitra, S.; Alghamdi, S.; Babalghith, A.O.; Theyab, A.; Rahman, M.T.; Akter, A.; et al. Investigating Polyphenol Nanoformulations for Therapeutic Targets against Diabetes Mellitus. Evid.-Based Complement. Altern. Med. 2022, 2022, 5649156. [Google Scholar] [CrossRef]
- Ferreira, M.; Costa, D.; Sousa, Â. Flavonoids-Based Delivery Systems towards Cancer Therapies. Bioengineering 2022, 9, 197. [Google Scholar] [CrossRef]
- Pecorini, G.; Ferraro, E.; Puppi, D. Polymeric Systems for the Controlled Release of Flavonoids. Pharmaceutics 2023, 15, 628. [Google Scholar] [CrossRef]
- Chen, L.; Cao, H.; Huang, Q.; Xiao, J.; Teng, H. Absorption, Metabolism and Bioavailability of Flavonoids: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 7730–7742. [Google Scholar] [CrossRef]
- Walle, T. Absorption and Metabolism of Flavonoids. Free Radic. Biol. Med. 2004, 36, 829–837. [Google Scholar] [CrossRef]
- Dobrzynska, M.; Napierala, M.; Florek, E. Flavonoid Nanoparticles: A Promising Approach for Cancer Therapy. Biomolecules 2020, 10, 1268. [Google Scholar] [CrossRef]
- Wahnou, H.; Limami, Y.; Oudghiri, M. Flavonoids and Flavonoid-Based Nanoparticles for Osteoarthritis and Rheumatoid Arthritis Management. BioChem 2024, 4, 38–61. [Google Scholar] [CrossRef]
- Rasel, M.S.I.; Mohona, F.A.; Akter, W.; Kabir, S.; Chowdhury, A.A.; Chowdhury, J.A.; Hassan, M.A.; Al Mamun, A.; Ghose, D.K.; Ahmad, Z.; et al. Exploration of Site-Specific Drug Targeting—A Review on EPR-, Stimuli-, Chemical-, and Receptor-Based Approaches as Potential Drug Targeting Methods in Cancer Treatment. J. Oncol. 2022, 2022, 9396760. [Google Scholar] [CrossRef] [PubMed]
- Soares Mateus, A.R.; Pena, A.; Sendón, R.; Almeida, C.; Nieto, G.A.; Khwaldia, K.; Sanches Silva, A. By-Products of Dates, Cherries, Plums and Artichokes: A Source of Valuable Bioactive Compounds. Trends Food Sci. Technol. 2023, 131, 220–243. [Google Scholar] [CrossRef]
- Anusha Siddiqui, S.; Redha, A.A.; Esmaeili, Y.; Mehdizadeh, M. Novel Insights on Extraction and Encapsulation Techniques of Elderberry Bioactive Compounds. Crit. Rev. Food Sci. Nutr. 2023, 63, 5937–5952. [Google Scholar] [CrossRef]
- Borah, M.S.; Tiwari, A.; Sridhar, K.; Narsaiah, K.; Nayak, P.K.; Stephen Inbaraj, B. Recent Trends in Valorization of Food Industry Waste and By-Products: Encapsulation and In Vitro Release of Bioactive Compounds. Foods 2023, 12, 3823. [Google Scholar] [CrossRef]
- Yetisgin, A.A.; Cetinel, S.; Zuvin, M.; Kosar, A.; Kutlu, O. Therapeutic Nanoparticles and Their Targeted Delivery Applications. Molecules 2020, 25, 2193. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-Based Nanoparticles: An Overview of Biomedical Applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef]
- Zhang, P.; Li, Y.; Tang, W.; Zhao, J.; Jing, L.; McHugh, K.J. Theranostic Nanoparticles with Disease-Specific Administration Strategies. Nano Today 2022, 42, 101335. [Google Scholar] [CrossRef]
- Liu, J.; Cabral, H.; Mi, P. Nanocarriers Address Intracellular Barriers for Efficient Drug Delivery, Overcoming Drug Resistance, Subcellular Targeting and Controlled Release. Adv. Drug Deliv. Rev. 2024, 207, 115239. [Google Scholar] [CrossRef]
- Lammers, T.; Kiessling, F.; Hennink, W.E.; Storm, G. Drug Targeting to Tumors: Principles, Pitfalls and (Pre-) Clinical Progress. J. Control. Release 2012, 161, 175–187. [Google Scholar] [CrossRef]
- Moradi Kashkooli, F.; Soltani, M.; Souri, M. Controlled Anti-Cancer Drug Release through Advanced Nano-Drug Delivery Systems: Static and Dynamic Targeting Strategies. J. Control. Release 2020, 327, 316–349. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Hu, Y.; Li, X. Polymer Mechanochemistry in Drug Delivery: From Controlled Release to Precise Activation. J. Control. Release 2024, 365, 259–273. [Google Scholar] [CrossRef]
- Alavi, S.E.; Alharthi, S.; Alavi, S.Z.; Raza, A.; Ebrahimi Shahmabadi, H. Bioresponsive Drug Delivery Systems. Drug Discov. Today 2024, 29, 103849. [Google Scholar] [CrossRef]
- Alavi, S.E.; Alharthi, S.; Alavi, S.F.; Alavi, S.Z.; Zahra, G.E.; Raza, A.; Ebrahimi Shahmabadi, H. Microfluidics for Personalized Drug Delivery. Drug Discov. Today 2024, 29, 103936. [Google Scholar] [CrossRef]
- Trucillo, P. Biomaterials for Drug Delivery and Human Applications. Materials 2024, 17, 456. [Google Scholar] [CrossRef]
- Lee, H.K.; Park, J.; Kim, B.-R.; Jun, I.; Kim, T.; Namkung, W. Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice. Int. J. Mol. Sci. 2021, 22, 3954. [Google Scholar] [CrossRef]
- Mayr, L.M.; Bojanic, D. Novel Trends in High-Throughput Screening. Curr. Opin. Pharmacol. 2009, 9, 580–588. [Google Scholar] [CrossRef]
- Wildey, M.J.; Haunso, A.; Tudor, M.; Webb, M.; Connick, J.H. Chapter Five—High-Throughput Screening. In Platform Technologies in Drug Discovery and Validation; Goodnow Robert, A., Ed.; Annual Reports in Medicinal Chemistry; Academic Press: London, UK, 2017; Volume 50, pp. 149–195. ISBN 0065-7743. [Google Scholar]
- Gao, M.; Lan, J.; Zhang, Y.; Yu, S.; Bao, B.; Yao, W.; Cao, Y.; Shan, M.; Cheng, F.; Zhang, L.; et al. Discovery of Processing-Associated Q-Marker of Carbonized Traditional Chinese Medicine: An Integrated Strategy of Metabolomics, Systems Pharmacology and in Vivo High-Throughput Screening Model. Phytomedicine 2022, 102, 154152. [Google Scholar] [CrossRef]
- Bajorath, J. Integration of Virtual and High-Throughput Screening. Nat. Rev. Drug Discov. 2002, 1, 882–894. [Google Scholar] [CrossRef]
- Mikhaylova, E.V. Insight into the Flavonoids Enrichment in Plants by Genome Engineering. In Applications of Genome Engineering in Plants; Upadhyay, S.K., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2024; pp. 242–259. ISBN 9781394183913. [Google Scholar]
- Selvakumar, S.C.; Preethi, K.A.; Ross, K.; Tusubira, D.; Khan, M.W.A.; Mani, P.; Rao, T.N.; Sekar, D. CRISPR/Cas9 and next Generation Sequencing in the Personalized Treatment of Cancer. Mol. Cancer 2022, 21, 83. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, D.; Yadav, P.K.; Sultana, N.; Agarwal, A.; Verma, S.; Chourasia, M.K.; Gayen, J.R. Advancements in Nanotechnology for the Delivery of Phytochemicals. J. Integr. Med. 2024, 22, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Zhao, Y.-G.; Zhang, Y. Enhancing Drug Solubility, Bioavailability, and Targeted Therapeutic Applications through Magnetic Nanoparticles. Molecules 2024, 29, 4854. [Google Scholar] [CrossRef]
- Yassin, M.T.; Al-Otibi, F.O.; Al-Sahli, S.A.; El-Wetidy, M.S.; Mohamed, S. Metal Oxide Nanoparticles as Efficient Nanocarriers for Targeted Cancer Therapy: Addressing Chemotherapy-Induced Disabilities. Cancers 2024, 16, 4234. [Google Scholar] [CrossRef]
- Attia, M.F.; Anton, N.; Wallyn, J.; Omran, Z.; Vandamme, T.F. An Overview of Active and Passive Targeting Strategies to Improve the Nanocarriers Efficiency to Tumour Sites. J. Pharm. Pharmacol. 2019, 71, 1185–1198. [Google Scholar] [CrossRef]
- Malone, E.R.; Oliva, M.; Sabatini, P.J.B.; Stockley, T.L.; Siu, L.L. Molecular Profiling for Precision Cancer Therapies. Genome Med. 2020, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Saud, S.M.; Young, M.R.; Jones-Hall, Y.L.; Ileva, L.; Evbuomwan, M.O.; Wise, J.; Colburn, N.H.; Kim, Y.S.; Bobe, G. Chemopreventive Activity of Plant Flavonoid Isorhamnetin in Colorectal Cancer Is Mediated by Oncogenic Src and β-Catenin. Cancer Res. 2013, 73, 5473–5484. [Google Scholar] [CrossRef]

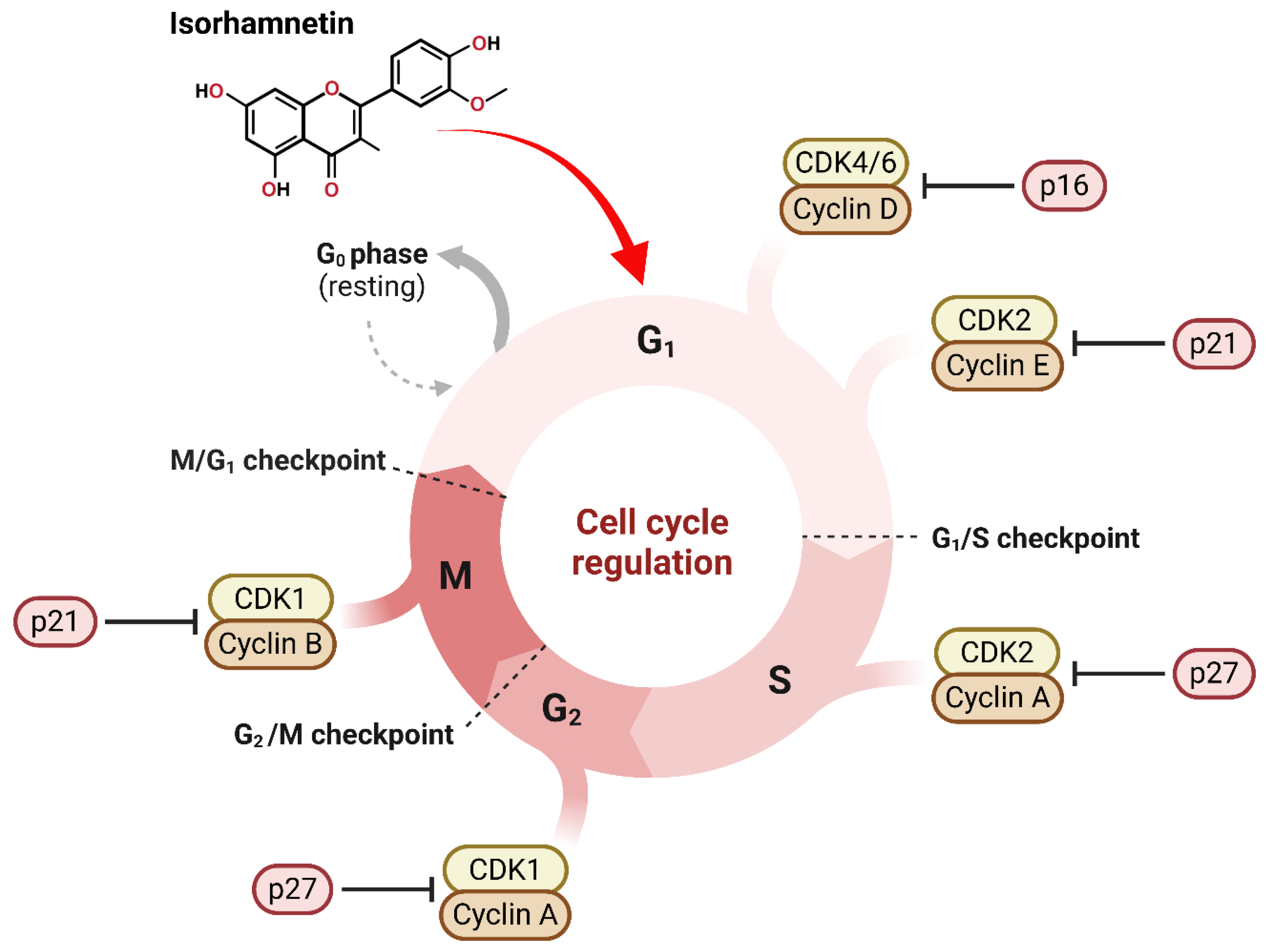


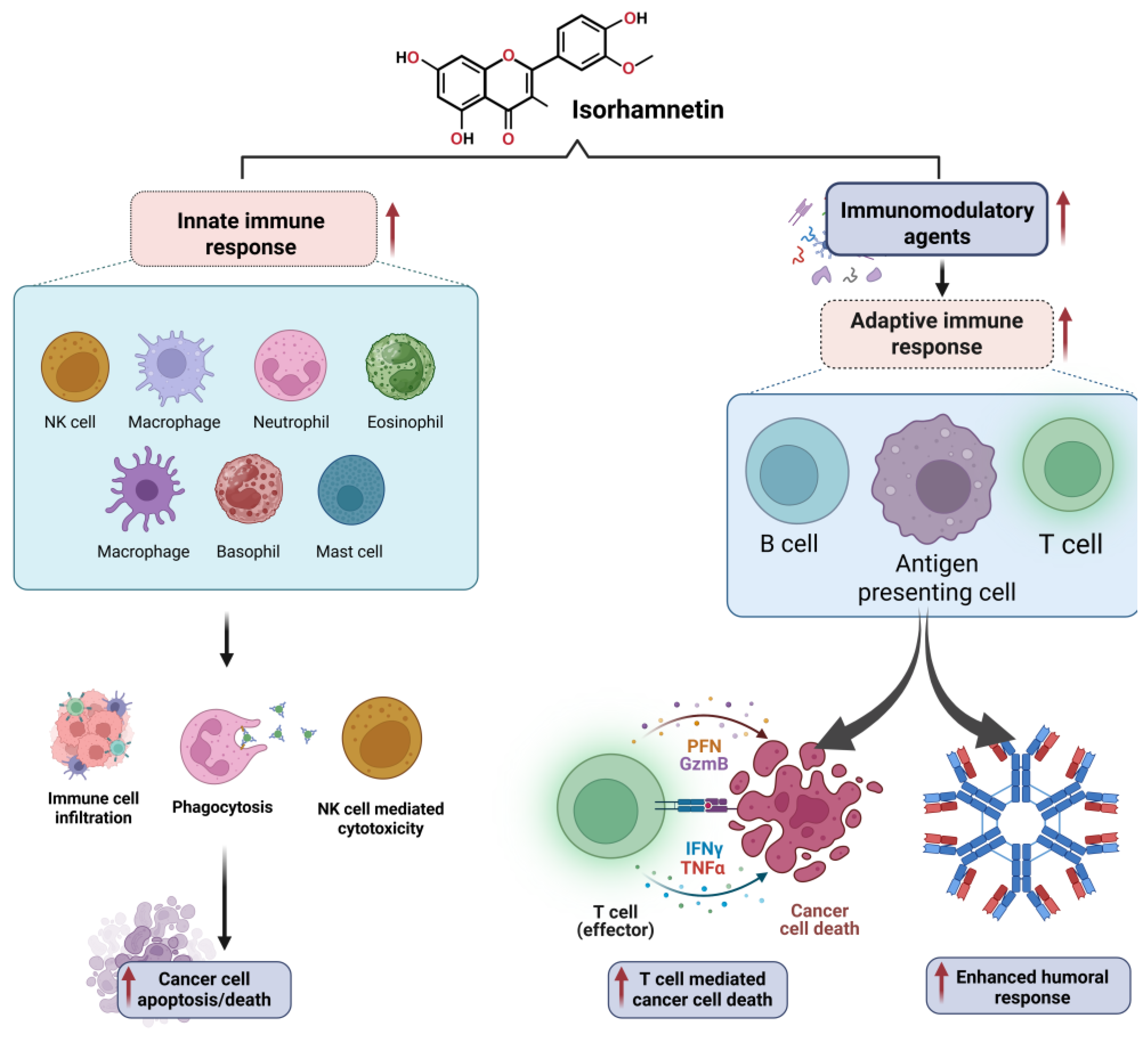
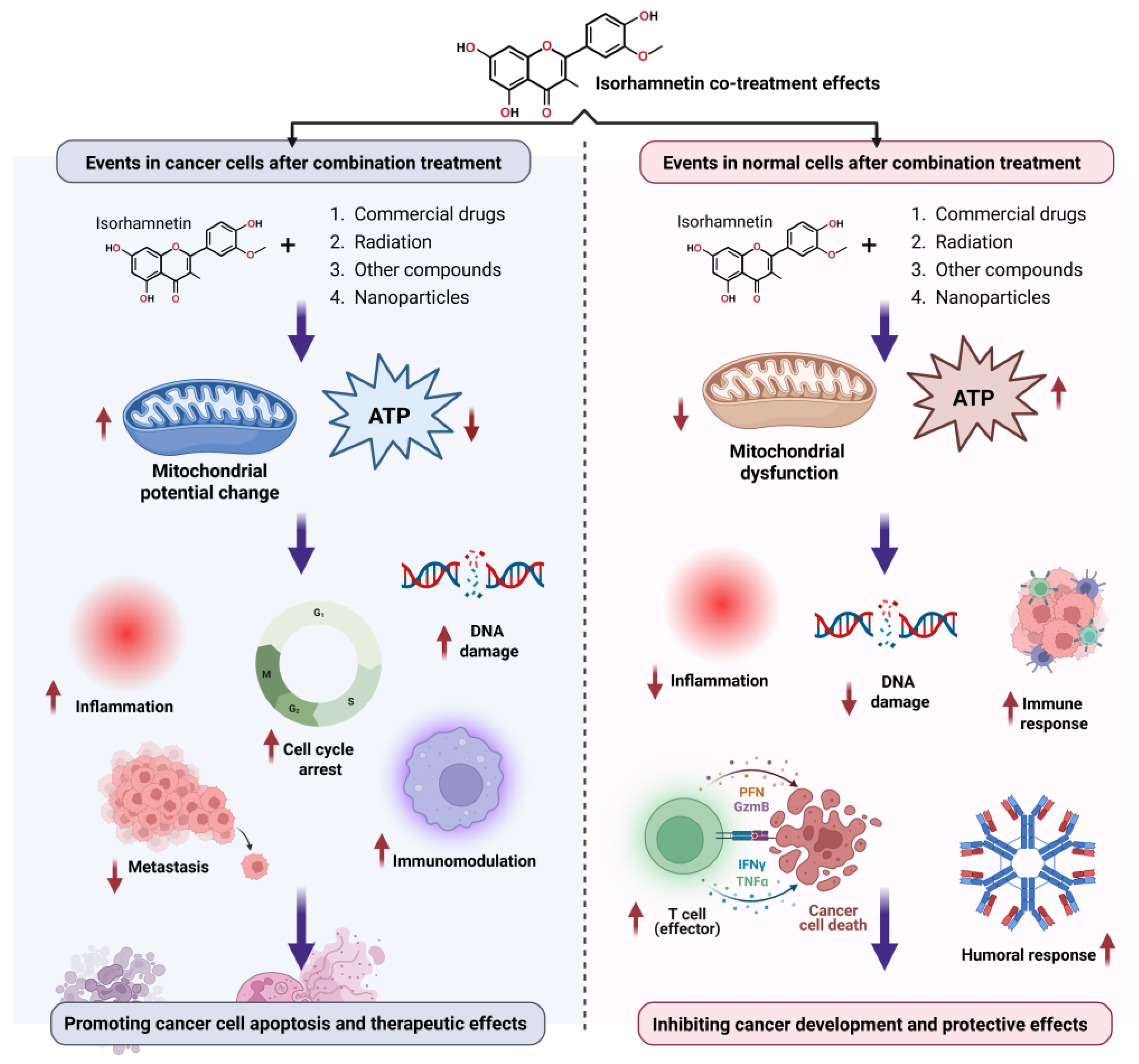
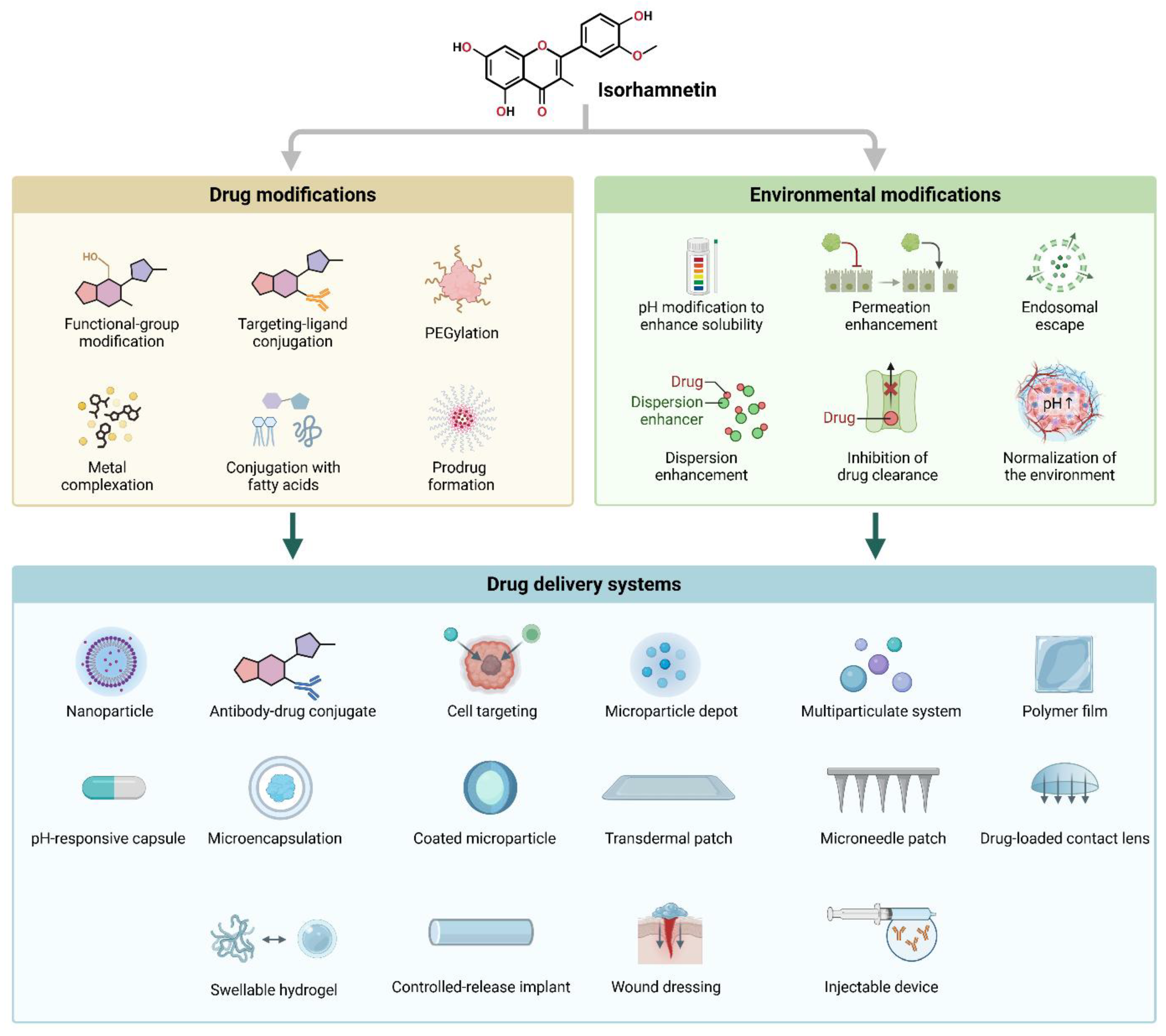
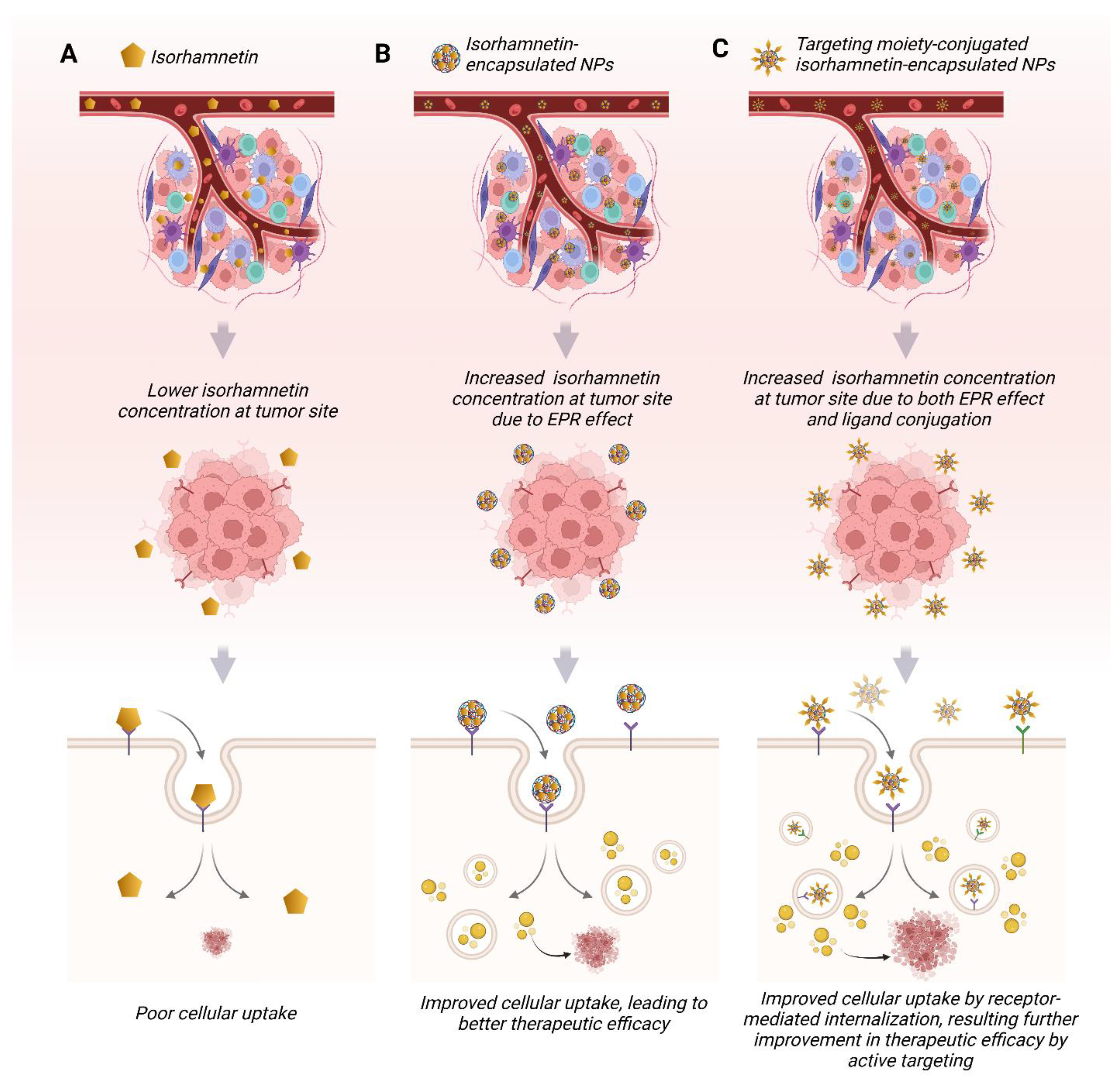

| Delivery Method | Advantages | Challenges |
|---|---|---|
| Nanoparticles | Improved solubility, targeted delivery, enhanced stability | Potential toxicity, complex formulation, high production costs |
| Cell Targeting | Specific cell delivery, minimal side effects, targeted therapy | Requires identification of specific biomarkers, complex design |
| Antibody-Drug Conjugate | Precise targeting, reduced off-target effects, higher therapeutic index | Complex design, risk of immune reactions, expensive |
| Microparticle Depot | Sustained release, controlled release profile, long-term therapy | Slow drug release, potential drug degradation, and formulation challenges |
| Multiparticulate System | Improved bioavailability, reduced side effects, controlled release | Requires precise formulation, potential for uneven drug distribution |
| Polymer Film | Controlled release protects the drug from degradation, versatile | May require large doses, limited release rate, potential skin irritation (for transdermal) |
| pH-Responsive Capsule | Site-specific release protects from stomach acid, enhances absorption | Limited to the gastrointestinal tract, there is potential for incomplete release |
| Microencapsulation | Enhanced stability, controlled release, protects from degradation | Slow release, complex preparation, limited for rapid onset |
| Coated Microparticles | Controlled release protects the drug from degradation and enhances stability. | Complex formulation, potential for incomplete release, high production costs |
| Transdermal Patch | Non-invasive, steady drug release, convenient for chronic conditions | Limited skin permeability, slow absorption, skin irritation |
| Microneedle Patch | Pain-free targeted drug delivery, easy to use | Limited drug load, potential for skin irritation, expensive |
| Drug-Loaded Contact Lens | Direct drug delivery to the eye, localized treatment, non-invasive | Limited to ocular diseases, potential irritation, short duration of effect |
| Controlled Release Implant | Prolonged, consistent release, reduced dosing frequency, localized delivery | Invasive, local tissue irritation, difficult to remove |
| Swellable Hydrogel | Responsive to fluids, sustained release can be used for wound care | Limited to topical applications, swelling issues, possible irritation |
| Wound Dressing | Direct drug application to wounds accelerates healing and protects from infection. | Requires proper formulation for sustained release, may need frequent changes |
| Injectable Device | Rapid onset, precise control of drug dose, targeted delivery | Invasive, potential local irritation, requires medical supervision |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rana, J.N.; Gul, K.; Mumtaz, S. Isorhamnetin: Reviewing Recent Developments in Anticancer Mechanisms and Nanoformulation-Driven Delivery. Int. J. Mol. Sci. 2025, 26, 7381. https://doi.org/10.3390/ijms26157381
Rana JN, Gul K, Mumtaz S. Isorhamnetin: Reviewing Recent Developments in Anticancer Mechanisms and Nanoformulation-Driven Delivery. International Journal of Molecular Sciences. 2025; 26(15):7381. https://doi.org/10.3390/ijms26157381
Chicago/Turabian StyleRana, Juie Nahushkumar, Kainat Gul, and Sohail Mumtaz. 2025. "Isorhamnetin: Reviewing Recent Developments in Anticancer Mechanisms and Nanoformulation-Driven Delivery" International Journal of Molecular Sciences 26, no. 15: 7381. https://doi.org/10.3390/ijms26157381
APA StyleRana, J. N., Gul, K., & Mumtaz, S. (2025). Isorhamnetin: Reviewing Recent Developments in Anticancer Mechanisms and Nanoformulation-Driven Delivery. International Journal of Molecular Sciences, 26(15), 7381. https://doi.org/10.3390/ijms26157381