
Molecular Detection and Genotyping of Enterocytozoon bieneusi in Environmental Sources near Cattle Farms in Korea

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
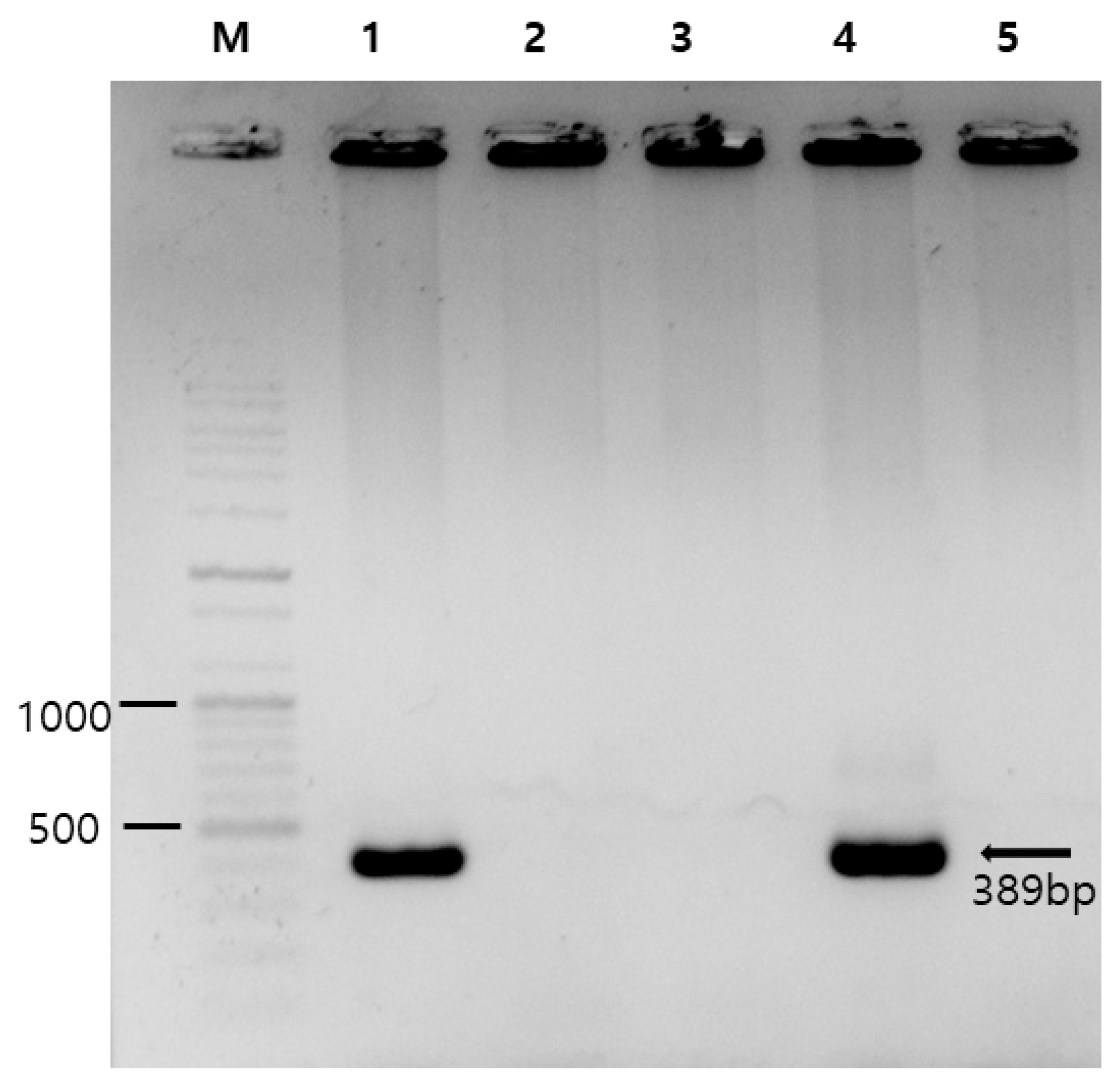
2.1. Polymerase Chain Reaction-Based Prevalence
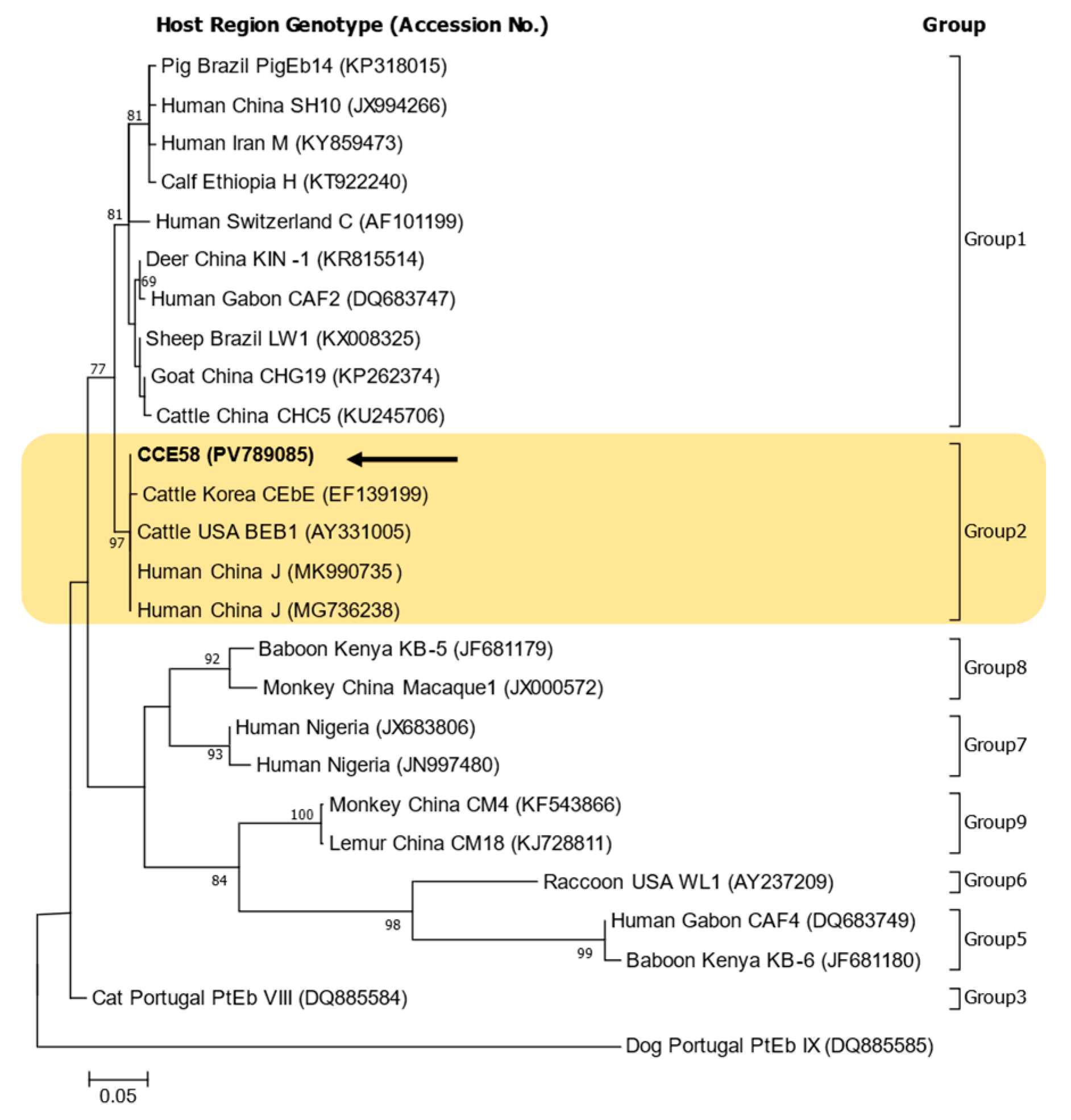
2.2. Sequencing and Phylogenetic Analysis
3. Discussion
4. Materials and Methods
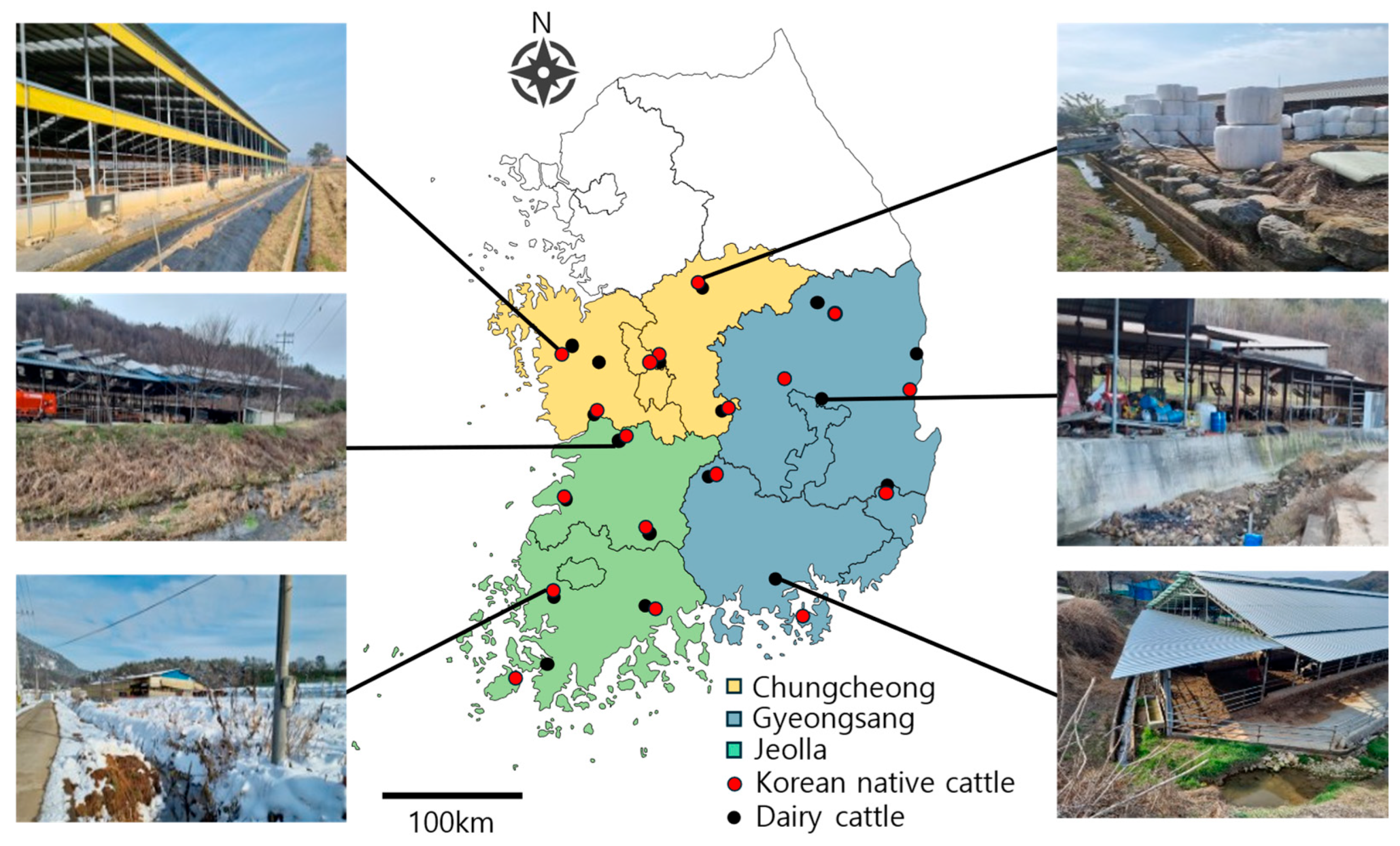
4.1. Sampling Site and Period
4.2. DNA Extraction and PCR
4.3. Phylogenetic Data Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ITS | Internal transcribed spacer |
| PCR | Polymerase chain reaction |
| Q3 | Third Quarter |
| SPSS | Statistical Package for the Social Sciences |
| NCBI | National Center for Biotechnology Information |
References
- Xue, N.Y.; Li, Z.Y.; Wang, H.T.; Qin, Y.; Li, X.M.; Hou, Q.Y.; Jiang, J.; Yang, X.; Ni, H.B. High genetic diversity of Enterocytozoon bieneusi in minks and raccoon dogs in northern China. Parasite 2024, 31, 71. [Google Scholar] [CrossRef]
- Dashti, A.; Santin, M.; Cano, L.; de Lucio, A.; Bailo, B.; de Mingo, M.H.; Koster, P.C.; Fernandez-Basterra, J.A.; Aramburu-Aguirre, J.; Lopez-Molina, N.; et al. Occurrence and genetic diversity of Enterocytozoon bieneusi (Microsporidia) in owned and sheltered dogs and cats in Northern Spain. Parasitol. Res. 2019, 118, 2979–2987. [Google Scholar] [CrossRef]
- Zhang, Y.; Koehler, A.V.; Wang, T.; Haydon, S.R.; Gasser, R.B. First detection and genetic characterisation of Enterocytozoon bieneusi in wild deer in Melbourne’s water catchments in Australia. Parasit. Vectors 2018, 11, 2. [Google Scholar] [CrossRef]
- Zheng, X.L.; Zhou, H.H.; Ren, G.; Ma, T.M.; Cao, Z.X.; Wei, L.M.; Liu, Q.W.; Wang, F.; Zhang, Y.; Liu, H.L.; et al. Genotyping and zoonotic potential of Enterocytozoon bieneusi in cattle farmed in Hainan Province, the southernmost region of China. Parasite 2020, 27, 65. [Google Scholar] [CrossRef]
- Zhao, W.; Ren, G.X.; Qiang, Y.; Li, J.; Pu, J.; Zhang, Y.; Tan, F.; Huang, H.; Liang, S.; Lu, G. Molecular-Based Detection of Enterocytozoon bieneusi in Farmed Masked Palm Civets (Paguma larvata) in Hainan, China: A High-Prevalence, Specificity, and Zoonotic Potential of ITS Genotypes. Front. Vet. Sci. 2021, 8, 714249. [Google Scholar] [CrossRef]
- Zhou, K.; Liu, M.; Wu, Y.; Zhang, R.; Wang, R.; Xu, H.; Wang, Y.; Yao, L.; Yu, H.; Liu, A. Enterocytozoon bieneusi in patients with diarrhea and in animals in the northeastern Chinese city of Yichun: Genotyping and assessment of potential zoonotic transmission. Parasite 2022, 29, 40. [Google Scholar] [CrossRef]
- Feng, K.; Yang, S.; Xu, Y.; Wen, L.; Chen, J.; Zhang, W.; Chen, S.; Shen, Y.; Xiao, L.; Guo, Y.; et al. Molecular characterization of Cryptosporidium spp., Giardia spp. and Enterocytozoon bieneusi in eleven wild rodent species in China: Common distribution, extensive genetic diversity and high zoonotic potential. One Health 2024, 18, 100750. [Google Scholar] [CrossRef]
- Jiang, S.; Yu, S.; Feng, Y.; Zhang, L.; Santin, M.; Xiao, L.; Li, W. Widespread distribution of human-infective Enterocytozoon bieneusi genotypes in small rodents in northeast China and phylogeny and zoonotic implications revisited. Acta Trop. 2024, 253, 107160. [Google Scholar] [CrossRef]
- Song, H.Y.; Wang, K.S.; Yang, J.F.; Mao, H.M.; Pu, L.H.; Zou, Y.; Ma, J.; Zhu, X.Q.; Zou, F.C.; He, J.J. Prevalence and Novel Genotypes Identification of Enterocytozoon bieneusi in Dairy Cattle in Yunnan Province, China. Animals 2021, 11, 3014. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.H.; Lee, Y.R.; Kim, H.Y.; Moon, B.Y.; Han, J.E.; Rhee, M.H.; Kwon, O.D.; Kwak, D. Enterocytozoon bieneusi Genotypes and Infections in the Horses in Korea. Korean J. Parasitol. 2021, 59, 639–643. [Google Scholar] [CrossRef]
- Hwang, S.; Shin, S.U.; Kim, S.; Ryu, J.H.; Choi, K.S. Zoonotic potential of Enterocytozoon bieneusi in pre-weaned Korean native calves. Parasit. Vectors 2020, 13, 300. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Qin, R.L.; Mei, J.J.; Zou, Y.; Zhang, Z.H.; Zheng, W.B.; Liu, Q.; Zhu, X.Q.; Gao, W.W.; Xie, S.C. Molecular Detection and Genotyping of Enterocytozoon bieneusi in Beef Cattle in Shanxi Province, North China. Animals 2022, 12, 2961. [Google Scholar] [CrossRef]
- Fan, W.J.; Zhang, S.; Wang, L.F.; Ding, Y.L.; Liu, H.X.; Wang, M.Y.; Wang, Y.; Chai, H.L.; Zhang, Z.S.; Yi, C.; et al. Prevalence and genotyping of Enterocytozoon bieneusi in cattle from Shanxi and Inner Mongolia, China. Sci. Rep. 2025, 15, 6818. [Google Scholar] [CrossRef]
- Zhou, H.H.; Zheng, X.L.; Ma, T.M.; Qi, M.; Zhou, J.G.; Liu, H.J.; Lu, G.; Zhao, W. Molecular detection of Enterocytozoon bieneusi in farm-raised pigs in Hainan Province, China: Infection rates, genotype distributions, and zoonotic potential. Parasite 2020, 27, 12. [Google Scholar] [CrossRef]
- Lee, J.H. Prevalence and molecular characteristics of Enterocytozoon bieneusi in cattle in Korea. Parasitol. Res. 2007, 101, 391–396. [Google Scholar] [CrossRef]
- Lee, S.H.; Oem, J.K.; Lee, S.M.; Son, K.; Jo, S.D.; Kwak, D. Molecular detection of Enterocytozoon bieneusi from bats in South Korea. Med. Mycol. 2018, 56, 1033–1037. [Google Scholar] [CrossRef]
- Amer, S.; Kim, S.; Han, J.I.; Na, K.J. Prevalence and genotypes of Enterocytozoon bieneusi in wildlife in Korea: A public health concern. Parasit. Vectors 2019, 12, 160. [Google Scholar] [CrossRef]
- Lee, H.; Seo, M.G.; Lee, S.H.; Oem, J.K.; Kim, S.H.; Jeong, H.; Kim, Y.; Jheong, W.H.; Kwon, O.D.; Kwak, D. Distribution and genotypic analysis of Enterocytozoon bieneusi from wild boars in Korea. Med. Mycol. 2021, 59, 934–938. [Google Scholar] [CrossRef]
- Noh, G.; Lee, H.; Lee, S.H.; Seo, M.G.; Kim, K.T.; Lee, J.; Nazim, K.; Park, S.J.; Rhee, M.H.; Kwak, D. Genotypic analysis of zoonotic Enterocytozoon bieneusi in wild deer in Korea. Parasites Hosts Dis. 2024, 62, 484–489. [Google Scholar] [CrossRef]
- Park, H.M.; Lee, H.; Sung, S.Y.; Nazim, K.; Jang, B.Y.; Sung, K.C.; Lee, S.H.; Seo, M.G.; Rhee, M.H.; Kwak, D. Distribution and genotypic analysis of Enterocytozoon bieneusi from cats in Korea. Parasites Hosts Dis. 2025, 63, 188–194. [Google Scholar] [CrossRef]
- Jiang, Y.; Yuan, Z.; Wang, Y.; Zhang, J.; Shen, Y.; Cao, J. Wastewater-based intestinal protozoa monitoring in Shanghai, China. Microbiol. Spectr. 2024, 12, e0403223. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, X.; Yang, R.; Zhao, W.; Li, N.; Guo, Y.; Xiao, L.; Feng, Y. Molecular characterization of the waterborne pathogens Cryptosporidium spp., Giardia duodenalis, Enterocytozoon bieneusi, Cyclospora cayetanensis and Eimeria spp. in wastewater and sewage in Guangzhou, China. Parasit. Vectors 2021, 14, 66. [Google Scholar] [CrossRef]
- Guo, Y.; Alderisio, K.A.; Yang, W.; Cama, V.; Feng, Y.; Xiao, L. Host specificity and source of Enterocytozoon bieneusi genotypes in a drinking source watershed. Appl. Environ. Microbiol. 2014, 80, 218–225. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, Y.; Huang, C.; Xiao, L. Occurrence, source, and human infection potential of Cryptosporidium and Enterocytozoon bieneusi in drinking source water in Shanghai, China, during a pig carcass disposal incident. Environ. Sci. Technol. 2014, 48, 14219–14227. [Google Scholar] [CrossRef]
- Rural Development Administration National Institute of Animal Science. Livestock Statistics 30. Available online: https://www.nias.go.kr/front/soboarddown.do?cmCode=M090814150936066&boardSeqNum=334&fileSeqNum=349 (accessed on 1 June 2025).
- Statistics Korea. Statistical Database. Available online: https://kosis.kr/eng/ (accessed on 1 June 2025).
- Petersen, H.H.; Jianmin, W.; Katakam, K.K.; Mejer, H.; Thamsborg, S.M.; Dalsgaard, A.; Olsen, A.; Enemark, H.L. Cryptosporidium and Giardia in Danish organic pig farms: Seasonal and age-related variation in prevalence, infection intensity and species/genotypes. Vet. Parasitol. 2015, 214, 29–39. [Google Scholar] [CrossRef]
- Zahedi, A.; Ryan, U.; Rawlings, V.; Greay, T.; Hancock, S.; Bruce, M.; Jacobson, C. Cryptosporidium and Giardia in dam water on sheep farms—An important source of transmission? Vet. Parasitol. 2020, 288, 109281. [Google Scholar] [CrossRef]
- Miambo, R.D.; Laitela, B.; Malatji, M.P.; De Santana Afonso, S.M.; Junior, A.P.; Lindh, J.; Mukaratirwa, S. Prevalence of Giardia and Cryptosporidium in young livestock and dogs in Magude District of Maputo Province, Mozambique. Onderstepoort J. Vet. Res. 2019, 86, e1–e6. [Google Scholar] [CrossRef]
- Yamashiro, S.; Fiuza, V.; Teixeira, A.; Branco, N.; Levy, C.E.; Castro, I.; Franco, R.M.B. Enterocytozoon bieneusi detected by molecular methods in raw sewage and treated effluent from a combined system in Brazil. Mem. Inst. Oswaldo Cruz 2017, 112, 403–410. [Google Scholar] [CrossRef]
- Tekeli, N.T.F.; Al-Jarjary, S.A.A.; Sheet, O.H. Molecular detection and phylogenetic analysis of microsporidia in water and soil in Mosul city. Open Vet. J. 2024, 14, 2421–2432. [Google Scholar] [CrossRef]
- Wnuk, E.; Wasko, A.; Walkiewicz, A.; Bartminski, P.; Bejger, R.; Mielnik, L.; Bieganowski, A. The effects of humic substances on DNA isolation from soils. PeerJ 2020, 8, e9378. [Google Scholar] [CrossRef]
- Yu, J.R.; Lee, J.K.; Seo, M.; Kim, S.I.; Sohn, W.M.; Huh, S.; Choi, H.Y.; Kim, T.S. Prevalence of cryptosporidiosis among the villagers and domestic animals in several rural areas of Korea. Korean J. Parasitol. 2004, 42, 1–6. [Google Scholar] [CrossRef]
- Park, J.H.; Guk, S.M.; Han, E.T.; Shin, E.H.; Kim, J.L.; Chai, J.Y. Genotype analysis of Cryptosporidium spp. prevalent in a rural village in Hwasun-gun, Republic of Korea. Korean J. Parasitol. 2006, 44, 27–33. [Google Scholar] [CrossRef]
- Chai, J.Y.; Lee, S.H.; Guk, S.M.; Lee, S.H. An epidemiological survey of Cryptosporidium parvum infection in randomly selected inhabitants of Seoul and Chollanam-do. Korean J. Parasitol. 1996, 34, 113–119. [Google Scholar] [CrossRef]
- Alegbeleye, O.O.; Sant’Ana, A.S. Manure-borne pathogens as an important source of water contamination: An update on the dynamics of pathogen survival/transport as well as practical risk mitigation strategies. Int. J. Hyg. Environ. Health 2020, 227, 113524. [Google Scholar] [CrossRef]
- Abdugheni, R.; Li, L.; Yang, Z.N.; Huang, Y.; Fang, B.Z.; Shurigin, V.; Mohamad, O.A.A.; Liu, Y.H.; Li, W.J. Microbial Risks Caused by Livestock Excrement: Current Research Status and Prospects. Microorganisms 2023, 11, 1897. [Google Scholar] [CrossRef]
- Li, W.; Xiao, L. Ecological and public health significance of Enterocytozoon bieneusi. One Health 2021, 12, 100209. [Google Scholar] [CrossRef]
- Ye, J.; Ji, Y.; Xu, J.; Ma, K.; Yang, X. Zoonotic Enterocytozoon bieneusi in raw wastewater in Zhengzhou, China. Folia Parasitol. 2017, 64, 1. [Google Scholar] [CrossRef]
- Zhang, Y.; Koehler, A.V.; Wang, T.; Haydon, S.R.; Gasser, R.B. Enterocytozoon bieneusi Genotypes in Cattle on Farms Located within a Water Catchment Area. J. Eukaryot. Microbiol. 2019, 66, 553–559. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, M.; Jing, B.; Yu, F.; Wu, Y.; Chang, Y.; Zhao, A.; Wei, Z.; Dong, H.; Zhang, L. Molecular Characterization of Cryptosporidium spp., Giardia duodenalis, and Enterocytozoon bieneusi in Rabbits in Xinjiang, China. J. Eukaryot. Microbiol. 2018, 65, 854–859. [Google Scholar] [CrossRef]
- Santin, M.; Fayer, R. Enterocytozoon bieneusi genotype nomenclature based on the internal transcribed spacer sequence: A consensus. J. Eukaryot. Microbiol. 2009, 56, 34–38. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, L.; Duan, L.; Ye, J.; Guo, Y.; Guo, M.; Liu, L.; Feng, Y. Concurrent infections of Giardia duodenalis, Enterocytozoon bieneusi, and Clostridium difficile in children during a cryptosporidiosis outbreak in a pediatric hospital in China. PLoS Negl. Trop. Dis. 2013, 7, e2437. [Google Scholar] [CrossRef]
- Thrusfield, M.V. Veterinary Epidemiology, 3rd ed.; Blackwell Science: Ames, IA, USA, 2005. [Google Scholar]
- Vallejo, A.; Muniesa, A.; Ferreira, C.; de Blas, I. New method to estimate the sample size for calculation of a proportion assuming binomial distribution. Res. Vet. Sci. 2013, 95, 405–409. [Google Scholar] [CrossRef]
- Lwanga, S.K.; Lemeshow, S. Sample Size Determination in Health Studies: A Practical Manual; World Health Organization: Geneva, Switzerland, 1991; p. 80. [Google Scholar]
- Karim, M.R.; Dong, H.; Li, T.; Yu, F.; Li, D.; Zhang, L.; Li, J.; Wang, R.; Li, S.; Li, X.; et al. Predomination and new genotypes of Enterocytozoon bieneusi in captive nonhuman primates in zoos in China: High genetic diversity and zoonotic significance. PLoS ONE 2015, 10, e0117991. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Variable | No. Tested | No. Positive (%) | p-Value | |
|---|---|---|---|---|
| Region | Chungcheong | 124 | 1 (0.8) | 0.643 |
| Gyeongsang | 130 | 0 | ||
| Jeolla | 110 | 0 | ||
| Season | Spring | 76 | 0 | 1.000 |
| Summer | 88 | 0 | ||
| Autumn | 120 | 1 (0.8) | ||
| Winter | 80 | 0 | ||
| Farm type | Dairy cattle | 177 | 0 | 1.000 |
| Korean native cattle | 187 | 1 (0.5) | ||
| Source | Soil | 185 | 0 | 0.492 |
| Water | 179 | 1 (0.6) | ||
| Total | 364 | 1 (0.3) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Jo, M.; Kim, H.; Nazim, K.; Lee, S.-H.; Seo, M.-G.; Park, S.-J.; Rhee, M.H.; Kwak, D. Molecular Detection and Genotyping of Enterocytozoon bieneusi in Environmental Sources near Cattle Farms in Korea. Int. J. Mol. Sci. 2025, 26, 7270. https://doi.org/10.3390/ijms26157270
Lee H, Jo M, Kim H, Nazim K, Lee S-H, Seo M-G, Park S-J, Rhee MH, Kwak D. Molecular Detection and Genotyping of Enterocytozoon bieneusi in Environmental Sources near Cattle Farms in Korea. International Journal of Molecular Sciences. 2025; 26(15):7270. https://doi.org/10.3390/ijms26157270
Chicago/Turabian StyleLee, Haeseung, Myungji Jo, Hyeyeon Kim, Kaifa Nazim, Seung-Hun Lee, Min-Goo Seo, Sang-Joon Park, Man Hee Rhee, and Dongmi Kwak. 2025. "Molecular Detection and Genotyping of Enterocytozoon bieneusi in Environmental Sources near Cattle Farms in Korea" International Journal of Molecular Sciences 26, no. 15: 7270. https://doi.org/10.3390/ijms26157270
APA StyleLee, H., Jo, M., Kim, H., Nazim, K., Lee, S.-H., Seo, M.-G., Park, S.-J., Rhee, M. H., & Kwak, D. (2025). Molecular Detection and Genotyping of Enterocytozoon bieneusi in Environmental Sources near Cattle Farms in Korea. International Journal of Molecular Sciences, 26(15), 7270. https://doi.org/10.3390/ijms26157270