
Large-Scale Profiling of Coding and Long Noncoding Transcriptomes in the Hippocampus of Mice Acutely Exposed to Vaporized CBD or THC

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Histological Assessment of Hippocampal Changes Following Single Exposure to CBD or THC
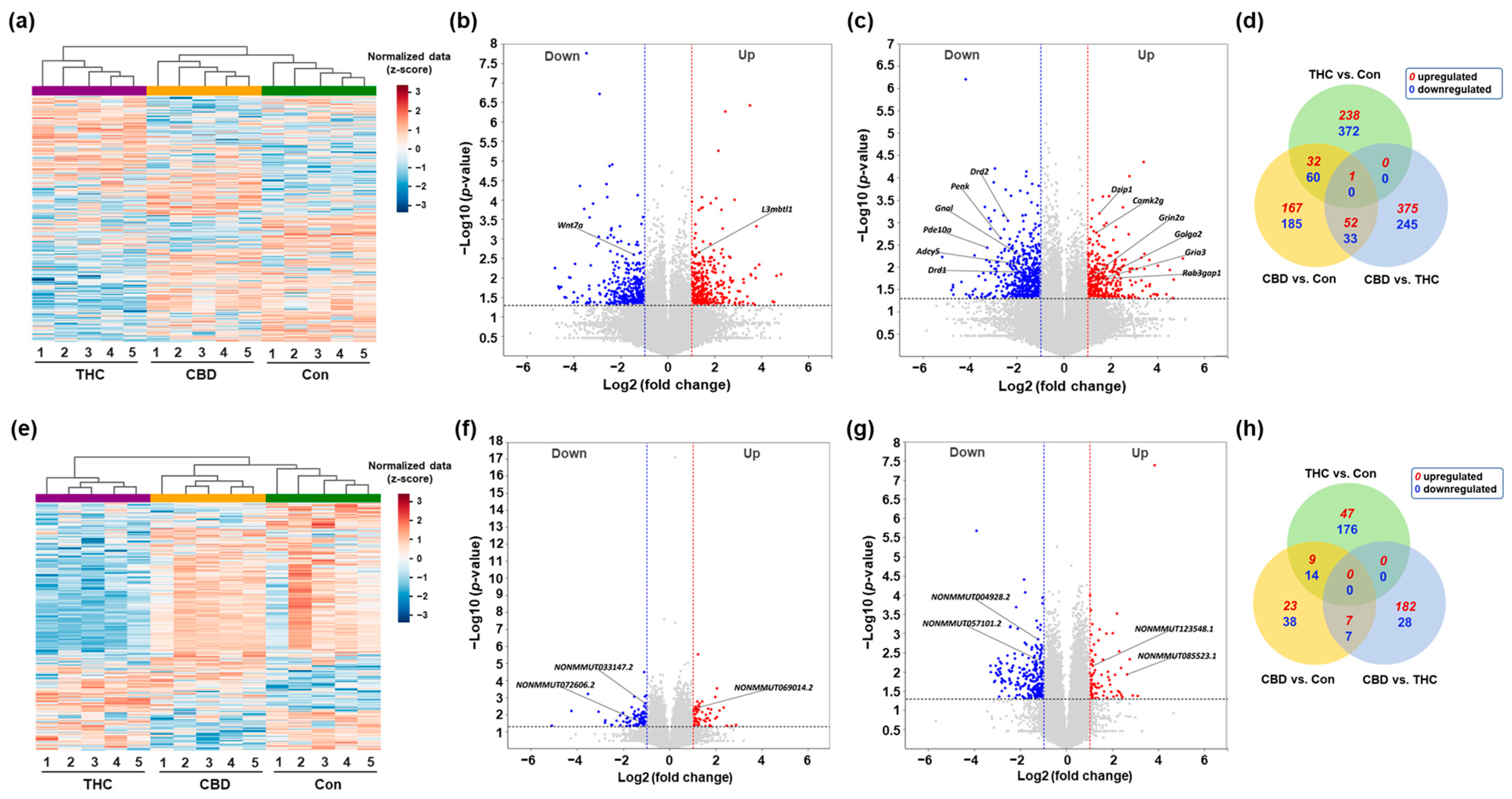
2.2. Differential Expression Profiling of mRNAs and lncRNAs in the Hippocampus After CBD or THC Exposure
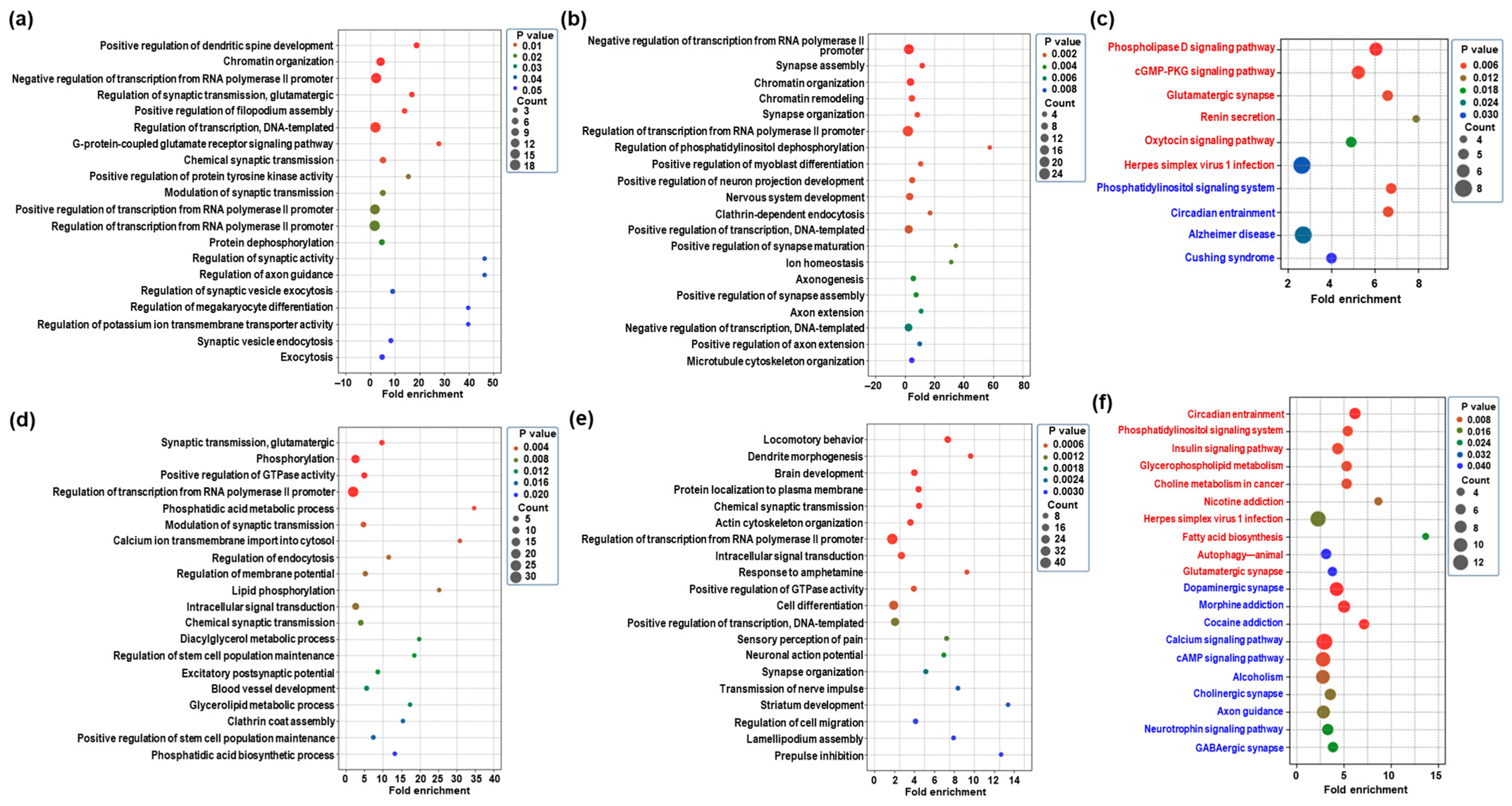
2.3. GO Enrichment and KEGG Pathway of Differentially Expressed mRNAs in the Hippocampus Following CBD or THC Exposure
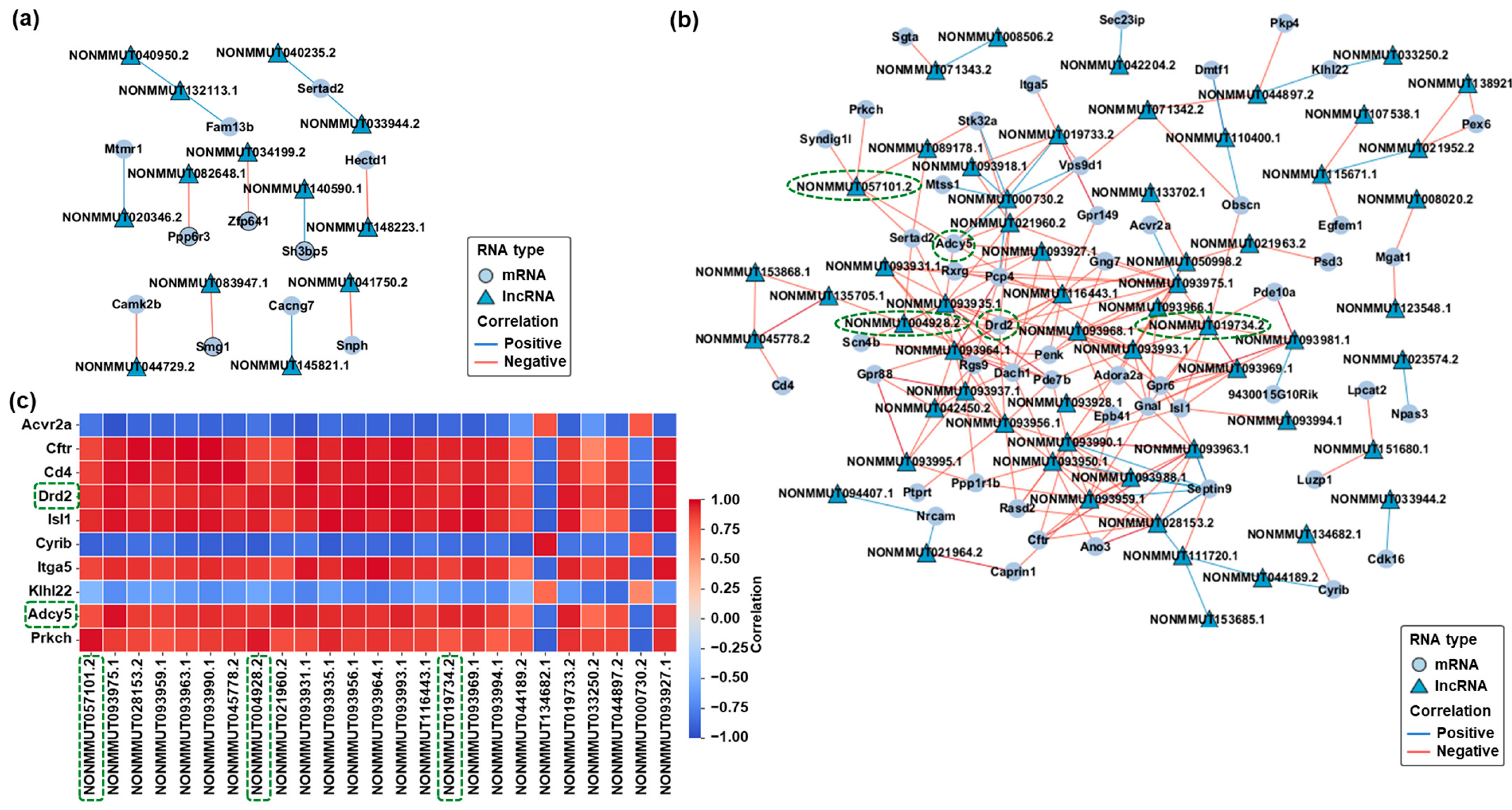
2.4. Prediction and Visualization of mRNA-lncRNA Interaction Networks in Response to CBD and THC Exposure
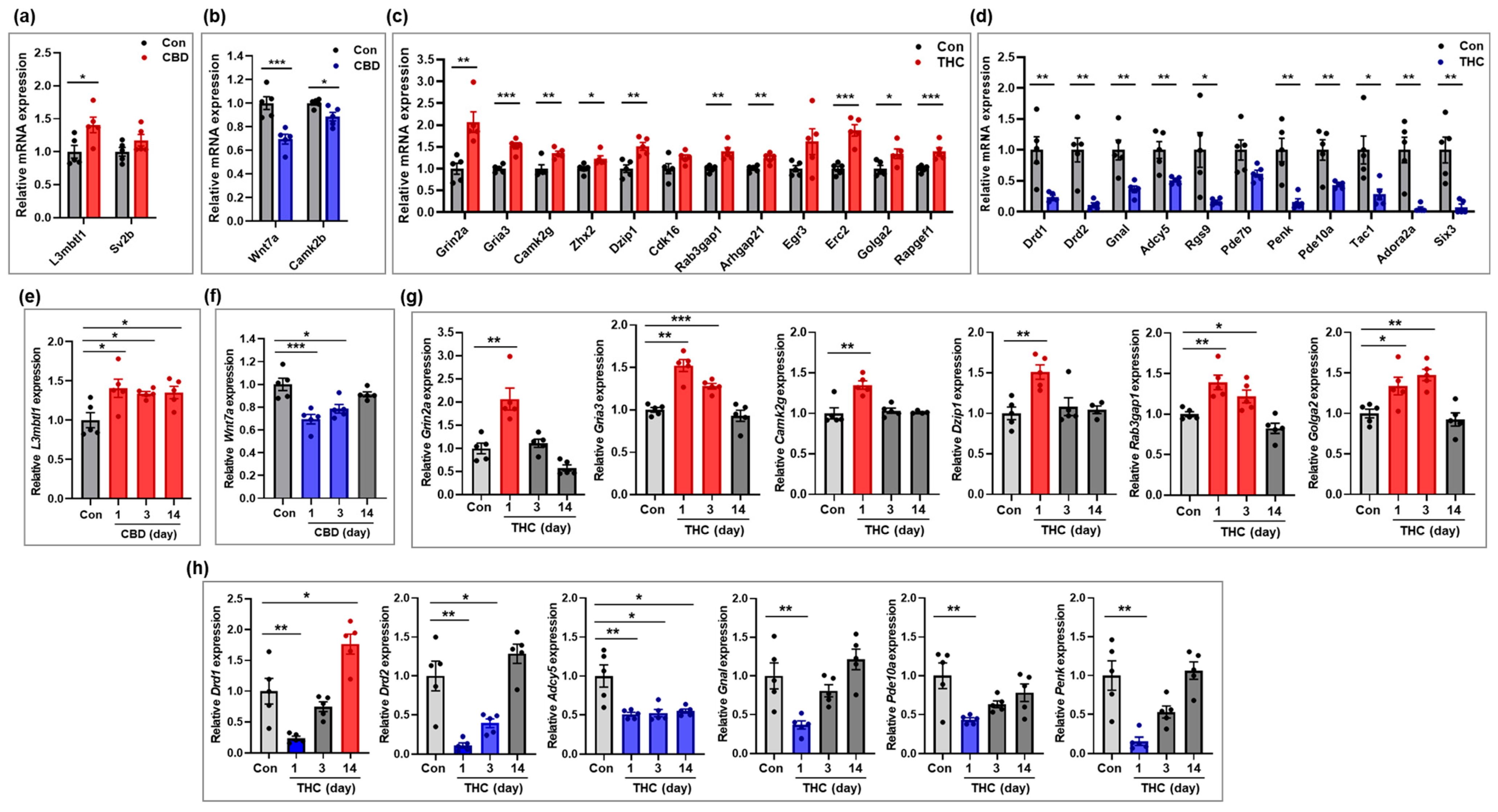
2.5. Validation of Selected mRNAs Based on qRT-PCR
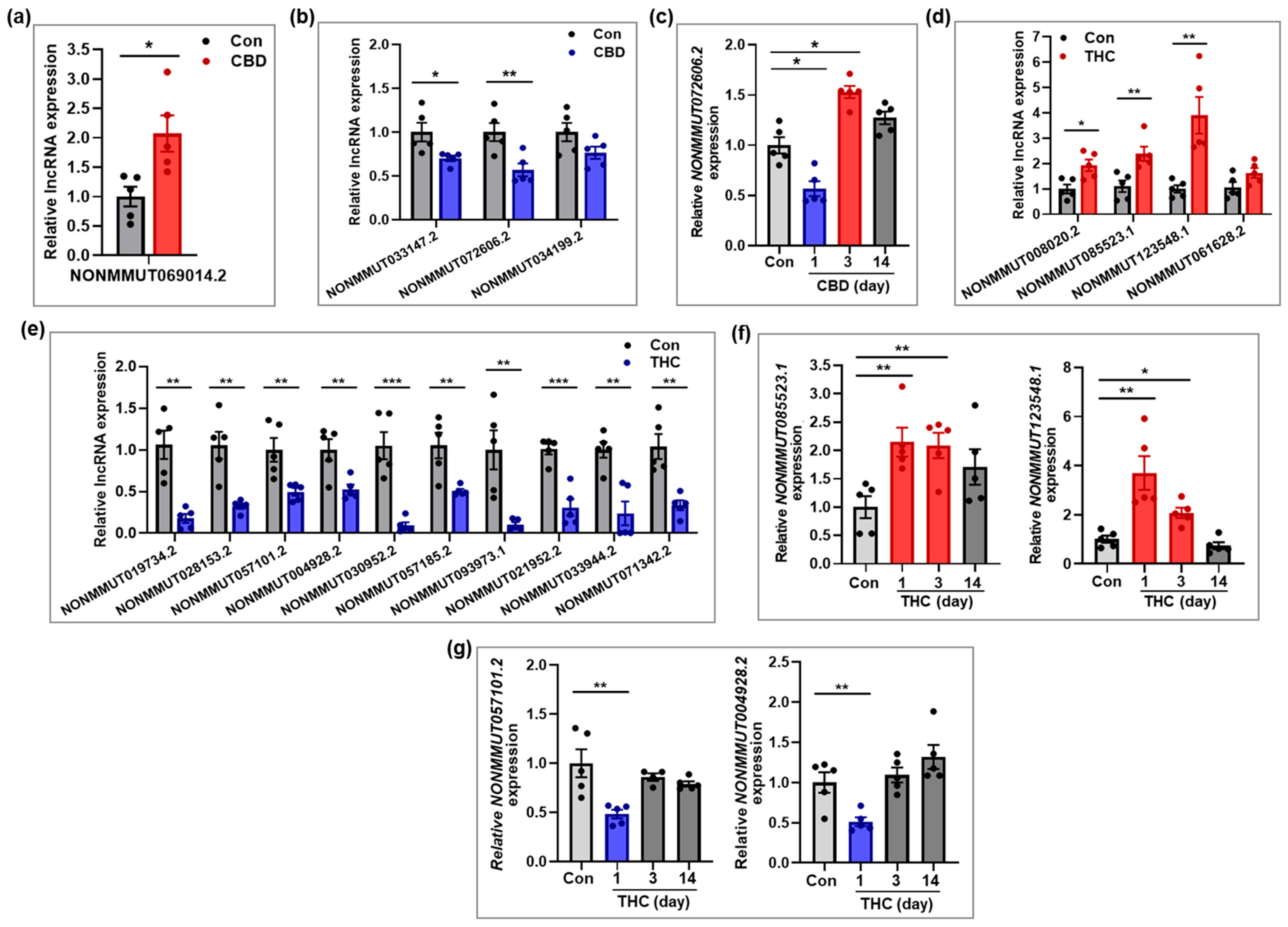
2.6. Validation of Selected lncRNAs Based on qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Vaping Exposure
4.3. Histological Analysis
4.4. Library Preparation and Sequencing
4.5. Differential Gene Expression Analysis and Functional Annotation
4.6. qRT-PCR
4.7. mRNA-lncRNA Co-Expression Network
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CBD | Cannabidiol |
| THC | Delta-9-tetrahydrocannabinol |
| lncRNAs | Long noncoding RNAs |
| EVALI | E-cigarette, or vaping, product use associated lung injury |
| qRT-PCR | Quantitative RT-PCR |
| H&E | Hematoxylin and eosin |
| ExDEGA | Excel-based differentially expressed gene analysis |
| GO | Gene ontology |
| KEGG | Kyto Encyclopedia of Genes and Genomes |
| BP | Biological process |
| PRC1 | Polycomb repressive complex 1 |
| PG | Propylene glycol |
| VG | Vegetable glycerin |
| rRNA | Ribosomal RNA |
| K-BDS | Korea BioData Station |
| FPKM | Fragments per kilobase of transcript per million mapped reads |
| CC | Cellular component |
| MF | Molecular function |
References
- Volkow, N.D.; Baler, R.D.; Compton, W.M.; Weiss, S.R. Adverse health effects of marijuana use. N. Engl. J. Med. 2014, 370, 2219–2227. [Google Scholar] [CrossRef]
- Chen, G.; Rahman, S.; Lutfy, K. E-cigarettes may serve as a gateway to conventional cigarettes and other addictive drugs. Adv. Drug Alcohol Res. 2023, 3, 11345. [Google Scholar] [CrossRef] [PubMed]
- Lubman, D.I.; Cheetham, A.; Yücel, M. Cannabis and adolescent brain development. Pharmacol. Ther. 2015, 148, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mersiades, A.J.; Tognela, A.; Haber, P.S.; Stockler, M.; Lintzeris, N.; Simes, J.; McGregor, I.; Olver, I.; Allsop, D.J.; Gedye, C.; et al. Oral cannabinoid-rich THC/CBD cannabis extract for secondary prevention of chemotherapy-induced nausea and vomiting: A study protocol for a pilot and definitive randomised double-blind placebo-controlled trial (CannabisCINV). BMJ Open 2018, 8, e020745. [Google Scholar] [CrossRef] [PubMed]
- Rajan, T.S.; Giacoppo, S.; Iori, R.; De Nicola, G.R.; Grassi, G.; Pollastro, F.; Bramanti, P.; Mazzon, E. Anti-inflammatory and antioxidant effects of a combination of cannabidiol and moringin in LPS-stimulated macrophages. Fitoterapia 2016, 112, 104–115. [Google Scholar] [CrossRef]
- Capucciati, A.; Bini, A.; Mannucci, B.; Porta, A.; Profumo, A.; Merli, D. CBD-Containing Liquids for e-Cigarettes: Formation of Psychotropic and Secondary Cannabinoids and Amount of CBD Surviving the Smoking Procedure. Forensic Sci. 2023, 3, 258–272. [Google Scholar] [CrossRef]
- Czégény, Z.; Nagy, G.; Babinszki, B.; Bajtel, Á.; Sebestyén, Z.; Kiss, T.; Csupor-Löffler, B.; Tóth, B.; Csupor, D. CBD, a precursor of THC in e-cigarettes. Sci. Rep. 2021, 11, 8951. [Google Scholar] [CrossRef]
- Bloomfield, M.A.P.; Green, S.F.; Hindocha, C.; Yamamori, Y.; Yim, J.L.L.; Jones, A.P.M.; Walker, H.R.; Tokarczuk, P.; Statton, B.; Howes, O.D.; et al. The effects of acute cannabidiol on cerebral blood flow and its relationship to memory: An arterial spin labelling magnetic resonance imaging study. J. Psychopharmacol. 2020, 34, 981–989. [Google Scholar] [CrossRef]
- Atwood, B.K.; Mackie, K. CB2: A cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 2010, 160, 467–479. [Google Scholar] [CrossRef]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Campos, A.C.; Fogaça, M.V.; Sonego, A.B.; Guimarães, F.S. Cannabidiol, neuroprotection and neuropsychiatric disorders. Pharmacol. Res. 2016, 112, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Scuderi, C.; Valenza, M.; Togna, G.I.; Latina, V.; De Filippis, D.; Cipriano, M.; Carratù, M.R.; Iuvone, T.; Steardo, L. Cannabidiol reduces Aβ-induced neuroinflammation and promotes hippocampal neurogenesis through PPARγ involvement. PLoS ONE 2011, 6, e28668. [Google Scholar] [CrossRef] [PubMed]
- Melas, P.A.; Scherma, M.; Fratta, W.; Cifani, C.; Fadda, P. Cannabidiol as a Potential Treatment for Anxiety and Mood Disorders: Molecular Targets and Epigenetic Insights from Preclinical Research. Int. J. Mol. Sci. 2021, 22, 1863. [Google Scholar] [CrossRef]
- Campos, A.C.; Ortega, Z.; Palazuelos, J.; Fogaça, M.V.; Aguiar, D.C.; Díaz-Alonso, J.; Ortega-Gutiérrez, S.; Vázquez-Villa, H.; Moreira, F.A.; Guzmán, M.; et al. The anxiolytic effect of cannabidiol on chronically stressed mice depends on hippocampal neurogenesis: Involvement of the endocannabinoid system. Int. J. Neuropsychopharmacol. 2013, 16, 1407–1419. [Google Scholar] [CrossRef]
- Wolf, S.A.; Bick-Sander, A.; Fabel, K.; Leal-Galicia, P.; Tauber, S.; Ramirez-Rodriguez, G.; Müller, A.; Melnik, A.; Waltinger, T.P.; Ullrich, O.; et al. Cannabinoid receptor CB1 mediates baseline and activity-induced survival of new neurons in adult hippocampal neurogenesis. Cell Commun. Signal. 2010, 8, 12. [Google Scholar] [CrossRef]
- Busquets-Garcia, A.; Gomis-González, M.; Salgado-Mendialdúa, V.; Galera-López, L.; Puighermanal, E.; Martín-García, E.; Maldonado, R.; Ozaita, A. Hippocampal Protein Kinase C Signaling Mediates the Short-Term Memory Impairment Induced by Delta9-Tetrahydrocannabinol. Neuropsychopharmacology 2018, 43, 1021–1031. [Google Scholar] [CrossRef]
- Wise, L.E.; Thorpe, A.J.; Lichtman, A.H. Hippocampal CB(1) receptors mediate the memory impairing effects of Δ9-tetrahydrocannabinol. Neuropsychopharmacology 2009, 34, 2072–2080. [Google Scholar] [CrossRef]
- D’Souza, D.C.; Perry, E.; MacDougall, L.; Ammerman, Y.; Cooper, T.; Wu, Y.T.; Braley, G.; Gueorguieva, R.; Krystal, J.H. The psychotomimetic effects of intravenous delta-9-tetrahydrocannabinol in healthy individuals: Implications for psychosis. Neuropsychopharmacology 2004, 29, 1558–1572. [Google Scholar] [CrossRef]
- Lee, K.; Vanin, S.; Nashed, M.; Sarikahya, M.H.; Laviolette, S.R.; Natale, D.R.C.; Hardy, D.B. Cannabidiol Exposure During Gestation Leads to Adverse Cardiac Outcomes Early in Postnatal Life in Male Rat Offspring. Cannabis Cannabinoid Res. 2024, 9, 781–796. [Google Scholar] [CrossRef]
- Xu, Y.; Yao, L.; Guo, Y.; Shi, C.; Zhou, J.; Hua, M. The Potential Antinociceptive Effect and Mechanism of Cannabis sativa L. Extract on Paclitaxel-Induced Neuropathic Pain in Rats Uncovered by Multi-Omics Analysis. Molecules 2024, 29, 1958. [Google Scholar] [CrossRef]
- Rokicki, M.; Żurowski, J.; Sawicki, S.; Ocłoń, E.; Szmatoła, T.; Jasielczuk, I.; Mizera-Szpilka, K.; Semik-Gurgul, E.; Gurgul, A. Impact of Long-Term Cannabidiol (CBD) Treatment on Mouse Kidney Transcriptome. Genes 2024, 15, 1640. [Google Scholar] [CrossRef]
- Carty, D.R.; Thornton, C.; Gledhill, J.H.; Willett, K.L. Developmental Effects of Cannabidiol and Δ9-Tetrahydrocannabinol in Zebrafish. Toxicol. Sci. 2018, 162, 137–145. [Google Scholar] [CrossRef]
- Pandelides, Z.; Aluru, N.; Thornton, C.; Watts, H.E.; Willett, K.L. Transcriptomic Changes and the Roles of Cannabinoid Receptors and PPARγ in Developmental Toxicities Following Exposure to Δ9-Tetrahydrocannabinol and Cannabidiol. Toxicol. Sci. 2021, 182, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Philippot, G.; Forsberg, E.; Tahan, C.; Viberg, H.; Fredriksson, R. A Single δ9-Tetrahydrocannabinol (THC) Dose During Brain Development Affects Markers of Neurotrophy, Oxidative Stress, and Apoptosis. Front. Pharmacol. 2019, 10, 1156. [Google Scholar] [CrossRef] [PubMed]
- Barry, G. Integrating the roles of long and small non-coding RNA in brain function and disease. Mol. Psychiatry 2014, 19, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Lin, L.; Soh, B.S.; Stanton, L.W. Long noncoding RNAs in development and disease of the central nervous system. Trends Genet. 2013, 29, 461–468. [Google Scholar] [CrossRef]
- Qureshi, I.A.; Mehler, M.F. Emerging roles of non-coding RNAs in brain evolution, development, plasticity and disease. Nat. Rev. Neurosci. 2012, 13, 528–541. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, L.; Wang, Y.; Li, H.; Ren, X.; Wei, F.; Yu, W.; Wang, X.; Zhang, L.; Yu, J.; et al. Long noncoding RNA HOTAIR involvement in cancer. Tumor Biol. 2014, 35, 9531–9538. [Google Scholar] [CrossRef]
- Li, J.; Gao, F.; Wei, L.; Chen, L.; Qu, N.; Zeng, L.; Luo, Y.; Huang, X.; Jiang, H. Predict the role of lncRNA in kidney aging based on RNA sequencing. BMC Genom. 2022, 23, 254. [Google Scholar] [CrossRef]
- Fan, C.; Xiong, F.; Tang, Y.; Li, P.; Zhu, K.; Mo, Y.; Wang, Y.; Zhang, S.; Gong, Z.; Liao, Q.; et al. Construction of a lncRNA–mRNA Co-Expression Network for Nasopharyngeal Carcinoma. Front. Oncol. 2022, 12, 809760. [Google Scholar] [CrossRef]
- Bloomfield, M.A.P.; Ashok, A.H.; Volkow, N.D.; Howes, O.D. The effects of Δ9-tetrahydrocannabinol on the dopamine system. Nature 2016, 539, 369–377. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Nady, N.; Qi, C.; Allali-Hassani, A.; Zhu, H.; Pan, P.; Adams-Cioaba, M.A.; Amaya, M.F.; Dong, A.; Vedadi, M.; et al. Methylation-state-specific recognition of histones by the MBT repeat protein L3MBTL2. Nucleic Acids Res. 2009, 37, 2204–2210. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Salzberg, A.C.; Uchigashima, M.; Hasegawa, Y.; Hock, H.; Watanabe, M.; Akbarian, S.; Kawasawa, Y.I.; Futai, K. Activity-Induced Regulation of Synaptic Strength through the Chromatin Reader L3mbtl1. Cell Rep. 2018, 23, 3209–3222. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Li, G.; Sims, R.J., 3rd; Vaquero, A.; Kalakonda, N.; Boccuni, P.; Lee, D.; Erdjument-Bromage, H.; Tempst, P.; Nimer, S.D.; et al. L3MBTL1, a histone-methylation-dependent chromatin lock. Cell 2007, 129, 915–928. [Google Scholar] [CrossRef]
- Ciani, L.; Boyle, K.A.; Dickins, E.; Sahores, M.; Anane, D.; Lopes, D.M.; Gibb, A.J.; Salinas, P.C. Wnt7a signaling promotes dendritic spine growth and synaptic strength through Ca2+/Calmodulin-dependent protein kinase II. Proc. Natl. Acad. Sci. USA 2011, 108, 10732–10737. [Google Scholar] [CrossRef]
- Marchalant, Y.; Brothers, H.M.; Norman, G.J.; Karelina, K.; DeVries, A.C.; Wenk, G.L. Cannabinoids attenuate the effects of aging upon neuroinflammation and neurogenesis. Neurobiol. Dis. 2009, 34, 300–307. [Google Scholar] [CrossRef]
- Palazuelos, J.; Ortega, Z.; Díaz-Alonso, J.; Guzmán, M.; Galve-Roperh, I. CB2 cannabinoid receptors promote neural progenitor cell proliferation via mTORC1 signaling. J. Biol. Chem. 2012, 287, 1198–1209. [Google Scholar] [CrossRef]
- Hosseini, H.; Evans-Martin, S.; Bogomilsky, E.; Pritchett, D.L.; Jones, K.S. Grin2a Hypofunction Impairs Spatial Working Memory and Disrupts Hippocampal Network Oscillations and Excitatory-Inhibitory Balance. Biol. Psychiatry Glob. Open Sci. 2025, 5, 100500. [Google Scholar] [CrossRef]
- Italia, M.; Ferrari, E.; Di Luca, M.; Gardoni, F. GluA3-containing AMPA receptors: From physiology to synaptic dysfunction in brain disorders. Neurobiol. Dis. 2021, 161, 105539. [Google Scholar] [CrossRef]
- Alger, B.E. Retrograde signaling in the regulation of synaptic transmission: Focus on endocannabinoids. Prog. Neurobiol. 2002, 68, 247–286. [Google Scholar] [CrossRef] [PubMed]
- Castillo, P.E.; Younts, T.J.; Chávez, A.E.; Hashimotodani, Y. Endocannabinoid signaling and synaptic function. Neuron 2012, 76, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Low, W.C.; Liu, A.; Wang, B. Centrosomal protein DZIP1 regulates Hedgehog signaling by promoting cytoplasmic retention of transcription factor GLI3 and affecting ciliogenesis. J. Biol. Chem. 2013, 288, 29518–29529. [Google Scholar] [CrossRef]
- Wilson, C.W.; Stainier, D.Y. Vertebrate Hedgehog signaling: Cilia rule. BMC Biol. 2010, 8, 102. [Google Scholar] [CrossRef]
- Huang, B.; Li, X.; Zhu, X. The Role of GM130 in Nervous System Diseases. Front. Neurol. 2021, 12, 743787. [Google Scholar] [CrossRef] [PubMed]
- Munro, S. The golgin coiled-coil proteins of the Golgi apparatus. Cold Spring Harb. Perspect. Biol. 2011, 3, a005256. [Google Scholar] [CrossRef]
- Shamseldin, H.E.; Bennett, A.H.; Alfadhel, M.; Gupta, V.; Alkuraya, F.S. GOLGA2, encoding a master regulator of golgi apparatus, is mutated in a patient with a neuromuscular disorder. Hum. Genet. 2016, 135, 245–251. [Google Scholar] [CrossRef]
- Sim-Selley, L.J. Regulation of cannabinoid CB1 receptors in the central nervous system by chronic cannabinoids. Crit. Rev. Neurobiol. 2003, 15, 91–119. [Google Scholar] [CrossRef]
- Rubino, T.; Realini, N.; Braida, D.; Alberio, T.; Capurro, V.; Viganò, D.; Guidali, C.; Sala, M.; Fasano, M.; Parolaro, D. The depressive phenotype induced in adult female rats by adolescent exposure to THC is associated with cognitive impairment and altered neuroplasticity in the prefrontal cortex. Neurotox. Res. 2009, 15, 291–302. [Google Scholar] [CrossRef]
- Taylor, A.; Nweke, A.; Vincent, V.; Oke, M.; Kulkarni, P.; Ferris, C.F. Chronic exposure to inhaled vaporized cannabis high in Δ9-THC alters brain structure in adult female mice. Front. Neurosci. 2023, 17, 1139309. [Google Scholar] [CrossRef]
- Simon, A. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 24 May 2024).
- Lab, H. FASTX-Toolkit. Available online: https://www.hannonlab.org/resources/ (accessed on 24 April 2024).
- Bushnell, B. BBMap. Available online: https://sourceforge.net/projects/bbmap (accessed on 3 May 2024).
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Choi, M.R.; Jung, K.H.; Park, J.H.; Das, N.D.; Chung, M.K.; Choi, I.G.; Lee, B.C.; Park, K.S.; Chai, Y.G. Ethanol-induced small heat shock protein genes in the differentiation of mouse embryonic neural stem cells. Arch. Toxicol. 2011, 85, 293–304. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.R.; Kim, J.; Park, C.; Chang, S.H.; Kim, H.-N.; Jin, Y.B.; Lee, S.-R. Large-Scale Profiling of Coding and Long Noncoding Transcriptomes in the Hippocampus of Mice Acutely Exposed to Vaporized CBD or THC. Int. J. Mol. Sci. 2025, 26, 7106. https://doi.org/10.3390/ijms26157106
Choi MR, Kim J, Park C, Chang SH, Kim H-N, Jin YB, Lee S-R. Large-Scale Profiling of Coding and Long Noncoding Transcriptomes in the Hippocampus of Mice Acutely Exposed to Vaporized CBD or THC. International Journal of Molecular Sciences. 2025; 26(15):7106. https://doi.org/10.3390/ijms26157106
Chicago/Turabian StyleChoi, Mi Ran, Jihun Kim, Chaeeun Park, Seok Hwan Chang, Han-Na Kim, Yeung Bae Jin, and Sang-Rae Lee. 2025. "Large-Scale Profiling of Coding and Long Noncoding Transcriptomes in the Hippocampus of Mice Acutely Exposed to Vaporized CBD or THC" International Journal of Molecular Sciences 26, no. 15: 7106. https://doi.org/10.3390/ijms26157106
APA StyleChoi, M. R., Kim, J., Park, C., Chang, S. H., Kim, H.-N., Jin, Y. B., & Lee, S.-R. (2025). Large-Scale Profiling of Coding and Long Noncoding Transcriptomes in the Hippocampus of Mice Acutely Exposed to Vaporized CBD or THC. International Journal of Molecular Sciences, 26(15), 7106. https://doi.org/10.3390/ijms26157106