
What a Modern Physician Should Know About microRNAs in the Diagnosis and Treatment of Diabetic Kidney Disease

 , , and
, , and
Abstract
1. Introduction
2. MiRNA Biosynthesis
MiRNA—Mechanism of Action
3. MiRNAs in DKD
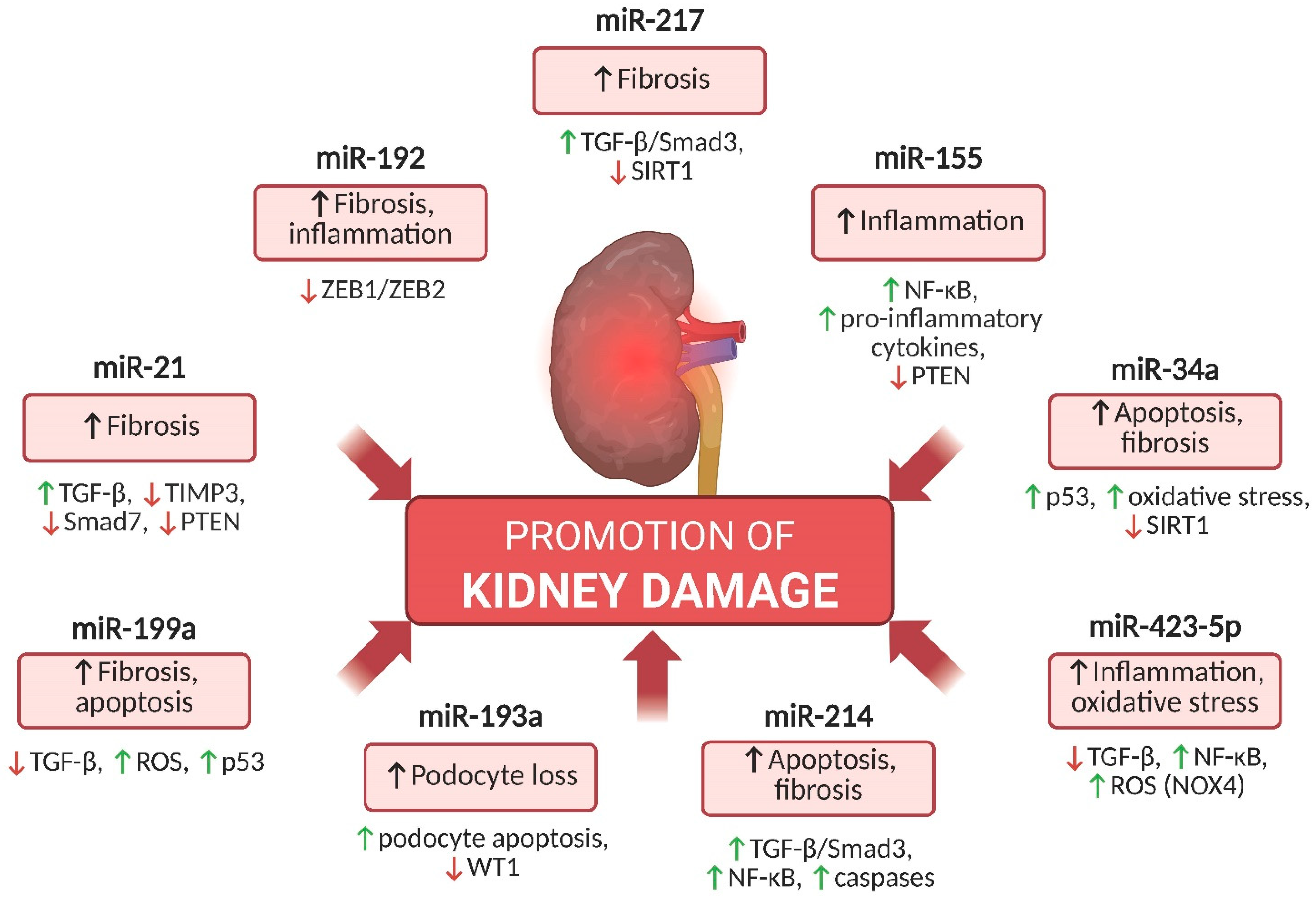
4. Key miRNAs Involved in DKD Pathogenesis That Promote Kidney Injury
4.1. miR-21
4.2. miR-192
4.3. miR-155
4.4. miR-34a
4.5. miR-217
4.6. miR-423-5p
4.7. mir-214
4.8. miR-199a
4.9. miR-193a
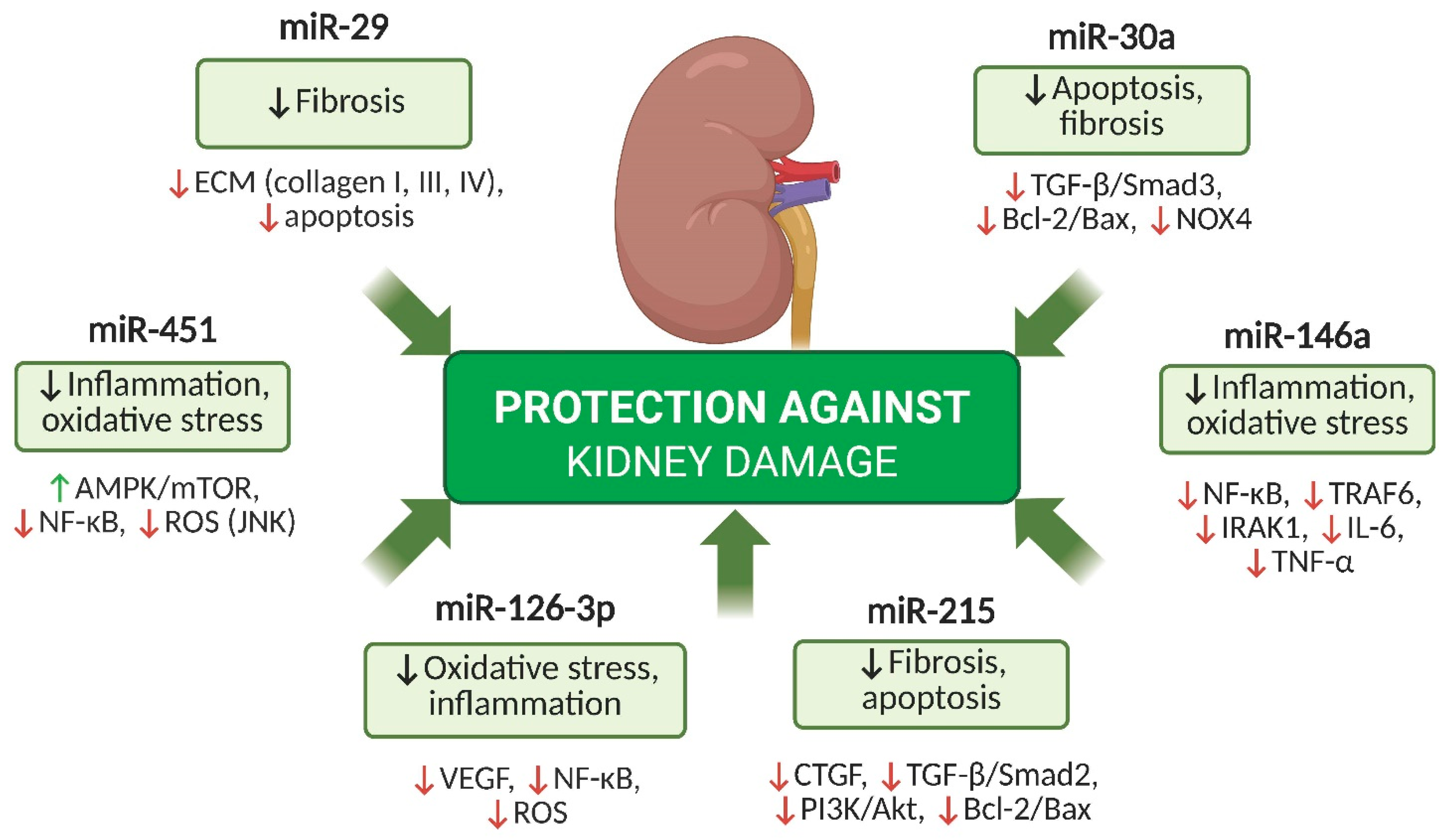
5. Key miRNAs with Protective Potential in DKD
5.1. miR-126-3p
5.2. miR-29
5.3. miR-451
5.4. miR-30a
5.5. miR-146a
5.6. miR-215
6. MiRNAs as Potential Biomarkers in DKD
7. Preclinical Models and Functional Validation of miRNAs in DKD
8. MiRNAs as Therapeutic Targets in DKD: Perspectives and Challenges
9. The Impact of Commonly Available Drugs and Dietary Compounds on miRNA Modulation in DKD
9.1. SGLT2 Inhibitors
9.2. GLP-1 Receptor Agonists
9.3. Statins
9.4. Metformin
9.5. Polyphenols
9.6. Sulforaphane
9.7. The Role of Gut Microbiota in miRNA Modulation in DKD
10. Summary
Funding
Conflicts of Interest
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef] [PubMed]
- Vallon, V.; Thomson, S.C. The tubular hypothesis of nephron filtration and diabetic kidney disease. Nat. Rev. Nephrol. 2020, 16, 317–336. [Google Scholar] [CrossRef]
- Kidney Disease: Improving Global Outcomes (KDIGO) Diabetes Work Group. KDIGO 2022 Clinical Practice Guideline for Diabetes Management in Chronic Kidney Disease. Kidney Int. 2022, 102 (Suppl. S5), S1–S127. [Google Scholar] [CrossRef]
- Ubara, Y.; Sawa, N.; Yamanouchi, M.; Kono, K.; Ohashi, K. New interpretation of diabetic nephropathy or diabetic kidney disease from kidney biopsy: Review article. Clin. Exp. Nephrol. 2025, 29, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, J.; Zhang, J.; Chen, S. Serum VEGF as a predictive marker of glycemic control and diabetic nephropathy in Chinese older adults with type 2 diabetes mellitus. Front. Endocrinol. 2023, 14, 1274025. [Google Scholar] [CrossRef]
- Margaritis, K.; Margioula-Siarkou, G.; Giza, S.; Tsakoumaki, F.; Goulis, D.G.; Koliakos, G.; Kotanidou, E.P. Micro-RNA implications in type 1 diabetes mellitus: A review of literature. Int. J. Mol. Sci. 2021, 22, 12165. [Google Scholar] [CrossRef]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-based diagnosis and therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Ren, H.; Wang, Q. Non-coding RNA and diabetic kidney disease. DNA Cell Biol. 2021, 40, 553–567. [Google Scholar] [CrossRef]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.-P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef]
- Wan, X.; Liao, J.; Lai, H.; Zhang, S.; Cui, J.; Chen, C. Roles of microRNA-192 in diabetic nephropathy: The clinical applications and mechanisms of action. Front. Endocrinol. 2023, 14, 1179161. [Google Scholar] [CrossRef]
- Liu, S.; Wu, W.; Liao, J.; Tang, F.; Gao, G.; Peng, J.; Fu, X.; Zhan, Y.; Chen, Z.; Xu, W.; et al. MicroRNA-21: A critical pathogenic factor of diabetic nephropathy. Front. Endocrinol. 2022, 13, 895010. [Google Scholar] [CrossRef] [PubMed]
- Dhas, Y.; Arshad, N.; Biswas, N.; Jones, L.D.; Ashili, S. MicroRNA-21 silencing in diabetic nephropathy: Insights on therapeutic strategies. Biomedicines 2023, 11, 2583. [Google Scholar] [CrossRef]
- Chen, F.; Wei, G.; Zhou, Y.; Ma, X.; Wang, Q. The mechanism of miR-192 in regulating high glucose-induced MCP-1 expression in rat glomerular mesangial cells. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; He, F.; Zhang, C. Molecular therapeutics for diabetic kidney disease: An update. Int. J. Mol. Sci. 2024, 25, 10051. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, J.; Wang, W.; Xu, M.; Gao, H.; Fang, Y.; Sun, W. Epigenetic modification in diabetic kidney disease. Front. Endocrinol. 2023, 14, 1133970. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Gambardella, J.; Sardu, C.; Lombardi, A.; Santulli, G. Functional role of miR-155 in the pathogenesis of diabetes mellitus and its complications. Noncoding RNA 2021, 7, 39. [Google Scholar] [CrossRef]
- Prieto, I.; Jimenez-Castilla, L.; Lazaro, I.; Bernal-Uribe, S.; Lopez-Sanz, L.; Flores-Muñoz, M.; Egido, J.; Lopez-Franco, O.; Gomez-Guerrero, C. MiR-155/SOCS1 regulatory loop influences diabetic kidney disease by JAK/STAT pathway modulation. Nephrol. Dial. Transplant. 2021, 36 (Suppl. S1), gfab143.004. [Google Scholar] [CrossRef]
- Prieto, I.; Kavanagh, M.; Jimenez-Castilla, L.; Pardines, M.; Lazaro, I.; Herrero del Real, I.; Flores-Muñoz, M.; Egido, J.; Lopez-Franco, O.; Gomez-Guerrero, C. Amutual regulatory loop between miR-155 and SOCS1 influences renal inflammation and diabetic kidney disease. Mol. Ther. Nucleic Acids 2023, 34, 102041. [Google Scholar] [CrossRef]
- Zhao, M.; Qi, Q.; Liu, S.; Huang, R.; Shen, J.; Zhu, Y.; Chai, J.; Zheng, H.; Wu, H.; Liu, H. MicroRNA-34a: A Novel Therapeutic Target in Fibrosis. Front. Physiol. 2022, 13, 895242. [Google Scholar] [CrossRef]
- Qi, W.; Hu, C.; Zhao, D.; Li, X. SIRT1–SIRT7 in diabetic kidney disease: Biological functions and molecular mechanisms. Front. Endocrinol. 2023, 14, 1151378. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Lv, C.; Wu, C.; Zhou, Y.; Wang, Q. miR-217 promotes inflammation and fibrosis in high glucose cultured rat glomerular mesangial cells via Sirt1/HIF-1α signaling pathway. Diabetes Metab. Res. Rev. 2016, 32, 534–543. [Google Scholar] [CrossRef]
- Chen, H.; Guo, Y.; Cheng, X. Long non-coding RNA XIST promoted cell proliferation and suppressed apoptosis by miR-423-5p/HMGA2 axis in diabetic nephropathy. Mol. Cell Biochem. 2021, 476, 4517–4528. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Fan, D.; Chen, B. LncRNA NEAT1 accelerates the proliferation, oxidative stress, inflammation, and fibrosis and suppresses the apoptosis through the miR-423-5p/GLIPR2 axis in diabetic nephropathy. J. Cardiovasc. Pharmacol. 2022, 79, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, J.; Fan, L.; He, X. miR-423-5p suppresses high-glucose-induced podocyte injury by targeting Nox4. Biochem. Biophys. Res. Commun. 2018, 505, 339–345. [Google Scholar] [CrossRef]
- Ma, Z.; Li, L.; Livingston, M.J.; Zhang, D.; Mi, Q.; Zhang, M.; Ding, H.F.; Huo, Y.; Mei, C.; Dong, Z. p53/microRNA-214/ULK1 axis impairs renal tubular autophagy in diabetic kidney disease. J. Clin. Investig. 2020, 130, 5011–5026. [Google Scholar] [CrossRef]
- Szostak, J.; Gorący, A.; Durys, D.; Dec, P.; Modrzewski, A.; Pawlik, A. The role of microRNA in the pathogenesis of diabetic nephropathy. Int. J. Mol. Sci. 2023, 24, 6214. [Google Scholar] [CrossRef]
- Yang, S.; Fei, X.; Lu, Y.; Xu, B.; Ma, Y.; Wan, H. miRNA-214 suppresses oxidative stress in diabetic nephropathy via the ROS/Akt/mTOR signaling pathway and uncoupling protein 2. Exp. Ther. Med. 2019, 17, 3530–3538. [Google Scholar] [CrossRef]
- Yi, Z.; Liu, P.; Zhang, Y.; Mamuti, D.; Zhou, W.; Liu, Z.; Chen, Z. METTL3 aggravates renal fibrogenesis in obstructive nephropathy by promoting the maturation of pri-miR-199a-3p. Eur. J. Pharmacol. 2024, 943, 175628. [Google Scholar]
- Yang, R.; Xu, X.; Li, H.; Chen, J.; Xiang, X.; Dong, Z.; Zhang, D. p53 induces miR-199a-3p to suppress SOCS7 for STAT3 activation in renal fibrosis. Sci. Rep. 2017, 7, 43409. [Google Scholar]
- Wang, P.; Yang, J.; Dai, S.; Gao, P.; Qi, Y.; Zhao, X.; Liu, J.; Wang, Y.; Gao, Y. miRNA-193a-mediated WT1 suppression triggers podocyte injury through activation of the EZH2/β-catenin/NLRP3 pathway in children with diabetic nephropathy. Exp. Cell Res. 2024, 442, 114238. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Kaneko, S.; Yanai, K.; Aomatsu, A.; Hirai, K.; Ookawara, S.; Ishibashi, K.; Morishita, Y. MicroRNAs in podocyte injury in diabetic nephropathy. Front. Genet. 2020, 11, 993. [Google Scholar] [CrossRef] [PubMed]
- Jordan, N.P.; Tingle, S.J.; Shuttleworth, V.G.; Cooke, K.; Redgrave, R.E.; Singh, E.; Glover, E.K.; Tajuddin, H.B.A.; Kirby, J.A.; Arthur, H.M.; et al. MiR-126-3p is dynamically regulated in endothelial-to-mesenchymal transition during fibrosis. Int. J. Mol. Sci. 2021, 22, 8629. [Google Scholar] [CrossRef]
- Assmann, T.S.; Recamonde-Mendoza, M.; Costa, A.R.; de Souza, B.M.; Bauer, A.C.; Crispim, D. MicroRNAs and diabetic kidney disease: Systematic review and bioinformatic analysis. J. Diabetes Complicat. 2018, 32, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Gao, H.; Dai, L.; Han, Y.; Lei, Z.; Wang, X.; Chang, H.; Liu, S.; Wang, Z.; Tong, H.; et al. miR-126 regulates angiogenesis in myocardial ischemia by targeting HIF-1α. Exp. Cell Res. 2021, 409, 112925. [Google Scholar] [CrossRef]
- Gu, Y.-Y.; Liu, X.-S.; Huang, X.-R.; Yu, X.-Q.; Lan, H.-Y. Diverse role of TGF-β in kidney disease. Front. Cell Dev. Biol. 2020, 8, 123. [Google Scholar] [CrossRef]
- Gluba-Sagr, A.; Franczyk, B.; Rysz-Górzyńska, M.; Rysz, J. The role of miRNA in renal fibrosis leading to chronic kidney disease. Biomedicines 2023, 11, 2358. [Google Scholar] [CrossRef]
- Kumari, M.; Mohan, A.; Ecelbarger, C.M.; Saxena, A.; Gupta, A.; Prasad, N.; Tiwari, S. miR-451 loaded exosomes are released by the renal cells in response to injury and associated with reduced kidney function in human. Front. Physiol. 2020, 11, 234. [Google Scholar]
- Garmaa, G.; Bunduc, S.; Kóti, T.; Hegyi, P.; Csupor, D.; Ganbat, D.; Dembrovszky, F.; Meznerics, F.A.; Nasirzadeh, A.; Barbagallo, C.; et al. A systematic review and meta-analysis of microRNA profiling studies in chronic kidney diseases. Noncoding RNA 2024, 10, 30. [Google Scholar] [CrossRef]
- Ning, Y.; Zhou, X.; Wang, G.; Zhang, L.; Wang, J. Exosome miR-30a-5p regulates glomerular endothelial cells’ EndMT and angiogenesis by modulating Notch1/VEGF signaling pathway. Curr. Gene Ther. 2024, 24, 159–177. [Google Scholar] [CrossRef]
- Wu, Q.; Huang, F. LncRNA H19: A novel player in the regulation of diabetic kidney disease. Front. Endocrinol. 2023, 14, 1238981. [Google Scholar] [CrossRef] [PubMed]
- Gilyazova, I.; Asadullina, D.; Kagirova, E.; Sikka, R.; Mustafin, A.; Ivanova, E.; Bakhtiyarova, K.; Gilyazova, G.; Gupta, S.; Khusnutdinova, E.; et al. MiRNA-146a—A key player in immunity and diseases. Int. J. Mol. Sci. 2023, 24, 12767. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Wang, M.; Chen, Z.; Bhatt, K.; Oh, H.J.; Lanting, L.; Deshpande, S.; Jia, Y.; Lai, J.Y.C.; O’Connor, C.L.; et al. An endoplasmic reticulum stress-regulated lncRNA hosting a microRNA megacluster induces early features of diabetic nephropathy. Nat. Commun. 2016, 7, 12864. [Google Scholar] [CrossRef]
- Mohandes, S.; Doke, T.; Hu, H.; Mukhi, D.; Dhillon, P.; Susztak, K. Molecular pathways that drive diabetic kidney disease. J. Clin. Investig. 2023, 133, e165654. [Google Scholar] [CrossRef]
- Negeem, Z.R.; Abdel Moneim, A.A.; Mahmoud, B.; Ahmed, A.E.; Hasona, N.A. Association of microRNA-192, pentraxin-3, and transforming growth factor β1 with diabetic nephropathy: A case-control study. J. Diabetes Metab. Disord. 2023, 22, 123–131. [Google Scholar]
- Sasso, C.V.; Lhamyani, S.; Hevilla, F.; Padial, M.; Blanca, M.; Barril, G.; Jiménez-Salcedo, T.; Sanz Martínez, E.; Nogueira, Á.; Lago-Sampedro, A.M.; et al. Modulation of miR-29a and miR-29b expression and their target genes related to inflammation and renal fibrosis by an oral nutritional supplement with probiotics in malnourished hemodialysis patients. Int. J. Mol. Sci. 2024, 25, 1132. [Google Scholar] [CrossRef] [PubMed]
- Theofilis, P.; Oikonomou, E.; Vogiatzi, G.; Sagris, M.; Antonopoulos, A.S.; Siasos, G.; Iliopoulos, D.C.; Perrea, D.; Vavouranakis, M.; Tsioufis, K.; et al. The Role of MicroRNA-126 in Atherosclerotic Cardiovascular Diseases. Curr. Med. Chem. 2023, 30, 1902–1921. [Google Scholar] [CrossRef]
- Bhatt, K.; Lanting, L.L.; Jia, Y.; Yadav, S.; Reddy, M.A.; Magilnick, N.; Boldin, M.; Natarajan, R. Anti-inflammatory role of microRNA-146a in the pathogenesis of diabetic nephropathy. J. Am. Soc. Nephrol. 2016, 27, 2277–2288. [Google Scholar] [CrossRef]
- Zang, J.; Maxwell, A.P.; Simpson, D.A.; McKay, G.J. Differential expression of urinary exosomal microRNAs miR-21-5p and miR-30b-5p in individuals with diabetic kidney disease. Sci. Rep. 2019, 9, 10900. [Google Scholar] [CrossRef]
- Lee, C.-C.; Chen, C.-C.; Hsu, C.-K.; Chen, Y.-T.; Chen, C.-Y.; Yang, K.-J.; Hung, M.-J.; Wu, I.-W. Urinary microRNA in diabetic kidney disease: A literature review. Medicina 2023, 59, 354. [Google Scholar] [CrossRef]
- Zapała, B.; Kamińska, A.; Piwowar, M.; Paziewska, A.; Gala-Błądzińska, A.; Stępień, E.Ł. miRNA signature of urine extracellular vesicles shows the involvement of inflammatory and apoptotic processes in diabetic chronic kidney disease. Pharm. Res. 2023, 40, 817–832. [Google Scholar] [CrossRef]
- Satake, E.; Krolewski, B.; Kobayashi, H.; Md Dom, Z.I.; Ricca, J.; Wilson, J.M.; Hoon, D.S.B.; Duffin, K.L.; Pezzolesi, M.G.; Krolewski, A.S. Preanalytical considerations in quantifying circulating miRNAs that predict progression to end-stage kidney disease in diabetes. JCI Insight 2024, 9, e174153. [Google Scholar] [CrossRef]
- Hung, P.H.; Hsu, Y.C.; Chen, T.H.; Lin, C.L. Recent advances in diabetic kidney diseases: From kidney injury to kidney fibrosis. Int. J. Mol. Sci. 2021, 22, 11857. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Miao, C.; Wang, J. LncRNA ZEB1-AS1 inhibits renal fibrosis in diabetic nephropathy by regulating the miR-217/MAFB axis. RSC Adv. 2019, 9, 30389–30397. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.Y.; Lu, F.H.; Huang, X.R.; Meng, X.M.; Lan, H.Y. Non-coding RNAs as biomarkers and therapeutic targets for diabetic kidney disease. Front. Pharmacol. 2021, 11, 583528. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, B.; Xue, H.; Zhou, Q.Q.; Peng, L. miR-217 is a useful diagnostic biomarker and regulates human podocyte cells apoptosis via targeting TNFSF11 in membranous nephropathy. Biomed. Res. Int. 2017, 2017, 2168767. [Google Scholar] [CrossRef]
- Zou, C. Advances in the study of miRNAs in chronic kidney disease with cardiovascular complications. Front. Physiol. 2023, 14, 1283597. [Google Scholar] [CrossRef]
- Kölling, M.; Kaucsar, T.; Schauerte, C.; Hübner, A.; Dettling, A.; Park, J.K.; Busch, M.; Wulff, X.; Meier, M.; Scherf, K.; et al. Therapeutic miR-21 silencing ameliorates diabetic kidney disease in mice. Mol. Ther. 2017, 25, 165–180. [Google Scholar] [CrossRef]
- Seyhan, A.A. Trials and tribulations of microRNA therapeutics. Int. J. Mol. Sci. 2024, 25, 1469. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Y.; Tang, J.-Y.; Chen, J.; Li, G.-Q.; Feng, B.; Mu, J. MiR-29a-3p inhibits fibrosis of diabetic kidney disease in diabetic mice via downregulation of DNA methyl transferase 3A and 3B. World J. Diabetes 2024, 15, 123–135. [Google Scholar] [CrossRef]
- Paul, P.; Chacko, L.; Dua, T.K.; Chakraborty, P.; Paul, U.; Phulchand, V.V.; Jha, N.K.; Jha, S.K.; Kandimalla, R.; Dewanjee, S. Nanomedicines for the management of diabetic nephropathy: Present progress and prospects. Front. Endocrinol. 2023, 14, 1236686. [Google Scholar] [CrossRef]
- Fluitt, M.B.; Mohit, N.; Gambhir, K.K.; Nunlee-Bland, G. To the future: The role of exosome-derived microRNAs as markers, mediators, and therapies for endothelial dysfunction in type 2 diabetes mellitus. J. Diabetes Res. 2022, 2022, 5126968. [Google Scholar] [CrossRef] [PubMed]
- Putta, S.; Lanting, L.; Sun, G.; Lawson, G.; Kato, M.; Natarajan, R. Inhibiting microRNA-192 ameliorates renal fibrosis in diabetic nephropathy. J. Am. Soc. Nephrol. 2012, 23, 458–469. [Google Scholar] [CrossRef]
- Xu, C.; Ha, X.; Yang, S.; Tian, X.; Jiang, H. Advances in understanding and treating diabetic kidney disease: Focus on tubulointerstitial inflammation mechanisms. Front. Endocrinol. 2023, 14, 1232790. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, Y.; Guo, Q.; Zhang, L.; Liu, H.; Wang, S.; Wu, X.; Shen, X.; Tao, L. MiR-126-loaded immunoliposomes against vascular endothelial inflammation: In vitro and in vivo evaluation. Pharmaceutics 2023, 15, 1379. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, C.; Jin, Y. The role of exosomes in the pathogenesis and management of diabetic kidney disease: A systematic review and meta-analysis. Front. Endocrinol. 2024, 15, 1398382. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Huo, Z.; He, X.; Liu, F.; Liang, J.; Wu, L.; Yang, D. The role of miR-29 in the mechanism of fibrosis. Mini Rev. Med. Chem. 2023, 23, 1846–1858. [Google Scholar] [CrossRef]
- Jensen, D.M.; Han, P.; Mangala, L.S.; Lopez-Berestein, G.; Sood, A.K.; Liu, J.; Kriegel, A.J.; Usa, K.; Widlansky, M.E.; Liang, M. Broad-acting therapeutic effects of miR-29b-chitosan on hypertension and diabetic complications. Mol. Ther. Nucleic Acids 2022, 29, 1–18. [Google Scholar] [CrossRef]
- Klen, J.; Dolžan, V. SGLT2 inhibitors in the treatment of diabetic kidney disease: More than just glucose regulation. Pharmaceutics 2023, 15, 1995. [Google Scholar] [CrossRef]
- Dai, Z.-C.; Chen, J.-X.; Zou, R.; Liang, X.-B.; Tang, J.-X.; Yao, C.-W. Role and mechanisms of SGLT-2 inhibitors in the treatment of diabetic kidney disease. Front. Immunol. 2023, 14, 1213473. [Google Scholar] [CrossRef]
- Kawanami, D.; Takashi, Y. GLP-1 receptor agonists in diabetic kidney disease: From clinical outcomes to mechanisms. Front. Pharmacol. 2020, 11, 967. [Google Scholar] [CrossRef] [PubMed]
- Górriz, J.L.; Soler, M.J.; Navarro-González, J.F.; García-Carro, C.; Puchades, M.J.; D’Marco, L.; Martínez Castelao, A.; Fernández-Fernández, B.; Ortiz, A.; Górriz-Zambrano, C.; et al. GLP-1 receptor agonists and diabetic kidney disease: A call of attention to nephrologists. J. Clin. Med. 2020, 9, 947. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Zhang, H.; Zhou, X.; Wang, D.; Chen, R.; Tan, W.; Liang, L.; Shi, M.; Zhang, F.; Xiao, Y.; et al. Atorvastatin restores PPARα inhibition of lipid metabolism disorders by downregulating miR-21 expression to improve mitochondrial function and alleviate diabetic nephropathy progression. Front. Pharmacol. 2022, 13, 819787. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ma, X.-Y.; Han, J.-Y.; Yang, M.; Lv, C.; Shao, Y.; Wang, Y.-L.; Kang, J.-Y.; Wang, Q.-Y. Metformin regulates inflammation and fibrosis in diabetic kidney disease through TNC/TLR4/NF-κB/miR-155-5p inflammatory loop. World J. Diabetes 2021, 12, 19–46. [Google Scholar] [CrossRef]
- Lewis, K.A.; Stroebel, B.; Zhang, L.; Aouizerat, B.; Mattis, A.; Flowers, E. MicroRNAs associated with metformin treatment in the diabetes prevention program. Int. J. Mol. Sci. 2024, 25, 5684. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyl, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary polyphenols and their role in oxidative stress-induced human diseases: Insights into protective effects, antioxidant potentials and mechanism(s) of action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef]
- Shabbir, U.; Rubab, M.; Daliri, E.B.; Oh, D.H.; Paik, H.D. Curcumin, quercetin, catechins and metabolic diseases: The role of gut microbiota. Nutrients 2021, 13, 206. [Google Scholar] [CrossRef]
- Cione, E.; La Torre, C.; Cannataro, R.; Caroleo, M.C.; Plastina, P.; Gallelli, L. Quercetin, epigallocatechin gallate, curcumin, and resveratrol: From dietary sources to human microRNA modulation. Molecules 2019, 25, 63. [Google Scholar] [CrossRef]
- Yu, Z.; He, W.; Shi, W. Sulforaphane (Sul) reduces renal interstitial fibrosis (RIF) by controlling the inflammation and TGF-β/Smad signaling pathway. Appl. Biol. Chem. 2024, 67, 8. [Google Scholar] [CrossRef]
- Mao, Z.H.; Gao, Z.X.; Liu, D.W.; Liu, Z.S.; Wu, P. Gut microbiota and its metabolites—Molecular mechanisms and management strategies in diabetic kidney disease. Front. Immunol. 2023, 14, 1124704. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, L.; Zhang, Y.; Li, K.; Yang, J. The role and mechanism of the gut microbiota in the development and treatment of diabetic kidney disease. Front. Physiol. 2023, 14, 1166685. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Liu, N.; Zheng, B.; Guo, F.; Zeng, X.; Huang, X.; Ouyang, D. Roles of gut microbial metabolites in diabetic kidney disease. Front. Endocrinol. 2021, 12, 636175. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, L.; Wei, W.; Fu, P. Efficacy of probiotics/synbiotics supplementation in patients with chronic kidney disease: A systematic review and meta-analysis of randomized controlled trials. Front. Nutr. 2024, 11, 1434613. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, C.; Lv, M.; Hu, Q.; Guo, L.; Xiong, D. Correlation between alterations of gut microbiota and miR-122-5p expression in patients with type 2 diabetes mellitus. Ann. Transl. Med. 2020, 8, 1481. [Google Scholar] [CrossRef]
- Lei, Z.; Xu, M.; Li, Y.; Chen, L.; Li, H. probiotics and nutrients in cardiovascular and kidney disease. Nutrients 2023, 15, 4284. [Google Scholar] [CrossRef]
- Paquette, S.; Thomas, S.C.; Venkataraman, K.; Appanna, V.D.; Tharmalingam, S. The effects of oral probiotics on type 2 diabetes mellitus (T2DM): A clinical trial systematic literature review. Nutrients 2023, 15, 4690. [Google Scholar] [CrossRef]
- Matz, L.M.; Geesala, R.; Gaddam, R.R.; Shi, X.Z. Editorial: Understanding the role of gut hormones, microbiota, and miRNAs in metabolic regulation and glucose homeostasis in obesity and type-2-diabetes. Front. Endocrinol. 2023, 14, 1255942. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| miRNA | Biomarker Function | Clinical Application in Diagnostic DKD |
|---|---|---|
| miR-192 | Regulator of fibrosis | High expression in early stages; marker for early kidney damage; potential therapeutic target |
| miR-21 | Regulator of fibrosis and inflammation | Indicator of disease progression and fibrosis; potential therapeutic target |
| miR-29 | Anti-fibrotic activity | Decreases with disease progression; potential early-stage marker |
| miR-126-3p | Endothelial function and vascular health marker | Associated with vascular complications and endothelial health; potential indicator of vascular damage in DKD |
| miR-146a | Anti-inflammatory function | Reduced levels in advanced DKD; potential therapeutic target |
| miR-215 | Fibrosis regulator | Promising early-stage biomarker; indicator of disease progression |
| miR-217 | Cell apoptosis regulator | Associated with various DKD stages; potential marker for cellular health and disease progression |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodzoń-Norwicz, M.; Kogut, P.; Sowa-Kućma, M.; Gala-Błądzińska, A. What a Modern Physician Should Know About microRNAs in the Diagnosis and Treatment of Diabetic Kidney Disease. Int. J. Mol. Sci. 2025, 26, 6662. https://doi.org/10.3390/ijms26146662
Rodzoń-Norwicz M, Kogut P, Sowa-Kućma M, Gala-Błądzińska A. What a Modern Physician Should Know About microRNAs in the Diagnosis and Treatment of Diabetic Kidney Disease. International Journal of Molecular Sciences. 2025; 26(14):6662. https://doi.org/10.3390/ijms26146662
Chicago/Turabian StyleRodzoń-Norwicz, Małgorzata, Patryk Kogut, Magdalena Sowa-Kućma, and Agnieszka Gala-Błądzińska. 2025. "What a Modern Physician Should Know About microRNAs in the Diagnosis and Treatment of Diabetic Kidney Disease" International Journal of Molecular Sciences 26, no. 14: 6662. https://doi.org/10.3390/ijms26146662
APA StyleRodzoń-Norwicz, M., Kogut, P., Sowa-Kućma, M., & Gala-Błądzińska, A. (2025). What a Modern Physician Should Know About microRNAs in the Diagnosis and Treatment of Diabetic Kidney Disease. International Journal of Molecular Sciences, 26(14), 6662. https://doi.org/10.3390/ijms26146662