
Altered HDL Phospholipid and Fatty Acid Profile in MASLD: A Possible Explanation for the Increased CVD Risk

 ,
,  ,
,
Abstract

1. Introduction
2. Results
2.1. Characteristics of Study Population
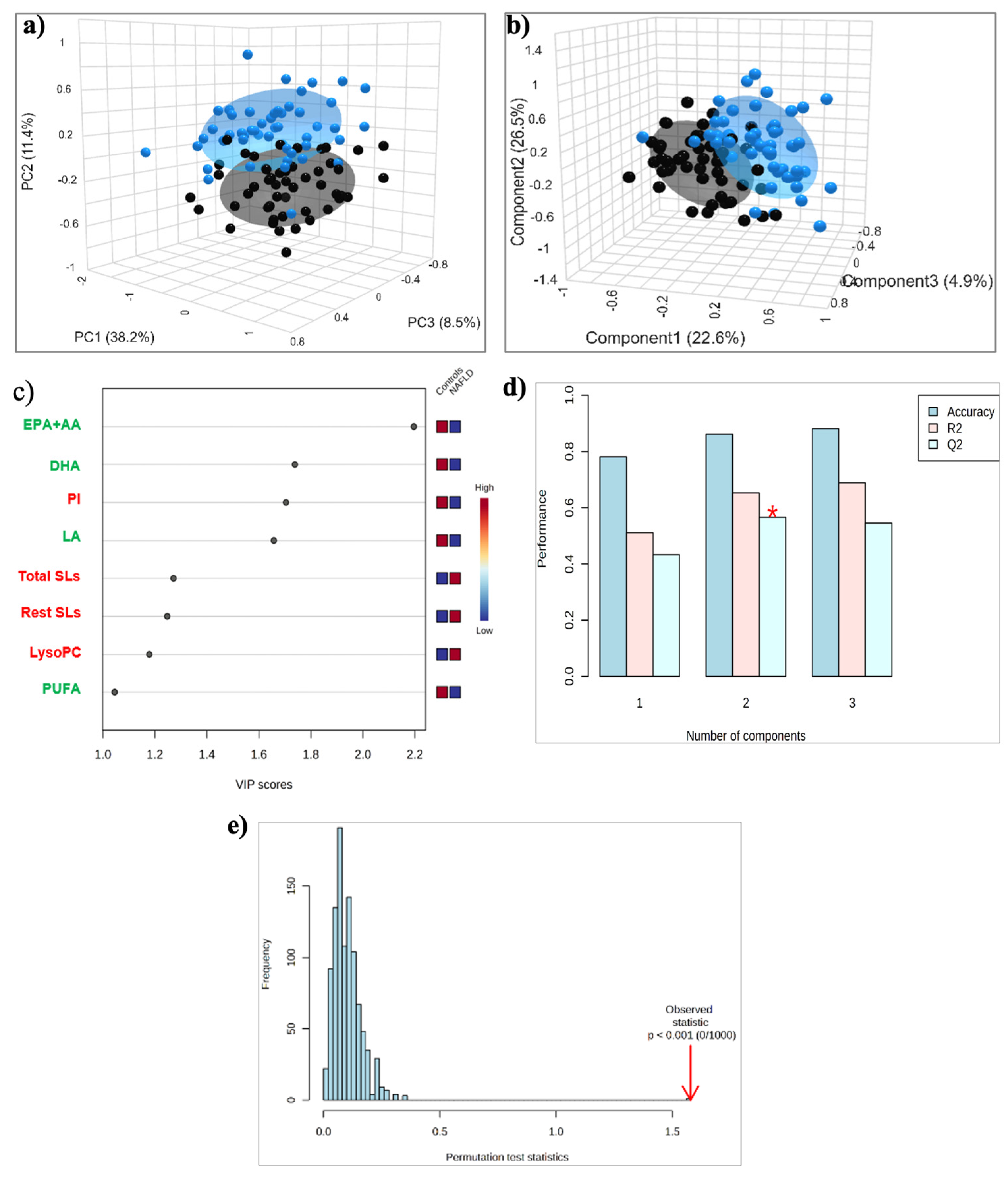
2.2. HDL Lipidome: MASLD vs. Control Group
2.2.1. Targeted Lipid Analysis of HDLs
2.2.2. Untargeted Lipid Analysis of HDLs
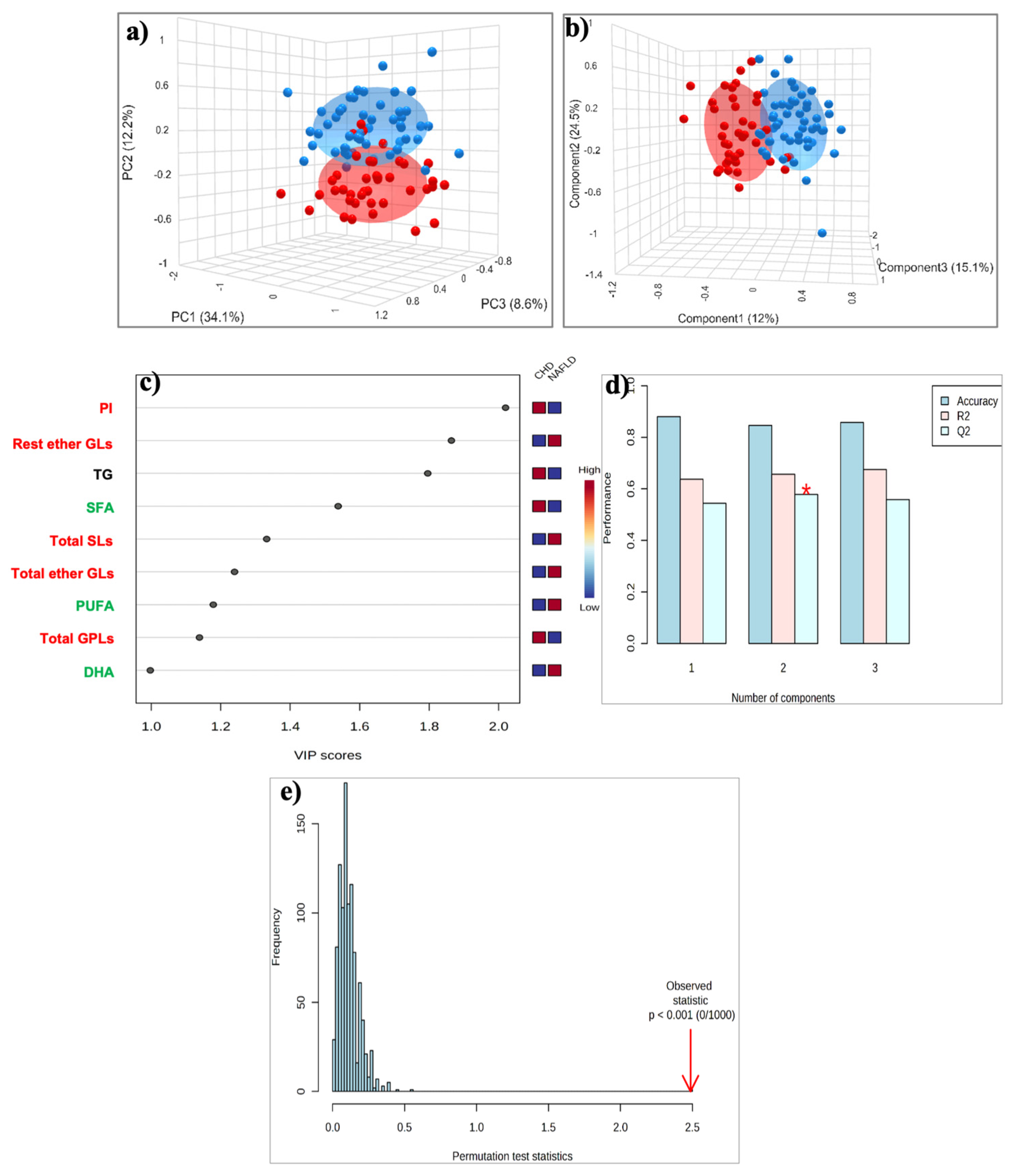
2.3. HDL Lipidome: MASLD vs. CAD Group
2.3.1. Targeted Lipid Analysis
2.3.2. Untargeted Lipid Analysis
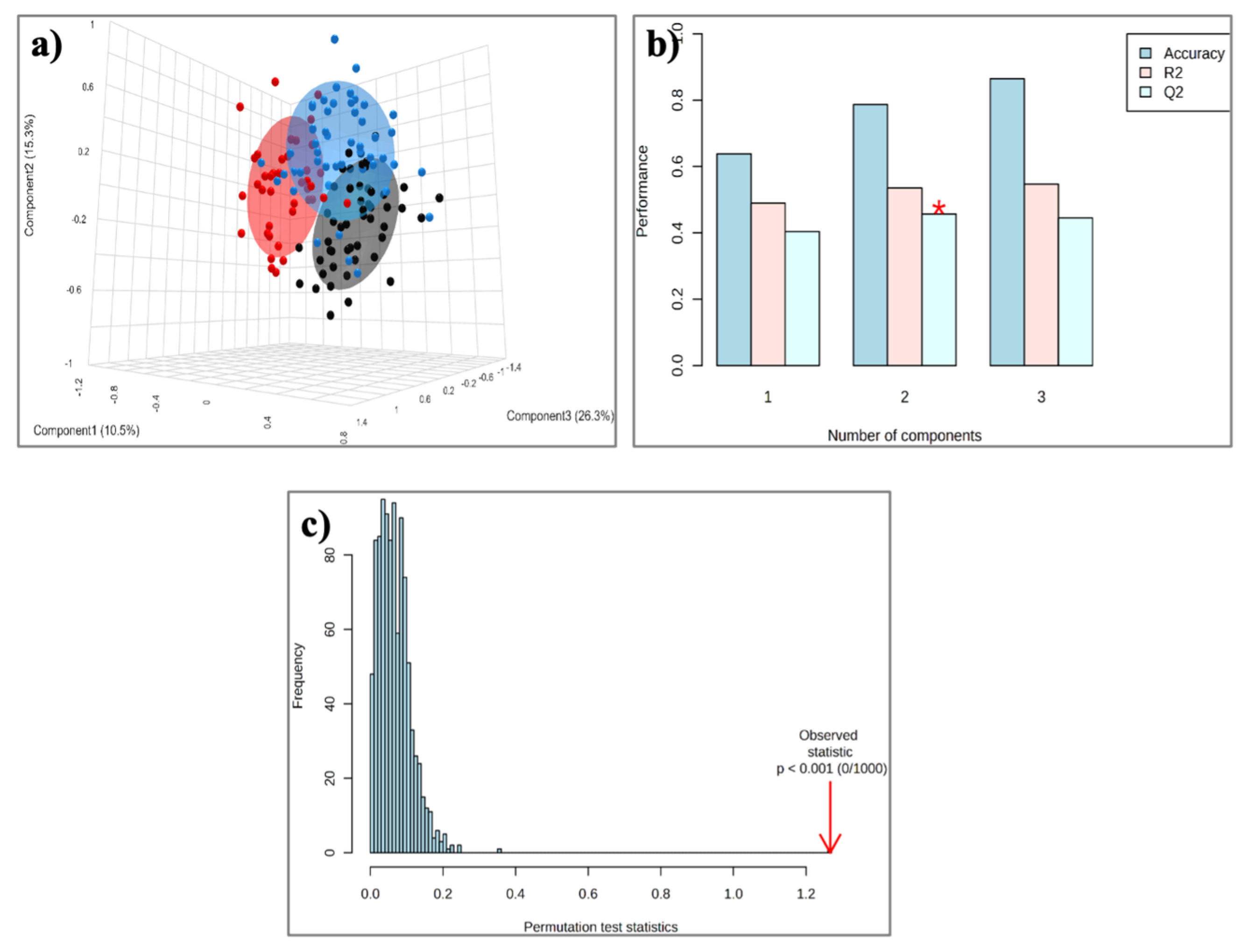
2.4. HDL Lipidome: Control Group vs. MASLD vs. CAD Group
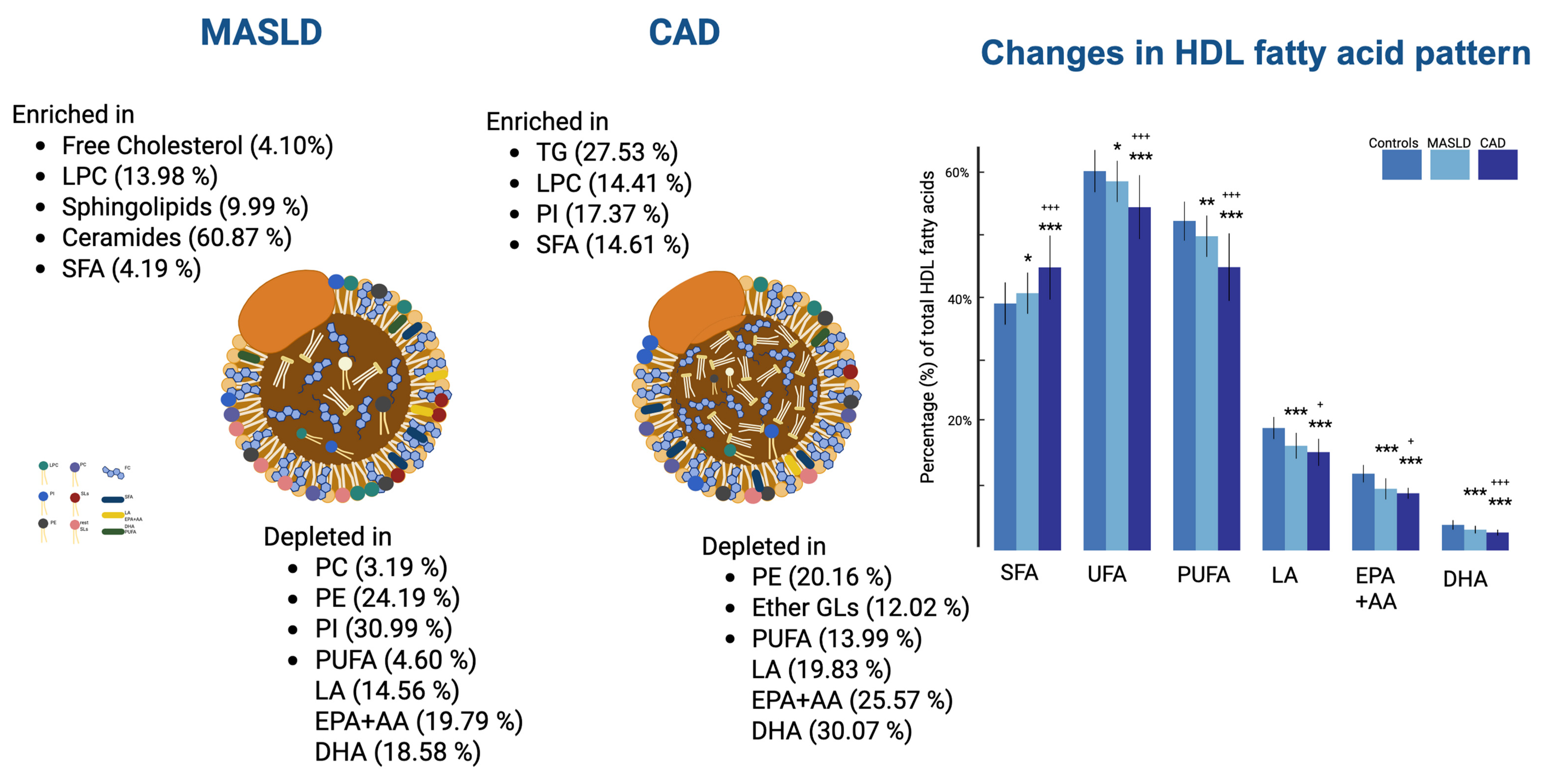
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Sample Collection and Preparation
4.3. Biochemical Parameters
4.4. Isolation and Lipid Extraction of HDL Lipoproteins
H NMR Spectroscopy
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1H NMR | proton nuclear magnetic resonance |
| ABCA1 | ATP-binding cassette subfamily A member 1 |
| apoAI | apolipoprotein-AI |
| CE | cholesteryl ester |
| Cer | ceramide |
| CETP | cholesteryl ester transfer protein |
| CVD | cardiovascular disease |
| HDL | high-density lipoprotein |
| LCAT | lecithin-cholesterol acyltransferase |
| LDL | low-density lipoprotein |
| LPC | lysophosphatidylcholine |
| LpPLA2 | lipoprotein phospholipase A2 |
| MASLD | metabolic dysfunction-associated steatotic liver disease |
| MASH | metabolic dysfunction-associated steatohepatitis |
| PC | phosphatidylcholine |
| PE | phosphatidylethanolamine |
| PI | phosphatidylinositol |
| PUFA | polyunsaturated fatty acids |
| SFA | saturated fatty acids |
| SM | sphingomyelin |
| SR-B1 | scavenger receptor class B type 1 |
| SREBPs | sterol regulatory element-binding proteins |
| TG | triglycerides |
| VLDL | very-low-density lipoprotein |
References
- Younossi, Z.M.; Kalligeros, M.; Henry, L. Epidemiology of Metabolic Dysfunction-Associated Steatotic Liver Disease. Clin. Mol. Hepatol. 2024, 31, S32. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines on the Management of Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD). J. Hepatol. 2024, 81, 492–542. [Google Scholar] [CrossRef] [PubMed]
- Meikle, P.J.; Summers, S.A. Sphingolipids and Phospholipids in Insulin Resistance and Related Metabolic Disorders. Nat. Rev. Endocrinol. 2017, 13, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Targher, G.; Byrne, C.D.; Lonardo, A.; Zoppini, G.; Barbui, C. Non-Alcoholic Fatty Liver Disease and Risk of Incident Cardiovascular Disease: A Meta-Analysis. J. Hepatol. 2016, 65, 589–600. [Google Scholar] [CrossRef]
- Martino, F.; Bassareo, P.P.; Martino, E.; Romeo, F.; Calcaterra, G.; Perrone Filardi, P.; Indolfi, C.; Nodari, S.; Montemurro, V.; Guccione, P.; et al. Cardiovascular Prevention in Childhood: A Consensus Document of the Italian Society of Cardiology Working Group on Congenital Heart Disease and Cardiovascular Prevention in Paediatric Age. J. Cardiovasc. Med. 2023, 24, 492–505. [Google Scholar] [CrossRef]
- Rye, K.-A.; Barter, P.J. Cardioprotective Functions of HDLs. J. Lipid Res. 2014, 55, 168–179. [Google Scholar] [CrossRef]
- Sviridov, D.; Mukhamedova, N.; Remaley, A.T.; Chin-Dusting, J.; Nestel, P. Antiatherogenic Functionality of High Density Lipoprotein: How Much versus How Gooden-Subtitle=. J. Atheroscler. Thromb. 2008, 15, 52–62. [Google Scholar] [CrossRef]
- Ferretti, G.; Bacchetti, T.; Nègre-Salvayre, A.; Salvayre, R.; Dousset, N.; Curatola, G. Structural Modifications of HDL and Functional Consequences. Atherosclerosis 2006, 184, 1–7. [Google Scholar] [CrossRef]
- de la Llera Moya, M.; McGillicuddy, F.C.; Hinkle, C.C.; Byrne, M.; Joshi, M.R.; Nguyen, V.; Tabita-Martinez, J.; Wolfe, M.L.; Badellino, K.; Pruscino, L.; et al. Inflammation Modulates Human HDL Composition and Function in Vivo. Atherosclerosis 2012, 222, 390–394. [Google Scholar] [CrossRef]
- Mocciaro, G.; Allison, M.; Jenkins, B.; Azzu, V.; Huang-Doran, I.; Herrera-Marcos, L.V.; Hall, Z.; Murgia, A.; Susan, D.; Frontini, M.; et al. Non-Alcoholic Fatty Liver Disease Is Characterised by a Reduced Polyunsaturated Fatty Acid Transport via Free Fatty Acids and High-Density Lipoproteins (HDL). Mol. Metab. 2023, 73, 101728. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Global Analyses of Cellular Lipidomes Directly from Crude Extracts of Biological Samples by ESI Mass Spectrometry: A Bridge to Lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Oresic, M.; Hänninen, V.A.; Vidal-Puig, A. Lipidomics: A New Window to Biomedical Frontiers. Trends Biotechnol. 2008, 26, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Vosegaard, T.; Guo, Z. Applications of Nuclear Magnetic Resonance in Lipid Analyses: An Emerging Powerful Tool for Lipidomics Studies. Prog. Lipid Res. 2017, 68, 37–56. [Google Scholar] [CrossRef]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A Lipidomic Analysis of Nonalcoholic Fatty Liver Disease. Hepatology 2007, 46, 1081–1090. [Google Scholar] [CrossRef]
- Wattacheril, J.; Seeley, E.H.; Angel, P.; Chen, H.; Bowen, B.P.; Lanciault, C.; Caprioli, R.M.; Abumrad, N.; Flynn, C.R. Differential Intrahepatic Phospholipid Zonation in Simple Steatosis and Nonalcoholic Steatohepatitis. PLoS ONE 2013, 8, e57165. [Google Scholar] [CrossRef]
- Chiappini, F.; Coilly, A.; Kadar, H.; Gual, P.; Tran, A.; Desterke, C.; Samuel, D.; Duclos-Vallée, J.-C.; Touboul, D.; Bertrand-Michel, J.; et al. Metabolism Dysregulation Induces a Specific Lipid Signature of Nonalcoholic Steatohepatitis in Patients. Sci. Rep. 2017, 7, 46658. [Google Scholar] [CrossRef]
- Mocciaro, G. High-Density Lipoprotein Lipidome: A Neglected Source of Hepatic Lipids. Nat. Rev. Gastroenterol. Hepatol. 2025, 22, 8. [Google Scholar] [CrossRef]
- Singaraja, R.R.; Van Eck, M.; Bissada, N.; Zimetti, F.; Collins, H.L.; Hildebrand, R.B.; Hayden, A.; Brunham, L.R.; Kang, M.H.; Fruchart, J.-C.; et al. Both Hepatic and Extrahepatic ABCA1 Have Discrete and Essential Functions in the Maintenance of Plasma High-Density Lipoprotein Cholesterol Levels in Vivo. Circulation 2006, 114, 1301–1309. [Google Scholar] [CrossRef]
- Cole, L.K.; Vance, J.E.; Vance, D.E. Phosphatidylcholine Biosynthesis and Lipoprotein Metabolism. Biochim. Biophys. Acta 2012, 1821, 754–761. [Google Scholar] [CrossRef]
- Walker, A.K.; Jacobs, R.L.; Watts, J.L.; Rottiers, V.; Jiang, K.; Finnegan, D.M.; Shioda, T.; Hansen, M.; Yang, F.; Niebergall, L.J.; et al. A Conserved SREBP-1/Phosphatidylcholine Feedback Circuit Regulates Lipogenesis in Metazoans. Cell 2011, 147, 840–852. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The Critical Role of Phosphatidylcholine and Phosphatidylethanolamine Metabolism in Health and Disease. Biochim. Biophys. Acta BBA Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef] [PubMed]
- Kartsoli, S.; Kostara, C.E.; Tsimihodimos, V.; Bairaktari, E.T.; Christodoulou, D.K. Lipidomics in Non-Alcoholic Fatty Liver Disease. World J. Hepatol. 2020, 12, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Shama, S.; Jang, H.; Wang, X.; Zhang, Y.; Shahin, N.N.; Motawi, T.K.; Kim, S.; Gawrieh, S.; Liu, W. Phosphatidylethanolamines Are Associated with Nonalcoholic Fatty Liver Disease (NAFLD) in Obese Adults and Induce Liver Cell Metabolic Perturbations and Hepatic Stellate Cell Activation. Int. J. Mol. Sci. 2023, 24, 1034. [Google Scholar] [CrossRef]
- Law, S.-H.; Chan, M.-L.; Marathe, G.K.; Parveen, F.; Chen, C.-H.; Ke, L.-Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef]
- Nass, K.J.; van den Berg, E.H.; Gruppen, E.G.; Dullaart, R.P.F. Plasma Lecithin:Cholesterol Acyltransferase and Phospholipid Transfer Protein Activity Independently Associate with Nonalcoholic Fatty Liver Disease. Eur. J. Clin. Investig. 2018, 48, e12988. [Google Scholar] [CrossRef]
- Colak, Y.; Senates, E.; Ozturk, O.; Doganay, H.L.; Coskunpinar, E.; Oltulu, Y.M.; Eren, A.; Sahin, O.; Ozkanli, S.; Enc, F.Y.; et al. Association of Serum Lipoprotein-Associated Phospholipase A2 Level with Nonalcoholic Fatty Liver Disease. Metab. Syndr. Relat. Disord. 2012, 10, 103–109. [Google Scholar] [CrossRef]
- Gorden, D.L.; Ivanova, P.T.; Myers, D.S.; McIntyre, J.O.; VanSaun, M.N.; Wright, J.K.; Matrisian, L.M.; Brown, H.A. Increased Diacylglycerols Characterize Hepatic Lipid Changes in Progression of Human Nonalcoholic Fatty Liver Disease; Comparison to a Murine Model. PLoS ONE 2011, 6, e22775. [Google Scholar] [CrossRef]
- Pagadala, M.; Kasumov, T.; McCullough, A.J.; Zein, N.N.; Kirwan, J.P. Role of Ceramides in Nonalcoholic Fatty Liver Disease. Trends Endocrinol. Metab. 2012, 23, 365–371. [Google Scholar] [CrossRef]
- Kontush, A.; Lhomme, M.; Chapman, M.J. Unraveling the Complexities of the HDL Lipidome1. J. Lipid Res. 2013, 54, 2950–2963. [Google Scholar] [CrossRef]
- Rye, K.A.; Duong, M.N. Influence of Phospholipid Depletion on the Size, Structure, and Remodeling of Reconstituted High Density Lipoproteins. J. Lipid Res. 2000, 41, 1640–1650. [Google Scholar] [CrossRef]
- Yancey, P.G.; de la Llera-Moya, M.; Swarnakar, S.; Monzo, P.; Klein, S.M.; Connelly, M.A.; Johnson, W.J.; Williams, D.L.; Rothblat, G.H. High Density Lipoprotein Phospholipid Composition Is a Major Determinant of the Bi-Directional Flux and Net Movement of Cellular Free Cholesterol Mediated by Scavenger Receptor BI. J. Biol. Chem. 2000, 275, 36596–36604. [Google Scholar] [CrossRef] [PubMed]
- Rached, F.; Lhomme, M.; Camont, L.; Gomes, F.; Dauteuille, C.; Robillard, P.; Santos, R.D.; Lesnik, P.; Serrano, C.V.; Chapman, M.J.; et al. Defective Functionality of Small, Dense HDL3 Subpopulations in ST Segment Elevation Myocardial Infarction: Relevance of Enrichment in Lysophosphatidylcholine, Phosphatidic Acid and Serum Amyloid A. Biochim. Biophys. Acta 2015, 1851, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Fadaei, R.; Poustchi, H.; Meshkani, R.; Moradi, N.; Golmohammadi, T.; Merat, S. Impaired HDL Cholesterol Efflux Capacity in Patients with Non-Alcoholic Fatty Liver Disease Is Associated with Subclinical Atherosclerosis. Sci. Rep. 2018, 8, 11691. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.W.; Rye, K.A.; Gamble, J.R.; Vadas, M.A.; Barter, P.J. Phospholipid Composition of Reconstituted High Density Lipoproteins Influences Their Ability to Inhibit Endothelial Cell Adhesion Molecule Expression. J. Lipid Res. 2000, 41, 1261–1267. [Google Scholar] [CrossRef]
- Stamler, C.J.; Breznan, D.; Neville, T.A.-M.; Viau, F.J.; Camlioglu, E.; Sparks, D.L. Phosphatidylinositol Promotes Cholesterol Transport in Vivo. J. Lipid Res. 2000, 41, 1214–1221. [Google Scholar] [CrossRef]
- Walle, P.; Takkunen, M.; Männistö, V.; Vaittinen, M.; Lankinen, M.; Kärjä, V.; Käkelä, P.; Ågren, J.; Tiainen, M.; Schwab, U.; et al. Fatty Acid Metabolism Is Altered in Non-Alcoholic Steatohepatitis Independent of Obesity. Metabolism 2016, 65, 655–666. [Google Scholar] [CrossRef]
- Musso, G.; Cassader, M.; Paschetta, E.; Gambino, R. Bioactive Lipid Species and Metabolic Pathways in Progression and Resolution of Nonalcoholic Steatohepatitis. Gastroenterology 2018, 155, 282–302.e8. [Google Scholar] [CrossRef]
- Papathanasiou, A.; Kostara, C.; Cung, M.-T.; Seferiadis, K.; Elisaf, M.; Bairaktari, E.; Goudevenos, I.A. Analysis of the Composition of Plasma Lipoproteins in Patients with Extensive Coronary Heart Disease Using 1H NMR Spectroscopy. Hell. J. Cardiol. 2008, 49, 72–78. [Google Scholar]
- Kostara, C.E.; Papathanasiou, A.; Psychogios, N.; Cung, M.T.; Elisaf, M.S.; Goudevenos, J.; Bairaktari, E.T. NMR-Based Lipidomic Analysis of Blood Lipoproteins Differentiates the Progression of Coronary Heart Disease. J. Proteome Res. 2014, 13, 2585–2598. [Google Scholar] [CrossRef]
- Kostara, C.E.; Papathanasiou, A.; Cung, M.T.; Elisaf, M.S.; Goudevenos, J.; Bairaktari, E.T. Evaluation of Established Coronary Heart Disease on the Basis of HDL and Non-HDL NMR Lipid Profiling. J. Proteome Res. 2010, 9, 897–911. [Google Scholar] [CrossRef]
- Park, K.-H.; Shin, D.-G.; Kim, J.-R.; Hong, J.-H.; Cho, K.-H. The Functional and Compositional Properties of Lipoproteins Are Altered in Patients with Metabolic Syndrome with Increased Cholesteryl Ester Transfer Protein Activity. Int. J. Mol. Med. 2010, 25, 129–136. [Google Scholar] [PubMed]
- Hansel, B.; Giral, P.; Nobecourt, E.; Chantepie, S.; Bruckert, E.; Chapman, M.J.; Kontush, A. Metabolic Syndrome Is Associated with Elevated Oxidative Stress and Dysfunctional Dense High-Density Lipoprotein Particles Displaying Impaired Antioxidative Activity. J. Clin. Endocrinol. Metab. 2004, 89, 4963–4971. [Google Scholar] [CrossRef] [PubMed]
- Kostara, C.E.; Ferrannini, E.; Bairaktari, E.T.; Papathanasiou, A.; Elisaf, M.; Tsimihodimos, V. Early Signs of Atherogenic Features in the HDL Lipidomes of Normolipidemic Patients Newly Diagnosed with Type 2 Diabetes. Int. J. Mol. Sci. 2020, 21, 8835. [Google Scholar] [CrossRef] [PubMed]
- Adeli, K.; Taghibiglou, C.; Van Iderstine, S.C.; Lewis, G.F. Mechanisms of Hepatic Very Low-Density Lipoprotein Overproduction in Insulin Resistance. Trends Cardiovasc. Med. 2001, 11, 170–176. [Google Scholar] [CrossRef]
- Curtiss, L.K.; Bonnet, D.J.; Rye, K.-A. The Conformation of Apolipoprotein A-I in High-Density Lipoproteins Is Influenced by Core Lipid Composition and Particle Size: A Surface Plasmon Resonance Study. Biochemistry 2000, 39, 5712–5721. [Google Scholar] [CrossRef]
- Lamarche, B.; Uffelman, K.D.; Carpentier, A.; Cohn, J.S.; Steiner, G.; Barrett, P.H.; Lewis, G.F. Triglyceride Enrichment of HDL Enhances in Vivo Metabolic Clearance of HDL Apo A-I in Healthy Men. J. Clin. Investig. 1999, 103, 1191–1199. [Google Scholar] [CrossRef]
- Skeggs, J.W.; Morton, R.E. LDL and HDL Enriched in Triglyceride Promote Abnormal Cholesterol Transport. J. Lipid Res. 2002, 43, 1264–1274. [Google Scholar] [CrossRef]
- Greene, D.J.; Skeggs, J.W.; Morton, R.E. Elevated Triglyceride Content Diminishes the Capacity of High Density Lipoprotein to Deliver Cholesteryl Esters via the Scavenger Receptor Class B Type I (SR-BI). J. Biol. Chem. 2001, 276, 4804–4811. [Google Scholar] [CrossRef]
- Rallidis, L.S.; Rizos, C.V.; Papathanasiou, K.A.; Liamis, G.; Skoumas, I.; Garoufi, A.; Kolovou, G.; Tziomalos, K.; Skalidis, E.; Kotsis, V.; et al. Physical Signs and Atherosclerotic Cardiovascular Disease in Familial Hypercholesterolemia: The HELLAS-FH Registry. J. Cardiovasc. Med. 2024, 25, 370–378. [Google Scholar] [CrossRef]
- Roffi, M.; Patrono, C.; Collet, J.-P.; Mueller, C.; Valgimigli, M.; Andreotti, F.; Bax, J.J.; Borger, M.A.; Brotons, C.; Chew, D.P.; et al. 2015 ESC Guidelines for the Management of Acute Coronary Syndromes in Patients Presenting without Persistent ST-Segment Elevation: Task Force for the Management of Acute Coronary Syndromes in Patients Presenting without Persistent ST-Segment Elevation of the European Society of Cardiology (ESC). Eur. Heart J. 2016, 37, 267–315. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Kostara, C.E.; Bairaktari, E.T. Lipid Profiling in Health and Disease. In Methodologies for Metabolomics: Experimental Strategies and Techniques; Lutz, N.W., Sweedler, J.V., Wevers, R.A., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. 317–332. ISBN 978-0-521-76590-9. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (n = 141) | Controls (n = 50) | MASLD 1 (n = 51) | CAD 2 (n = 40) | p-Value |
| Age (years) | 54.3 ± 12.4 | 51.4 ± 12.9 | 55.2 ± 8.5 | NS |
| Gender (Males/Females) | 26/24 | 23/28 | 22/18 | |
| Serum Lipid Parameters (mg/dL) | ||||
| Total Cholesterol | 197.9 ± 30.5 | 207.6 ± 47.7 | 204.3 ± 45.4 | NS |
| Triglycerides | 121.9 ± 33.7 | 132.4 ± 48.6 | 131.4 ± 64.8 | NS |
| HDL Cholesterol | 48.9 ± 11.5 | 51.3 ± 12.4 | 48.4 ± 10.1 | NS |
| LDL Cholesterol | 124.7 ± 29.4 | 130.0 ± 39.9 | 129.5 ± 43.2 | NS |
| non-HDL Cholesterol | 149.0 ± 28.4 | 156.2 ± 43.5 | 155.8 ± 42.3 | NS |
| Total (n = 141) | Controls (n = 50) | MASLD 1 (n = 51) | CAD 2 (n = 40) |
| % Cholesterol, total | 41.87 ± 1.54 | 42.13 ± 1.79 | 40.63 ± 1.87 **,+++ |
| Esterified (EC) | 32.12 ± 1.37 | 31.98 ± 1.29 | 30.89 ± 1.69 ***,+++ |
| Free (FC) | 9.75 ± 0.84 | 10.15 ± 0.96 * | 9.74 ± 0.86 + |
| % Triglycerides (TG) | 4.65 ± 1.12 | 4.74 ± 1.72 | 5.93 ± 1.11 ***,+++ |
| % Phospholipids (PLs), total | 53.47 ± 1.66 | 53.14 ± 2.06 | 53.44 ± 1.83 |
| % Core Lipids, total | 36.77 ± 1.75 | 36.72 ± 1.82 | 36.82 ± 1.85 |
| % Surface Lipids, total | 63.23 ± 1.75 | 63.28 ± 1.82 | 63.18 ± 1.85 |
| FC/PLs | 0.18 ± 0.02 | 0.19 ± 0.02 * | 0.18 ± 0.02 + |
| EC/FC | 3.32 ± 0.34 | 3.18 ± 0.30 * | 3.20 ± 0.33 |
| EC/Total Cholesterol | 0.77 ± 0.02 | 0.76 ± 0.02 * | 0.76 ± 0.02 |
| TG/EC | 0.14 ± 0.04 | 0.15 ± 0.06 | 0.19 ± 0.04 ***,+++ |
| Total (n = 141) | Controls (n = 50) | MASLD 1 (n = 51) | CAD 2 (n = 40) |
| Glycerophospholipids (GPLs), total | 41.37 ± 1.55 | 39.98 ± 2.22 ** | 41.99 ± 2.14 +++ |
| Phosphatidylcholine (PC) | 34.22 ± 1.35 | 33.13 ± 2.77 * | 33.41 ± 2.17 |
| Lysophosphatidylcholine (LPC) | 2.36 ± 0.40 | 2.69 ± 0.56 ** | 2.70 ± 0.53 ** |
| Phosphatidylethanolamine (PE) | 1.24 ± 0.29 | 0.94 ± 0.26 *** | 0.99 ± 0.28 *** |
| Phosphatidylinositol (PI) | 2.13 ± 0.58 | 1.47 ± 0.43 *** | 2.50 ± 1.21 *,+++ |
| Rest of the GPLs a | 1.59 ± 1.27 | 1.78 ± 1.76 | 2.40 ± 1.74 * |
| Ether Glycerolipids (Ether GLs), total | 4.99 ± 0.84 | 5.34 ± 1.03 | 4.39 ± 0.94 **,+++ |
| Plasmalogens | 1.77 ± 0.30 | 1.71 ± 0.37 | 1.88 ± 0.48 + |
| Rest of the ether GLs b | 3.22 ± 0.82 | 3.63 ± 0.96 * | 2.51 ± 0.83 ***,+++ |
| Sphingolipids (SLs), total | 7.11 ± 0.69 | 7.82 ± 0.71 *** | 7.06 ± 0.89 ***,+++ |
| Sphingomyelin (SM) | 6.19 ± 0.97 | 6.34 ± 1.01 | 5.91 ± 0.89 + |
| Rest of the SLs c | 0.92 ± 0.72 | 1.48 ± 0.78 *** | 1.15 ± 0.59 + |
| PC/SM | 4.61 ± 0.50 | 5.01 ± 0.78 ** | 4.56 ± 0.55 ++ |
| PC/LPC | 14.87 ± 2.45 | 12.84 ± 2.76 *** | 12.87 ± 2.75 ** |
| SM/LPC | 2.67 ± 0.54 | 2.49 ± 0.84 | 2.23 ± 0.39 ** |
| Total (n = 141) | Controls (n = 50) | MASLD 1 (n = 51) | CAD 2 (n = 40) |
| SFA | 39.42 ± 3.37 | 41.07 ± 3.30 * | 45.18 ± 5.11 ***,+++ |
| UFA | 60.58 ± 3.37 | 58.93 ± 3.30 * | 54.82 ± 5.11 ***,+++ |
| MUFA | 7.98 ± 2.17 | 8.75 ± 3.04 | 9.58 ± 2.26 ** |
| PUFA | 52.60 ± 3.12 | 50.18 ± 3.29 ** | 45.24 ± 5.41 ***,+++ |
| LA | 19.57 ± 1.76 | 16.72 ± 2.07 *** | 15.69 ± 2.17 ***,+ |
| EPA + AA | 12.28 ± 1.39 | 9.85 ± 1.68 *** | 9.14 ± 0.88 ***,+ |
| DHA | 4.09 ± 0.76 | 3.33 ± 0.61 *** | 2.86 ± 0.47 ***,+++ |
| SFA/UFA | 0.66 ± 0.10 | 0.70 ± 0.10 | 0.84 ± 0.18 ***,+++ |
| SFA/PUFA | 0.76 ± 0.11 | 0.82 ± 0.11 * | 1.03 ± 0.25 ***,+++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kartsoli, S.; Kostara, C.E.; Papathanasiou, A.; Tsimihodimos, V.; Bairaktari, E.T.; Christodoulou, D.K. Altered HDL Phospholipid and Fatty Acid Profile in MASLD: A Possible Explanation for the Increased CVD Risk. Int. J. Mol. Sci. 2025, 26, 6148. https://doi.org/10.3390/ijms26136148
Kartsoli S, Kostara CE, Papathanasiou A, Tsimihodimos V, Bairaktari ET, Christodoulou DK. Altered HDL Phospholipid and Fatty Acid Profile in MASLD: A Possible Explanation for the Increased CVD Risk. International Journal of Molecular Sciences. 2025; 26(13):6148. https://doi.org/10.3390/ijms26136148
Chicago/Turabian StyleKartsoli, Sofia, Christina E. Kostara, Athanasios Papathanasiou, Vasilis Tsimihodimos, Eleni T. Bairaktari, and Dimitrios K. Christodoulou. 2025. "Altered HDL Phospholipid and Fatty Acid Profile in MASLD: A Possible Explanation for the Increased CVD Risk" International Journal of Molecular Sciences 26, no. 13: 6148. https://doi.org/10.3390/ijms26136148
APA StyleKartsoli, S., Kostara, C. E., Papathanasiou, A., Tsimihodimos, V., Bairaktari, E. T., & Christodoulou, D. K. (2025). Altered HDL Phospholipid and Fatty Acid Profile in MASLD: A Possible Explanation for the Increased CVD Risk. International Journal of Molecular Sciences, 26(13), 6148. https://doi.org/10.3390/ijms26136148