
Targeting PAD4: A Promising Strategy to Combat β-Cell Loss in Type 1 Diabetes

 ,
,
Abstract
1. Introduction
2. Results
2.1. Verifying That STZ Injured the Pancreatic Tissue, Not the Liver, at the SIXTH Week in Exp A
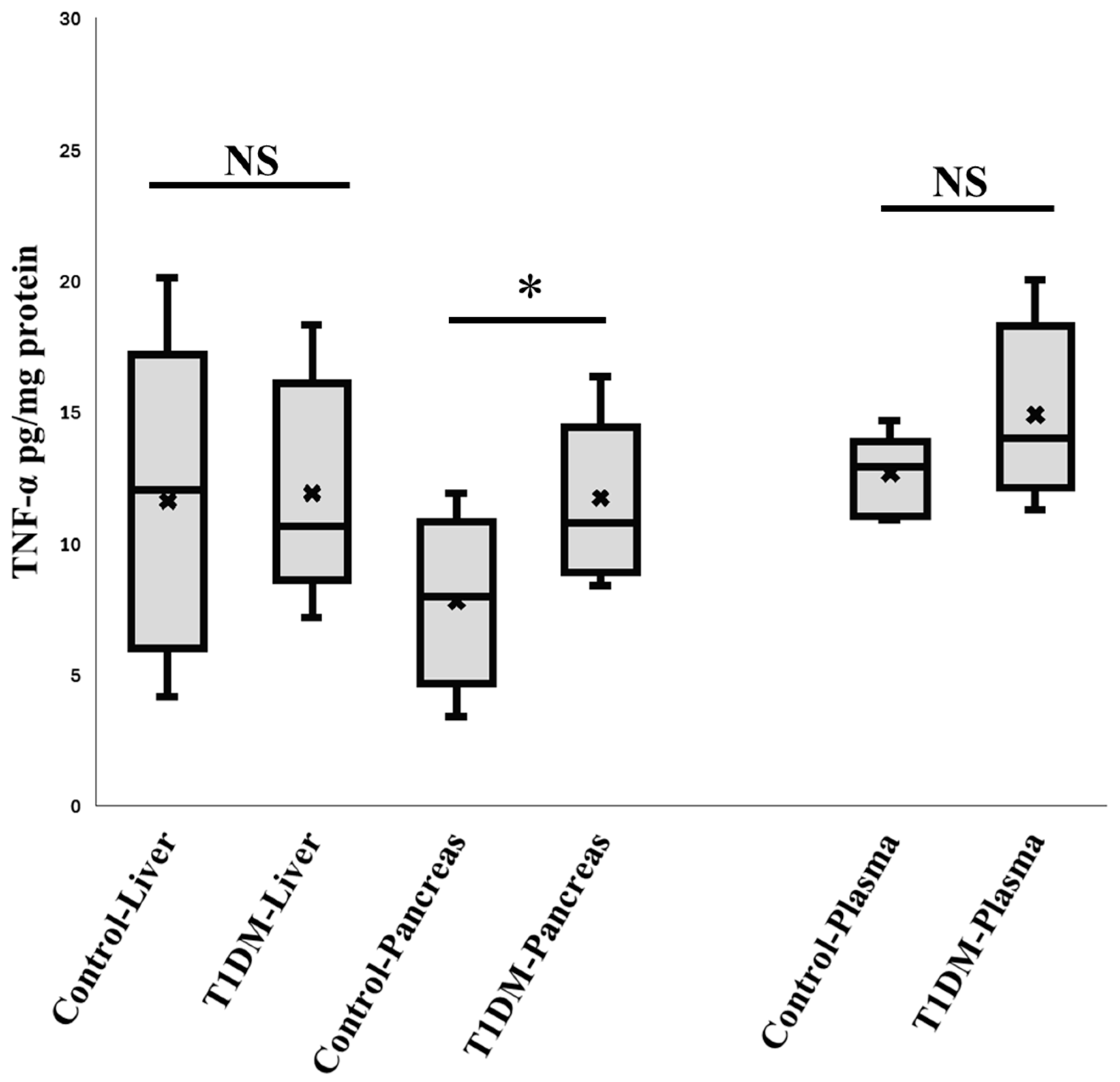
2.2. Validating the Inflammation in Pancreas During the Sixth Week in Exp A
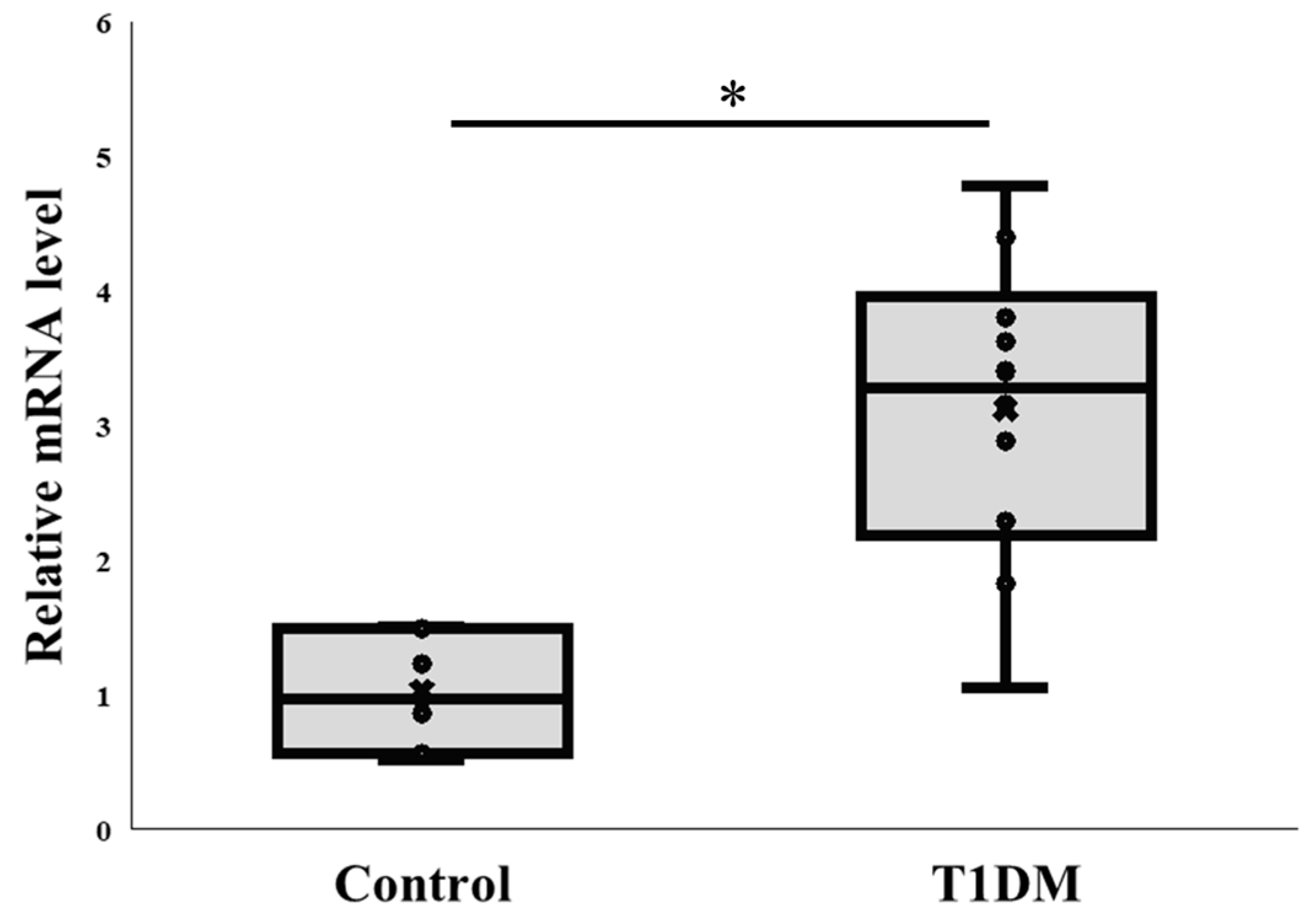
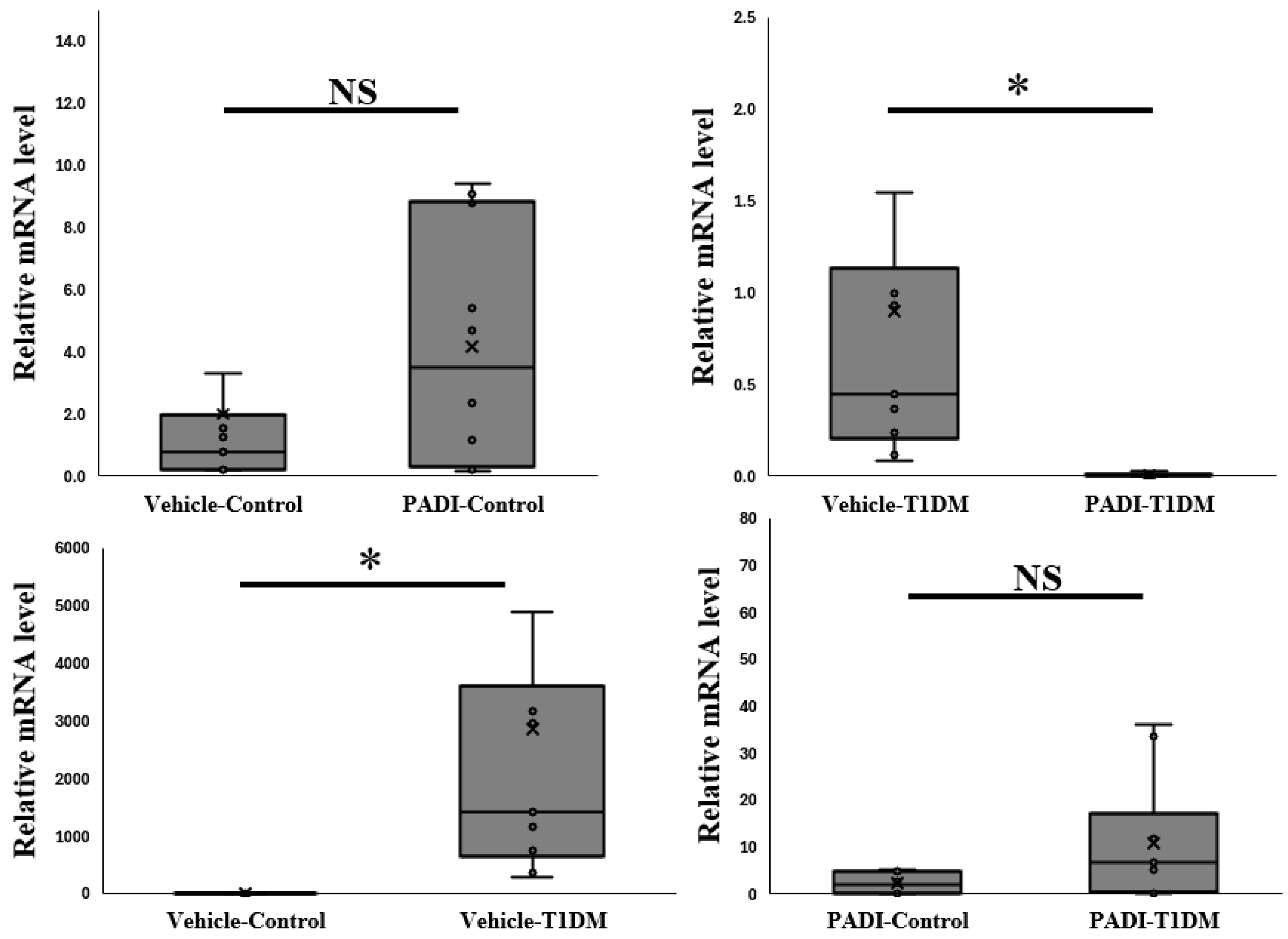
2.3. PAD4 mRNA Level in the Pancreas at the Sixth Week in Exp A
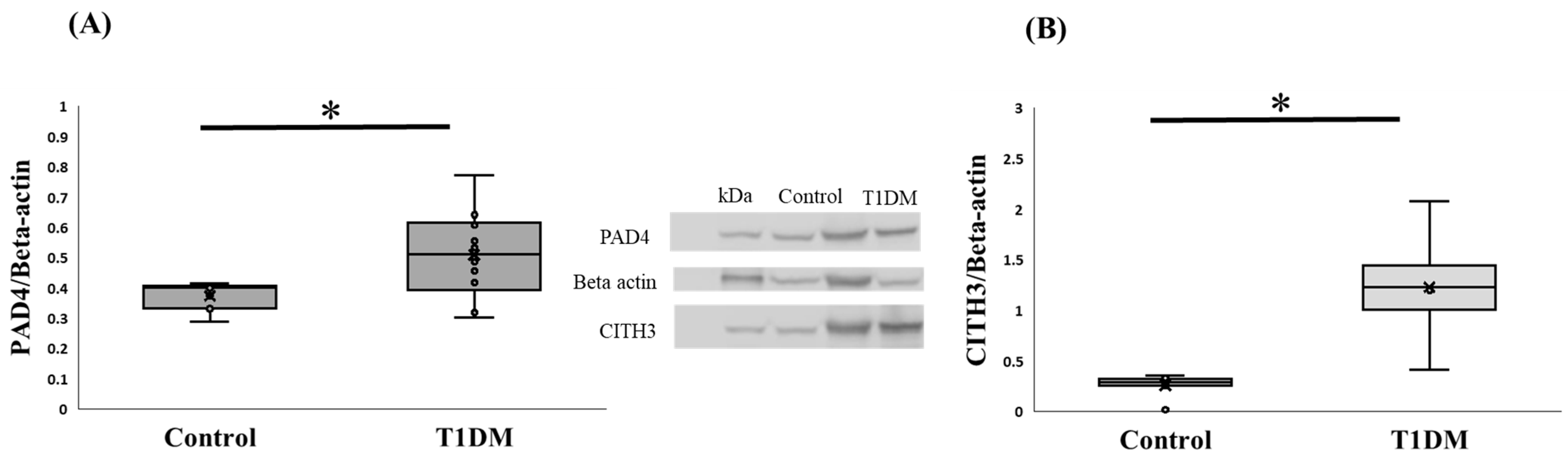
2.4. The Expression of PAD4 and Citrullinated Protein in the Pancreas at the Sixth Week in Exp A
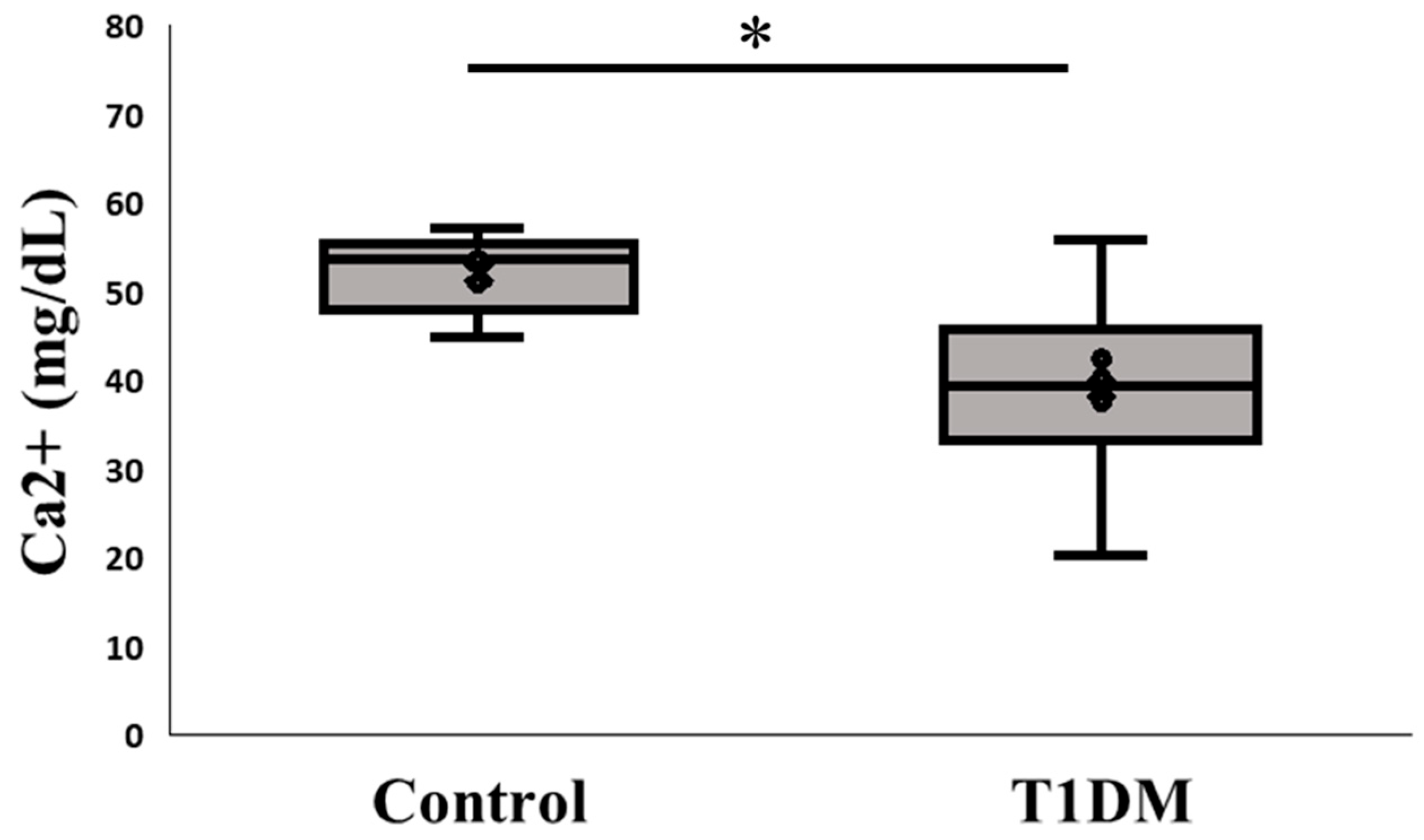
2.5. Ca2+: A Prerequisite Cofactor for Citrullination Among PADs in the Sixth Week in Exp A
2.6. Blood Glucose Measurement Before Termination in Exp A and B
2.7. The Expression of PAD4 mRNA in the Pancreas in Exp B
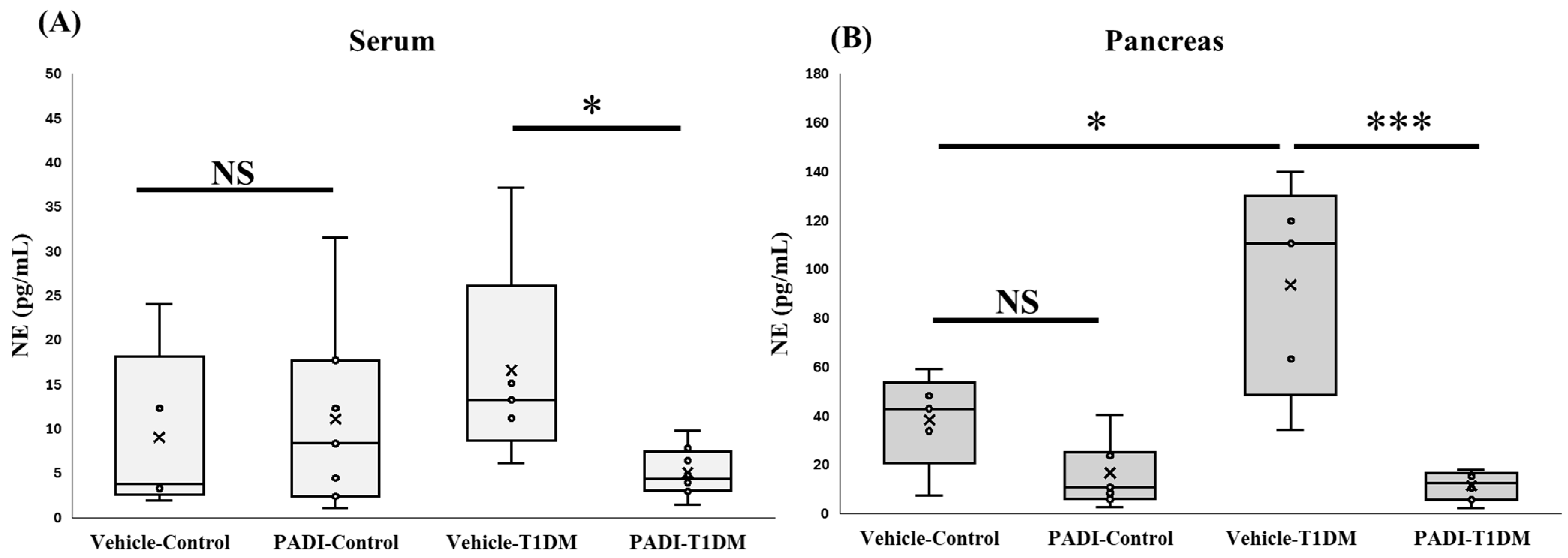
2.8. NETosis Level in the Serum and Pancreas in Exp B
3. Discussion
Limitations of This Study
4. Materials and Methods
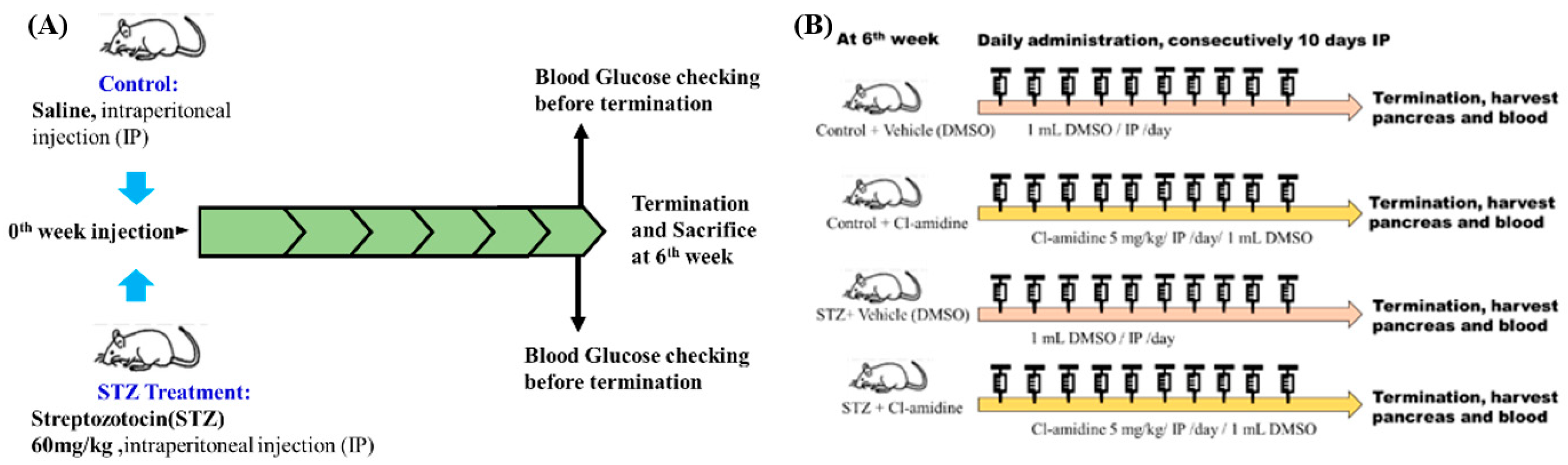
4.1. Experimental Protocol
4.2. Measurement of Calcium Ion in the Pancreas in Exp A
4.3. Western Blot of CITH3, PAD4 in Exp A
4.4. Measurement of Pancreatic TNF-α, IFN-γ, IL-6, IL-10, IL-18, NETosis, and IL-33 Concentrations in Exp A
4.5. RNA Extraction, Reverse Transcription, and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
- PAD4 sense: GCTCCCTCTCATCAGTTCCA.
- PAD4 antisense: GGCTTGTCACTCGAGTTTTGA.
- HPRT sense: CATTAATATTTAACGATGTGGATGCGTTTCA.
- HPRT antisense: GCCTACCATCTTTAAACTGCACAAT.
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- György, B.; Tóth, E.; Tarcsa, E.; Falus, A.; Buzás, E.I. Citrullination: A Posttranslational Modification in Health and Disease. Int. J. Biochem. Cell Biol. 2006, 38, 1662–1677. [Google Scholar] [CrossRef]
- Ramazi, S.; Zahiri, J. Post-Translational Modifications in Proteins: Resources, Tools and Prediction Methods. Database 2021, 2021, baab012. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, R.J.; Spindler, M.P.; Van Lummel, M.; Roep, B.O. Where, How, and When: Positioning Posttranslational Modification Within Type 1 Diabetes Pathogenesis. Curr. Diab. Rep. 2016, 16, 63. [Google Scholar] [CrossRef] [PubMed]
- Mauracher, L.-M.; Posch, F.; Martinod, K.; Grilz, E.; Däullary, T.; Hell, L.; Brostjan, C.; Zielinski, C.; Ay, C.; Wagner, D.D.; et al. Citrullinated Histone H3, a Biomarker of Neutrophil Extracellular Trap Formation, Predicts the Risk of Venous Thromboembolism in Cancer Patients. J. Thromb. Haemost. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- Guo, G.; Liu, Z.; Yu, J.; You, Y.; Li, M.; Wang, B.; Tang, J.; Han, P.; Wu, J.; Shen, H. Neutrophil Function Conversion Driven by Immune Switchpoint Regulator against Diabetes-Related Biofilm Infections. Adv. Mater. 2024, 36, e2310320. [Google Scholar] [CrossRef]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes Primes Neutrophils to Undergo NETosis, Which Impairs Wound Healing. Nat. Med. 2015, 21, 815–819. [Google Scholar] [CrossRef]
- Yang, M.-L.; Sodré, F.M.C.; Mamula, M.J.; Overbergh, L. Citrullination and PAD Enzyme Biology in Type 1 Diabetes—Regulators of Inflammation, Autoimmunity, and Pathology. Front. Immunol. 2021, 12, 678953. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; You, Q.; Wu, Y.; Wu, J. Inhibition of PAD4-Mediated NET Formation by Cl-Amidine Prevents Diabetes Development in Nonobese Diabetic Mice. Eur. J. Pharmacol. 2022, 916, 174623. [Google Scholar] [CrossRef]
- Yang, C.; Dong, Z.-Z.; Zhang, J.; Teng, D.; Luo, X.; Li, D.; Zhou, Y. Peptidylarginine Deiminases 4 as a Promising Target in Drug Discovery. Eur. J. Med. Chem. 2021, 226, 113840. [Google Scholar] [CrossRef]
- He, W.; Xi, Q.; Cui, H.; Zhang, P.; Huang, R.; Wang, T.; Wang, D. Forsythiaside B Ameliorates Coagulopathies in a Rat Model of Sepsis through Inhibition of the Formation of PAD4-Dependent Neutrophil Extracellular Traps. Front. Pharmacol. 2022, 13, 1022985. [Google Scholar] [CrossRef]
- Streptozotocin-Induced Diabetic Models in Mice and Rats-Furman-2021-Current Protocols-Wiley Online Library. Available online: https://currentprotocols.onlinelibrary.wiley.com/doi/full/10.1002/cpz1.78 (accessed on 22 March 2025).
- Zhu, Y.; Xia, X.; He, Q.; Xiao, Q.-A.; Wang, D.; Huang, M.; Zhang, X. Diabetes-Associated Neutrophil NETosis: Pathogenesis and Interventional Target of Diabetic Complications. Front. Endocrinol. 2023, 14, 1202463. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Song, W.; Jiang, Z.; Zhou, H.; Wang, S. Citrullination: A Modification Important in the Pathogenesis of Autoimmune Diseases. Clin. Immunol. 2022, 245, 109134. [Google Scholar] [CrossRef] [PubMed]
- Kraaij, T.; Tengström, F.C.; Kamerling, S.W.A.; Pusey, C.D.; Scherer, H.U.; Toes, R.E.M.; Rabelink, T.J.; van Kooten, C.; Teng, Y.K.O. A Novel Method for High-Throughput Detection and Quantification of Neutrophil Extracellular Traps Reveals ROS-Independent NET Release with Immune Complexes. Autoimmun. Rev. 2016, 15, 577–584. [Google Scholar] [CrossRef]
- Chrysanthopoulou, A.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Mikroulis, D.; Konstantinidis, T.; Sivridis, E.; Koffa, M.; Giatromanolaki, A.; Boumpas, D.T.; et al. Neutrophil Extracellular Traps Promote Differentiation and Function of Fibroblasts. J. Pathol. 2014, 233, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Öğüten, P.N.; Öztürk, S.E.; Dikmen, M. The Investigation of Cytotoxic and Apoptotic Activity of Cl-Amidine on the Human U-87 MG Glioma Cell Line. Medicine 2024, 103, e37015. [Google Scholar] [CrossRef]
- Yao, H.; Cao, G.; Liu, Z.; Zhao, Y.; Yan, Z.; Wang, S.; Wang, Y.; Guo, Z.; Wang, Y. Inhibition of Netosis with PAD Inhibitor Attenuates Endotoxin Shock Induced Systemic Inflammation. Int. J. Mol. Sci. 2022, 23, 13264. [Google Scholar] [CrossRef]
- Du, J.; Wang, N.; Sun, H.; Zheng, L.; Qi, X. Cl-Amidine Attenuates Lipopolysaccharide-Induced Inflammation in Human Gingival Fibroblasts via the JNK/MAPK, NF-κB, and Nrf2 Signalling Pathways. Hum. Cell 2023, 36, 223–233. [Google Scholar] [CrossRef]
- Long, X.; Yuan, Q.; Tian, R.; Zhang, W.; Liu, L.; Yang, M.; Yuan, X.; Deng, Z.; Li, Q.; Sun, R. Efficient Healing of Diabetic Wounds by MSC-EV-7A Composite Hydrogel via Suppression of Inflammation and Enhancement of Angiogenesis. Biomater. Sci. 2024, 12, 1750–1760. [Google Scholar] [CrossRef]
- Yang, S.; Wang, S.; Chen, L.; Wang, Z.; Chen, J.; Ni, Q.; Guo, X.; Zhang, L.; Xue, G. Neutrophil Extracellular Traps Delay Diabetic Wound Healing by Inducing Endothelial-to-Mesenchymal Transition via the Hippo Pathway. Int. J. Biol. Sci. 2023, 19, 347–361. [Google Scholar] [CrossRef]
- Altamura, S.; Lombardi, F.; Palumbo, P.; Cinque, B.; Ferri, C.; Del Pinto, R.; Pietropaoli, D. The Evolving Role of Neutrophils and Neutrophil Extracellular Traps (NETs) in Obesity and Related Diseases: Recent Insights and Advances. Int. J. Mol. Sci. 2024, 25, 13633. [Google Scholar] [CrossRef]
- Sun, B.; Chen, H.; Xue, J.; Li, P.; Fu, X. The Role of GLUT2 in Glucose Metabolism in Multiple Organs and Tissues. Mol. Biol. Rep. 2023, 50, 6963–6974. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Thompson, P.R. Protein Arginine Deiminases (PADs): Biochemistry and Chemical Biology of Protein Citrullination. Acc. Chem. Res. 2019, 52, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, B.; Mittereder, N.; Chaerkady, R.; Strain, M.; An, L.-L.; Rahman, S.; Ma, W.; Low, C.P.; Chan, D. Spontaneous Secretion of the Citrullination Enzyme PAD2 and Cell Surface Exposure of PAD4 by Neutrophils. Front. Immunol. 2017, 8, 1200. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Hamza, A.; Boyle, E.; Donu, D.; Cen, Y. Post-Translational Modifications and Diabetes. Biomolecules 2024, 14, 310. [Google Scholar] [CrossRef]
- Battaglia, M.; Petrelli, A.; Vecchio, F. Neutrophils and Type 1 Diabetes: Current Knowledge and Suggested Future Directions. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Ciesielski, O.; Biesiekierska, M.; Panthu, B.; Soszyński, M.; Pirola, L.; Balcerczyk, A. Citrullination in the Pathology of Inflammatory and Autoimmune Disorders: Recent Advances and Future Perspectives. Cell. Mol. Life Sci. 2022, 79, 94. [Google Scholar] [CrossRef]
- Aukrust, S.G.; Holte, K.B.; Opstad, T.B.; Seljeflot, I.; Berg, T.J.; Helseth, R. NETosis in Long-Term Type 1 Diabetes Mellitus and Its Link to Coronary Artery Disease. Front. Immunol. 2022, 12, 799539. [Google Scholar] [CrossRef]
- Ramadan, J.W.; Steiner, S.R.; O’Neill, C.M.; Nunemaker, C.S. The Central Role of Calcium in the Effects of Cytokines on Beta-Cell Function: Implications for Type 1 and Type 2 Diabetes. Cell Calcium 2011, 50, 481–490. [Google Scholar] [CrossRef]
- Garley, M.; Jabłońska, E.; Surażyński, A.; Grubczak, K.; Ratajczak-Wrona, W.; Iwaniuk, A.; Dąbrowska, D.; Pałka, J.A.; Moniuszko, M. Original Article Cytokine Network & NETs. Folia Biol. 2017, 63, 182–189. [Google Scholar]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of Interleukin (IL)-6-Type Cytokine Signalling and Its Regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef]
- Xiao, J.; Li, J.; Cai, L.; Chakrabarti, S.; Li, X. Cytokines and Diabetes Research. J. Diabetes Res. 2014, 2014, 920613. [Google Scholar] [CrossRef] [PubMed]
- Glowacka, E.; Banasik, M.; Lewkowicz, P.; Tchorzewski, H. The Effect of LPS on Neutrophils from Patients with High Risk of Type 1 Diabetes Mellitus in Relation to IL-8, IL-10 and IL-12 Production and Apoptosis In Vitro: Polymorphonuclear Neutrophils Apoptosis in DM1. Scand. J. Immunol. 2002, 55, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Sodré, F.M.C.; Bissenova, S.; Bruggeman, Y.; Tilvawala, R.; Cook, D.P.; Berthault, C.; Mondal, S.; Callebaut, A.; You, S.; Scharfmann, R.; et al. Peptidylarginine Deiminase Inhibition Prevents Diabetes Development in NOD Mice. Diabetes 2021, 70, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Abacar, K.; Macleod, T.; Direskeneli, H.; McGonagle, D. How Underappreciated Autoinflammatory (Innate Immunity) Mechanisms Dominate Disparate Autoimmune Disorders. Front. Immunol. 2024, 15, 1439371. [Google Scholar] [CrossRef]
- Ghasemi, A.; Jeddi, S. Streptozotocin as a Tool for Induction of Rat Models of Diabetes: A Practical Guide. Excli J. 2023, 22, 274–294. [Google Scholar] [CrossRef]
- Melo, Z.; Gutierrez-Mercado, Y.K.; Garcia-Martínez, D.; Portilla-de-Buen, E.; Canales-Aguirre, A.A.; Gonzalez-Gonzalez, R.; Franco-Acevedo, A.; Palomino, J.; Echavarria, R. Sex-Dependent Mechanisms Involved in Renal Tolerance to Ischemia-Reperfusion: Role of Inflammation and Histone H3 Citrullination. Transpl. Immunol. 2020, 63, 101331. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | CXCL-1 | IFN-γ | IL-6 | IL-18 | IL-33 | IL-10 |
|---|---|---|---|---|---|---|
| Control | 33.02 ±3.34 | 21.62 ± 2.44 | 40.81 ± 5.63 | 77.67 ± 4.83 | 708.38 ± 76.19 | 451.95 ± 152.31 * |
| STZ | 46.85 ± 3.32 * | 29.46 ± 2.00 * | 55.44 ± 3.29 * | 130.75 ± 17.46 * | 1027.26 ± 113.47 * | 153.71 ± 18.73 |
| Cytokine | inflammatory | inflammatory | inflammatory | inflammatory | inflammatory | anti-inflammatory |
| Rats (Weight: 180~200 g) | Mice (Weight: ~25 g) | Note | |
|---|---|---|---|
| Strains | Sprague Dawley, Wistar | CD-1, C57BL-6 | Male mice or rats are more susceptible to STZ than females |
| STZ-induced protocol and STZ dose | Single STZ dose: 40~70 mg/kg | 1. Low STZ dose: 40~50 mg/kg/day, 5 consecutive day 2. Single, high STZ dose: 200 mg/kg | Caution: Mice that undergo STZ-induced experiments are more likely to die (Figure 1) |
| Cl-amidine dose | In the current trial, we administered 5 mg/kg/day via IP injection for ten days in a row | Oral gavage or IP: 5~75 mg/kg/day for 30 consecutive days in accordance with other articles | Caution: rats are heavier than mice, hence more Cl-amidine will be consumed by them during the experiment Consequently, the cost is increasing |
| PAD4 expression and the capacity of the inhibition of Cl-amidine | In the current work, we confirmed that PAD4 expression and Cl-amidine inhibition are significant between the control and STZ-T1DM groups | To date, the majority of the research has used mice to treat Cl-amidine and study any disorders connected to PAD4 expression | Some high STZ-sensitive strains may treat low-dose Cl-amidine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.L.; Szász, A.; Valkusz, Z.; Várkonyi, T.; Pósa, A.; Kupai, K. Targeting PAD4: A Promising Strategy to Combat β-Cell Loss in Type 1 Diabetes. Int. J. Mol. Sci. 2025, 26, 6113. https://doi.org/10.3390/ijms26136113
Kang HL, Szász A, Valkusz Z, Várkonyi T, Pósa A, Kupai K. Targeting PAD4: A Promising Strategy to Combat β-Cell Loss in Type 1 Diabetes. International Journal of Molecular Sciences. 2025; 26(13):6113. https://doi.org/10.3390/ijms26136113
Chicago/Turabian StyleKang, Hsu Lin, András Szász, Zsuzsanna Valkusz, Tamás Várkonyi, Anikó Pósa, and Krisztina Kupai. 2025. "Targeting PAD4: A Promising Strategy to Combat β-Cell Loss in Type 1 Diabetes" International Journal of Molecular Sciences 26, no. 13: 6113. https://doi.org/10.3390/ijms26136113
APA StyleKang, H. L., Szász, A., Valkusz, Z., Várkonyi, T., Pósa, A., & Kupai, K. (2025). Targeting PAD4: A Promising Strategy to Combat β-Cell Loss in Type 1 Diabetes. International Journal of Molecular Sciences, 26(13), 6113. https://doi.org/10.3390/ijms26136113