


Osteopontin: Its Properties, Recent Studies, and Potential Applications




Abstract
1. Introduction
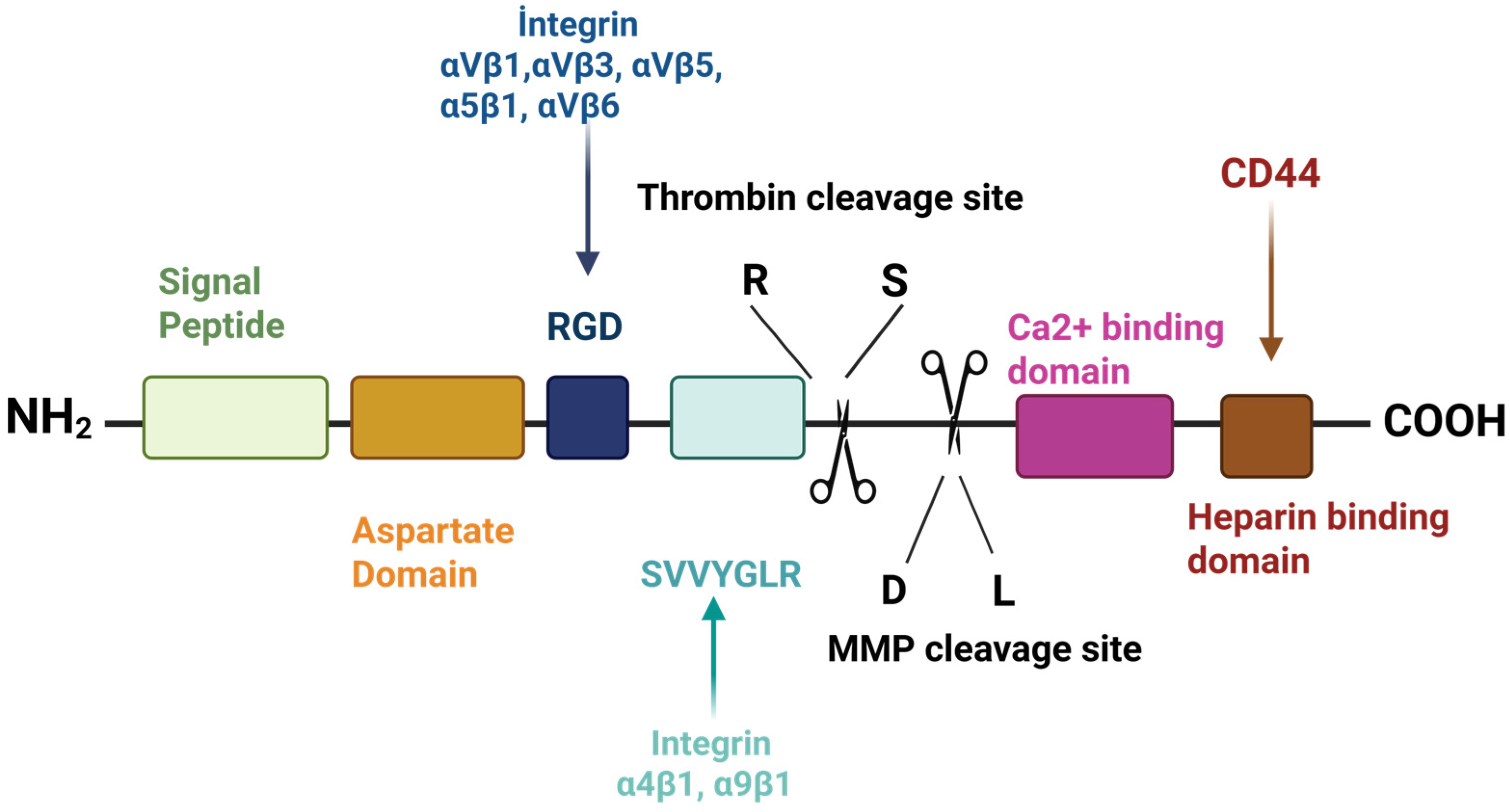
2. Structure and Chemical Properties of Osteopontin
Biosynthesis of Osteopontin
3. Effects of Environmental Conditions and Genetic Factors on OPN Levels
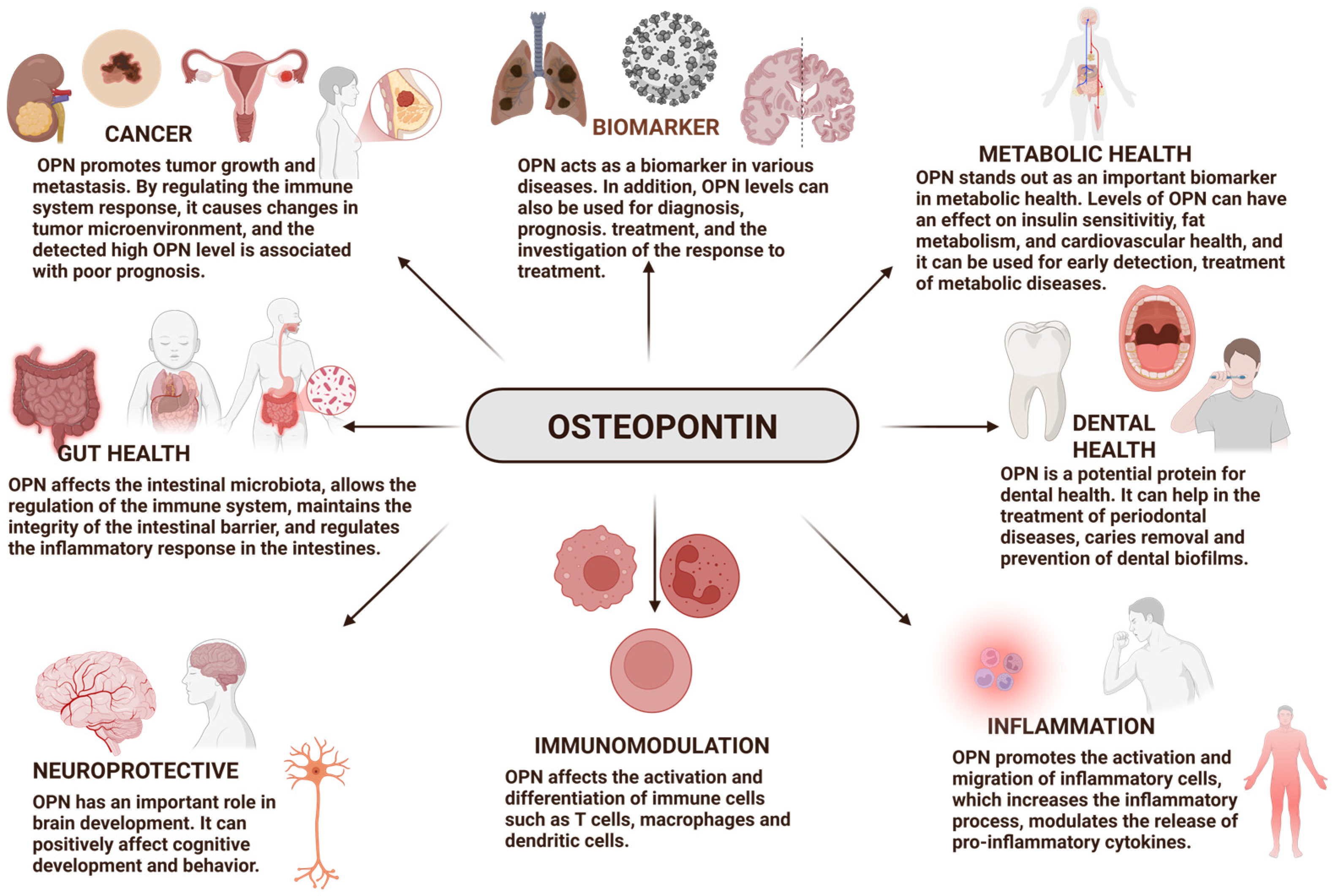
4. Biological Functions of Osteopontin
4.1. Osteopontin as a Biomarker
4.2. Osteopontin and Cancer
4.3. Osteopontin and Gut Health
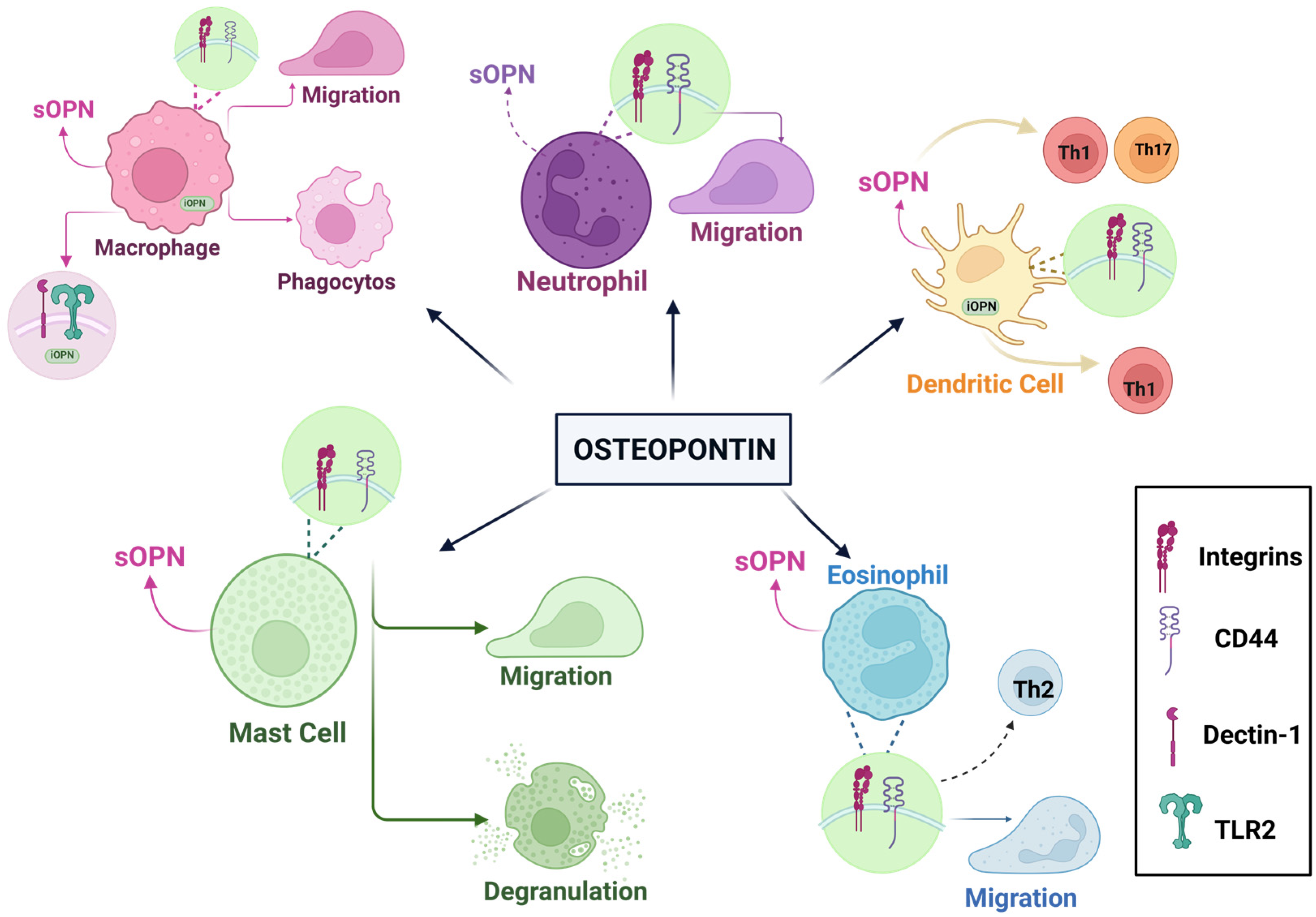
4.4. Osteopontin and Immunological Effects
4.5. Dental Health
4.6. Osteopontin and Brain Development: Cognitive Function
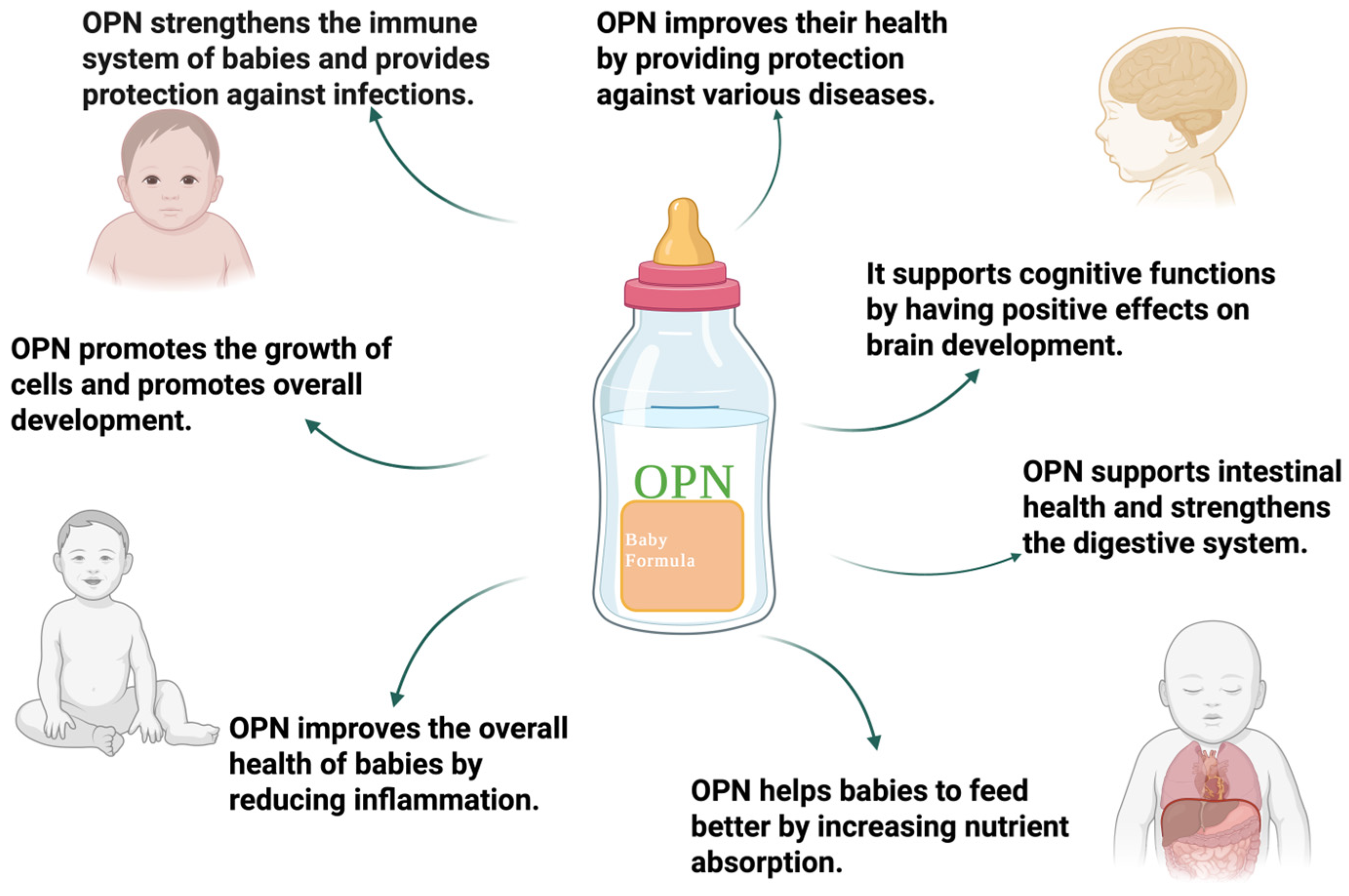
5. Nutritional Potential of Osteopontin
6. Conclusions and Future Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kläning, E.; Christensen, B.; Sørensen, E.S.; Vorup-Jensen, T.; Jensen, J.K. Osteopontin Binds Multiple Calcium Ions with High Affinity and Independently of Phosphorylation Status. Bone 2014, 66, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Senger, D.F.; Wirth, D.F.; Hynes, R. Transformed Mammalian Cells Secrete Specific Proteins and Phosphoproteins. Cell 1979, 16, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Franzitn, A.; Heinegard, D. Isolation and Characterization of Two Sialoproteins Present Only in Bone Calcified Matrix. Biochem. J. 1985, 232, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Oldberg, A.; Franzfn, A.; Heinegard, D. Cloning and Sequence Analysis of Rat Bone Sialoprotein (Osteopontin) CDNA Reveals an Arg-Gly-Asp Cell-Binding Sequence (Cell Adhesion/Bone Matrix Proteins/Glycosylation). Biochemistry 1986, 83, 8819–8823. [Google Scholar]
- Jia, Q.; Ouyang, Y.; Yang, Y.; Yao, S.; Chen, X.; Hu, Z. Osteopontin: A Novel Therapeutic Target for Respiratory Diseases. Lung 2024, 202, 25–39. [Google Scholar] [CrossRef]
- Clemente, N.; Raineri, D.; Cappellano, G.; Boggio, E.; Favero, F.; Soluri, M.F.; Dianzani, C.; Comi, C.; Dianzani, U.; Chiocchetti, A. Osteopontin Bridging Innate and Adaptive Immunity in Autoimmune Diseases. J. Immunol. Res. 2016, 2016, 7675437. [Google Scholar] [CrossRef]
- Wai, P.Y.; Kuo, P.C. The Role of Osteopontin in Tumor Metastasis. J. Surg. Res. 2004, 121, 228–241. [Google Scholar] [CrossRef]
- Shimodaira, T.; Matsuda, K.; Uchibori, T.; Sugano, M.; Uehara, T.; Honda, T. Upregulation of Osteopontin Expression via the Interaction of Macrophages and Fibroblasts under IL-1b Stimulation. Cytokine 2018, 110, 63–69. [Google Scholar] [CrossRef]
- Craig, A.M.; Denhardt, D.T. The Murine Gene Encoding Secreted Phosphoprotein 1 (Osteopontin): Promoter Structure, Activity, and Induction in Vivo by Estrogen and Progesterone (Promoter Analysis; Gene Structure; Bone Sialoprotein; Transformation-Associated Phosphoprotein; Early T-Lymphocyte Activation 1; Tumor Promoter Induction). Gene 1991, 100, 163–171. [Google Scholar]
- Attur, M.G.; Dave, M.N.; Stuchin, S.; Kowalski, A.J.; Steiner, G.; Abramson, S.B.; Denhardt, D.T.; Amin, A.R. Osteopontin: An Intrinsic Inhibitor of Inflammation in Cartilage. Arthritis Rheum. 2001, 44, 578–584. [Google Scholar] [CrossRef]
- Hwang, S.-M.; Wilson, P.D.; Laskin, J.D.; Denmardt, D.T. Age and Development-Related Changes in Osteopontin and Nitric Oxide Synthase MRNA Levels in Human Kidney Proximal Tubule Epithelial Cells: Contrasting Responses to Hypoxia and Reoxygenation. J. Cell Physiol. 1994, 160, 6148. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.M.; Lopez, C.A.; Heck, D.E.; Gardner, C.R.; Laskin, D.L.; Laskin, J.D.; Denhardt, D.T. Osteopontin Inhibits Induction of Nitric Oxide Synthase Gene Expression by Inflammatory Mediators in Mouse Kidney Epithelial Cells. J. Biol. Chem. 1994, 269, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.A.; Lynn Weir, M.; Wilson, S.M.; Xuan, J.W.; Chambers, A.F.; Mccormack, D.G.; Burton, A.C.; Weir, M.L. Osteopontin Inhibits Inducible Nitric Oxide Synthase Activity in Rat Vascular Tissue. Am. J. Physiol.-Heart Circ. Physiol. 1998, 275, H2258–H2265. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.; Guo, X. Osteopontin: A Protein with Diverse Functions. FASEB J. 1993, 7, 1475–1482. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M. Osteopontin Expression and Function: Role in Bone Remodeling. J. Cell Biochem. 1998, 72, 92–102. [Google Scholar] [CrossRef]
- O’Regan, A.; Berman, J.S. Osteopontin: A Key Cytokine in Cell-Mediated and Granulomatous Inflammation. Int. J. Exp. Pathol. 2000, 81, 373–390. [Google Scholar] [CrossRef]
- Weber, G.F. The Metastasis Gene Osteopontin: A Candidate Target for Cancer Therapy. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2001, 1552, 61–85. [Google Scholar] [CrossRef]
- Tenen, D.G.; Senger, D.R.; Perruzzi, C.A.; Gracey, C.F.; Papadopoulos, A. Secreted Phosphoproteins Associated with Neoplastic Transformation: Close Homology with Plasma Proteins Cleaved during Blood Coagulation. Cancer Res. 1988, 48, 5770–5774. [Google Scholar]
- Senger, D.R.; Perruzzi, C.A.; Papadopoulos, A.; Tenen, D.G. Purification of a Human Milk Protein Closely Similar to Tumor-Secreted Phosphoproteins and Osteopontin. Biochim. Biophys. Acta (BBA)/Protein Struct. Mol. 1989, 996, 43–48. [Google Scholar] [CrossRef]
- Bautista, D.S.; Denstedt, J.; Chambers, A.F.; Harris, J.F. Low-Molecular-Weight Variants of Osteopontin Generated by Serine Proteinases in Urine of Patients with Kidney Stones. J. Cell Biochem. 1996, 61, 402–409. [Google Scholar] [CrossRef]
- Moorman, H.R.; Poschel, D.; Klement, J.D.; Lu, C.; Redd, P.S.; Liu, K. Osteopontin: A Key Regulator of Tumor Progression and Immunomodulation. Cancers 2020, 12, 3379. [Google Scholar] [CrossRef] [PubMed]
- Toyonaga, T.; Nakase, H.; Ueno, S.; Matsuura, M.; Yoshino, T.; Honzawa, Y.; Itou, A.; Namba, K.; Minami, N.; Yamada, S.; et al. Osteopontin Deficiency Accelerates Spontaneous Colitis in Mice with Disrupted Gut Microbiota and Macrophage Phagocytic Activity. PLoS ONE 2015, 10, e0135552. [Google Scholar] [CrossRef] [PubMed]
- Chunder, R.; Schropp, V.; Marzin, M.; Amor, S.; Kuerten, S. A Dual Role of Osteopontin in Modifying B Cell Responses. Biomedicines 2023, 11, 1969. [Google Scholar] [CrossRef] [PubMed]
- Schack, L.; Lange, A.; Kelsen, J.; Agnholt, J.; Christensen, B.; Petersen, T.E.; Sørensen, E.S. Considerable Variation in the Concentration of Osteopontin in Human Milk, Bovine Milk, and Infant Formulas. J. Dairy Sci. 2009, 92, 5378–5385. [Google Scholar] [CrossRef]
- West, C.E.; Kvistgaard, A.S.; Peerson, J.M.; Donovan, S.M.; Peng, Y.M.; Lönnerdal, B. Effects of Osteopontin-Enriched Formula on Lymphocyte Subsets in the First 6 Months of Life: A Randomized Controlled Trial. Pediatr. Res. 2017, 82, 63–71. [Google Scholar] [CrossRef]
- Huang, Y.; Lu, Z.; Liu, F.; Lane, J.; Chen, J.; Fu, X.; Huang, Q.; Hu, R.; Zhang, B. Osteopontin Enhances the Probiotic Viability of Bifidobacteria in Pectin-Based Microencapsulation Subjected to in Vitro Infant Gastrointestinal Digestion. Food Hydrocoll. 2024, 149, 109634. [Google Scholar] [CrossRef]
- Si, J.; Wang, C.; Zhang, D.; Wang, B.; Hou, W.; Zhou, Y. Osteopontin in Bone Metabolism and Bone Diseases. Med. Sci. Monit. 2020, 26, e919159-1. [Google Scholar] [CrossRef]
- Kristensen, M.F.; Sørensen, E.S.; Del Rey, Y.C.; Schlafer, S. Prevention of Initial Bacterial Attachment by Osteopontin and Other Bioactive Milk Proteins. Biomedicines 2022, 10, 1922. [Google Scholar] [CrossRef]
- Nurrohman, H.; Carter, L.; Barnes, N.; Zehra, S.; Singh, V.; Tao, J.; Marshall, S.J.; Marshall, G.W. The Role of Process-Directing Agents on Enamel Lesion Remineralization: Fluoride Boosters. Biomimetics 2022, 7, 54. [Google Scholar] [CrossRef]
- Burling, H.; Sørensen, E.S.; Bertelsen, H.; Jørgensen, A.S.; Graverholt, G. Use of Osteopontin in Dental Formulations. WO 2005/053628 A1; World Intellectual Property Organization: Geneva, Switzerland, 2005. Available online: https://patentscope.wipo.int/search/en/WO2005053628 (accessed on 15 June 2025).
- Joung, S.; Fil, J.E.; Heckmann, A.B.; Kvistgaard, A.S.; Dilger, R.N. Early-Life Supplementation of Bovine Milk Osteopontin Supports Neurodevelopment and Influences Exploratory Behavior. Nutrients 2020, 12, 2206. [Google Scholar] [CrossRef]
- Chen, W.; Ma, Q.; Suzuki, H.; Hartman, R.; Tang, J.; Zhang, J.H. Osteopontin Reduced Hypoxia-Ischemia Neonatal Brain Injury by Suppression of Apoptosis in a Rat Pup Model. Stroke 2011, 42, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Lu, F.; Li, P.; Jian, Y.; Xu, J.; Zhong, T.; Guo, Q.; Yang, Y. Osteopontin Promotes Angiogenesis in the Spinal Cord and Exerts a Protective Role Against Motor Function Impairment and Neuropathic Pain After Spinal Cord Injury. Spine 2024, 49, E142–E151. [Google Scholar] [CrossRef] [PubMed]
- Iturbe-Fernández, D.; Pulito-Cueto, V.; Mora-Cuesta, V.M.; Remuzgo-Martínez, S.; Ferrer-Pargada, D.J.; Genre, F.; Alonso-Lecue, P.; López-Mejías, R.; Atienza-Mateo, B.; González-Gay, M.A.; et al. Osteopontin as a Biomarker in Interstitial Lung Diseases. Biomedicines 2024, 12, 1108. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Lönnerdal, B. Evaluation of Bioactivities of Bovine Milk Osteopontin Using a Knockout Mouse Model. J. Pediatr. Gastroenterol. Nutr. 2020, 71, 125–131. [Google Scholar] [CrossRef]
- Othman, O.A.; Qasem, A.; Taha, G.; Amer, H. Evaluation of Osteopontin as a Potential Biomarker in Hepatocellular Carcinoma in a Sample of Egyptian Patients. Egypt. J. Cancer Biomed. Res. 2024, 8, 47–53. [Google Scholar] [CrossRef]
- Kadoglou, N.P.E.; Khattab, E.; Velidakis, N.; Gkougkoudi, E. The Role of Osteopontin in Atherosclerosis and Its Clinical Manifestations (Atherosclerotic Cardiovascular Diseases)—A Narrative Review. Biomedicines 2023, 11, 3178. [Google Scholar] [CrossRef]
- Günaydın, S.; Özlü, T.; Özsu, S.S.; Örem, A. Role of Osteopontin and NGAL in Differential Diagnosis of Acute Exacerbations of COPD and Pneumonia. Med. Sci. Discov. 2024, 11, 150–156. [Google Scholar] [CrossRef]
- Pappas, A.G.; Eleftheriou, K.; Vlahakos, V.; Magkouta, S.F.; Riba, T.; Dede, K.; Siampani, R.; Kompogiorgas, S.; Polydora, E.; Papalampidou, A.; et al. High Plasma Osteopontin Levels Are Associated with Serious Post-Acute-COVID-19-Related Dyspnea. J. Clin. Med. 2024, 13, 392. [Google Scholar] [CrossRef]
- Quesnel, M.J.; Labonté, A.; Picard, C.; Bowie, D.C.; Zetterberg, H.; Blennow, K.; Brinkmalm, A.; Villeneuve, S.; Poirier, J. Osteopontin: A Novel Marker of Pre-Symptomatic Sporadic Alzheimer’s Disease. Alzheimer’s Dement. 2024, 20, 6008–6031. [Google Scholar] [CrossRef]
- Ardizzone, A.; Siracusa, R.; Capra, A.P.; Di Paola, R.; Esposito, E.; Cuzzocrea, S.; D’Amico, R.; Impellizzeri, D. Evaluating Osteopontin as a Biomarker of Obesity Related Complications before and after Metabolic and Bariatric Surgery: A Systematic Review and Meta-Analysis. Obes. Res. Clin. Pract. 2025, 19, 19–27. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The Multiple Functions and Mechanisms of Osteopontin. Clin Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.T. The Nature and Significance of Osteopontin. Connect. Tissue Res. 1989, 23, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Bayless, K.J.; Davis, G.E. Identification of Dual A4β1 Integrin Binding Sites within a 38 Amino Acid Domain in the N-Terminal Thrombin Fragment of Human Osteopontin. J. Biol. Chem. 2001, 276, 13483–13489. [Google Scholar] [CrossRef]
- Christensen, B.; Petersen, T.E.; Sørensen, E.S. Post-Translational Modification and Proteolytic Processing of Urinary Osteopontin. Biochem. J. 2008, 411, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.; Kazanecki, C.C.; Petersen, T.E.; Rittling, S.R.; Denhardt, D.T.; Sørensen, E.S. Cell Type-Specific Post-Translational Modifications of Mouse Osteopontin Are Associated with Different Adhesive Properties. J. Biol. Chem. 2007, 282, 19463–19472. [Google Scholar] [CrossRef]
- Katayama, Y.; House, C.M.; Udagawa, N.; Kazama, J.J.; McFarland, R.J.; Martin, T.J.; Findlay, D.M. Casein Kinase 2 Phosphorylation of Recombinant Rat Osteopontin Enhances Adhesion of Osteoclasts but Not Osteoblasts. J. Cell Physiol. 1998, 176, 179–187. [Google Scholar] [CrossRef]
- Ek-Rylander, B.; Flores, M.; Wendel, M.; Heinegård, D.; Andersson, G. Dephosphorylation of Osteopontin and Bone Sialoprotein by Osteoclastic Tartrate-Resistant Acid Phosphatase: Modulation of Osteoclast Adhesion in Vitro. J. Biol. Chem. 1994, 269, 14853–14856. [Google Scholar] [CrossRef]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (Osteopontin): An Early Component of Type-1 (Cell-Mediated) Immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef]
- Al-Shami, R.; Sorensen, E.S.; Ek-Rylander, B.; Andersson, G.; Carson, D.D.; Farach-Carson, M.C. Phosphorylated Osteopontin Promotes Migration of Human Choriocarcinoma Cells via a P70 S6 Kinase-Dependent Pathway. J. Cell Biochem. 2005, 94, 1218–1233. [Google Scholar] [CrossRef]
- Boskey, A.L.; Maresca, M.; Ullrich, W.; Doty, S.B.; Butler, W.T.; Prince, C.W. Osteopontin-Hydroxyapatite Interactions in Vitro: Inhibition of Hydroxyapatite Formation and Growth in a Gelatin-Gel. Bone Miner. 1993, 22, 147–159. [Google Scholar] [CrossRef]
- Hunter, G.K.; Kyle, C.L.; Goldberg, H.A. Modulation of Crystal Formation by Bone Phosphoproteins: Structural Specificity of the Osteopontin-Mediated Inhibition of Hydroxyapatite Formation. Biochem. J. 1994, 300, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Kazanecki, C.C. Osteopontin and Cell Adhesion: Role of Post-Translational Modifications and The C-Terminal Region. Ph.D. Thesis, Rutgers, The State University of New Jersey, Rutgers University Community Repository, New Brunswick, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Tan, Y.; Zhao, L.; Yang, Y.G.; Liu, W. The Role of Osteopontin in Tumor Progression Through Tumor-Associated Macrophages. Front. Oncol. 2022, 12, 953283. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, E.S.; Christensen, B. Milk Osteopontin and Human Health. Nutrients 2023, 15, 2423. [Google Scholar] [CrossRef] [PubMed]
- Standal, T.; Borset, M.; Sundan, A. Role of Osteopontin in Adhesion, Migration, Cell Survival and Bone Remodeling. Exp. Oncol. 2004, 26, 179–184. [Google Scholar]
- Yokosaki, Y.; Matsuura, N.; Sasaki, T.; Murakami, I.; Schneider, H.; Higashiyama, S.; Saitoh, Y.; Yamakido, M.; Taooka, Y.; Sheppard, D. The Integrin A9β1 Binds to a Novel Recognition Sequence (SVVYGLR) in the Thrombin-Cleaved Amino-Terminal Fragment of Osteopontin. J. Biol. Chem. 1999, 274, 36328–36334. [Google Scholar] [CrossRef]
- Christensen, B.; Nielsen, M.S.; Haselmann, K.F.; Petersen, T.E.; Sørensen, E.S. Post-Translationally Modified Residues of Native Human Osteopontin Are Located in Clusters: Identification of 36 Phosphorylation and Five O-Glycosylation Sites and Their Biological Implications. Biochem. J. 2005, 390, 285–292. [Google Scholar] [CrossRef]
- Tagliabracci, V.S.; Wiley, S.E.; Guo, X.; Kinch, L.N.; Durrant, E.; Wen, J.; Xiao, J.; Cui, J.; Nguyen, K.B.; Engel, J.L.; et al. A Single Kinase Generates the Majority of the Secreted Phosphoproteome. Cell 2015, 161, 1619–1632. [Google Scholar] [CrossRef]
- Ishikawa, H.O.; Xu, A.; Ogura, E.; Manning, G.; Irvine, K.D. The Raine Syndrome Protein FAM20C Is a Golgi Kinase That Phosphorylates Bio-Mineralization Proteins. PLoS ONE 2012, 7, e42988. [Google Scholar] [CrossRef]
- Kon, S.; Ikesue, M.; Kimura, C.; Aoki, M.; Nakayama, Y.; Saito, Y.; Kurotaki, D.; Diao, H.; Matsui, Y.; Segawa, T.; et al. Syndecan-4 Protects against Osteopontin-Mediated Acute Hepatic Injury by Masking Functional Domains of Osteopontin. J. Exp. Med. 2008, 205, 25–33. [Google Scholar] [CrossRef]
- Kazanecki, C.C.; Uzwiak, D.J.; Denhardt, D.T. Control of Osteopontin Signaling and Function by Post-Translational Phosphorylation and Protein Folding. J. Cell Biochem. 2007, 102, 912–924. [Google Scholar] [CrossRef]
- Lin, E.Y.H.; Xi, W.; Aggarwal, N.; Shinohara, M.L. Osteopontin (OPN)/SPP1: From Its Biochemistry to Biological Functions in the Innate Immune System and the Central Nervous System (CNS). Int. Immunol. 2023, 35, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.L.; Kim, H.-J.; Kim, J.-H.; Garcia, V.A.; Cantor, H. Alternative Translation of Osteopontin Generates Intracellular and Secreted Isoforms That Mediate Distinct Biological Activities in Dendritic Cells. Proc. Natl. Acad. Sci. USA 2008, 105, 7235–7239. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Li, Y.; Yao, R.; Jiang, M. Osteopontin in Chronic Inflammatory Diseases: Mechanisms, Biomarker Potential, and Therapeutic Strategies. Biology 2025, 14, 428. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.; Karlsen, N.J.; Jørgensen, S.D.S.; Jacobsen, L.N.; Ostenfeld, M.S.; Petersen, S.V.; Müllertz, A.; Sørensen, E.S. Milk Osteopontin Retains Integrin-Binding Activity after in Vitro Gastrointestinal Transit. J. Dairy Sci. 2020, 103, 42–51. [Google Scholar] [CrossRef]
- Schytte, G.N.; Christensen, B.; Bregenov, I.; Kjøge, K.; Scavenius, C.; Petersen, S.V.; Enghild, J.J.; Sørensen, E.S. FAM20C Phosphorylation of the RGDSVVYGLR Motif in Osteopontin Inhibits Interaction with the Avβ3 Integrin. J. Cell Biochem. 2020, 121, 4809–4818. [Google Scholar] [CrossRef]
- WHO. Evidence for the Ten Steps to Successful Breastfeeding; WHO: Geneva, Switzerland, 1998. [Google Scholar]
- WHO. Promoting and Supporting Breast-Feeding: The Special Role of Maternity Services. A Joint WHO/UNICEF Statement; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Aksan, A.; Erdal, I.; Yalcin, S.S.; Stein, J.; Samur, G. Osteopontin Levels in Human Milk Are Related to Maternal Nutrition and Infant Health and Growth. Nutrients 2021, 13, 2670. [Google Scholar] [CrossRef]
- Arslan, A.; Kaplan, M.; Duman, H.; Bayraktar, A.; Ertürk, M.; Henrick, B.M.; Frese, S.A.; Karav, S. Bovine Colostrum and Its Potential for Human Health and Nutrition. Front. Nutr. 2021, 8, 651721. [Google Scholar] [CrossRef]
- Sarıtaş, S.; Portocarrero, A.C.M.; Miranda López, J.M.; Lombardo, M.; Koch, W.; Raposo, A.; El-Seedi, H.R.; de Brito Alves, J.L.; Esatbeyoglu, T.; Karav, S.; et al. The Impact of Fermentation on the Antioxidant Activity of Food Products. Molecules 2024, 29, 3941. [Google Scholar] [CrossRef]
- Karav, S.; Casaburi, G.; Frese, S.A. Reduced Colonic Mucin Degradation in Breastfed Infants Colonized by Bifidobacterium longum subsp. Infantis EVC001. FEBS Open Bio 2018, 8, 1649–1657. [Google Scholar] [CrossRef]
- Bolat, E.; Karagöz, Z.; Alves, J.L.d.B.; Neto, J.P.R.C.; Witkowska, A.M.; El-Seedi, H.; Lombardo, M.; Karav, S. The Potential Applications of Natural Colostrum in Skin Health. Cosmetics 2024, 11, 197. [Google Scholar] [CrossRef]
- Bezirtzoglou, E.; Tsiotsias, A.; Welling, G.W. Microbiota Profile in Feces of Breast- and Formula-Fed Newborns by Using Fluorescence in Situ Hybridization (FISH). Anaerobe 2011, 17, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Another Reason to Favor Exclusive Breastfeeding: Microbiome Resilience. J. Pediatr. 2018, 94, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.Å.; Korotkova, M. The Role of Breastfeeding in Prevention of Neonatal Infection. Semin. Neonatol. 2002, 7, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Park, H.; Ha, E.; Hong, Y.C.; Ha, M.; Park, H.; Kim, B.N.; Lee, B.; Lee, S.J.; Lee, K.Y.; et al. Effect of Breastfeeding Duration on Cognitive Development in Infants: 3-Year Follow-up Study. J. Korean Med. Sci. 2016, 31, 579–584. [Google Scholar] [CrossRef]
- Demmelmair, H.; Prell, C.; Timby, N.; Lönnerdal, B. Benefits of Lactoferrin, Osteopontin and Milk Fat Globule Membranes for Infants. Nutrients 2017, 9, 817. [Google Scholar] [CrossRef]
- Ruan, H.; Tang, Q.; Zhao, X.; Zhang, Y.; Zhao, X.; Xiang, Y.; Geng, W.; Feng, Y.; Cai, W. The Levels of Osteopontin in Human Milk of Chinese Mothers and Its Associations with Maternal Body Composition. Food Sci. Hum. Wellness 2022, 11, 1419–1427. [Google Scholar] [CrossRef]
- Jiang, R.; Lönnerdal, B. Osteopontin in Human Milk and Infant Formula Affects Infant Plasma Osteopontin Concentrations. Pediatr. Res. 2019, 85, 502–505. [Google Scholar] [CrossRef]
- Goonatilleke, E.; Huang, J.; Xu, G.; Wu, L.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Human Milk Proteins and Their Glycosylation Exhibit Quantitative Dynamic Variations during Lactation. J. Nutr. 2019, 149, 1317–1325. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, A.; Lai, S.; Yuan, Q.; Jia, X.; Wang, P.; Zhang, Y. Longitudinal Changes in the Concentration of Major Human Milk Proteins in the First Six Months of Lactation and Their Effects on Infant Growth. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, X.; Wang, Y.; Bai, S.; Lai, J.; Tong, X.; Xing, Y. Longitudinal Changes of Lactopontin (Milk Osteopontin) in Term and Preterm Human Milk. Front. Nutr. 2022, 9, 962802. [Google Scholar] [CrossRef]
- Joung, K.E.; Christou, H.; Park, K.H.; Mantzoros, C.S. Cord Blood Levels of Osteopontin as a Phenotype Marker of Gestational Age and Neonatal Morbidities. Obesity 2014, 22, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Ueno, H.M.; Yamaide, F.; Nakano, T.; Shiko, Y.; Kawasaki, Y.; Mitsuishi, C.; Shimojo, N. Comparison of 30 Cytokines in Human Breast Milk between 1989 and 2013 in Japan. Nutrients 2023, 15, 1735. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolomeo, M.; Pietrantonio, F.; Pellegrinelli, A.; Martinetti, A.; Mariani, L.; Daidone, M.G.; Bajetta, E.; Pelosi, G.; de Braud, F.; Floriani, I.; et al. Osteopontin, E-Cadherin, and β-Catenin Expression as Prognostic Biomarkers in Patients with Radically Resected Gastric Cancer. Gastric Cancer 2016, 19, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Tsuda, H.; Bandera, C.A.; Nishimura, S.; Inoue, T.; Kawamura, N.; Berkowitz, R.S.; Mok, S.C. Comparison of Osteopontin Expression in Endometrioid Endometrial Cancer and Ovarian Endometrioid Cancer. Med. Oncol. 2006, 23, 205–212. [Google Scholar] [CrossRef]
- Kundu, G.; Elangovan, S. Investigating the Role of Osteopontin (OPN) in the Progression of Breast, Prostate, Renal and Skin Cancers. Biomedicines 2025, 13, 173. [Google Scholar] [CrossRef]
- Mack, P.C.; Redman, M.W.; Chansky, K.; Williamson, S.K.; Farneth, N.C.; Lara, P.N.; Franklin, W.A.; Le, Q.T.; Crowley, J.J.; Gandara, D.R. Lower Osteopontin Plasma Levels Are Associated with Superior Outcomes in Advanced Non-Small-Cell Lung Cancer Patients Receiving Platinum-Based Chemotherapy: SWOG Study S0003. J. Clin. Oncol. 2008, 26. [Google Scholar] [CrossRef]
- Das, S.; Song, Z.; Han, H.; Ge, X.; Desert, R.; Athavale, D.; Babu Komakula, S.S.; Magdaleno, F.; Chen, W.; Lantvit, D.; et al. Intestinal Osteopontin Protects from Alcohol-Induced Liver Injury by Preserving the Gut Microbiome and the Intestinal Barrier Function. CMGH 2022, 14, 813–839. [Google Scholar] [CrossRef]
- Aasmul-Olsen, K.; Henriksen, N.L.; Nguyen, D.N.; Heckmann, A.B.; Thymann, T.; Sangild, P.T.; Bering, S.B. Milk Osteopontin for Gut, Immunity and Brain Development in Preterm Pigs. Nutrients 2021, 13, 2675. [Google Scholar] [CrossRef]
- Han, L.; Li, Q.; Du, M.; Mao, X. Bovine Milk Osteopontin Improved Intestinal Health of Pregnant Rats Fed a High-Fat Diet through Improving Bile Acid Metabolism. J. Dairy Sci. 2024, 107, 24–39. [Google Scholar] [CrossRef]
- Ferreres-Serafini, L.; Martín-Orúe, S.M.; Sadurní, M.; Jiménez, J.; Moreno-Muñoz, J.A.; Castillejos, L. Supplementing Infant Milk Formula with a Multi-Strain Synbiotic and Osteopontin Enhances Colonic Microbial Colonization and Modifies Jejunal Gene Expression in Lactating Piglets. Food Funct. 2024, 15, 6536–6552. [Google Scholar] [CrossRef]
- Donovan, S.M.; Monaco, M.H.; Drnevich, J.; Kvistgaard, A.S.; Hernell, O.; Lönnerdal, B. Bovine Osteopontin Modifies the Intestinal Transcriptome of Formula-Fed Infant Rhesus Monkeys to Be More Similar to Those That Were Breastfed. J. Nutr. 2014, 144, 1910–1919. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zeng, P.; Gong, L.; Zhang, X.; Ling, Z.; Bi, K.; Shi, F.; Wang, K.; Zhang, Q.; Jiang, J.; et al. Osteopontin Exacerbates High-Fat Diet-Induced Metabolic Disorders in a Microbiome-Dependent Manner. mBio 2022, 13, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Layton, G.R.; Antoun, I.; Copperwheat, A.; Khan, Z.L.; Bhandari, S.S.; Somani, R.; Ng, A.; Zakkar, M. Osteopontin as a Biomarker for Coronary Artery Disease. Cells 2025, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a Means to Cope with Environmental Insults: Regulation of Inflammation, Tissue Remodeling, and Cell Survival. J. Clin. Investig. 2001, 107, 1055–1061. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Furger, K.A.; Menon, R.K.; Tuck, A.B.; Bramwell, V.H.; Chambers, A.F. The Functional and Clinical Roles of Osteopontin in Cancer and Metastasis. Curr. Mol. Med. 2001, 1, 621–632. [Google Scholar] [CrossRef]
- Alpana Kumari, D.K.V.K.G. Osteopontin in Cancer. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2024; Volume 118, pp. 87–110. [Google Scholar]
- Tilli, T.M.; Thuler, L.C.; Matos, A.R.; Coutinho-Camillo, C.M.; Soares, F.A.; da Silva, E.A.; Neves, A.F.; Goulart, L.R.; Gimba, E.R. Expression Analysis of Osteopontin MRNA Splice Variants in Prostate Cancer and Benign Prostatic Hyperplasia. Exp. Mol. Pathol. 2012, 92, 13–19. [Google Scholar] [CrossRef]
- Gassler, N.; Autschbach, F.; Gauer, S.; Bohn, J.; Sido, B.; Otto, H.F.; Geiger, H.; Obermüller, N. Expression of Osteopontin (Eta-1) in Crohn Disease of the Terminal Ileum. Scand. J. Gastroenterol. 2002, 37, 1286–1295. [Google Scholar] [CrossRef]
- Agnholt, J.; Kelsen, J.; Schack, L.; Hvas, C.L.; Dahlerup, J.F.; Sørensen, E.S. Osteopontin, a Protein with Cytokine-like Properties, Is Associated with Inflammation in Crohn’s Disease. Scand. J. Immunol. 2007, 65, 453–460. [Google Scholar] [CrossRef]
- Da Silva, A.P.B.; Pollett, A.; Rittling, S.R.; Denhardt, D.T.; Sodek, J.; Zohar, R. Exacerbated Tissue Destruction in DSS-Induced Acute Colitis of OPN-Null Mice Is Associated with Downregulation of TNF-α Expression and Non-Programmed Cell Death. J. Cell Physiol. 2006, 208, 629–639. [Google Scholar] [CrossRef]
- Patarca, R.; Freeman, G.J.; Singh, R.P.; Wei, F.Y.; Durfee, T.; Blattner, F.; Regnier, D.C.; Kozak, C.A.; Mock, B.A.; Morse, H.C.; et al. Structural and Functional Studies of the Early T Lymphocyte Activation 1 (Eta-1) Gene. Definition of a Novel T Cell-Dependent Response Associated with Genetic Resistance to Bacterial Infection. J. Exp. Med. 1989, 170, 145–161. [Google Scholar] [CrossRef] [PubMed]
- Puxeddu, I.; Berkman, N.; Ribatti, D.; Bader, R.; Haitchi, H.M.; Davies, D.E.; Howarth, P.H.; Levi-Schaffer, F. Osteopontin Is Expressed and Functional in Human Eosinophils. Allergy Eur. J. Allergy Clin. Immunol. 2010, 65, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Macneil, R.L.; Berry, J.; D’errico, J.; Strayhorn, C.; Piotrowski, B.; Somerman, M.J. Role of Two Mineral-Associated Adhesion Molecules, Osteopontin and Bone Sialoprotein, during Cementogenesis. Connect. Tissue Res. 1995, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- McKee, M.D.; Nanci, A. Osteopontin at Mineralized Tissue Interfaces in Bone, Teeth, and Osseointegrated Implants: Ultrastructural Distribution and Implications for Mineralized Tissue Formation, Turnover, and Repair. Microsc. Res. Tech. 1996, 33, 141–164. [Google Scholar] [CrossRef]
- Duman, H.; Kaplan, M.; Arslan, A.; Sahutoglu, A.S.; Kayili, H.M.; Frese, S.A.; Karav, S. Potential Applications of Endo-β-N-Acetylglucosaminidases from Bifidobacterium longum Subspecies Infantis in Designing Value-Added, Next-Generation Infant Formulas. Front. Nutr. 2021, 8, 646275. [Google Scholar] [CrossRef]
- Yu, H.; Liu, X.; Zhong, Y. The Effect of Osteopontin on Microglia. Biomed. Res. Int. 2017, 2017, 1879437. [Google Scholar] [CrossRef]
- Stampanoni Bassi, M.; Buttari, F.; Gilio, L.; Iezzi, E.; Galifi, G.; Carbone, F.; Micillo, T.; Dolcetti, E.; Azzolini, F.; Bruno, A.; et al. Osteopontin Is Associated with Multiple Sclerosis Relapses. Biomedicines 2023, 11, 178. [Google Scholar] [CrossRef]
- Chiocchetti, A.; Comi, C.; Indelicato, M.; Castelli, L.; Mesturini, R.; Bensi, T.; Mazzarino, M.C.; Giordano, M.; D’Alfonso, S.; Momigliano-Richiardi, P.; et al. Osteopontin Gene Haplotypes Correlate with Multiple Sclerosis Development and Progression. J. Neuroimmunol. 2005, 163, 172–178. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, M.; Dai, W.; Guo, W.; Qiu, J.; Zhang, Z.; Mo, M.; Ding, L.; Ye, P.; Wu, Y.; et al. Bone-Derived Factors as Potential Biomarkers for Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 634213. [Google Scholar] [CrossRef]
- Wirths, O.; Breyhan, H.; Marcello, A.; Cotel, M.C.; Brück, W.; Bayer, T.A. Inflammatory Changes Are Tightly Associated with Neurodegeneration in the Brain and Spinal Cord of the APP/PS1KI Mouse Model of Alzheimer’s Disease. Neurobiol. Aging 2010, 31, 747–757. [Google Scholar] [CrossRef]
- Gehrmann, J.; Matsumoto, Y.; Kreutzberg, G.W. Full-Length Review Microglia: Intrinsic Immuneffector Cell of the Brain. Brain Res. Rev. 1995, 20, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Luo, X.; Zhang, J.; Liu, Y.; Luo, H.; Huang, Q.; Cheng, Y.; Xie, Z. Osteopontin as a Potential Therapeutic Target for Ischemic Stroke. Curr. Drug Deliv. 2017, 14, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Prell, C.; Lönnerdal, B. Milk Osteopontin Promotes Brain Development by Up-Regulating Osteopontin in the Brain in Early Life. FASEB J. 2019, 33, 1681–1694. [Google Scholar] [CrossRef] [PubMed]
- Eker, F.; Akdaşçi, E.; Duman, H.; Yalçıntaş, Y.M.; Canbolat, A.A.; Kalkan, A.E.; Karav, S.; Šamec, D. Antimicrobial Properties of Colostrum and Milk. Antibiotics 2024, 13, 251. [Google Scholar] [CrossRef]
- Karav, S.; Le Parc, A.; Leite Nobrega de Moura Bell, J.M.; Frese, S.A.; Kirmiz, N.; Block, D.E.; Barile, D.; Mills, D.A. Oligosaccharides Released from Milk Glycoproteins Are Selective Growth Substrates for Infant-Associated Bifidobacteria. Appl. Environ. Microbiol. 2016, 82, 3622–3630. [Google Scholar] [CrossRef]
- Kaplan, M.; Şahutoğlu, A.S.; Sarıtaş, S.; Duman, H.; Arslan, A.; Pekdemir, B.; Karav, S. Role of Milk Glycome in Prevention, Treatment, and Recovery of COVID-19. Front. Nutr. 2022, 9, 1033779. [Google Scholar] [CrossRef]
- Kaplan, M.; Baydemir, B.; Günar, B.B.; Arslan, A.; Duman, H.; Karav, S. Benefits of A2 Milk for Sports Nutrition, Health and Performance. Front. Nutr. 2022, 9, 935344. [Google Scholar] [CrossRef]
- Duman, H.; Bechelany, M.; Karav, S. Human Milk Oligosaccharides: Decoding Their Structural Variability, Health Benefits, and the Evolution of Infant Nutrition. Nutrients 2025, 17, 118. [Google Scholar] [CrossRef]
- Parc, A.L.; Karav, S.; Rouquié, C.; Maga, E.A.; Bunyatratchata, A.; Barile, D. Characterization of Recombinant Human Lactoferrin N-Glycans Expressed in the Milk of Transgenic Cows. PLoS ONE 2017, 12, e0171477. [Google Scholar] [CrossRef]
- Mondragon Portocarrero, A.d.C.; Lopez-Santamarina, A.; Lopez, P.R.; Ortega, I.S.I.; Duman, H.; Karav, S.; Miranda, J.M. Substitutive Effects of Milk vs. Vegetable Milk on the Human Gut Microbiota and Implications for Human Health. Nutrients 2024, 16, 3108. [Google Scholar] [CrossRef]
- Karav, S.; Salcedo, J.; Frese, S.A.; Barile, D. Thoroughbred Mare’s Milk Exhibits a Unique and Diverse Free Oligosaccharide Profile. FEBS Open Bio 2018, 8, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Kutluay Özhan, H.; Duman, H.; Bechelany, M.; Karav, S. Lactoperoxidase: Properties, Functions, and Potential Applications. Int. J. Mol. Sci. 2025, 26, 5055. [Google Scholar] [CrossRef] [PubMed]
- Mosca, F.; Giannì, M.L. Human Milk: Composition and Health Benefits. Pediatr. Medica Chir. 2017, 39, 155. [Google Scholar] [CrossRef]
- Dewey, K.G.; Heinig, M.J.; Nommsen-Rivers, L.A. Differences in Morbidity between Breast-Fed and Formula-Fed Infants. J. Pediatr. 1995, 126, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Chatterton, D.E.W.; Rasmussen, J.T.; Heegaard, C.W.; Sørensen, E.S.; Petersen, T.E. In Vitro Digestion of Novel Milk Protein Ingredients for Use in Infant Formulas: Research on Biological Functions. Trends Food Sci. Technol. 2004, 15, 373–383. [Google Scholar] [CrossRef]
- Ge, X.; Lu, Y.; Leung, T.-M.; Sørensen, E.S.; Nieto, N. Milk Osteopontin, a Nutritional Approach to Prevent Alcohol-Induced Liver Injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, 929–939. [Google Scholar] [CrossRef]
- Kvistgaard, A.S.; Matulka, R.A.; Dolan, L.C.; Ramanujam, K.S. Pre-Clinical in Vitro and in Vivo Safety Evaluation of Bovine Whey Derived Osteopontin, Lacprodan® OPN-10. Food Chem. Toxicol. 2014, 73, 59–70. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Dose | Effect | Study Design | Ref |
|---|---|---|---|---|
| Neuroprotective | The OPN-supplemented formula was designed to use 250 mg/L OPN. | BmOPN supplementation enhanced the relative volume of various brain regions and changed behaviors in the novel object recognition task. | In vivo | [31] |
| Neuroprotective | OPN (0.03 μg or 0.1 μg) was administered intracerebroventricularly at 1 h post-HI. | Exogenous OPN reduced infarct volume and augmented neurological outcomes 7 weeks after hypoxic-ischemic (HI) injury, while an integrin antagonist inhibited OPN-induced neuroprotective. | In vivo | [32] |
| Neuroprotective | - | The possible advantages of an OPN increase in improving locomotor function and alleviating neuropathic pain subsequent to a spinal cord injury (SCI). | In vivo | [33] |
| Gut Health | - | OPN deficiency worsened alcohol-related disease (ALD). Increasing OPN in intestinal epithelial cells may help maintain the intestinal microbiome, protect barrier function, and combat ALD. | In vivo | [91] |
| Gut, Immunity, and Brain Development | Sixteen premature pigs were fed diets with 46 mg/(kg·g) of OPN added. | Pigs fed OPN supplementation showed increased villus-crypt depth, higher monocytes and lymphocytes, similar cognitive development, and improved intestinal structure and immunity, according to T-maze test results. | In vivo | [92] |
| Gut Health | BmOPN was enhanced at a dose of 6 mg/kg body weight. | OPN supplements in pregnant women prevented colon inflammation and enhanced intestinal barrier function. They regulated intestinal population through short-chain fatty acid (SCFA) production and supported bile acid secretion. | In vivo | [93] |
| Gut Health and Immune Function | In total, 0.436 g/L OPN was added to the milk formulation. | OPN improved digestion, microbiota, and the immune system in piglets. Synbiotic tablets enhanced benefical bacteria and reduced harmful genera, lowering diarrhea. SCFA increased and reduced ammonia, preventing pathogen growth. | In vivo | [94] |
| Gut Health | Formula fed supplemented with 25 mg/L bovine OPN. | Newborn rhesus monkeys fed breast milk or OPN-supplemented bovine milk showed similar growth and gene expression, suggesting OPN may affect intestinal development. | In vivo | [95] |
| Gut Health and Brain Development | In total, 12 µg/g OPN was used as a supplement every morning. | BmOPN enhanced small intestine growth, inhibited TNF-α secretion, increased brain myelination, and supported cognitive development. | In vivo | [35] |
| Biomarker | - | OPN was associated with atherosclerotic cardiovascular diseases (ASCVD) and vascular calcifications (VC), had varying effects on diseases, and may be a biomarker for coronary artery disease (CAD). | In vitro and in vivo | [37] |
| Biomarker | - | OPN levels increased post-MBS, suggesting they may indicate inflammation and bone health in bariatric patients, and suggesting OPN-targeting treatments could address these issues. | In vivo | [41] |
| Biomarker | - | The study highlights the significance of examining OPN levels in COPD and pneumonia, suggesting that OPN may have potential biomarker properties. | In vivo | [38] |
| Biomarker | - | COVID-19 patients with increased OPN levels in their circulation and severe symptoms have been suggested as potential biomarkers. | In vitro | [39] |
| Biomarker | - | Elevated CSF OPN predicted faster AD progression and was increased in a frontal cortex of AD brains. | In vivo | [40] |
| Biomarker | - | Interstitial lung disorders (ILDs) patients had higher OPN levels than healthy individuals. Elevated OPN was linked to lower vital capacity and higher mortality or lung transplant rates, indicating the potential for identifying ILD patients. | In vivo | [34] |
| Biomarker | - | In Egypt, OPN was studied as a biomarker for diagnosing hepatocellular carcinoma (HCC) in HCV patients. It proved more effective and sensitive than other markers, indicating its potential for early diagnosis. | In vivo | [36] |
| Dental Health | - | OPN was effective as process-directing agents utilized in the polymerinduced liquid-precursor (PILP) process to stabilize and convey mineral ions, and the regulated remineralization had the potential to boost the performance of fluoride. | In vitro | [29] |
| Dental Health | Previous studies used 46 µM OPN, but in this study, 50 µM was used due to the unobserved optimal effect. | Research indicates that milk proteins, particularly OPN, effectively inhibited bacterial adhesion to saliva-coated surfaces, potentially enhancing oral health and preventing dental biofilm. | In vitro | [28] |
| Dental Health | The study determined the optimal OPN dosage for use, ranging from 100 mg/kg to 1000 mg/kg, with the most suitable amount being approximately 350 mg/kg. | It has been found that OPN could bind to hydroxyapatite surfaces, as well as prevent bacteria from adhering to the tooth surface. | In vitro | [30] |
| Immunomodulation | - | Colitis progressed faster in OPN/Interleukin-10 (IL-10) double knockout (DKO) mice than in IL-10 knockout (KO) mice, indicating that OPN deficiency worsens colitis. OPN expression was higher in IL-10 KO epithelial cells. In OPN/IL-10 DKO mice, Clostridium subset XIVa decreased, while cluster XVIII increased. | In vivo | [22] |
| Immunomodulation | In total, 2 µg/well (1 µg/mL) recombinant OPN was added to B cells. | OPN reduced B cell clusters in MS brain tissue, increasing neuroinflammation and IL-6 and autoantibodies. Human recombinant OPN down-regulated IL-6 and up-regulated IL-10 in B cells. | In vitro | [23] |
| Immunomodulation | - | A 3-month-old baby’s umbilical cord OPN plasma was 7–10 times higher than normal, indicating low OPN concentration in infants’ intestinal immune systems, limiting cytokine expression. | In vitro | [24] |
| Immunomodulation | Milk formulas with 65 mg OPN/L or 130 mg OPN/L added were used. | Infants fed 130 mg of OPN/L showed increased T cell and monocyte levels, potentially promoting immunity development and progression. | In vivo | [25] |
| Cancer | - | High OPN levels can lead to cancer, promote tumor growth, and spread, with gastric cancer patients’ survival predicted by OPN levels. | In vitro | [87] |
| Cancer | - | In particular, the OPN levels of EEC (endometrioid endometrial cancer) and OEC (ovarian endometrioid cancer) cancer types were similar to each other. It was found that the OPN level was higher in advanced tumors. | In vitro | [88] |
| Cancer | - | OPN expression was high in breast, prostate, kidney, and skin cancers, with even higher levels in advanced kidney and skin cancers. Treatment targeting these cancers resulted in decreased OPN levels. | In vitro | [89] |
| Cancer | - | OPN expression in non-small cell lung cancer (NSCLC) patients was linked to survival, disease development, and treatment responses, with strong OPN expression resulting in shorter lifespans. | In vivo | [90] |
| Metabolic Health | - | OPN played a crucial role in high-fat-diet-induced lipid deposition, regulating intestinal lipid and fatty acid metabolism, and potentially preventing dyslipidemia by altering gut flora. | In vitro and in vivo | [96] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karasalih, B.; Duman, H.; Bechelany, M.; Karav, S. Osteopontin: Its Properties, Recent Studies, and Potential Applications. Int. J. Mol. Sci. 2025, 26, 5868. https://doi.org/10.3390/ijms26125868
Karasalih B, Duman H, Bechelany M, Karav S. Osteopontin: Its Properties, Recent Studies, and Potential Applications. International Journal of Molecular Sciences. 2025; 26(12):5868. https://doi.org/10.3390/ijms26125868
Chicago/Turabian StyleKarasalih, Büşra, Hatice Duman, Mikhael Bechelany, and Sercan Karav. 2025. "Osteopontin: Its Properties, Recent Studies, and Potential Applications" International Journal of Molecular Sciences 26, no. 12: 5868. https://doi.org/10.3390/ijms26125868
APA StyleKarasalih, B., Duman, H., Bechelany, M., & Karav, S. (2025). Osteopontin: Its Properties, Recent Studies, and Potential Applications. International Journal of Molecular Sciences, 26(12), 5868. https://doi.org/10.3390/ijms26125868