
Effects of Essential Oil Inhalation on the Enhancement of Plasma and Liver Lipid Metabolism in Mice

 ,
,
Abstract
1. Introduction
2. Results
2.1. Effects of Essential Oil Inhalation on Mouse (C57BL/6J) Food Intake, Body Weight, and Fat Mass
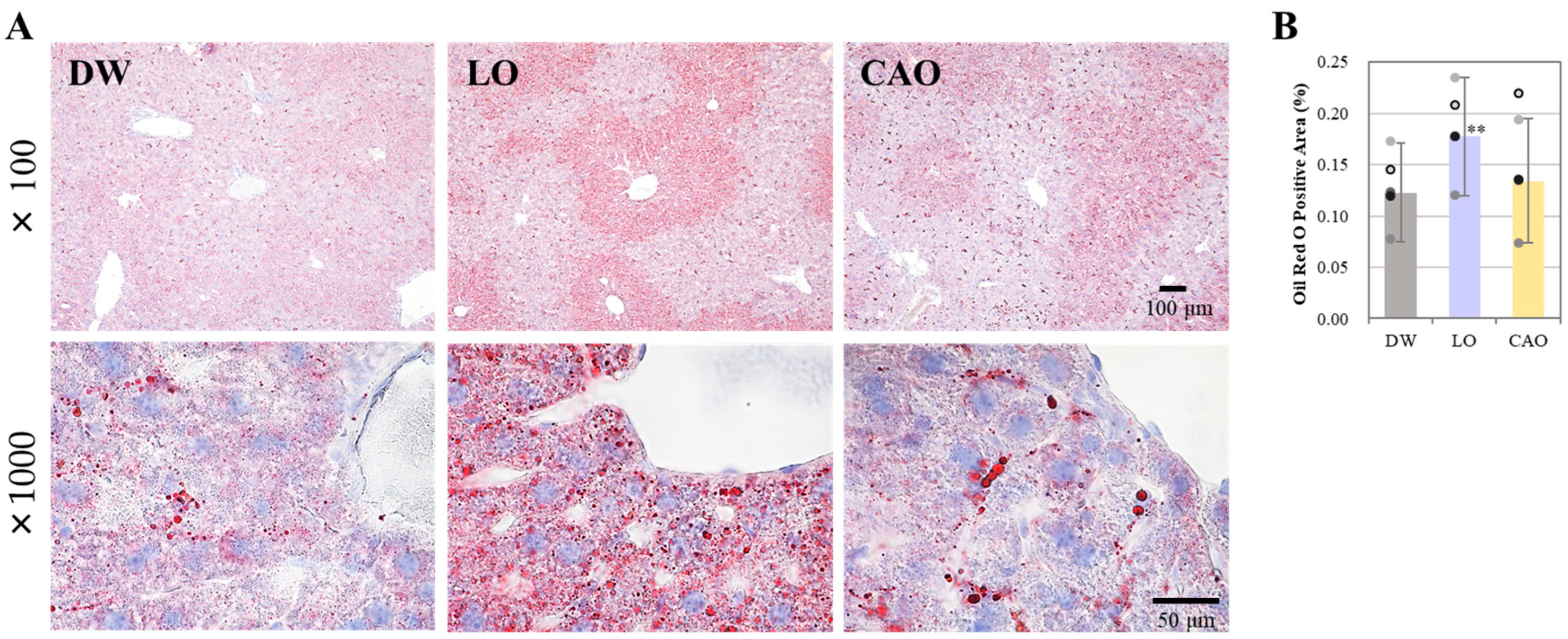
2.2. Effect of Essential Oil Inhalation on Liver Lipid Accumulation in Mice
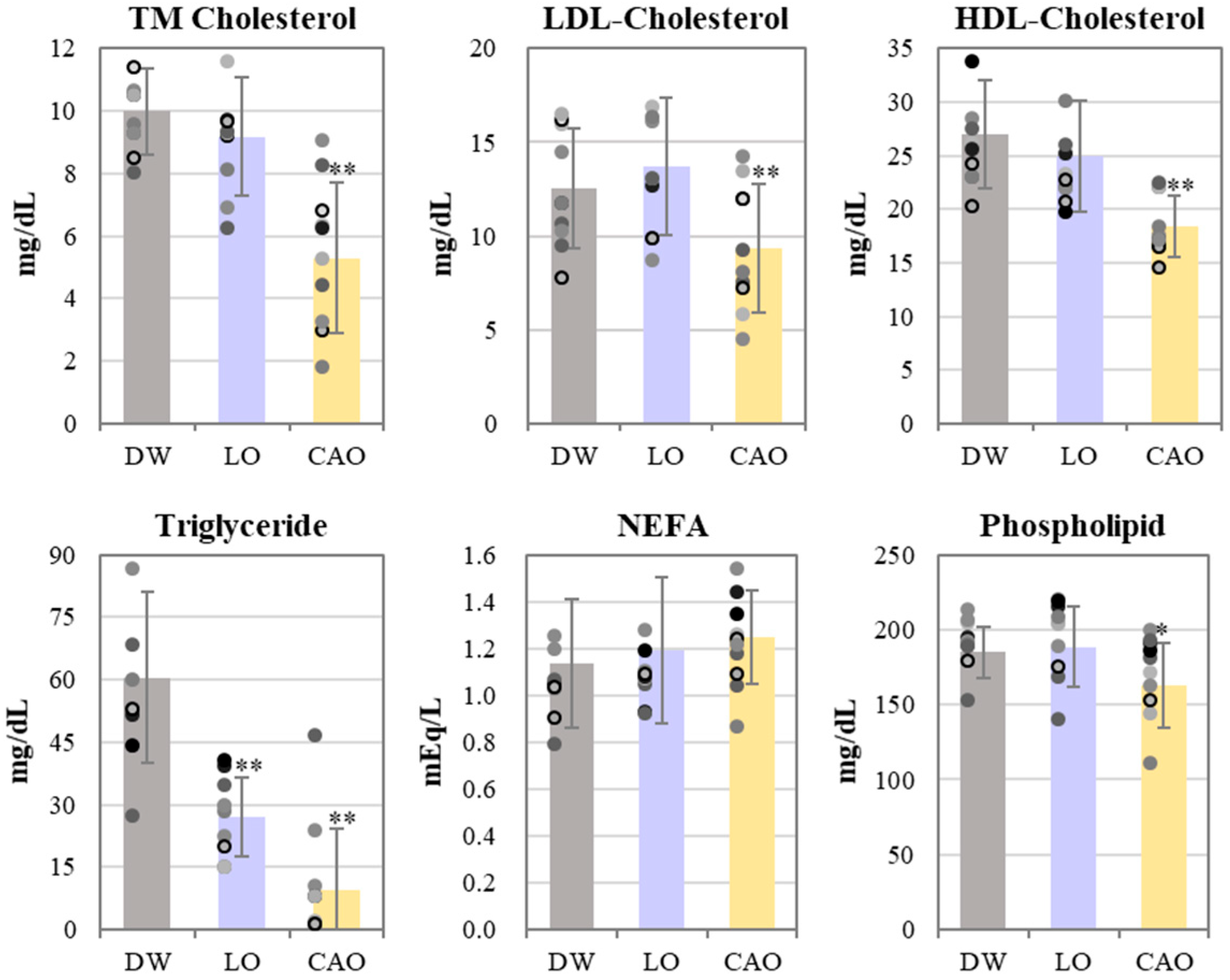
2.3. Effect of Essential Oil Inhalation on Blood Lipid Profiles of Mice
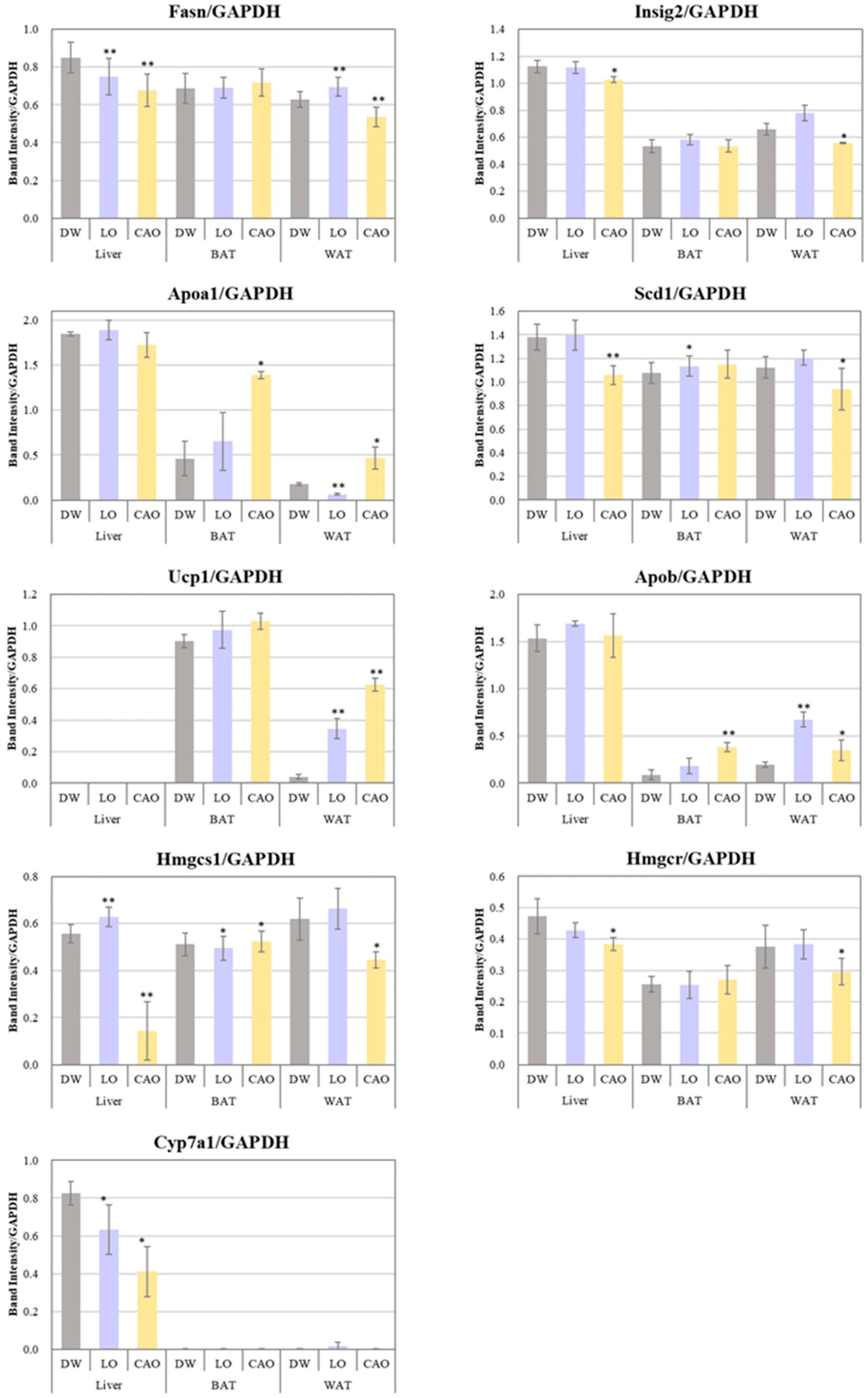
2.4. Effects of Essential Oil Inhalation on Genes Associated with Adipogenesis
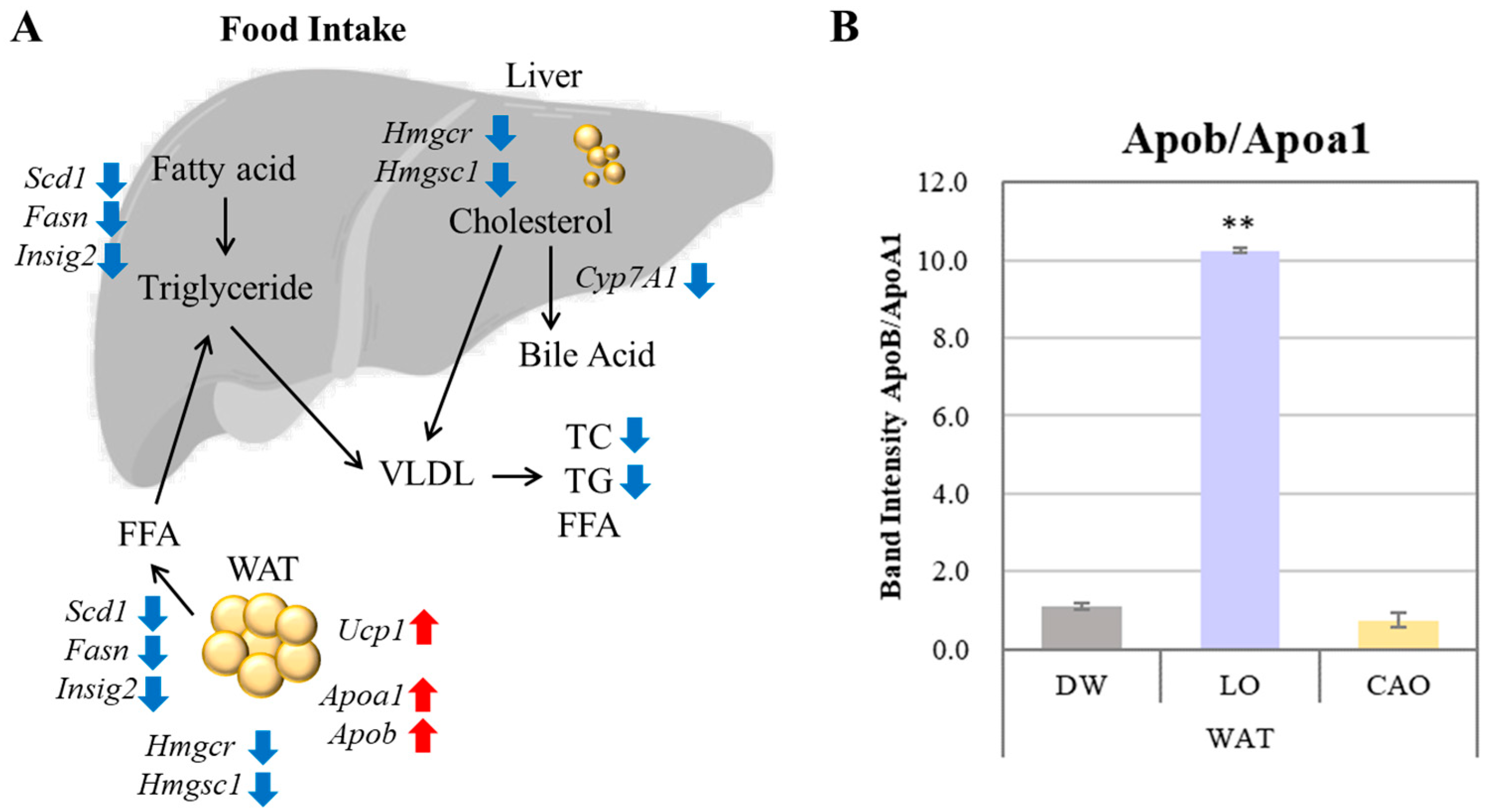
3. Discussion
4. Materials and Methods
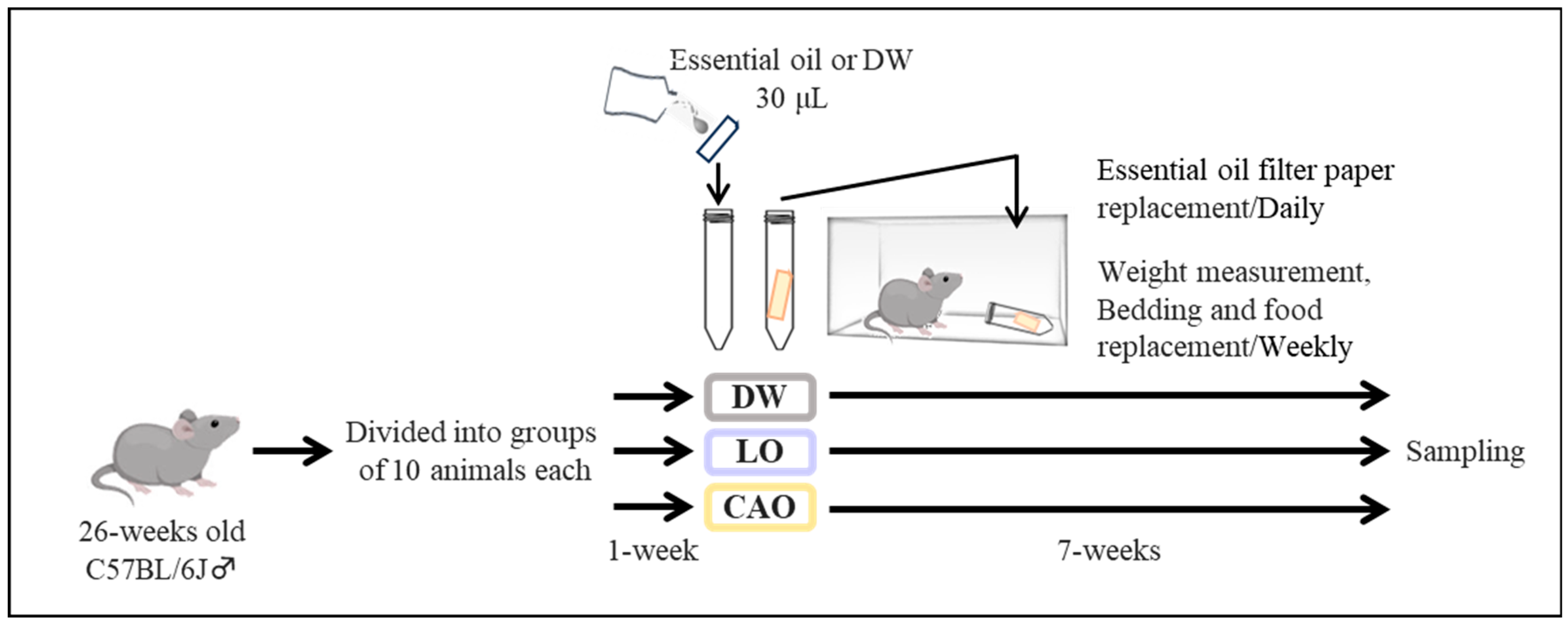
4.1. Mouse Maintenance and Essential Oil Inhalation
4.2. Sampling
4.3. Oil Red O Staining
4.4. LabAssay
4.5. Total RNA Extraction for Gene Expression Analysis
4.6. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Spisni, E.; Valerii, M.C.; Massimino, M.L. Essential Oil Molecules Can Break the Loop of Oxidative Stress in Neurodegenerative Diseases. Biology 2023, 12, 1504. [Google Scholar] [CrossRef]
- Lizarraga-Valderrama, L.R. Effects of essential oils on the central nervous system: Focus on mental health. Phytother. Res. 2021, 35, 657–679. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Dong, W.Y.; Guo, J.R.; Huang, J.Y.; Cheng, P.K.; Yang, Y.; Liu, A.; Yang, X.L.; Zhu, X.; Zhang, Z.; et al. A Neural Circuit For Bergamot Essential Oil-Induced Anxiolytic Effects. Adv. Sci. 2025, 12, e2406766. [Google Scholar] [CrossRef]
- Zhang, Y.; Long, Y.; Yu, S.; Li, D.; Yang, M.; Guan, Y.; Zhang, D.; Wan, J.; Liu, S.; Shi, A.; et al. Natural volatile oils derived from herbal medicines: A promising therapeutic way for treating depressive disorder. Pharmacol. Res. 2021, 164, 105376. [Google Scholar] [CrossRef] [PubMed]
- Suntar, I.; Khan, H.; Patel, S.; Celano, R.; Rastrelli, L. An Overview on Citrus aurantium L.: Its Functions Food as Ingredient and Therapeutic Agent. Oxid. Med. Cell Longev. 2018, 2018, 7864269. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.P.; Fassin, J., Jr.; Baziloni, E.M.F.; Almeida, R.N.; Mattei, R.; Leite, J.R. Behavioral effects of essential oil of Citrus aurantium L. inhalation in rats. Braz. J. Pharmacogn. 2008, 18, 661–666. [Google Scholar] [CrossRef]
- de Moraes Pultrini, A.; Almeida Galindo, L.; Costa, M. Effects of the essential oil from Citrus aurantium L. in experimental anxiety models in mice. Life Sci. 2006, 78, 1720–1725. [Google Scholar] [CrossRef]
- Tan, L.; Liao, F.F.; Long, L.Z.; Ma, X.C.; Peng, Y.X.; Lu, J.M.; Qu, H.; Fu, C.G. Essential oils for treating anxiety: A systematic review of randomized controlled trials and network meta-analysis. Front. Public Health 2023, 11, 1144404. [Google Scholar] [CrossRef]
- Scandurra, C.; Mezzalira, S.; Cutillo, S.; Zapparella, R.; Statti, G.; Maldonato, N.M.; Locci, M.; Bochicchio, V. The effectiveness of neroli essential oil in relieving anxiety and perceived pain in women during labor: A Randomized controlled trial. Healthcare 2022, 10, 366. [Google Scholar] [CrossRef]
- Nascimento, J.C.; Gonçalves, V.S.D.S.; Souza, B.R.S.; Nascimento, L.C.; Carvalho, B.M.R.; Nogueira, P.C.L.; Santos, J.P.S.; Borges, L.P.; Goes, T.C.; Souza, J.B.; et al. Effectiveness of aromatherapy with sweet orange oil (Citrus sinensis L.) in relieving pain and anxiety during labor. Explore 2024, 21, 103081. [Google Scholar] [CrossRef] [PubMed]
- Bargi, S.; Bahraminejad, N.; Jafari, S.; Fallah, R. The Effect of Aromatherapy with Citrus aurantium Aroma on Pain after Orthopedic Surgery: A Randomized Clinical Trial. J. Caring Sci. 2023, 12, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.S.; Tu, Y.K.; Chou, F.H.; Fang, C.J.; Chang, S.L. Effect of inhaled aromatherapy on sleep quality in critically ill patients: A systematic review and network meta-analysis. J. Clin. Nurs. 2024, 34, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, F.; Moradi, M.; Niazi, A.; Jamali, J. The Impact of Aromatherapy with Citrus aurantium Essential Oil on Sleep Quality in Pregnant Women with Sleep Disorders: A Randomized Controlled Clinical Trial. Int. J. Community Based Nurs. Midwifery 2022, 10, 160–171. [Google Scholar]
- Hong, S.J.; Cho, J.; Boo, C.G.; Youn, M.Y.; Pan, J.H.; Kim, J.K.; Shin, E.-C. Inhalation of Patchouli (Pogostemon Cablin Benth.) essential oil improved metabolic parameters in obesity-induced Sprague Dawley rats. Nutrients 2020, 12, 2077. [Google Scholar] [CrossRef]
- Batubara, I.; Suparto, I.H.; Sa’diah, S.; Matsuoka, R.; Mitsunaga, T. Effects of inhaled citronella oil and related compounds on rat body weight and brown adipose tissue sympathetic nerve. Nutrients 2015, 7, 1859–1870. [Google Scholar] [CrossRef]
- Hong, S.J.; Yoon, S.; Jo, S.M.; Jeong, H.; Youn, M.Y.; Kim, Y.J.; Kim, J.K.; Shin, E.C. Olfactory Stimulation by Fennel (Foeniculum vulgare Mill.) Essential Oil Improves Lipid Metabolism and Metabolic Disorders in High Fat-Induced Obese Rats. Nutrients 2022, 14, 741. [Google Scholar] [CrossRef]
- Shen, J.; Niijima, A.; Tanida, M.; Horii, Y.; Maeda, K.; Nagai, K. Olfactory stimulation with scent of grapefruit oil affects autonomic nerves, lipolysis and appetite in rats. Neurosci. Lett. 2005, 380, 289–294. [Google Scholar] [CrossRef]
- Shen, J.; Niijima, A.; Tanida, M.; Horii, Y.; Maeda, K.; Nagai, K. Olfactory stimulation with scent of lavender oil affects autonomic nerves, lipolysis and appetite in rats. Neurosci. Lett. 2005, 383, 188–193. [Google Scholar] [CrossRef]
- Guimarães, A.C.; de Moura, E.G.; Silva, S.G.; Lopes, B.P.; Bertasso, I.M.; Pietrobon, C.B.; Quitete, F.T.; de Oliveira Malafaia, T.; Souza, É.P.G.; Lisboa, P.C.; et al. Citrus aurantium L. and synephrine improve brown adipose tissue function in adolescent mice programmed by early postnatal overfeeding. Front. Nutr. 2024, 10, 1278121. [Google Scholar] [CrossRef]
- Dodonova, S.A.; Zhidkova, E.M.; Kryukov, A.A.; Valiev, T.T.; Kirsanov, K.I.; Kulikov, E.P.; Budunova, I.V.; Yakubovskaya, M.G.; Lesovaya, E.A. Synephrine and Its Derivative Compound A: Common and Specific Biological Effects. Int. J. Mol. Sci. 2023, 24, 17537. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, R.; Harrigan, K.; Tremblay, J.F.; Hedrei, P.; Lamarche, M.; Morais, J.A. Increase in the thermic effect of food in women by adrenergic amines extracted from Citrus aurantium. Obes. Res. 2006, 13, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Sharpe Patricia, A.; Granner Michelle, L.; Conway Joan, M.; Ainsworth Barbara, E.; Dobre, M. Availability of weight-loss supplements: Results of an audit of retail outlets in a southeastern city. J. Am. Diet. Assoc. 2006, 106, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- National Center for Complementary and Integrative Health. Bitter Orange. 2008. Available online: https://www.nccih.nih.gov/health/bitter-orange (accessed on 3 November 2008).
- Wang, M.; Han, H.; Wan, F.; Zhong, R.; Do, Y.J.; Oh, S.I.; Lu, X.; Liu, L.; Yi, B.; Zhang, H. Dihydroquercetin Supplementation Improved Hepatic Lipid Dysmetabolism Mediated by Gut Microbiota in High-Fat Diet (HFD)-Fed Mice. Nutrients 2022, 14, 5214. [Google Scholar] [CrossRef]
- Yao, W.; Jiao, Y.; Zhou, Y.; Luo, X. KLF13 suppresses the proliferation and growth of colorectal cancer cells through transcriptionally inhibiting HMGCS1-mediated cholesterol biosynthesis. Cell Biosci. 2020, 10, 76. [Google Scholar] [CrossRef]
- Haaz, S.; Fontaine, K.R.; Cutter, G.; Limdi, N.; Perumean-Chaney, S.; Allison, D.B. Citrus aurantium and synephrine alkaloids in the treatment of overweight and obesity: An update. Obes. Rev. 2006, 7, 79–88. [Google Scholar] [CrossRef]
- Arbo, M.D.; Schmitt, G.C.; Limberger, M.F.; Charão, M.F.; Moro, A.M.; Ribeiro, G.L.; Dallegrave, E.; Garcia, S.C.; Leal, M.B.; Limberger, R.P. Subchronic toxicity of Citrus aurantium L. (Rutaceae) extract and p-synephrine in mice. Regul. Toxicol. Pharmacol. 2009, 54, 114–117. [Google Scholar] [CrossRef]
- Deng, F.; Li, D.; Lei, L.; Yang, Q.; Li, Q.; Wang, H.; Deng, J.; Zheng, Q.; Jiang, W. Association between apolipoprotein B/A1 ratio and coronary plaque vulnerability in patients with atherosclerotic cardiovascular disease: An intravascular optical coherence tomography study. Cardiovasc. Diabetol. 2021, 20, 188. [Google Scholar] [CrossRef]
- Song, T.; Wang, P.; Li, C.; Jia, L.; Liang, Q.; Cao, Y.; Dong, P.; Shi, H.; Jiang, M. Salidroside simultaneously reduces de novo lipogenesis and cholesterol biosynthesis to attenuate atherosclerosis in mice. Biomed. Pharmacother. 2021, 134, 111137. [Google Scholar] [CrossRef]
- Ma, X.; Bai, Y.; Liu, K.; Han, Y.; Zhang, J.; Liu, Y.; Hou, X.; Hao, E.; Hou, Y.; Bai, G. Ursolic acid inhibits the cholesterol biosynthesis and alleviates high fat diet-induced hypercholesterolemia via irreversible inhibition of HMGCS1 in vivo. Phytomedicine 2022, 103, 154233. [Google Scholar] [CrossRef]
- Chambers, K.F.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients 2019, 11, 2588. [Google Scholar] [CrossRef] [PubMed]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Jelinek, D.F.; Andersson, S.; Slaughter, C.A.; Russell, D.W. Cloning and regulation of cholesterol 7 alpha-hydroxylase, the rate-limiting enzyme in bile acid biosynthesis. J. Biol. Chem. 1990, 265, 8190–8197. [Google Scholar] [CrossRef]
- Attia, H.G.; El-Morshedy, S.M.; Nagy, A.M.; Ibrahim, A.M.; Aleraky, M.; Abdelrahman, S.S.; Osman, S.M.; Alasmari, S.M.; El Raey, M.A.; Abdelhameed, M.F. Citrus clementine Peel Essential Oil Ameliorates Potassium Dichromate-Induced Lung Injury: Insights into the PI3K/AKT Pathway. Metabolites 2024, 14, 68. [Google Scholar] [CrossRef]
- Patel, M.; Narke, D.; Kurade, M.; Frey, K.M.; Rajalingam, S.; Siddiquee, A.; Mustafa, S.J.; Ledent, C.; Ponnoth, D.S. Limonene-induced activation of A(2A) adenosine receptors reduces airway inflammation and reactivity in a mouse model of asthma. Purinergic Signal. 2020, 16, 415–426. [Google Scholar] [CrossRef]
- Wittmeier, P.; Hummel, S. Agarose gel electrophoresis to assess PCR product yield: Comparison with spectrophotometry, fluorometry and qPCR. Biotechniques 2022, 72, 155–158. [Google Scholar] [CrossRef]
- Yamamoto, T.; Inui, T.; Tsuji, T. The odour of Osmanthus fragrans attenuates food intake. Sci. Rep. 2013, 3, 1518. [Google Scholar] [CrossRef] [PubMed]
- Takenoya, F.; Yamashita, M.; Shibato, J.; Kimura, A.; Chiba, Y.; Hirabayashi, T.; Shioda, S. Histological observation of rat hypothalamic feeding-regulated neurons in peppermint and ginger essential oil exposure. J. Jpn. Soc. Aromather. 2020, 21, 40–48. [Google Scholar]
- Liao, J.T.; Huang, Y.W.; Hou, C.Y.; Wang, J.J.; Wu, C.C.; Hsieh, S.L. D-Limonene Promotes Anti-Obesity in 3T3-L1 Adipocytes and High-Calorie Diet-Induced Obese Rats by Activating the AMPK Signaling Pathway. Nutrients 2023, 15, 267. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Hsiao, S.Y.; Lin, Y.H.; Tsai, G.J. Effects of Fermented Citrus Peel on Ameliorating Obesity in Rats Fed with High-Fat Diet. Molecules 2022, 27, 8966. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi de Alvarenga, J.F.; Lei Preti, C.; Santos Martins, L.; Noronha Hernandez, G.; Genaro, B.; Lamesa Costa, B.; Gieseler Dias, C.; Purgatto, E.; Fiamoncini, J. Identification of D-Limonene Metabolites by LC-HRMS: An Exploratory Metabolic Switching Approach in a Mouse Model of Diet-Induced Obesity. Metabolites 2022, 12, 1246. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | ||
|---|---|---|---|
| Accession | Nucleotide Sequence (5′-3′) | Nucleotide Sequence (5′-3′) | Gene Name |
| NM_007988 | AATCCATCATCAACATCATCCA | CCACTGACTCTTCACAGACCAG | Fasn |
| NM_009692 | ACGAATTCCAGAAGAAATGGAA | GTGGTACTCGTTCAAGGTAGGG | Apoa1 |
| NM_009127 | CACCTTCTTGCGATACACTCTG | CTCCCGTCTCCAGTTCTCTTAA | Scd1 |
| NM_009463 | AACTCTCTGCCAGGACAGTACC | AACGGAGCTGTTCATTTGATTT | Ucp1 |
| NM_009693 | GTTGGTGAGTCCACAAGATTGA | GCTTGGTTGCAGGTATAGTTCC | Apob |
| NM_145942 | TGGTATCTGGTCAGAGTGGATG | GACCACAACAGGAAGCATGTTA | Hmgcs1 |
| NM_001360166 | TCACATGGTT CACAACAGAT CA | GCAC AGAGACTCCT CAGATGTG | Hmgcr |
| NM_153526 | ACCACGTCTGGAACTATCCAAG | CTCCCAGGTGACTGTCAATACA | Insig1 |
| NM_007824 | AAATACGACCGGTACCTTGATG | TAACGCTCAGCAGTCGTTACAT | Cyp7a1 |
| NM_001001303 | GCTACACTGAGGACCAGGTTGT | CTCCTGTTATTATGGGGGTCTG | GAPDH |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibato, J.; Kimura, A.; Yamashita, M.; Shioda, S.; Takenoya, F.; Rakwal, R. Effects of Essential Oil Inhalation on the Enhancement of Plasma and Liver Lipid Metabolism in Mice. Int. J. Mol. Sci. 2025, 26, 5674. https://doi.org/10.3390/ijms26125674
Shibato J, Kimura A, Yamashita M, Shioda S, Takenoya F, Rakwal R. Effects of Essential Oil Inhalation on the Enhancement of Plasma and Liver Lipid Metabolism in Mice. International Journal of Molecular Sciences. 2025; 26(12):5674. https://doi.org/10.3390/ijms26125674
Chicago/Turabian StyleShibato, Junko, Ai Kimura, Michio Yamashita, Seiji Shioda, Fumiko Takenoya, and Randeep Rakwal. 2025. "Effects of Essential Oil Inhalation on the Enhancement of Plasma and Liver Lipid Metabolism in Mice" International Journal of Molecular Sciences 26, no. 12: 5674. https://doi.org/10.3390/ijms26125674
APA StyleShibato, J., Kimura, A., Yamashita, M., Shioda, S., Takenoya, F., & Rakwal, R. (2025). Effects of Essential Oil Inhalation on the Enhancement of Plasma and Liver Lipid Metabolism in Mice. International Journal of Molecular Sciences, 26(12), 5674. https://doi.org/10.3390/ijms26125674