

Plakophilin 3 Is Involved in Basal Body Docking in Multiciliated Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
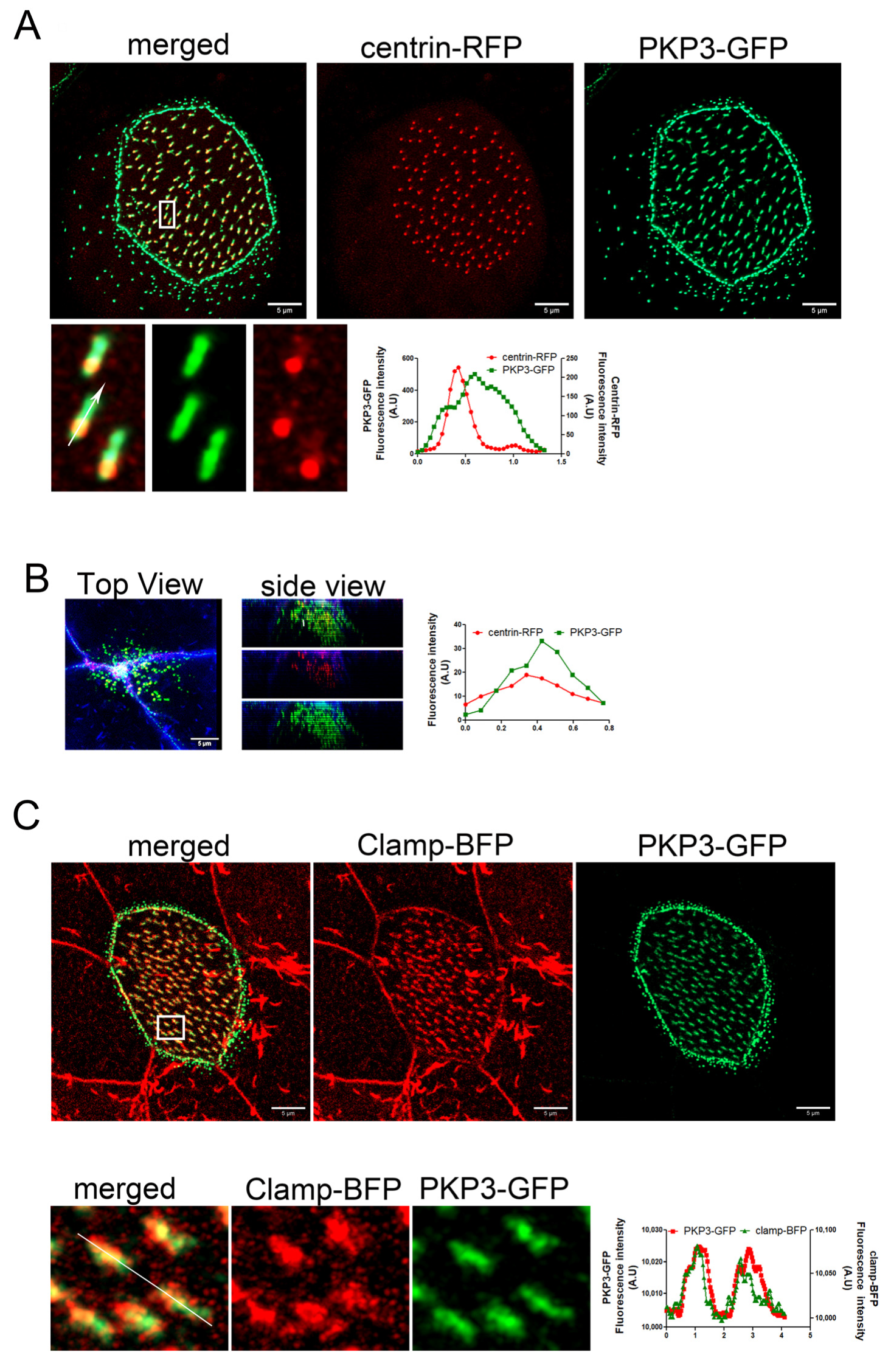
2.1. Plakophilin 3 Localizes at the Striated Rootlet
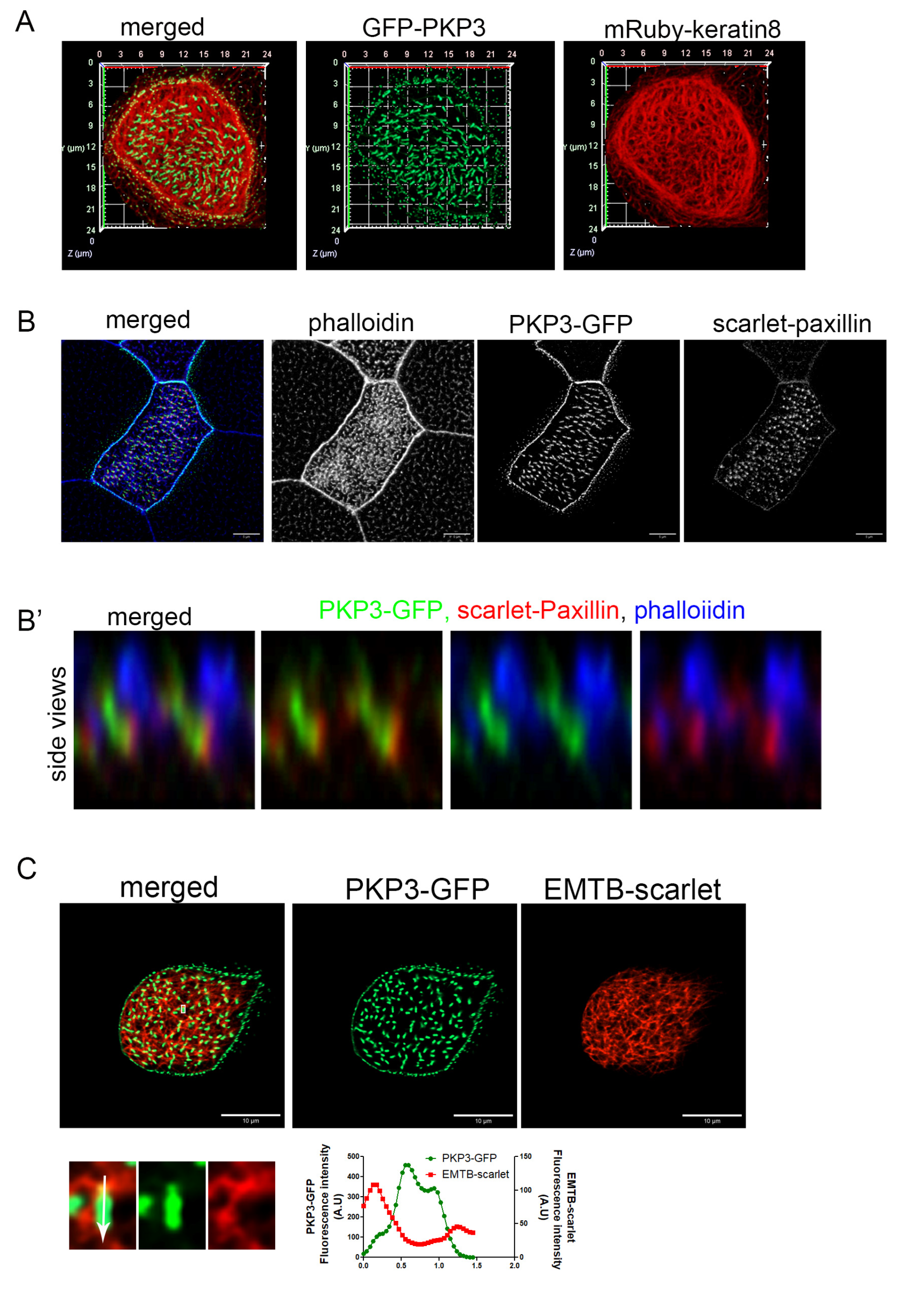
2.2. PKP3 Does Not Co-Localize with Keratin 8 Intermediate Filaments, Actin, or Microtubules
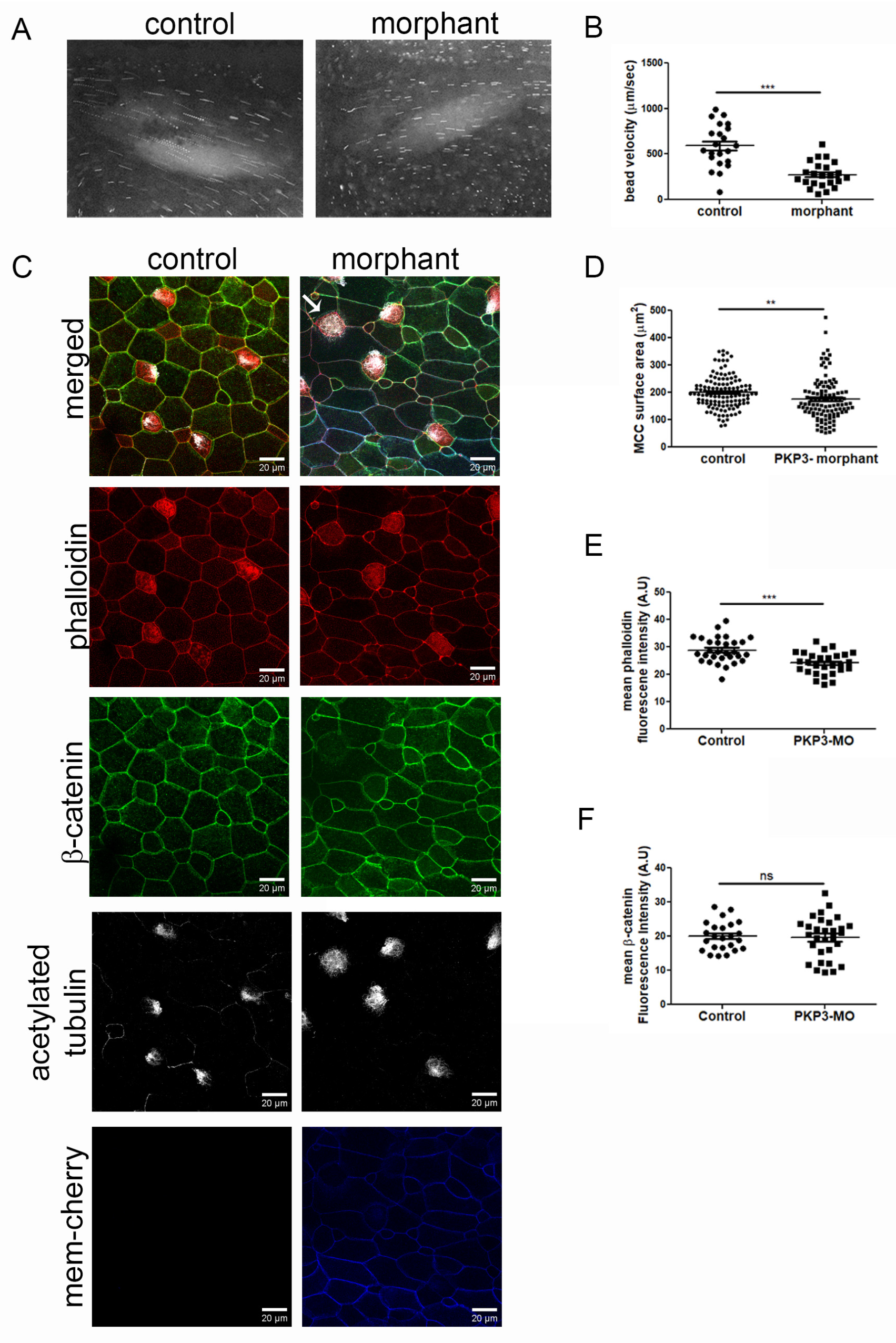
2.3. Downregulation of PKP3 Affects the Function of MCCs
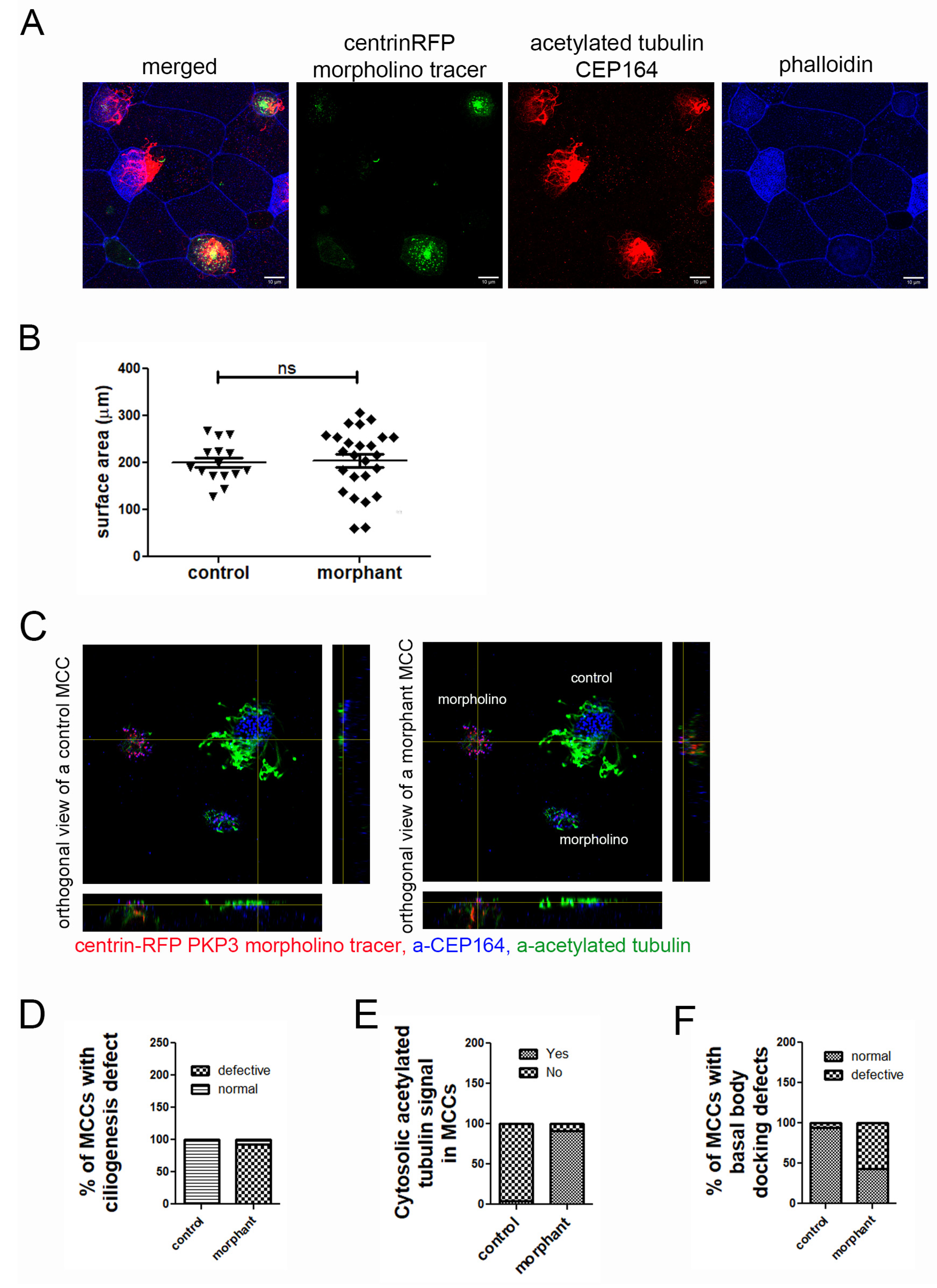
2.4. Basal Body Docking Defect in PKP3 Morphant MCCs Is Independent of MCC Intercalation
2.5. Basal Body Docking Defect Is Independent of the Actin Cytoskeleton
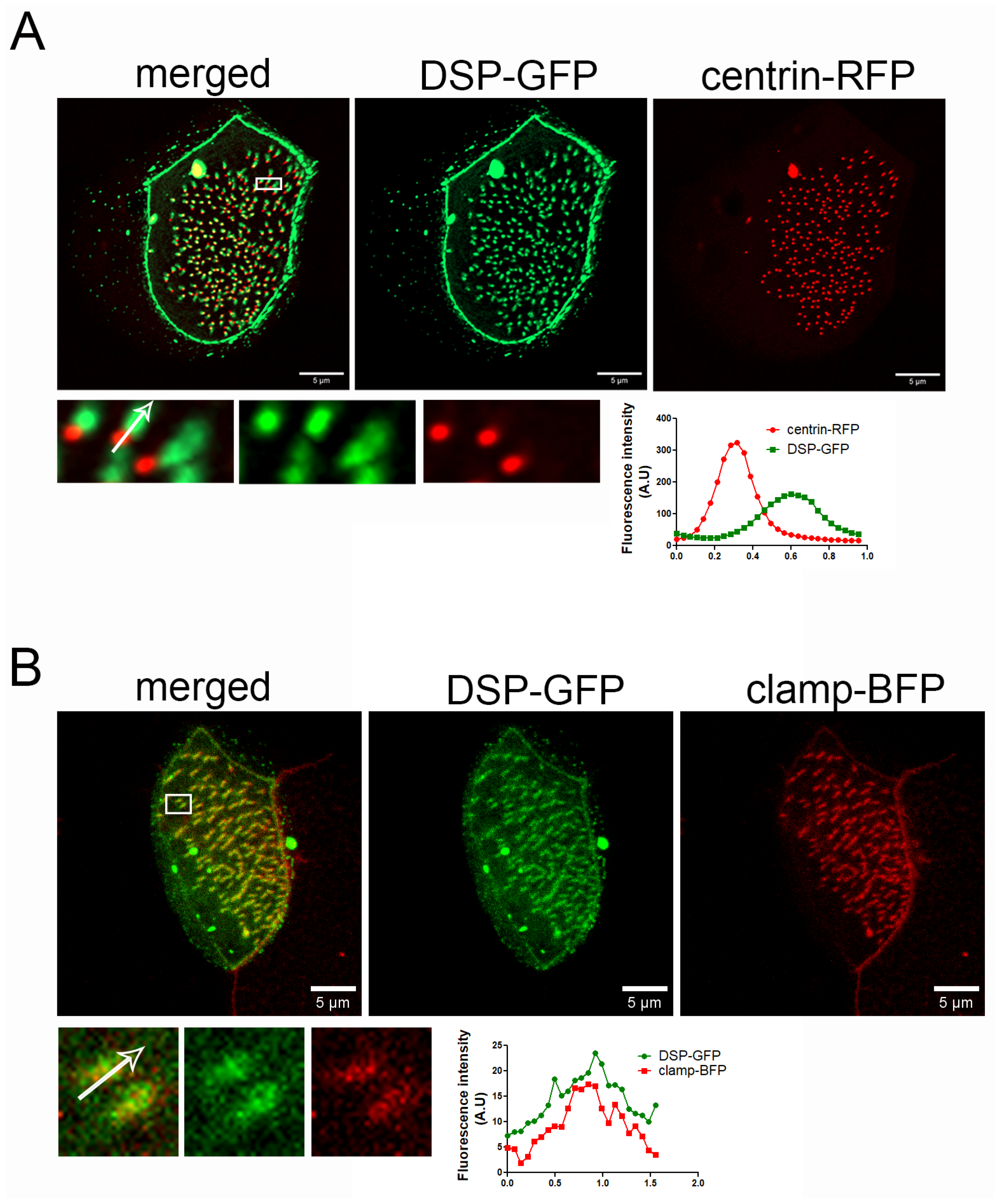
2.6. Downregulation of PKP3 Does Not Affect the Recruitment of DSP to Basal Bodies
3. Discussion
4. Materials and Methods
4.1. Embryo Manipulations and Microinjections
4.2. DNA Constructs and Morpholino Oligonucleotide
4.3. Fluid Flow Assay
4.4. Immunostaining
4.5. Image Analysis and Quantifications
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MCC | Multiciliated cell |
| PKP3 | Plakophilin 3 |
| DSP | Desmoplakin |
| CA | Ciliary adhesions |
References
- Despotes, K.A.; Zariwala, M.A.; Davis, S.D.; Ferkol, T.W. Primary Ciliary Dyskinesia: A Clinical Review. Cells 2024, 13, 974. [Google Scholar] [CrossRef] [PubMed]
- Afzelius, B.A. A Human Syndrome Caused by Immotile Cilia. Science 1976, 193, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, S.P. Reconstructions of centriole formation and ciliogenesis in mammalian lungs. J. Cell Sci. 1968, 3, 207–230. [Google Scholar] [CrossRef] [PubMed]
- Boisvieux-Ulrich, E.; Lainé, M.-C.; Sandoz, D. Cytochalasin D inhibits basal body migration and ciliary elongation in quail oviduct epithelium. Cell Tissue Res. 1990, 259, 443–454. [Google Scholar] [CrossRef]
- Dawe, H.R.; Farr, H.; Gull, K. Centriole/basal body morphogenesis and migration during ciliogenesis in animal cells. J. Cell Sci. 2007, 120, 7–15. [Google Scholar] [CrossRef]
- Lemullois, M.; Boisvieux-Ulrich, E.; Laine, M.; Chailley, B.; Sandoz, D. Development and functions of the cytoskeleton during ciliogenesis in metazoa. Biol. Cell 1988, 63, 195–208. [Google Scholar] [CrossRef]
- Pan, J.; You, Y.; Huang, T.; Brody, S.L. RhoA-mediated apical actin enrichment is required for ciliogenesis and promoted by Foxj1. J. Cell Sci. 2007, 120, 1868–1876. [Google Scholar] [CrossRef]
- Ioannou, A.; Santama, N.; Skourides, P.A. Xenopus laevis nucleotide binding protein 1 (xNubp1) is important for convergent extension movements and controls ciliogenesis via regulation of the actin cytoskeleton. Dev. Biol. 2013, 380, 243–258. [Google Scholar] [CrossRef]
- Drysdale, T.A.; Elinson, R.P. Cell Migration and Induction in the Development of the Surface Ectodermal Pattern of the Xenopus laevis Tadpole: (Xenopus /ciliated cell/hatching gland/cement gland/ectodermal differentiation). Dev. Growth Differ. 1992, 34, 51–59. [Google Scholar] [CrossRef]
- Deblandre, G.A.; Wettstein, D.A.; Koyano-Nakagawa, N.; Kintner, C. A two-step mechanism generates the spacing pattern of the ciliated cells in the skin of Xenopus embryos. Development 1999, 126, 4715–4728. [Google Scholar] [CrossRef]
- Kulkarni, S.; Marquez, J.; Date, P.; Ventrella, R.; Mitchell, B.J.; Khokha, M.K. Mechanical stretch scales centriole number to apical area via Piezo1 in multiciliated cells. eLife 2021, 10, e66076. [Google Scholar] [CrossRef] [PubMed]
- Sedzinski, J.; Hannezo, E.; Tu, F.; Biro, M.; Wallingford, J.B. Emergence of an Apical Epithelial Cell Surface In Vivo. Dev. Cell 2016, 36, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Garcia, G.; Reiter, J.F. A primer on the mouse basal body. Cilia 2016, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Tateishi, K.; Nishida, T.; Inoue, K.; Tsukita, S. Three-dimensional Organization of Layered Apical Cytoskeletal Networks Associated with Mouse Airway Tissue Development. Sci. Rep. 2017, 7, srep43783. [Google Scholar] [CrossRef]
- Werner, M.E.; Hwang, P.; Huisman, F.; Taborek, P.; Yu, C.C.; Mitchell, B.J. Actin and microtubules drive differential aspects of planar cell polarity in multiciliated cells. J. Cell Biol. 2011, 195, 19–26. [Google Scholar] [CrossRef]
- Klotz, C.; Bordes, N.; Laine, M.C.; Sandoz, D.; Bornens, M. Myosin at the apical pole of ciliated epithelial cells as revealed by a monoclonal antibody. J. Cell Biol. 1986, 103, 613–619. [Google Scholar] [CrossRef]
- Clare, D.K.; Magescas, J.; Piolot, T.; Dumoux, M.; Vesque, C.; Pichard, E.; Dang, T.; Duvauchelle, B.; Poirier, F.; Delacour, D. Basal foot MTOC organizes pillar MTs required for coordination of beating cilia. Nat. Commun. 2014, 5, 4888. [Google Scholar] [CrossRef]
- Kunimoto, K.; Yamazaki, Y.; Nishida, T.; Shinohara, K.; Ishikawa, H.; Hasegawa, T.; Okanoue, T.; Hamada, H.; Noda, T.; Tamura, A.; et al. Coordinated Ciliary Beating Requires Odf2-Mediated Polarization of Basal Bodies via Basal Feet. Cell 2012, 148, 189–200. [Google Scholar] [CrossRef]
- Vladar, E.K.; Lee, Y.L.; Stearns, T.; Axelrod, J.D. Observing planar cell polarity in multiciliated mouse airway epithelial cells. In Methods in Cell Biology [Internet]; Elsevier: Amsterdam, The Netherlands, 2015; pp. 37–54. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0091679X15000369 (accessed on 11 April 2025).
- Satir, P.; Dirksen, E.R. Function-Structure Correlations in Cilia from Mammalian Respiratory Tract. In Comprehensive Physiology [Internet], 1st ed.; Prakash, Y.S., Ed.; Wiley: Hoboken, NJ, USA, 1985; pp. 473–494. Available online: https://onlinelibrary.wiley.com/doi/10.1002/cphy.cp030115 (accessed on 11 April 2025).
- Gibbons, I.R. The relationship between the fine structure and direction of beat in gill cilia of a lamellibranch mollusc. J. Cell Biol. 1961, 11, 179–205. [Google Scholar] [CrossRef]
- Satir, P. Chirality of the cytoskeleton in the origins of cellular asymmetry. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150408. [Google Scholar] [CrossRef]
- Steinman, R.M. An electron microscopic study of ciliogenesis in developing epidermis and trachea in the embryo of Xenopus laevis. Am. J. Anat. 1968, 122, 19–55. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Yue, G.; Adamian, M.; Bulgakov, O.; Li, T. Rootletin, a novel coiled-coil protein, is a structural component of the ciliary rootlet. J. Cell Biol. 2002, 159, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gao, J.; Adamian, M.; Wen, X.-H.; Pawlyk, B.; Zhang, L.; Sanderson, M.J.; Zuo, J.; Makino, C.L.; Li, T. The Ciliary Rootlet Maintains Long-Term Stability of Sensory Cilia. Mol. Cell. Biol. 2005, 25, 4129–4137. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.V.; Kao, L.-R.; Jana, S.C.; Sivan-Loukianova, E.; Mendonça, S.; Cabrera, O.A.; Singh, P.; Cabernard, C.; Eberl, D.F.; Bettencourt-Dias, M.; et al. Rootletin organizes the ciliary rootlet to achieve neuron sensory function in Drosophila. J. Cell Biol. 2015, 211, 435–453. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Timbers, T.A.; Kennedy, J.; Blacque, O.E.; Leroux, M.R. Striated Rootlet and Nonfilamentous Forms of Rootletin Maintain Ciliary Function. Curr. Biol. 2013, 23, 2016–2022. [Google Scholar] [CrossRef]
- Antoniades, I.; Stylianou, P.; Skourides, P.A. Making the Connection: Ciliary Adhesion Complexes Anchor Basal Bodies to the Actin Cytoskeleton. Dev. Cell 2014, 28, 70–80. [Google Scholar] [CrossRef]
- Yasunaga, T.; Wiegel, J.; Bergen, M.D.; Helmstädter, M.; Epting, D.; Paolini, A.; Çiçek, Ö.; Radziwill, G.; Engel, C.; Brox, T.; et al. Microridge-like structures anchor motile cilia. Nat. Commun. 2022, 13, 2056. [Google Scholar] [CrossRef]
- Stark, C. BioGRID: A general repository for interaction datasets. Nucleic Acids Res. 2006, 34, D535–D539. [Google Scholar] [CrossRef]
- Liu, Q.; Tan, G.; Levenkova, N.; Li, T.; Pugh, E.N., Jr.; Rux, J.J.; Speicher, D.W.; Pierce, E.A. The Proteome of the Mouse Photoreceptor Sensory Cilium Complex. Mol. Cell. Proteom. 2007, 6, 1299–1317. [Google Scholar] [CrossRef]
- Sammeta, N.; Yu, T.; Bose, S.C.; McClintock, T.S. Mouse olfactory sensory neurons express 10,000 genes. J. Comp. Neurol. 2007, 502, 1138–1156. [Google Scholar] [CrossRef]
- Munoz, W.A.; Kloc, M.; Cho, K.; Lee, M.; Hofmann, I.; Sater, A.; Vleminckx, K.; McCrea, P.D. Plakophilin-3 Is Required for Late Embryonic Amphibian Development, Exhibiting Roles in Ectodermal and Neural Tissues. PLoS ONE 2012, 7, e34342. [Google Scholar] [CrossRef]
- Bharathan, N.K.; Dickinson, A.J. Desmoplakin is required for epidermal integrity and morphogenesis in the Xenopus laevis embryo. Dev. Biol. 2019, 450, 115–131. [Google Scholar] [CrossRef]
- Hatzfeld, M.; Wolf, A.; Keil, R. Plakophilins in Desmosomal Adhesion and Signaling. Cell Commun. Adhes. 2014, 21, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Briggs, J.A.; Weinreb, C.; Wagner, D.E.; Megason, S.; Peshkin, L.; Kirschner, M.W.; Klein, A.M. The dynamics of gene expression in vertebrate embryogenesis at single-cell resolution. Science 2018, 360, eaar5780. [Google Scholar] [CrossRef] [PubMed]
- Bonneé, S.; Gilbert, B.; Hatzfeld, M.; Chen, X.; Green, K.J.; van Roy, F. Defining desmosomal plakophilin-3 interactions. J. Cell Biol. 2003, 161, 403–416. [Google Scholar] [CrossRef]
- Shiratsuchi, G.; Konishi, S.; Yano, T.; Yanagihashi, Y.; Nakayama, S.; Katsuno, T.; Kashihara, H.; Tanaka, H.; Tsukita, K.; Suzuki, K.; et al. Dual-color live imaging unveils stepwise organization of multiple basal body arrays by cytoskeletons. Embo Rep. 2024, 25, 1176–1207. [Google Scholar] [CrossRef]
- Bulinski, J.C.; Odde, D.J.; Howell, B.J.; Salmon, T.D.; Waterman-Storer, C.M. Rapid dynamics of the microtubule binding of ensconsin in vivo. J. Cell Sci. 2001, 114, 3885–3897. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, J.L.; Vladar, E.K.; Axelrod, J.D.; Kintner, C. Multicilin promotes centriole assembly and ciliogenesis during multiciliate cell differentiation. Nat. Cell Biol. 2012, 14, 140–147. [Google Scholar] [CrossRef]
- Li, Z.; Aizenman, C.D.; Cline, H.T. Regulation of Rho GTPases by Crosstalk and Neuronal Activity In Vivo. Neuron 2002, 33, 741–750. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Griffin, J.N.; Date, P.P.; Liem, K.F.; Khokha, M.K. WDR5 Stabilizes Actin Architecture to Promote Multiciliated Cell Formation. Dev. Cell 2018, 46, 595–610.e3. [Google Scholar] [CrossRef]
- Park, T.J.; Mitchell, B.J.; Abitua, P.B.; Kintner, C.; Wallingford, J.B. Dishevelled controls apical docking and planar polarization of basal bodies in ciliated epithelial cells. Nat. Genet. 2008, 40, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Chatzifrangkeskou, M.; Kouis, P.; Skourides, P.A. JNK regulates ciliogenesis through the interflagellar transport complex and actin networks. J. Cell Biol. 2023, 222, e202303052. [Google Scholar] [CrossRef] [PubMed]
- Kozminski, K.G.; Beech, P.L.; Rosenbaum, J.L. The Chlamydomonas kinesin-like protein FLA10 is involved in motility associated with the flagellar membrane. J. Cell Biol. 1995, 131, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Cole, D.G.; Diener, D.R.; Himelblau, A.L.; Beech, P.L.; Fuster, J.C.; Rosenbaum, J.L. Chlamydomonas Kinesin-II–dependent Intraflagellar Transport (IFT): IFT Particles Contain Proteins Required for Ciliary Assembly in Caenorhabditis elegans Sensory Neurons. J. Cell Biol. 1998, 141, 993–1008. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.L.; Scholey, J.M. Heterotrimeric Kinesin-II Is Required for the Assembly of Motile 9+2 Ciliary Axonemes on Sea Urchin Embryos. J. Cell Biol. 1997, 138, 1009–1022. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Louka, P.; Kyriakou, C.; Diakourti, I.; Skourides, P. Plakophilin 3 Is Involved in Basal Body Docking in Multiciliated Cells. Int. J. Mol. Sci. 2025, 26, 5381. https://doi.org/10.3390/ijms26115381
Louka P, Kyriakou C, Diakourti I, Skourides P. Plakophilin 3 Is Involved in Basal Body Docking in Multiciliated Cells. International Journal of Molecular Sciences. 2025; 26(11):5381. https://doi.org/10.3390/ijms26115381
Chicago/Turabian StyleLouka, Panagiota, Chrysovalantou Kyriakou, Ioanna Diakourti, and Paris Skourides. 2025. "Plakophilin 3 Is Involved in Basal Body Docking in Multiciliated Cells" International Journal of Molecular Sciences 26, no. 11: 5381. https://doi.org/10.3390/ijms26115381
APA StyleLouka, P., Kyriakou, C., Diakourti, I., & Skourides, P. (2025). Plakophilin 3 Is Involved in Basal Body Docking in Multiciliated Cells. International Journal of Molecular Sciences, 26(11), 5381. https://doi.org/10.3390/ijms26115381