
Targeted Gut Microbiota Modulation Enhances Levodopa Bioavailability and Motor Recovery in MPTP Parkinson’s Disease Models
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Antibiotic-Induced Gut Microbiota Depletion Improved L-Dopa Bioavailability
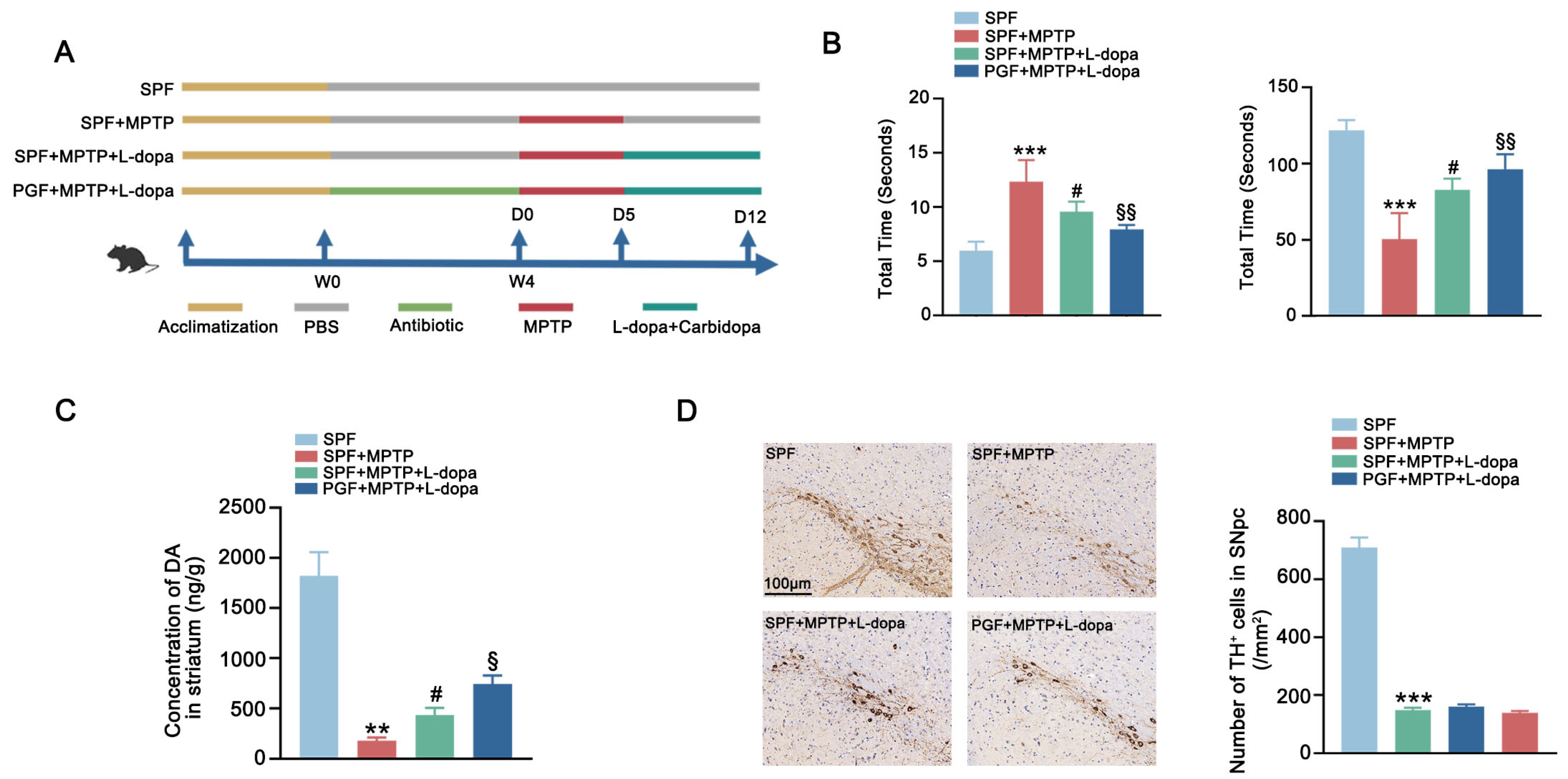
2.2. Antibiotic-Induced Gut Microbiota Depletion Enhanced Therapeutic Effect of L-Dopa in PD Mice
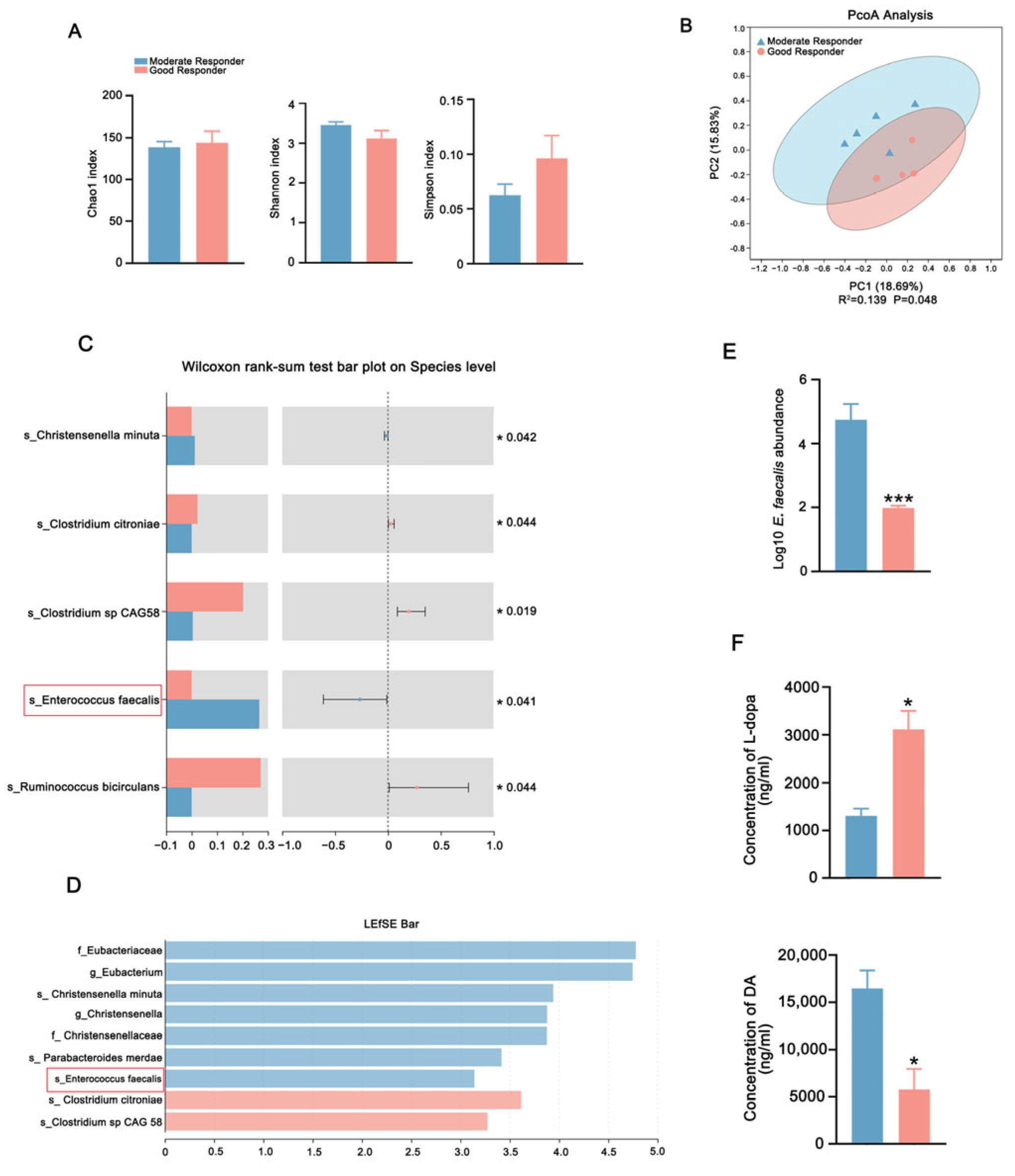
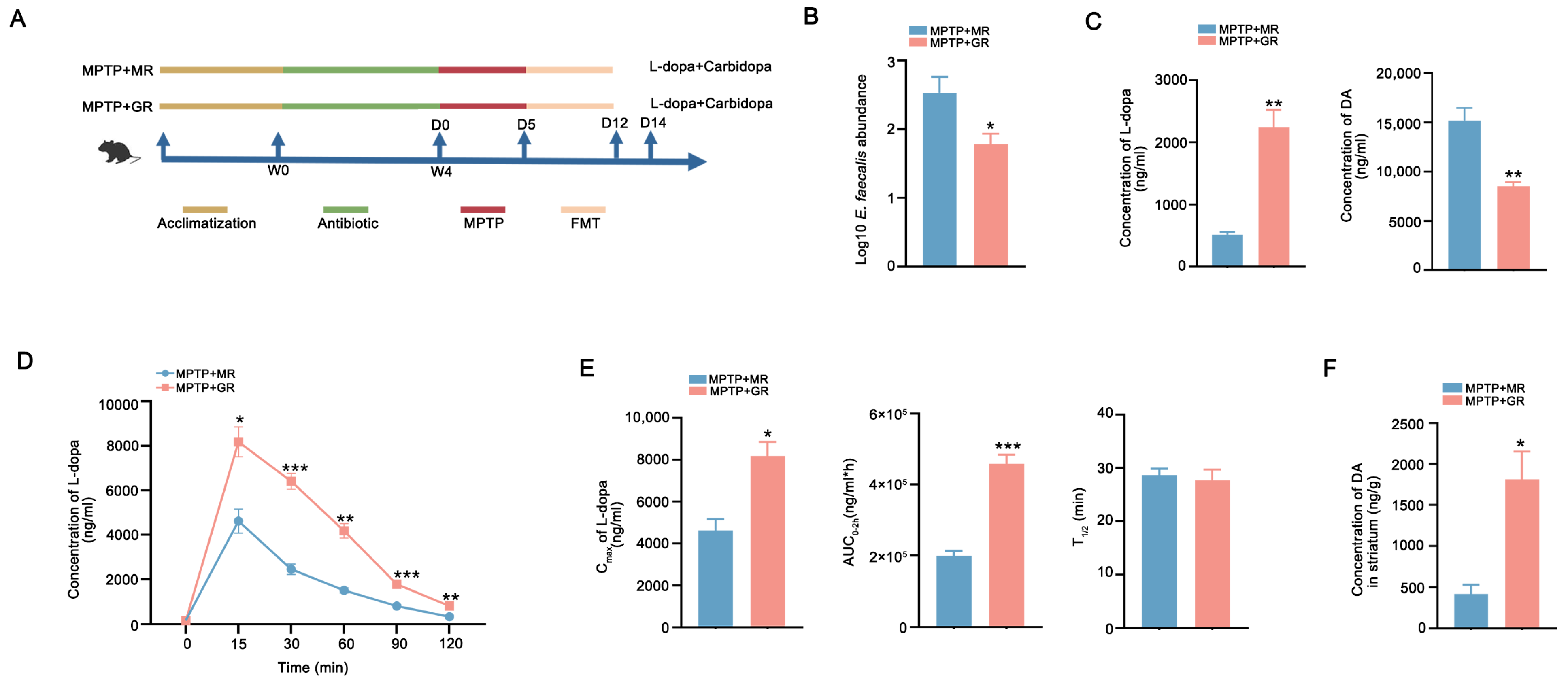
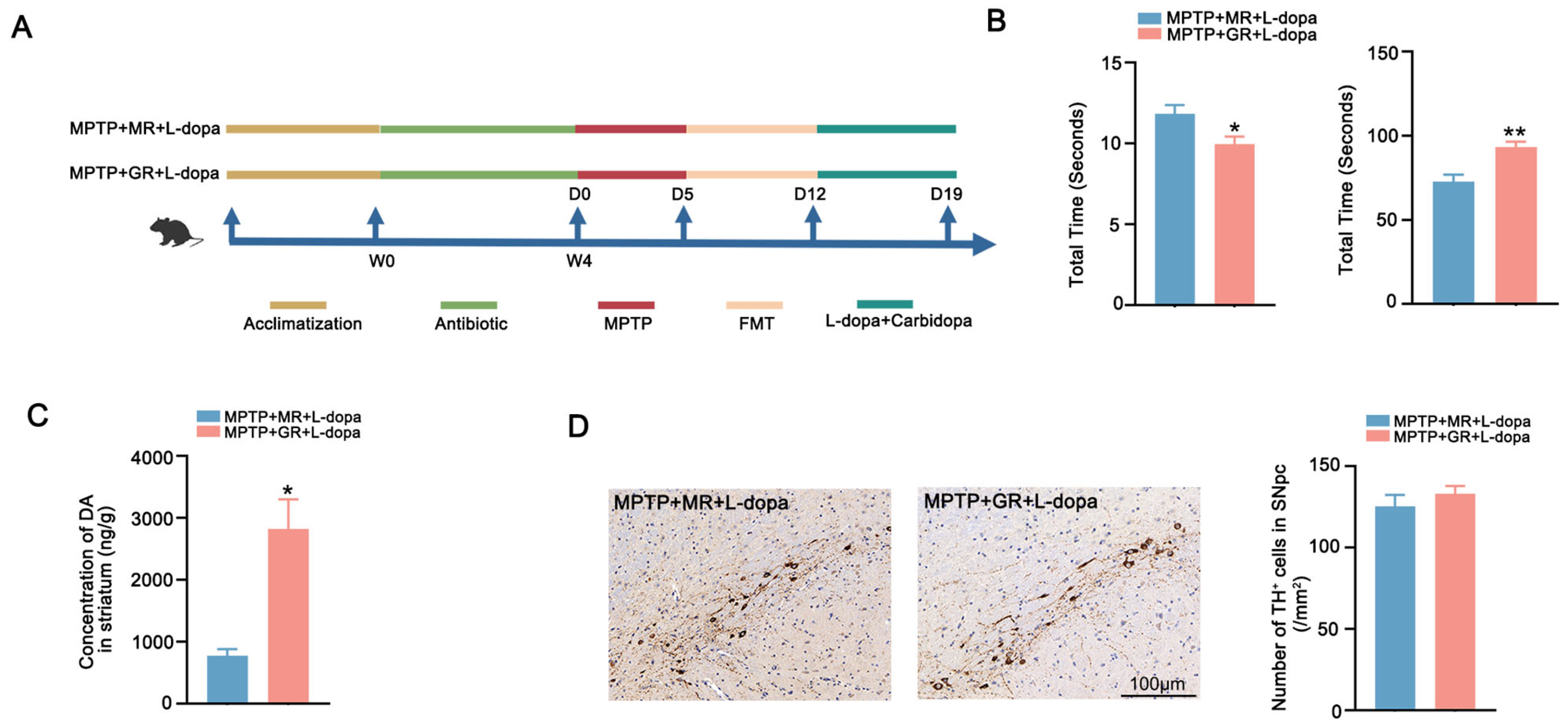
2.3. FMT from Good Responders Enhanced L-Dopa Bioavailability and Therapeutic Effect
3. Discussion
4. Materials and Methods
4.1. Study Approvals
4.2. Mouse Treatment
4.3. Quantitative Analysis of L-Dopa and DA
- (I)
- Standard Preparation
- Stock Solutions: L-dopa and DA single-component stock solutions (10 ppm) prepared in 10 mM HCl and acetonitrile, respectively.
- Standard Mixed External Standard Solution 1: We diluted L-dopa (1 ppm) and DA (1 ppm) with acetonitrile to prepare a mixed working solution.
- Standard Mixed External Standard Solution 2: We diluted L-dopa (100 ppb) and DA (100 ppb) with acetonitrile to prepare a mixed working solution.
- Standard Mixed Internal Standard Working Solution: We diluted L-dopa-d3 (1 ppm, MCE, America) and dopamine-d4 (1 ppm, MCE, America) with acetonitrile to prepare a mixed internal-standard working solution.
- Calibration Standards: We took 200 µL, 100 µL, and 50 µL of Standard Mixed External Standard Solution 1 and 200 µL and 100 µL of Standard Mixed External Standard Solution 2. We added 100 µL of the mixed internal-standard working solution to each tube, vortexed thoroughly, and analyzed using the instrument to obtain five points for the standard curve.
- (II)
- Sample Pre-treatment
- Culture Supernatant Sample Treatment: We centrifuged the culture supernatant at 13,000 rpm for 15 min at 4 °C. We collected 450 µL of the supernatant, added 50 µL of the mixed internal-standard working solution, vortexed thoroughly, filtered through a 0.22 µm membrane, and analyzed using the instrument.
- Mouse Serum Sample Treatment: We took 10 µL of serum sample, added 20 µL of the mixed internal-standard working solution, and added 70 µL of 40% acetonitrile trifluoroacetic acid solution. We vortexed for 1 min, centrifuged at 15,000 rpm for 20 min at 4 °C, collected the supernatant, and analyzed using the instrument.
- Mouse Brain Tissue Sample Treatment: Weigh 0.1 g of brain tissue, add 100 µL of 10 mM HCl solution, followed by 80 µL of 40% acetonitrile trifluoroacetic acid solution, and finally add 20 µL of the mixed internal-standard working solution. We homogenized using magnetic beads for 10 min, froze at −80 °C for 30 min, then centrifuged at 15,000 rpm for 20 min at 4 °C. We collected the supernatant, filtered through a 0.22 µm membrane, and analyzed using the instrument.
- (III)
- Ultra-High-Performance Liquid Chromatography–Tandem Mass Spectrometry (HPLC-MS/MS, Shimadzu LCMS-8050, Kyoto, Japan) Conditions
- Liquid Phase Conditions: Mobile phase A: 2.55 mM ammonium acetate, 0.25% formic acid in water; mobile phase B: acetonitrile. Chromatographic column: YMC TICLIC-C18 (4.6 × 100 mm, 1.9 μm). Injector temperature: 10 °C; injection volume: 10 µL; flow rate: 0.2 mL/min; gradient: started with 15% B phase.
- Mass Spectrometry Conditions: Ionization mode: electrospray ionization (ESI) in positive and negative ion modes; spray voltage: ±3.5 kV; nebulizer temperature: 560 °C; sheath gas pressure: 30 psi; auxiliary gas pressure: 20 psi; DL tube temperature: 350 °C; ion detection mode: multiple-reaction monitoring (MRM).
4.4. Behavioral Tests
4.4.1. Pole Test
4.4.2. Rotarod Test
4.5. Immunohistochemical (IHC) Staining
4.6. Participant Recruitment
4.7. Data and Sample Collection
4.8. L-Dopa Challenge Experiment
4.9. Metagenomic Sequencing
4.10. LEfSe
4.11. Fecal Sample DNA Extraction and Quantitative Real-Time PCR
4.12. Metabolic Assay of L-Dopa by Fecal Incubation In Vitro
4.13. Fecal Microbiota Transplantation
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dorsey, E.R.; Sherer, T.; Okun, M.S.; Bloem, B.R. The Emerging Evidence of the Parkinson Pandemic. J. Park. Dis. 2018, 8, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Okun, M.S. Diagnosis and Treatment of Parkinson Disease: A Review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.P.; Bianchine, J.R.; Spiegel, H.E.; Rivera-Calimlim, L.; Hersey, R.M. Metabolism of Levodopa in Patients with Parkinson’s Disease: Radioactive and Fluorometric Assays. Arch. Neurol. 1971, 25, 39–44. [Google Scholar] [CrossRef]
- Cacabelos, R. Parkinson’s Disease: From Pathogenesis to Pharmacogenomics. Int. J. Mol. Sci. 2017, 18, 551. [Google Scholar] [CrossRef]
- Maini Rekdal, V.; Bess, E.N.; Bisanz, J.E.; Turnbaugh, P.J.; Balskus, E.P. Discovery and inhibition of an interspecies gut bacterial pathway for Levodopa metabolism. Science 2019, 364, eaau6323. [Google Scholar] [CrossRef]
- Muleiro Alvarez, M.; Cano-Herrera, G.; Osorio Martínez, M.F.; Vega Gonzales-Portillo, J.; Monroy, G.R.; Murguiondo Pérez, R.; Torres-Ríos, J.A.; van Tienhoven, X.A.; Garibaldi Bernot, E.M.; Esparza Salazar, F.; et al. A Comprehensive Approach to Parkinson’s Disease: Addressing Its Molecular, Clinical, and Therapeutic Aspects. Int. J. Mol. Sci. 2024, 25, 7183. [Google Scholar] [CrossRef]
- Caputi, V.; Giron, M.C. Microbiome-Gut-Brain Axis and Toll-like Receptors in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 1689. [Google Scholar] [CrossRef]
- Tsunoda, S.M.; Gonzales, C.; Jarmusch, A.K.; Momper, J.D.; Ma, J.D. Contribution of the Gut Microbiome to Drug Disposition, Pharmacokinetic and Pharmacodynamic Variability. Clin. Pharmacokinet. 2021, 60, 971–984. [Google Scholar] [CrossRef]
- Haiser, H.J.; Gootenberg, D.B.; Chatman, K.; Sirasani, G.; Balskus, E.P.; Turnbaugh, P.J. Predicting and Manipulating Cardiac Drug Inactivation by the Human Gut Bacterium Eggerthella lenta. Science 2013, 341, 295–298. [Google Scholar] [CrossRef]
- Guo, Y.; Crnkovic, C.M.; Won, K.J.; Yang, X.; Lee, J.R.; Orjala, J.; Lee, H.; Jeong, H. Commensal Gut Bacteria Convert the Immunosuppressant Tacrolimus to Less Potent Metabolites. Drug Metab. Dispos. Biol. Fate Chem. 2019, 47, 194–202. [Google Scholar] [CrossRef]
- Lee, J.R.; Muthukumar, T.; Dadhania, D.; Taur, Y.; Jenq, R.R.; Toussaint, N.C.; Ling, L.; Pamer, E.; Suthanthiran, M. Gut microbiota and tacrolimus dosing in kidney transplantation. PLoS ONE 2015, 10, e0122399. [Google Scholar] [CrossRef] [PubMed]
- Claus Sandrine, P.; Ellero Sandrine, L.; Berger, B.; Krause, L.; Bruttin, A.; Molina, J.; Paris, A.; Want Elizabeth, J.; de Waziers, I.; Cloarec, O.; et al. Colonization-Induced Host-Gut Microbial Metabolic Interaction. mBio 2011, 2, 1–8. [Google Scholar] [CrossRef]
- Björkholm, B.; Bok, C.M.; Lundin, A.; Rafter, J.; Hibberd, M.L.; Pettersson, S. Intestinal Microbiota Regulate Xenobiotic Metabolism in the Liver. PLoS ONE 2009, 4, e6958. [Google Scholar] [CrossRef]
- van Kessel, S.P.; de Jong, H.R.; Winkel, S.L.; van Leeuwen, S.S.; Nelemans, S.A.; Permentier, H.; Keshavarzian, A.; El Aidy, S. Gut bacterial deamination of residual levodopa medication for Parkinson’s disease. BMC Biol. 2020, 18, 137. [Google Scholar] [CrossRef]
- Pierantozzi, M.; Pietroiusti, A.; Galante, A.; Sancesario, G.; Lunardi, G.; Fedele, E.; Giacomini, P.; Stanzione, P. Helicobacter pylori-induced reduction of acute levodopa absorption in Parkinson’s disease patients. Ann. Neurol. 2001, 50, 686–687. [Google Scholar] [CrossRef]
- Lee, W.Y.; Yoon, W.T.; Shin, H.Y.; Jeon, S.H.; Rhee, P.-L. Helicobacter pylori infection and motor fluctuations in patients with Parkinson’s disease. Mov. Disord. 2008, 23, 1696–1700. [Google Scholar] [CrossRef]
- Spencer, R.P. Intestinal absorption of amino acids. Current concepts. Am. J. Clin. Nutr. 1969, 22, 292–299. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.; Mo, C.; Liu, X.; Li, J.; Yan, Z.; Qian, Y.; Lai, Y.; Xu, S.; Yang, X.; et al. Association Between Microbial Tyrosine Decarboxylase Gene and Levodopa Responsiveness in Patients with Parkinson Disease. Neurology 2022, 99, e2443–e2453. [Google Scholar] [CrossRef]
- Menozzi, E.; Schapira, A.H.V. The Gut Microbiota in Parkinson Disease: Interactions with Drugs and Potential for Therapeutic Applications. CNS Drugs 2024, 38, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chengjun, M.; Penghui, A.; Xiaoqin, H.; Qin, X.; and Yang, X. Pharmacomicrobiomics: A new field contributing to optimizing drug therapy in Parkinson’s disease. Gut Microbes 2025, 17, 2454937. [Google Scholar] [CrossRef]
- Contin, M.; Martinelli, P. Pharmacokinetics of levodopa. J. Neurol. 2010, 257, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xiong, N.; Huang, J.; Guo, S.; Liu, L.; Han, C.; Zhang, G.; Jiang, H.; Ma, K.; Xia, Y.; et al. Protein-Restricted Diets for Ameliorating Motor Fluctuations in Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 206. [Google Scholar] [CrossRef]
- Warren Olanow, C.; Torti, M.; Kieburtz, K.; Leinonen, M.; Vacca, L.; Grassini, P.; Heller, A.; Heller, E.; Stocchi, F. Continuous versus intermittent oral administration of levodopa in Parkinson’s disease patients with motor fluctuations: A pharmacokinetics, safety, and efficacy study. Mov. Disord. 2019, 34, 425–429. [Google Scholar] [CrossRef]
- Leta, V.; Klingelhoefer, L.; Longardner, K.; Campagnolo, M.; Levent, H.Ç.; Aureli, F.; Metta, V.; Bhidayasiri, R.; Chung-Faye, G.; Falup-Pecurariu, C.; et al. Gastrointestinal barriers to levodopa transport and absorption in Parkinson’s disease. Eur. J. Neurol. 2023, 30, 1465–1480. [Google Scholar] [CrossRef]
- Fasano, A.; Bove, F.; Gabrielli, M.; Petracca, M.; Zocco, M.A.; Ragazzoni, E.; Barbaro, F.; Piano, C.; Fortuna, S.; Tortora, A.; et al. The role of small intestinal bacterial overgrowth in Parkinson’s disease. Mov. Disord. 2013, 28, 1241–1249. [Google Scholar] [CrossRef]
- Tan, A.H.; Mahadeva, S.; Thalha, A.M.; Gibson, P.R.; Kiew, C.K.; Yeat, C.M.; Ng, S.W.; Ang, S.P.; Chow, S.K.; Tan, C.T.; et al. Small intestinal bacterial overgrowth in Parkinson’s disease. Park. Relat. Disord. 2014, 20, 535–540. [Google Scholar] [CrossRef]
- Pierantozzi, M.; Pietroiusti, A.; Sancesario, G.; Lunardi, G.; Fedele, E.; Giacomini, P.; Frasca, S.; Galante, A.; Marciani, M.G.; Stanzione, P. Reduced L-dopa absorption and increased clinical fluctuations in Helicobacter pylori-infected Parkinson’s disease patients. Neurol. Sci. 2001, 22, 89–91. [Google Scholar] [CrossRef]
- Pierantozzi, M.; Pietroiusti, A.; Brusa, L.; Galati, S.; Stefani, A.; Lunardi, G.; Fedele, E.; Sancesario, G.; Bernardi, G.; Bergamaschi, A.; et al. Helicobacter pylori eradication and l-dopa absorption in patients with PD and motor fluctuations. Neurology 2006, 66, 1824–1829. [Google Scholar] [CrossRef]
- van Kessel, S.P.; Frye, A.K.; El-Gendy, A.O.; Castejon, M.; Keshavarzian, A.; van Dijk, G.; El Aidy, S. Gut bacterial tyrosine decarboxylases restrict levels of levodopa in the treatment of Parkinson’s disease. Nat. Commun. 2019, 10, 310. [Google Scholar] [CrossRef]
- Whitfield, A.C.; Moore, B.T.; Daniels, R.N. Classics in Chemical Neuroscience: Levodopa. ACS Chem. Neurosci. 2014, 5, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Flowers, S.A.; Bhat, S.; Lee, J.C. Potential Implications of Gut Microbiota in Drug Pharmacokinetics and Bioavailability. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2020, 40, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Nutt, J.G.; Carter, J.H.; Lea, E.S.; Sexton, G.J. Evolution of the response to levodopa during the first 4 years of therapy. Ann. Neurol. 2002, 51, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Sethi, K. Levodopa unresponsive symptoms in Parkinson disease. Mov. Disord. 2008, 23, S521–S533. [Google Scholar] [CrossRef]
- He, X.; Lai, Y.; Mo, C.; Zhang, Y.; Ai, P.; Xu, S.; Qian, Y.; Xiao, Q.; Yang, X. Association between Fecal Bile Acids and Levodopa Response in Patients with Parkinson’s Disease. Microorganisms 2024, 12, 1432. [Google Scholar] [CrossRef]
- Sampaio, T.F.; dos Santos, E.U.D.; de Lima, G.D.C.; dos Anjos, R.S.G.; da Silva, R.C.; Asano, A.G.C.; Asano, N.M.J.; Crovella, S.; de Souza, P.R.E. MAO-B and COMT Genetic Variations Associated with Levodopa Treatment Response in Patients with Parkinson’s Disease. J. Clin. Pharmacol. 2018, 58, 920–926. [Google Scholar] [CrossRef]
- Devos, D.; Lejeune, S.; Cormier-Dequaire, F.; Tahiri, K.; Charbonnier-Beaupel, F.; Rouaix, N.; Duhamel, A.; Sablonnière, B.; Bonnet, A.-M.; Bonnet, C.; et al. Dopa-decarboxylase gene polymorphisms affect the motor response to l-dopa in Parkinson’s disease. Park. Relat. Disord. 2014, 20, 170–175. [Google Scholar] [CrossRef]
- Moreau, C.; Meguig, S.; Corvol, J.-C.; Labreuche, J.; Vasseur, F.; Duhamel, A.; Delval, A.; Bardyn, T.; Devedjian, J.-C.; Rouaix, N.; et al. Polymorphism of the dopamine transporter type 1 gene modifies the treatment response in Parkinson’s disease. Brain 2015, 138, 1271–1283. [Google Scholar] [CrossRef]
- He, X.; Shi, K.; Mo, C.; Zhang, Y.; Xiao, Q.; Yang, X. Contribution of dopaminergic polymorphisms to levodopa treatment response and drug concentration in Chinese patients with Parkinson’s disease. Clin. Park. Relat. Disord. 2025, 12, 100333. [Google Scholar] [CrossRef]
- Koppel, N.; Maini Rekdal, V.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, eaag2770. [Google Scholar] [CrossRef]
- Spanogiannopoulos, P.; Bess, E.N.; Carmody, R.N.; Turnbaugh, P.J. The microbial pharmacists within us: A metagenomic view of xenobiotic metabolism. Nat. Rev. Microbiol. 2016, 14, 273–287. [Google Scholar] [CrossRef]
- Zhao, Z.; Ning, J.; Bao, X.-Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.-F.; Zhu, Y.-L.; Zhou, Z.-L.; Jia, X.-B.; Xu, Y.-D.; Yang, Q.; Cui, C.; Shen, Y.-Q. Neuroprotective effects of fecal microbiota transplantation on MPTP-induced Parkinson’s disease mice: Gut microbiota, glial reaction and TLR4/TNF-α signaling pathway. Brain Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Kuai, X.-Y.; Yao, X.-H.; Xu, L.-J.; Zhou, Y.-Q.; Zhang, L.-P.; Liu, Y.; Pei, S.-F.; Zhou, C.-L. Evaluation of fecal microbiota transplantation in Parkinson’s disease patients with constipation. Microb. Cell Factories 2021, 20, 98. [Google Scholar] [CrossRef]
- Xue, L.-J.; Yang, X.-Z.; Tong, Q.; Shen, P.; Ma, S.-J.; Wu, S.-N.; Zheng, J.-L.; Wang, H.-G. Fecal microbiota transplantation therapy for Parkinson’s disease: A preliminary study. Medicine 2020, 99, e22035. [Google Scholar] [CrossRef]
- Sadowski, K.; Zając, W.; Milanowski, Ł.; Koziorowski, D.; Figura, M. Exploring Fecal Microbiota Transplantation for Modulating Inflammation in Parkinson’s Disease: A Review of Inflammatory Markers and Potential Effects. Int. J. Mol. Sci. 2024, 25, 7741. [Google Scholar] [CrossRef]
- Domínguez Rojo, N.; Blanco Benítez, M.; Cava, R.; Fuentes, J.M.; Canales Cortés, S.; González Polo, R.A. Convergence of Neuroinflammation, Microbiota, and Parkinson’s Disease: Therapeutic Insights and Prospects. Int. J. Mol. Sci. 2024, 25, 11629. [Google Scholar] [CrossRef]
- Zhong, Z.; Ye, M.; Yan, F. A review of studies on gut microbiota and levodopa metabolism. Front. Neurol. 2023, 14, 1046910. [Google Scholar] [CrossRef]
- Daniel, S.E.; Lees, A.J. Parkinson’s Disease Society Brain Bank, London: Overview and research. J. Neural Transm. Suppl. 1993, 39, 165–172. [Google Scholar]
- Tomlinson, C.L.; Stowe, R.; Patel, S.; Rick, C.; Gray, R.; Clarke, C.E. Systematic review of levodopa dose equivalency reporting in Parkinson’s disease. Mov. Disord. 2010, 25, 2649–2653. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Beghini, F.; Mclver, L.J.; Blanco-Miguez, A.; Dubois, L.; Asnicar, F.; Maharjan, S.; Mailyan, A.; Manghi, P.; Scholz, M.; Thomas, A.M.; et al. Integrating taxonomic, functional, and strain-levelprofiling of diverse microbial communities with bioBakery 3. eLife 2021, 10, e65088. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Jiang, Y.; Wu, L.; Fu, J.; Du, J.; Luo, Z.; Guo, L.; Xu, J.; Liu, Y. Porphyromonas gingivalis aggravates colitis via a gut microbiota-linoleic acid metabolism-Th17/Treg cell balance axis. Nat. Commun. 2024, 15, 1617. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ai, P.; Xu, S.; Yuan, Y.; Xu, Z.; He, X.; Mo, C.; Zhang, Y.; Yang, X.; Xiao, Q. Targeted Gut Microbiota Modulation Enhances Levodopa Bioavailability and Motor Recovery in MPTP Parkinson’s Disease Models. Int. J. Mol. Sci. 2025, 26, 5282. https://doi.org/10.3390/ijms26115282
Ai P, Xu S, Yuan Y, Xu Z, He X, Mo C, Zhang Y, Yang X, Xiao Q. Targeted Gut Microbiota Modulation Enhances Levodopa Bioavailability and Motor Recovery in MPTP Parkinson’s Disease Models. International Journal of Molecular Sciences. 2025; 26(11):5282. https://doi.org/10.3390/ijms26115282
Chicago/Turabian StyleAi, Penghui, Shaoqing Xu, Yuan Yuan, Ziqi Xu, Xiaoqin He, Chengjun Mo, Yi Zhang, Xiaodong Yang, and Qin Xiao. 2025. "Targeted Gut Microbiota Modulation Enhances Levodopa Bioavailability and Motor Recovery in MPTP Parkinson’s Disease Models" International Journal of Molecular Sciences 26, no. 11: 5282. https://doi.org/10.3390/ijms26115282
APA StyleAi, P., Xu, S., Yuan, Y., Xu, Z., He, X., Mo, C., Zhang, Y., Yang, X., & Xiao, Q. (2025). Targeted Gut Microbiota Modulation Enhances Levodopa Bioavailability and Motor Recovery in MPTP Parkinson’s Disease Models. International Journal of Molecular Sciences, 26(11), 5282. https://doi.org/10.3390/ijms26115282