
The Molecular and Cellular Basis of Physiological Changes in Pregnancy and Its Implications in Neurologic and Ophthalmic Pathologies




Abstract
1. Introduction
2. Methods: Literature Review and Search Strategy
3. The Molecular and Cellular Basis of Physiological Changes in Pregnancy
3.1. Antepartum Neuroanatomical Changes in Mothers
3.2. Postpartum Neuroanatomical Changes in Mothers
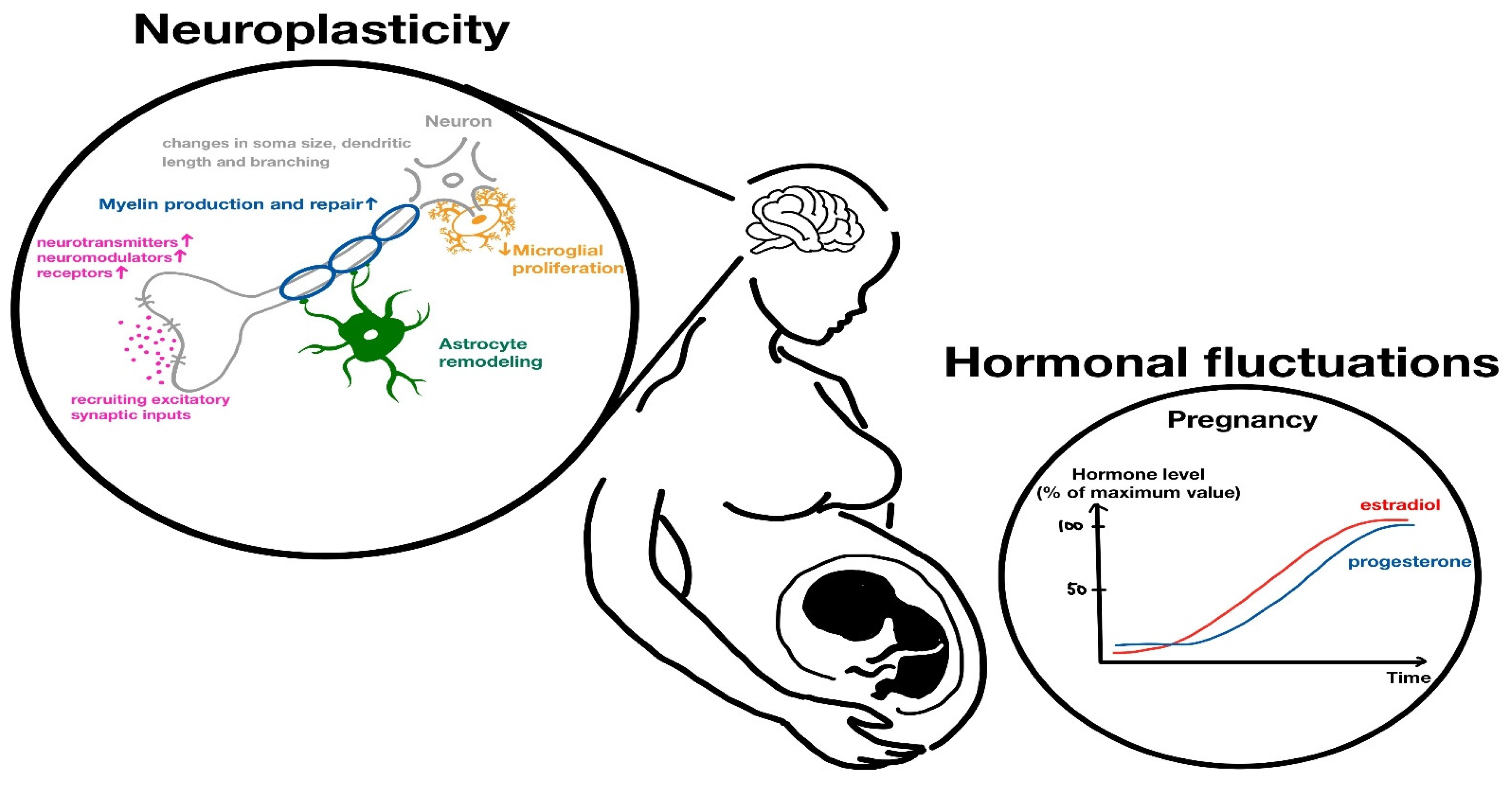
3.3. The Role of Pregnancy Hormones in Anatomy
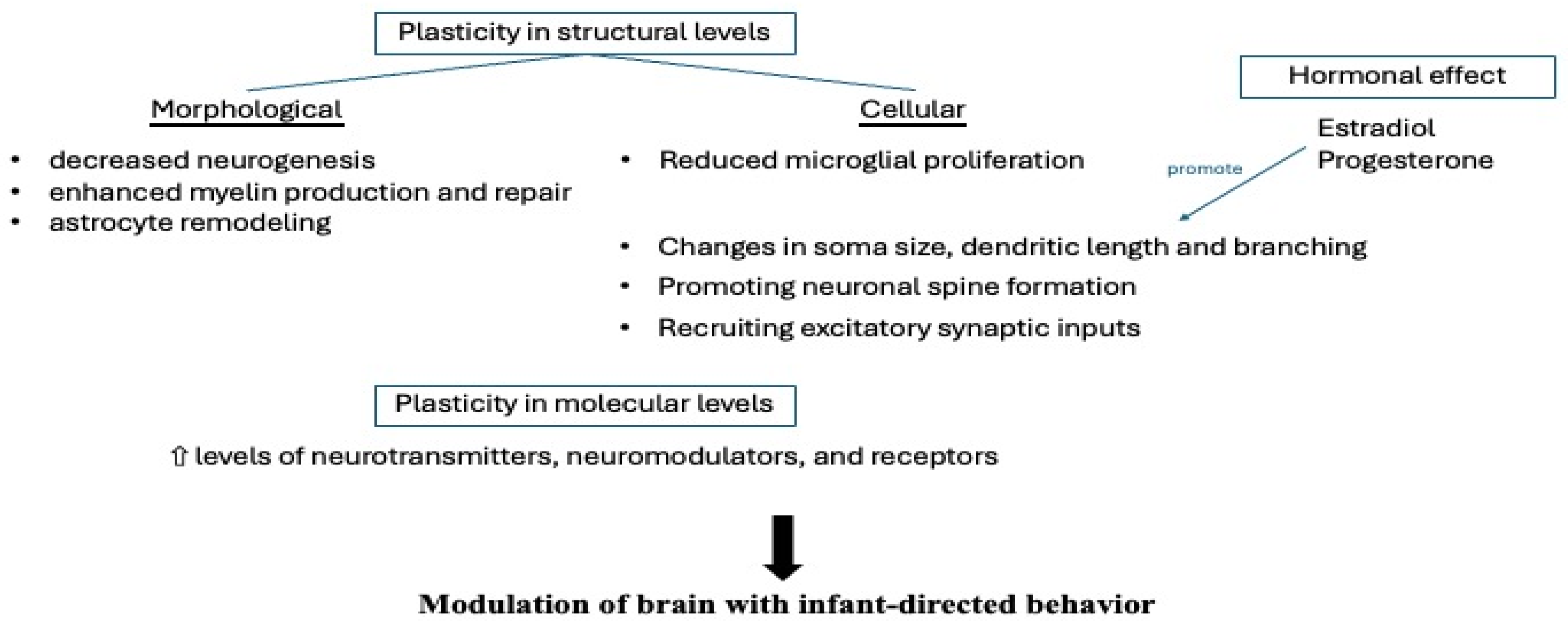
3.4. Hormonal-Mediated Neuroplasticity
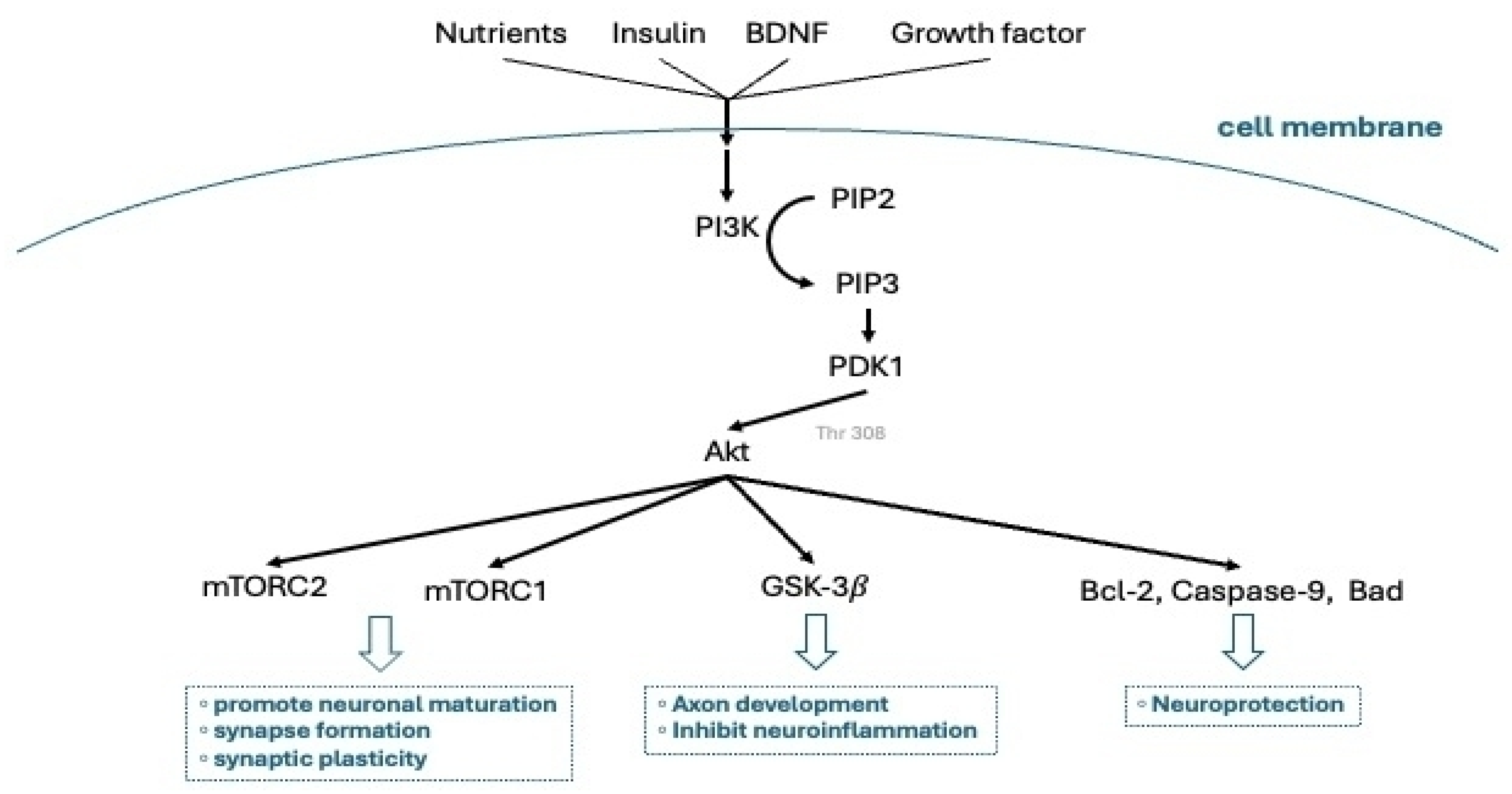
3.5. Neurogenesis
4. The Implications of Pregnant Physiological Changes in Neurologic and Ophthalmic Pathologies
4.1. Vascular Change—Hypercoagulable Status
4.2. Vascular Event—Cerebral Stroke
4.3. Pituitary Gland and Pituitary Adenoma
4.4. Meningioma
4.5. Preeclampsia and Eclampsia
4.6. Papilledema
4.7. Migraine
4.8. Cranial Neuropathy
4.9. Diabetic Retinopathy
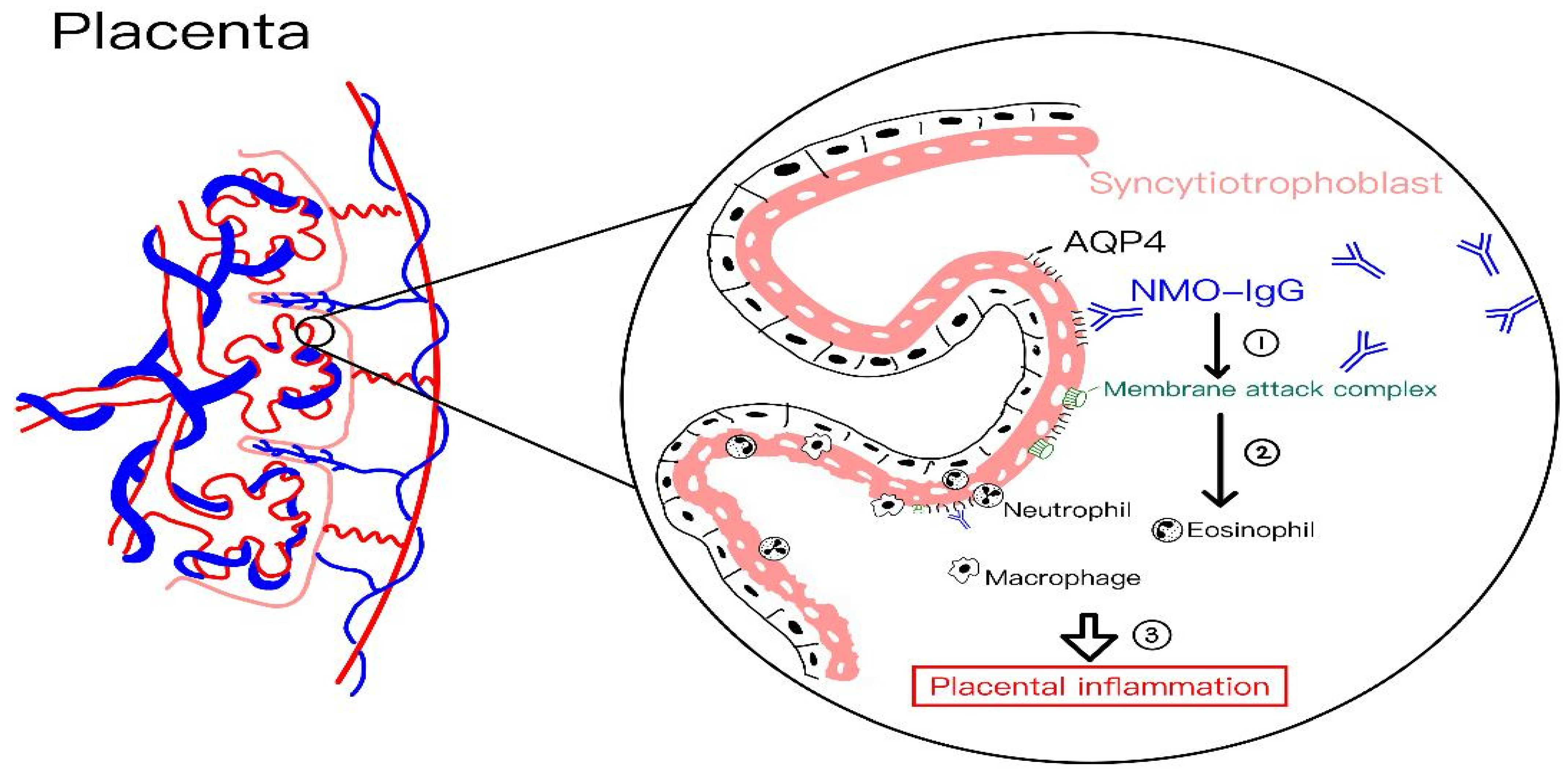
4.10. Neuromyelitis Optica Spectrum Disorder (NMOSD)
4.11. Multiple Sclerosis (MS)
4.12. Graves’ Disease and Thyroid Orbitopathy
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Carmona, S.; Martínez-García, M.; Paternina-Die, M.; Barba-Müller, E.; Wierenga, L.M.; Alemán-Gómez, Y.; Pretus, C.; Marcos-Vidal, L.; Beumala, L.; Cortizo, R.; et al. Pregnancy and Adolescence Entail Similar Neuroanatomical Adaptations: A Comparative Analysis of Cerebral Morphometric Changes. Hum. Brain Mapp. 2019, 40, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Hoekzema, E.; Barba-Müller, E.; Pozzobon, C.; Picado, M.; Lucco, F.; García-García, D.; Soliva, J.C.; Tobeña, A.; Desco, M.; A Crone, E.; et al. Pregnancy Leads to Long-Lasting Changes in Human Brain Structure. Nat. Neurosci. 2017, 20, 287–296. [Google Scholar] [CrossRef]
- Servin-Barthet, C.; Martínez-García, M.; Pretus, C.; Paternina-Die, M.; Soler, A.; Khymenets, O.; Pozo, Ó.J.; Leuner, B.; Vilarroya, O.; Carmona, S. The Transition to Motherhood: Linking Hormones, Brain and Behaviour. Nat. Rev. Neurosci. 2023, 24, 605–619. [Google Scholar] [CrossRef] [PubMed]
- Pritschet, L.; Taylor, C.M.; Cossio, D.; Faskowitz, J.; Santander, T.; Handwerker, D.A.; Grotzinger, H.; Layher, E.; Chrastil, E.R.; Jacobs, E.G. Neuroanatomical Changes Observed over the Course of a Human Pregnancy. Nat. Neurosci. 2024, 27, 2253–2260. [Google Scholar] [CrossRef]
- Hoekzema, E.; Tamnes, C.K.; Berns, P.; Barba-Müller, E.; Pozzobon, C.; Picado, M.; Lucco, F.; Martínez-García, M.; Desco, M.; Ballesteros, A.; et al. Becoming a Mother Entails Anatomical Changes in the Ventral Striatum of the Human Brain That Facilitate Its Responsiveness to Offspring Cues. Psychoneuroendocrinology 2020, 112, 104507. [Google Scholar] [CrossRef]
- Luders, E.; Kurth, F.; Gingnell, M.; Engman, J.; Yong, E.-L.; Poromaa, I.S.; Gaser, C. From Baby Brain to Mommy Brain: Widespread Gray Matter Gain after Giving Birth. Cortex 2020, 126, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Lisofsky, N.; Gallinat, J.; Lindenberger, U.; Kühn, S. Postpartal Neural Plasticity of the Maternal Brain: Early Renormalization of Pregnancy-Related Decreases? Neurosignals 2019, 27, 12–24. [Google Scholar] [CrossRef]
- Hoekzema, E.; van Steenbergen, H.; Straathof, M.; Beekmans, A.; Freund, I.M.; Pouwels, P.J.W.; Crone, E.A. Mapping the Effects of Pregnancy on Resting State Brain Activity, White Matter Microstructure, Neural Metabolite Concentrations and Grey Matter Architecture. Nat. Commun. 2022, 13, 6931. [Google Scholar] [CrossRef]
- Chechko, N.; Dukart, J.; Tchaikovski, S.; Enzensberger, C.; Neuner, I.; Stickel, S. The Expectant Brain—Pregnancy Leads to Changes in Brain Morphology in the Early Postpartum Period. Cereb. Cortex 2022, 32, 4025–4038. [Google Scholar] [CrossRef]
- Kim, P.; Leckman, J.F.; Mayes, L.C.; Feldman, R.; Wang, X.; Swain, J.E. The Plasticity of Human Maternal Brain: Longitudinal Changes in Brain Anatomy during the Early Postpartum Period. Behav. Neurosci. 2010, 124, 695–700. [Google Scholar] [CrossRef]
- Ammari, R.; Monaca, F.; Cao, M.; Nassar, E.; Wai, P.; Del Grosso, N.A.; Lee, M.; Borak, N.; Schneider-Luftman, D.; Kohl, J. Hormone-Mediated Neural Remodeling Orchestrates Parenting Onset during Pregnancy. Science 2023, 382, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Kohl, J.; Babayan, B.M.; Rubinstein, N.D.; Autry, A.E.; Marin-Rodriguez, B.; Kapoor, V.; Miyamishi, K.; Zweifel, L.S.; Luo, L.; Uchida, N.; et al. Functional Circuit Architecture Underlying Parental Behaviour. Nature 2018, 556, 326–331. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Musatov, S.; Shteyler, A.; Simanduyev, S.; Arrieta-Cruz, I.; Ogawa, S.; Pfaff, D.W. siRNA Silencing of Estrogen Receptor-α Expression Specifically in Medial Preoptic Area Neurons Abolishes Maternal Care in Female Mice. Proc. Natl. Acad. Sci. USA 2012, 109, 16324–16329. [Google Scholar] [CrossRef]
- Brown, R.S.E.; Aoki, M.; Ladyman, S.R.; Phillipps, H.R.; Wyatt, A.; Boehm, U.; Grattan, D.R. Prolactin Action in the Medial Preoptic Area Is Necessary for Postpartum Maternal Nursing Behavior. Proc. Natl. Acad. Sci. USA 2017, 114, 10779–10784. [Google Scholar] [CrossRef] [PubMed]
- Numan, M. The Parental Brain: Mechanisms, Development, and Evolution; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Keyser-Marcus, L.; Stafisso-Sandoz, G.; Gerecke, K.; Jasnow, A.; Nightingale, L.; Lambert, K.G.; Gatewood, J.; Kinsley, C.H. Alterations of Medial Preoptic Area Neurons Following Pregnancy and Pregnancy-like Steroidal Treatment in the Rat. Brain Res. Bull. 2001, 55, 737–745. [Google Scholar] [CrossRef]
- Uriarte, N.; Ferreño, M.; Méndez, D.; Nogueira, J. Reorganization of Perineuronal Nets in the Medial Preoptic Area During the Reproductive Cycle in Female Rats. Sci. Rep. 2020, 10, 5479. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Sabihi, S. The Birth of New Neurons in the Maternal Brain: Hormonal Regulation and Functional Implications. Front. Neuroendocrinol. 2016, 41, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Shingo, T.; Gregg, C.; Enwere, E.; Fujikawa, H.; Hassam, R.; Geary, C.; Cross, J.C.; Weiss, S. Pregnancy-Stimulated Neurogenesis in the Adult Female Forebrain Mediated by Prolactin. Science 2003, 299, 117–120. [Google Scholar] [CrossRef]
- Eid, R.S.; Chaiton, J.A.; Lieblich, S.E.; Bodnar, T.S.; Weinberg, J.; Galea, L.A.M. Early and Late Effects of Maternal Experience on Hippocampal Neurogenesis, Microglia, and the Circulating Cytokine Milieu. Neurobiol. Aging 2019, 78, 1–17. [Google Scholar] [CrossRef]
- Pawluski, J.L.; Paravatou, R.; Even, A.; Cobraiville, G.; Fillet, M.; Kokras, N.; Dalla, C.; Charlier, T.D. Effect of Sertraline on Central Serotonin and Hippocampal Plasticity in Pregnant and Non-Pregnant Rats. Neuropharmacology 2020, 166, 107950. [Google Scholar] [CrossRef]
- Pawluski, J.L.; Galea, L.A. Reproductive Experience Alters Hippocampal Neurogenesis During the Postpartum Period in the Dam. Neuroscience 2007, 149, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Mirescu, C.; Noiman, L.; Gould, E. Maternal Experience Inhibits the Production of Immature Neurons in the Hippocampus During the Postpartum Period Through Elevations in Adrenal Steroids. Hippocampus 2007, 17, 434–442. [Google Scholar] [CrossRef]
- Darnaudéry, M.; Perez-Martin, M.; Del Favero, F.; Gomez-Roldan, C.; Garcia-Segura, L.M.; Maccari, S. Early Motherhood in Rats Is Associated with a Modification of Hippocampal Function. Psychoneuroendocrinology 2007, 32, 803–812. [Google Scholar] [CrossRef]
- Hillerer, K.M.; Neumann, I.D.; Couillard-Despres, S.; Aigner, L.; Slattery, D.A. Lactation-Induced Reduction in Hippocampal Neurogenesis Is Reversed by Repeated Stress Exposure. Hippocampus 2014, 24, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Workman, J.L.; Gobinath, A.R.; Kitay, N.F.; Chow, C.; Brummelte, S.; Galea, L.A.M. Parity Modifies the Effects of Fluoxetine and Corticosterone on Behavior, Stress Reactivity, and Hippocampal Neurogenesis. Neuropharmacology 2016, 105, 443–453. [Google Scholar] [CrossRef]
- Szecsi, P.B.; Jørgensen, M.; Klajnbard, A.; Andersen, M.R.; Colov, N.P.; Stender, S. Haemostatic Reference Intervals in Pregnancy. Thromb. Haemost. 2010, 103, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Digre, K.B.; Kinard, K. Neuro-Ophthalmic Disorders in Pregnancy. Continuum 2014, 20, 162–176. [Google Scholar] [CrossRef]
- Yenerel, N.M.; Küçümen, R.B. Pregnancy and the Eye. Turk. J. Ophthalmol. 2015, 45, 213–219. [Google Scholar] [CrossRef]
- Anton, N.; Bogdănici, C.M.; Branișteanu, D.C.; Armeanu, T.; Ilie, O.D.; Doroftei, B. A Narrative Review on Neuro-Ophthalmological Manifestations That May Occur During Pregnancy. Life 2024, 14, 431. [Google Scholar] [CrossRef]
- Digre, K.B. Neuro-Ophthalmology and Pregnancy: What Does a Neuro-Ophthalmologist Need to Know? J. Neuroophthalmol. 2011, 31, 381–387. [Google Scholar] [CrossRef]
- Rantanen, K.; Tatlisumak, T. Stroke in Women—Oral Contraception, Pregnancy, and Hormone Replacement Therapy. Curr. Vasc. Pharmacol. 2013, 11, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Pařízek, A.; Koucký, M.; Dušková, M. Progesterone, Inflammation and Preterm Labor. J. Steroid Biochem. Mol. Biol. 2014, 139, 159–165. [Google Scholar] [CrossRef]
- Gilbert, A.L.; Prasad, S.; Mallery, R.M. Neuro-Ophthalmic Disorders in Pregnancy. Neurol. Clin. 2019, 37, 85–102. [Google Scholar] [CrossRef]
- Wang, N.; Shen, X.; Zhang, G.; Gao, B.; Lerner, A. Cerebrovascular Disease in Pregnancy and Puerperium: Perspectives from Neuroradiologists. Quant. Imaging Med. Surg. 2021, 11, 838–851. [Google Scholar] [CrossRef] [PubMed]
- Grear, K.E.; Bushnell, C.D. Stroke and Pregnancy: Clinical Presentation, Evaluation, Treatment, and Epidemiology. Clin. Obstet. Gynecol. 2013, 56, 350–359. [Google Scholar] [CrossRef]
- Zoli, M.; Guaraldi, F.; Zoia, C.; La Corte, E.; Asioli, S.; Bongetta, D.; Rustici, A.; Mazzatenta, D. Management of Sellar and Parasellar Tumors Becoming Symptomatic During Pregnancy: A Practical Algorithm Based on Multi-Center Experience and Systematic Literature Review. Pituitary 2021, 24, 269–283. [Google Scholar] [CrossRef]
- Dinç, H.; Esen, F.; Demirci, A.; Sari, A.; Resit Gümele, H. Pituitary Dimensions and Volume Measurements in Pregnancy and Post Partum. MR Assessment. Acta Radiol. 1998, 39, 64–69. [Google Scholar] [CrossRef]
- Inoue, T.; Hotta, A.; Awai, M.; Tanihara, H. Loss of Vision Due to a Physiologic Pituitary Enlargement During Normal Pregnancy. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 1049–1051. [Google Scholar] [CrossRef] [PubMed]
- Bachmeier, C.A.E.; Snell, C.; Morton, A. Visual Loss in Pregnancy. BMJ Case Rep. 2019, 12, e228323. [Google Scholar] [CrossRef]
- Lee, H.R.; Song, J.E.; Lee, K.Y. Developed diplopia and ptosis due to a nonfunctioning pituitary macroadenoma during pregnancy. Obstet. Gynecol. Sci. 2014, 57, 66–69. [Google Scholar] [CrossRef]
- Lambert, K.; Williamson, C. Review of Presentation, Diagnosis and Management of Pituitary Tumours in Pregnancy. Obstet. Med. 2013, 6, 13–19. [Google Scholar] [CrossRef]
- Laviv, Y.; Ohla, V.; Kasper, E.M. Unique features of pregnancy-related meningiomas: Lessons learned from 148 reported cases and theoretical implications of a prolactin modulated pathogenesis. Neurosurg. Rev. 2018, 41, 95–108. [Google Scholar] [CrossRef]
- Hortobágyi, T.; Bencze, J.; Murnyák, B.; Kouhsari, M.C.; Bognár, L.; Marko-Varga, G. Pathophysiology of meningioma growth in pregnancy. Open Med. 2017, 12, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Chow, M.S.; Mercier, P.A.; Omahen, D.A.; Wood, S.L.; Johnson, J.-A.M. Recurrent Exophytic Meningioma in Pregnancy. Obstet. Gynecol. 2013, 121, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.S.; Choi, E.Y.; Lee, M.; Kim, H.; Chung, H. Serous retinal detachment in preeclampsia and malignant hypertension. Eye 2019, 33, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Moss, H.E. Neuro-ophthalmology and Pregnancy. Continuum 2022, 28, 147–161. [Google Scholar] [CrossRef]
- Euser, A.G.; Cipolla, M.J. Magnesium sulfate for the treatment of eclampsia: A brief review. Stroke 2009, 40, 1169–1175. [Google Scholar] [CrossRef]
- Crowther, C.A.; Hiller, J.E.; Doyle, L.W.; Haslam, R.R.; Group for the ACToMSC. Effect of Magnesium Sulfate Given for Neuroprotection Before Preterm Birth: A Randomized Controlled Trial. JAMA 2003, 290, 2669–2676. [Google Scholar] [CrossRef]
- Digre, K.B.; Varner, M.W.; Schiffman, J.S. Neuroophthalmologic effects of intravenous magnesium sulfate. Am. J. Obstet. Gynecol. 1990, 163 Pt 1, 1848–1852. [Google Scholar] [CrossRef]
- Bain, E.S.; Middleton, P.F.; Crowther, C.A. Maternal adverse effects of different antenatal magnesium sulphate regimens for improving maternal and infant outcomes: A systematic review. BMC Pregnancy Childbirth 2013, 13, 195. [Google Scholar] [CrossRef]
- Broner, S.W.; Bobker, S.; Klebanoff, L. Migraine in Women. Semin. Neurol. 2017, 37, 601–610. [Google Scholar] [CrossRef]
- Frederick, I.O.; Qiu, C.; Enquobahrie, D.A.; Aurora, S.K.; Peterlin, B.L.; Gelaye, B.; Williams, M.A. Lifetime prevalence and correlates of migraine among women in a pacific northwest pregnancy cohort study. Headache 2014, 54, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Chauvel, V.; Schoenen, J.; Multon, S. Influence of ovarian hormones on cortical spreading depression and its suppression by L-kynurenine in rat. PLoS ONE 2013, 8, e82279. [Google Scholar] [CrossRef]
- Vrabec, J.T.; Isaacson, B.; Van Hook, J.W. Bell’s palsy and pregnancy. Otolaryngol. Head. Neck Surg. 2007, 137, 858–861. [Google Scholar] [CrossRef]
- Evangelista, V.; Gooding, M.S.; Pereira, L. Bell’s Palsy in Pregnancy. Obstet. Gynecol. Surv. 2019, 74, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.R.; Madanagopalan, V.G.; Narayanan, R. Diabetic retinopathy in pregnancy—A review. Indian. J. Ophthalmol. 2021, 69, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Rahman, W.; Rahman, F.Z.; Yassin, S.; Al-Suleiman, S.A.; Rahman, J. Progression of retinopathy during pregnancy in type 1 diabetes mellitus. Clin. Exp. Ophthalmol. 2007, 35, 231–236. [Google Scholar] [CrossRef]
- The Diabetes Control and Complications Trial Research Group. Effect of pregnancy on microvascular complications in the diabetes control and complications trial. Diabetes Care 2000, 23, 1084–1091. [Google Scholar] [CrossRef]
- Sheth, B.P. Does pregnancy accelerate the rate of progression of diabetic retinopathy?: An update. Curr. Diab. Rep. 2008, 8, 270–273. [Google Scholar] [CrossRef]
- Shumway, C.L.; Patel, B.C.; Tripathy, K.; De Jesus, O. Neuromyelitis Optica Spectrum Disorder (NMOSD). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Jacob, A.; McKeon, A.; Nakashima, I.; Sato, D.K.; Elsone, L.; Fujihara, K.; de Seze, J. Current concept of neuromyelitis optica (NMO) and NMO spectrum disorders. J. Neurol. Neurosurg. Psychiatry 2013, 84, 922–930. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin 4 and neuromyelitis optica. Lancet Neurol. 2012, 11, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Pandit, L.; Asgari, N.; Apiwattanakul, M.; Palace, J.; Paul, F.; Leite, M.; Kleiter, I.; Chitnis, T.; the GJCF International Clinical Consortium & Biorepository for Neuromyelitis Optica. Demographic and clinical features of neuromyelitis optica: A review. Mult. Scler. 2015, 21, 845–853. [Google Scholar] [CrossRef]
- De Falco, M.; Cobellis, L.; Torella, M.; Acone, G.; Varano, L.; Sellitti, A.; Ragucci, A.; Coppola, G.; Cassandro, R.; Laforgia, V.; et al. Down-regulation of aquaporin 4 in human placenta throughout pregnancy. In Vivo 2007, 21, 813–817. [Google Scholar]
- Saadoun, S.; Waters, P.; Leite, M.I.; Bennett, J.L.; Vincent, A.; Papadopoulos, M.C. Neuromyelitis optica IgG causes placental inflammation and fetal death. J. Immunol. 2013, 191, 2999–3005. [Google Scholar] [CrossRef]
- Klawiter, E.C.; Bove, R.; Elsone, L.; Alvarez, E.; Borisow, N.; Cortez, M.; Mateen, F.; Mealy, M.A.; Sorum, J.; Mutch, K.; et al. High risk of postpartum relapses in neuromyelitis optica spectrum disorder. Neurology 2017, 89, 2238–2244. [Google Scholar] [CrossRef]
- Fragoso, Y.D.; Adoni, T.; Bichuetti, D.B.; Brooks, J.B.B.; Ferreira, M.L.B.; Oliveira, E.M.L.; Oliveira, C.L.S.; Ribeiro, S.B.F.; Silva, A.E.; Siquineli, F. Neuromyelitis optica and pregnancy. J. Neurol. 2013, 260, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Bourre, B.; Marignier, R.; Zephir, H.; Papeix, C.; Brassat, D.; Castelnovo, G.; Collongues, N.; Vukusic, S.; Labauge, P.; Outteryck, O.; et al. Neuromyelitis optica and pregnancy. Neurology 2012, 78, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Kim, S.-H.; Nakashima, I.; Takai, Y.; Fujihara, K.; Leite, M.; Kitley, J.; Palace, J.; Santos, E.; Coutinho, E.; et al. Influence of pregnancy on neuromyelitis optica spectrum disorder. Neurology 2012, 78, 1264–1267. [Google Scholar] [CrossRef]
- Koskela, E.; Soinne, L.; Valanne, L.; Setälä, K. Pregnancy Associated Ischaemic Optic Neuropathy. Neuro-Ophthalmology 2011, 35, 202–206. [Google Scholar] [CrossRef]
- Pozzilli, C.; Pugliatti, M. An overview of pregnancy-related issues in patients with multiple sclerosis. Eur. J. Neurol. 2015, 22 (Suppl. S2), 34–39. [Google Scholar] [CrossRef]
- Hughes, S.E.; Spelman, T.; Gray, O.M.; Boz, C.; Trojano, M.; Lugaresi, A.; Izquierdo, G.; Duquette, P.; Girard, M.; Grand’maison, F.; et al. Predictors and dynamics of postpartum relapses in women with multiple sclerosis. Mult. Scler. 2014, 20, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Finkelsztejn, A.; Brooks, J.B.; Paschoal, F.M., Jr.; Fragoso, Y.D. What can we really tell women with multiple sclerosis regarding pregnancy? A systematic review and meta-analysis of the literature. Bjog 2011, 118, 790–797. [Google Scholar] [CrossRef]
- Alwan, S.; Sadovnick, A.D. Multiple sclerosis and pregnancy: Maternal considerations. Womens Health 2012, 8, 399–414. [Google Scholar] [CrossRef]
- Lazarus, J.H. Thyroid function in pregnancy. Br. Med. Bull. 2011, 97, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Brent, G.A. Graves’ Disease. N. Engl. J. Med. 2008, 358, 2594–2605. [Google Scholar] [CrossRef]
- Aagaard-Tillery, K.M.; Silver, R.; Dalton, J. Immunology of normal pregnancy. Semin. Fetal Neonatal Med. 2006, 11, 279–295. [Google Scholar] [CrossRef]
- Sykes, L.; MacIntyre, D.A.; Yap, X.J.; Ponnampalam, S.; Teoh, T.G.; Bennett, P.R. Changes in the Th1:Th2 cytokine bias in pregnancy and the effects of the anti-inflammatory cyclopentenone prostaglandin 15-deoxy-Δ(12,14)-prostaglandin J2. Mediat. Inflamm. 2012, 2012, 416739. [Google Scholar] [CrossRef]
- Bucci, I.; Giuliani, C.; Napolitano, G. Thyroid-Stimulating Hormone Receptor Antibodies in Pregnancy: Clinical Relevance. Front. Endocrinol. 2017, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Wall, J.R.; Lahooti, H.; Hibbert, E.J.; Champion, B. Relationship Between Clinical and Immunological Features of Thyroid Autoimmunity and Ophthalmopathy During Pregnancy. J. Thyroid Res. 2015, 2015, 698470. [Google Scholar] [CrossRef]
- Gopinath, B.; Musselman, R.; Adams, C.L.; Tani, J.; Beard, N.; Wall, J.R. Study of Serum Antibodies against Three Eye Muscle Antigens and the Connective Tissue Antigen Collagen XIII in Patients with Graves’ Disease with and without Ophthalmopathy: Correlation with Clinical Features. Thyroid 2006, 16, 967–974. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Region | Observed Changes During Pregnancy | Clinical and Functional Implications | Observed Changes Postpartum |
|---|---|---|---|
| Cortical gray matter |
|
|
|
| White matter |
|
| Returning to baseline levels |
| Ophthalmic Disorders | Disease Activity During Pregnancy | Underlying Mechanisms | Effects in Neuro-Ophthalmic Symptoms |
|---|---|---|---|
| Pituitary adenoma | Enlargement of pre-existing adenomas | 1. Lactotroph hyperplasia driven by estrogen and progesterone; 2. Estrogen receptor expression of the pituitary adenoma. | 1. Visual disturbances resulting from optic chiasm compression; 2. Headache. |
| Meningioma | More aggressive behavior | 1. Estrogen/progesterone-driven tumor growth; 2. VEGF-mediated vascularization; 3. Predominantly parasellar localization and clear cell or chordoid subtypes. | 1. Optic nerve compression; 2. Visual impairment. |
| Migraine | 1. Generally improved; 2. Aura worsened in subset patients. | 1. Estrogen fluctuations altering cortical excitability; 2. Cortical spreading depression. | 1. Reduced migraine frequency overall; 2. Increased aua frequency in subset of patients. |
| Diabetic retinopathy | Progression | 1. Increased cardiac output and impaired autoregulation causing vascular dilation; 2. Placental hormones (IGF, placental GH) induced angiogenesis; 3. Enhanced inflammation. | 1. Progression from non-proliferative to proliferative retinopathy; 2. Increased diabetic macular edema. |
| Neuromyelitis optica spectrum disorder | Increased disease activity | 1. High placental expression of AQP4; 2. Anti-AQP4 antibody-mediated placental inflammation. | 1. Exacerbations of optic neuritis and transverse myelitis; 2. Increased miscarriage risk; 3. Increased relapse postpartum. |
| Multiple sclerosis | Decreased disease activity | 1. Immunosuppressive Th2 dominance; 2. Postpartum immunologic rebound. | 1. Reduced optic neuritis attacks during pregnancy; 2. Increased postpartum relapse frequency. |
| Graves’ disease and thyroid orbitopathy | Initial aggravation; subsequent improvement | 1. Pregnancy-induced immune modulation; 2. hCG-induced thyroid stimulation; 3. Fluctuating thyroid receptor antibody (TRAb) levels. | Inconsistent association with the severity of ocular symptoms |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiang, Y.-T.; Chen, J.-H.; Chen, K.-H. The Molecular and Cellular Basis of Physiological Changes in Pregnancy and Its Implications in Neurologic and Ophthalmic Pathologies. Int. J. Mol. Sci. 2025, 26, 5220. https://doi.org/10.3390/ijms26115220
Chiang Y-T, Chen J-H, Chen K-H. The Molecular and Cellular Basis of Physiological Changes in Pregnancy and Its Implications in Neurologic and Ophthalmic Pathologies. International Journal of Molecular Sciences. 2025; 26(11):5220. https://doi.org/10.3390/ijms26115220
Chicago/Turabian StyleChiang, Yi-Ting, Jie-Hong Chen, and Kuo-Hu Chen. 2025. "The Molecular and Cellular Basis of Physiological Changes in Pregnancy and Its Implications in Neurologic and Ophthalmic Pathologies" International Journal of Molecular Sciences 26, no. 11: 5220. https://doi.org/10.3390/ijms26115220
APA StyleChiang, Y.-T., Chen, J.-H., & Chen, K.-H. (2025). The Molecular and Cellular Basis of Physiological Changes in Pregnancy and Its Implications in Neurologic and Ophthalmic Pathologies. International Journal of Molecular Sciences, 26(11), 5220. https://doi.org/10.3390/ijms26115220