
Endometriosis: An Immunologist’s Perspective

 , , and
, , and 
Abstract
1. Introduction
2. Endometriosis
2.1. Genetic and Epigenetic Changes in Endometriosis
2.2. Extracellular Vesicles
2.3. Microbiota and Endometriosis
3. Immune Response in Endometriosis
3.1. Pattern-Recognition Receptors (PRRs), Pathogen-Associated Molecular Patterns (PAMPs), Damage-Associated Molecular Patterns (DAMPs), and Endometriosis
3.2. Innate Immune Response in Endometriosis

{kind=link}
| Component | Characteristics | Ref |
|---|---|---|
| Complement pathway | Increased expression of the protein components of the pathway in human endometriosis. | [173] |
| Increased levels of C1q, C1 inhibitor, mannose-binding lectin (MBL), C3c, C4, and the membrane attack complex (SC5b-9) in the peritoneal fluid of endometriosis patients. Increased expression of 1QA, C1QB, C1R, C1S, C2, C3, C4A/B, C5, C6, C7, C8A, CFB, CFH, and CFI in ectopic endometrium. | [174] | |
| C1q levels are correlated with vessel formation in endometriosis (human). | [175] | |
| The lectin pathway may not be involved in endometriosis in humans. | [176] | |
| Defensins | There are no changes in defensin levels in women with endometriosis. | [177] |
| Neutrophils | Increased levels of human neutrophil peptides 1, 2, and 3 have been observed in the endometrial fluid of women with endometriosis. | [178] |
| In endometriosis, neutrophil phagocytosis is impaired. Neutrophils support the survival of endometrial cells and help create a microenvironment conducive to the development and growth of lesions (mouse and human). | [179,180,181] | |
| Neutrophil depletion in mice reduces the formation of endometriotic lesions. | [181] | |
| Macrophages | Elevated IL-8, C-C chemokine RANTES (CCL5), MCP-1, and MIF attracted more cells in advanced endometriosis lesions in humans and mice. | [182] |
| There are different types of macrophages present in endometriomas (mouse model). | [183] | |
| High iron levels in the peritoneal fluid impair the phagocytic response and increase oxygen radical formation (human). | [173,179,184] | |
| Extracellular vesicles modulate macrophage response in endometriosis. | [185,186] | |
| The expression of CD36 in macrophages is inhibited by the high concentration of PGE2 in the endometrioma (human). | [187] | |
| An increased expression of CD200 correlates with reduced phagocytic activity and decreased CD36 expression in endometriosis (human). | [188] | |
| TLR4 and RAGE expression in peritoneal fluid macrophages inversely correlate with endometriosis severity (human). | [189] | |
| Macrophages play a vital role in both fibrosis and mesenchymal transdifferentiation (in humans and mice). | [190,191] | |
| NK cells | There is a higher density of CD56 in uNK cells in patients with endometriosis undergoing IVF treatment. | [159,160,161] |
| Uterine NK cell amounts are higher in patients with endometriosis. | [159,161,192] | |
| There is a decrease in tissue immature CD56 cells following the surgical removal of endometriomas (human). | [159,193] | |
| NK cell cytotoxic activity is significantly reduced in women with moderate to severe endometriosis (peripheral, peritoneal, and uNK). | [194,195] | |
| Granzyme B and perforin secretion were reduced in NK cells from endometriosis patients. | [195,196] | |
| The increase in soluble MICA/B levels in the peritoneal fluid of patients with endometriosis negatively affects the cytotoxic function of NK cells. | [196] | |
| Elevated levels of IL-6 and TGF-β1 in the peritoneal fluid of endometriosis patients are responsible for the impaired cytotoxic activity of NK cells. | [197,198] | |
| High IL-15 levels produced by ectopic endometrial stromal cells can inhibit NK cell function (human). | [199] | |
| IL-10 produced from co-cultures of macrophages can also inhibit NK cell cytotoxic response (mouse and human in vitro). | [200] | |
| NK cells from patients with endometriosis have a high density of NK inhibitory receptors and ligands. However, NK-activating receptors are also expressed at high levels. | [201,202,203] | |
| Mast cells | High numbers of degranulated mast cells have been found in women with endometriotic lesions. | [204,205] |
| Increased concentrations of stem cell factor in the peritoneal fluid of women with endometriosis are responsible for increased mast cell migration. | [162] | |
| Mast cells express estrogen receptors and are highly activated by the estrogens in the ectopic endometrium in patients with endometriosis. | [205] | |
| Mast cells are involved in pain in women with endometriosis. | [206] | |
| Dendritic cells | Increased immature cells (CD80lowCD1ahigh) and fewer mature cells (CD80highCD1alow) in the peritoneal fluid (mouse and human). | [207,208] |
| The activity of enzyme 1-hydroxysteroid dehydrogenase type 1, which activates cortisol, impairs dendritic cell maturation in patients with endometriosis. | [209] | |
| CD1c expression on peripheral myeloid dendritic cells was higher during menstruation in patients with endometriosis. | [210] | |
| IL-10 produced by dendritic cells induces angiogenesis in patients with endometriosis | [211] |
3.3. Adaptive Immune Response
| Cell Type | Characteristics | Ref |
|---|---|---|
| T cells | In patients with endometriosis, circulating CD8+ cells and activated T cells increase, leading to higher secretion of proinflammatory cytokines and elevated autoantibody titers. | [214,215,216] |
| CD8 cell apoptosis is elevated in endometriosis patients due to Fas-FasL interaction. | [151] | |
| Foxp3+CD39+CD73+ Treg cells are decreased in the blood of women with deep infiltrating endometriosis but increased in the peritoneum and endometriotic lesions. | [217,218] | |
| Elevated levels of estrogen and thymus-expressed chemokine (TECK/CCL25) lead to an increase in Tregs, which in turn reduces immune surveillance in endometriosis patients. | [219] | |
| Disruption of Th17/Treg balance leads to heightened inflammation in ectopic and eutopic endometria in women with endometriosis. | [220,221] | |
| Peritoneal fluid from endometriosis patients promoted Treg cell generation and inhibited Th17 cell differentiation in CD4+ T cell cultures in vitro. | [221] | |
| Patients with endometriosis have higher amount of CD16+ CD8 T cells in their peripheral blood, and CD8 T cell cytotoxicity is increased in menstrual effluent. | [222] | |
| Patients with endometriosis show low amounts of perforin-CD8 T cells in peripheral blood. | [223] | |
| Potential T cell exhaustion indicated by PD-1 expression and increased PD-1L presence in tissues of endometriosis patients. | [224] | |
| The Th2 immune response (IL-4, IL-10) dominates later stages of endometriosis, whereas Th1 is present initially. CTLA-4 plays a role in chronic inflammation and endometriosis in humans and mice. | [178,225,226,227] | |
| Higher soluble circulating CTLA-4 levels in patients with endometriosis are associated with chronic inflammation. | [227] | |
| Estrogen plays a role in regulating the GATA3 transcription factor and Th2 differentiation in patients with endometriosis. | [228] | |
| The interleukin IL-4/IFN-γ, IL-10/IFN-γ, and IL-4/IL-2 ratios are higher in women with endometriosis, probably in the late stage. | [229] | |
| B cells | Increased circulating levels of activated B cells in patients with endometriosis. | [230,231] |
| Local B cells secrete high levels of IL-6 and IL-17, inducing local inflammation. They also produce anti-endometrial antibodies. | [231] | |
| The production of IL-35 by B cells is increased in patients with endometriosis. | [232] |
4. Cytokines and Endometriosis
5. Mechanisms of Pain in Endometriosis
6. Endometriosis and Autoimmunity
7. Immunological Therapies in Endometriosis
| Drug | Effects | References |
|---|---|---|
| Glucocorticoids | Inhibit the inflammatory milieu in endometriosis. Prevent the self-renewal, migration, and differentiation of endometrial stem cells and endometriosis formation. | [392,393,394] |
| Statins | Statins reduce inflammation and inhibit the formation of new blood vessels, acting as anti-angiogenic agents in the murine model. | [395,396] |
| In a pilot study, administering atorvastatin 10 mg daily for 7 days improved nitric oxide-mediated, endothelial-dependent cutaneous microvascular function in women with endometriosis. | [397] | |
| Pentoxifylline | It reduces inflammation by regulating the immune response. | [398] |
| There was an increased tendency for pregnancy after surgery in the group treated with pentoxifylline compared to the placebo. | [399] | |
| Patients who received pentoxifylline showed significantly improved visual analog scale scores after 3 months. There is insufficient evidence to recommend pentoxifylline for the treatment of subfertility and pain related to endometriosis. | [400,401,402] | |
| Peroxisome proliferator receptor γ (PPARγ) activators | The compounds inhibit cell proliferation, induce apoptosis in endometriotic epithelial and stromal cells, reduce vascularization, and repress VEGF, IL-6, IL-8, and TNF-α gene expression. | [403] |
| Ciglitazone decreased the size of ectopic endometriotic tissues in a rat model of endometriosis. | [404] | |
| In a baboon model of endometriosis, Rosiglitazone decreased the size of the endometriotic lesion. Pioglitazone improved embryo implantation rates in infertile women with endometriosis undergoing IVF by reducing serum RANTES. | [405] | |
| No clinical trial has been published. | [406] | |
| Rapamycin (mTOR inhibitor) | Rapamycin treatment reduced the volume of lesions in a mouse model of endometriosis. | [407] |
| In women with infertility due to endometriosis, rapamycin has improved rates of fertilization, implantation, clinical pregnancy, and live births. More clinical trials are needed to ascertain the possible benefit of rapamycin treatment. | [408] | |
| Bentamapimod (c-Jun N-terminal kinase inhibitor) | In a mouse and rat model of endometriosis, bentamapimod led to a reduction in lesion size. | [409] |
| In baboons with induced endometriosis, bentamapimod decreased the lesions’ area and volume. No clinical trial has been published. | [410] | |
| Ligustrazine (Tetramethylpyrazine) | A natural product has demonstrated a broad anti-inflammatory effect in preclinical trials. No clinical trial has been published. | [411] |
| Resveratrol | Several preclinical trials have published the anti-neoplastic, anti-inflammatory, anti-oxidative, anti-microbial, anti-atherogenic, and anti-angiogenic effects of resveratrol. | [412] |
| Prevents the progression of experimental endometriosis in living organisms and reduces the invasiveness of endometrial stromal cells in laboratory tests. | [413] | |
| Resveratrol reduced MMP-2 and MMP-9 levels in the endometrium and blood of women with endometriosis. | [414] | |
| Treatment with resveratrol reduced TNF-α2 and VEGF expression in patients with endometriosis. | [415] | |
| There is not enough evidence to support the use of resveratrol in humans. | [416] | |
| Astaxanthin (antioxidant) | Treatment with astaxanthin reduced serum levels of malondialdehyde, IL-1β, and TNF-α, decreasing IL-6 and TNF-α levels in follicular fluid in one triple-blind placebo-controlled clinical trial of patients undergoing assisted reproduction. | [417] |
| Curcumin | In ectopic endometrial stromal cells cultured in vitro, it suppresses the TNF-α-induced secretion of IL-6, IL-8, and MCP-1, and the mRNA expression of ICAM-1 and VCAM-1. | [418] |
| In eutopic endometrial stromal cells of patients with endometriosis, in vitro treatment inhibits the secretion of IL-6, IL-8, G-CSF, MCP-1, and RANTES. | [419] | |
| In a small trial involving nano-micellar curcumin, inflammatory and oxidative patterns linked to IVF treatment in patients with endometriosis showed improvement. | [420] | |
| There is not enough evidence to support the use of curcumin in humans. Well-designed clinical trials are needed. | [421] | |
| Quercetin | Experimental data on quercetin have demonstrated its antioxidant, anti-inflammatory, and anti-angiogenic properties. | [422] |
| It decreased the volume of endometriosis lesions in a mouse model. No clinical trials have been published. | [423] | |
| Epigallocatechin gallate (EGCG) | EGCG notably decreased the proliferation, migration, and invasion of endometrial and endometriotic stromal cells in vitro model of human endometriosis. In mouse models, it also reduced the growth of endometrial lesions. No clinical trials with the purified compound have been published, although trials utilizing green tea have shown some improvement. | [424,425] |
| N-palmitoyl ethanolamine plus trans-polydatin | It induces anti-inflammatory effects in women with endometriosis. It reduced pelvic pain in women after laparoscopy. | [426] |
| A meta-analysis showed no conclusive evidence. | [427] | |
| Cannabidiol | It reduced the diameter, volume, and area of lesions in rat models of endometriosis. It exhibited an anti-fibrotic effect, lowering IL-1β, TNF-α, and PGE2 levels in peritoneal fluids. | [428] |
| It alleviated pelvic pain and related symptoms. Long-term use may be linked to cannabis use disorder, psychosis, and mood disturbances. No clinical trials have been published. | [429,430] | |
| Fenretinide (synthetic retinoid) | Fenretinide reduces the levels of retinol fatty acid binding protein 4. It is used in cancer and cystic fibrosis, but there are no clinical trials in patients with endometriosis. | [431,432,433,434] |
| Vitamin D | The effects of vitamin D supplementation have produced controversial results that require further studies. | [435,436,437] |
| Treatment | Effects | References |
|---|---|---|
| Antibody-based (anti-fibronectin F8) pharmacological delivery of interleukin 4 (F8-IL4) | In a mouse model of endometriosis, F8-IL4 reduced the number and volume of lesions while lowering the expression of genes related to cell adhesion, invasion, and neovascularization, such as integrin β1, MMP-3, MMP-9, and VEGF, without affecting inflammatory cytokines. No clinical studies have been performed in humans. | [438] |
| IL-12 | IL-12 enhances cytokine production and increases NK cell activity. An intraperitoneal injection of IL-12 reduced lesion size in a mouse model by activating NK cells and inhibiting the development of endometriotic lesions. No studies have been performed in humans. | [439,440] |
| Interferon (IFN) I | In a rat model of endometriosis, the subcutaneous administration of IFN-α reduced the volume of endometriosis lesions. | [441] |
| IFN-β1a inhibited the in vitro growth and movement of endometrial stromal cells obtained from patients. | [442] | |
| IFNα2b treatment increased the later recurrence of endometriosis in a small clinical trial. | [443] | |
| IL-37 | Anti-inflammatory effects. In mouse models, IL-37 reduced the size and weight of endometriotic-like lesions and the expression of IL-1β, IL-6, IL-10, TNF-α, VEGF, and ICAM-1 in a murine model of endometriosis. No studies have been performed in humans. | [444,445] |
| Anti-TNF-α | In patients with endometriomas who were treated using assisted reproductive technology, etanercept was shown to increase the pregnancy rate and double the live birth rate. However, this result was not statistically significant. (p = 0.052). | [446] |
| In a retrospective study, peri-implantation treatment with TNF-α inhibitor increased the implantation rate and clinical pregnancy rate significantly compared with non-treated controls; however, no changes in the pregnancy rate of live birth were observed. Cochrane reviews of humans with endometriosis did not find conclusive evidence. | [447] | |
| Epidemiological data on young women treated with anti-TNFα therapy and endometriosis incidence have not been published. | [448] | |
| IL-1 antagonist (anakinra) | In a pilot study using anakinra, mild improvements were observed. A reduction in the inflammatory markers BDNF, IL-1RA, and IL-6 was reported. | [449] |
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Available online: https://www.who.int/news-room/fact-sheets/detail/endometriosis (accessed on 7 April 2025).
- Tsamantioti, E.S.; Mahdy, H. Endometriosis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK567777/ (accessed on 7 May 2025).
- Smolarz, B.; Szyłło, K.; Romanowicz, H. Endometriosis: Epidemiology, classification, pathogenesis, treatment and genetics. Int. J. Mol. Sci. 2021, 22, 10554. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.K.; Chapron, C.; Giudice, L.C.; Laufer, M.R.; Leyland, N.; Missmer, S.A.; Singh, S.S.; Taylor, H.S. Clinical Diagnosis of Endometriosis: A Call to Action. Am. J. Obstet. Gynecol. 2019, 220, 354.e1–354.e12. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Lim, W.; Kim, M.; Jang, H.; Park, S.J.; Song, G.; Park, S. Female reproductive disease, endometriosis: From inflammation to infertility. Mol. Cells 2025, 48, 100164. [Google Scholar] [CrossRef]
- Simoens, S.; Dunselman, G.; Dirksen, C.; Hummelshoj, L.; Bokor, A.; Brandes, I.; Brodszky, V.; Canis, M.; Colombo, G.L.; DeLeire, T.; et al. The burden of endometriosis: Costs and quality of life of women with endometriosis and treated in referral centres. Hum. Reprod. 2012, 27, 1292–1299. [Google Scholar] [CrossRef]
- Hadfield, R.; Mardon, H.; Barlow, D.; Kennedy, S. Delay in the diagnosis of endometriosis: A survey of women from the USA and the UK. Hum. Reprod. 1996, 11, 878–880. [Google Scholar] [CrossRef]
- Swift, B.; Taneri, B.; Becker, C.M.; Basarir, H.; Naci, H.; Missmer, S.A.; Zondervan, K.T.; Rahmioglu, N. Prevalence, diagnostic delay and economic burden of endometriosis and its impact on quality of life: Results from an Eastern Mediterranean population. Eur. J. Public Health 2024, 34, 244–252. [Google Scholar] [CrossRef]
- Taylor, H.S.; Kotlyar, A.M.; Flores, V.A. Endometriosis is a chronic systemic disease: Clinical challenges and novel innovations. Lancet 2021, 397, 839–852. [Google Scholar] [CrossRef]
- Tariverdian, N.; Siedentopf, F.; Rücke, M.; Blois, S.M.; Klapp, B.F.; Kentenich, H.; Arck, P.C. Intraperitoneal immune cell status in infertile women with and without endometriosis. J. Reprod. Immunol. 2009, 80, 80–90. [Google Scholar] [CrossRef]
- Amidifar, S.; Jafari, D.; Mansourabadi, A.H.; Sadaghian, S.; Esmaeilzadeh, A. Immunopathology of Endometriosis, Molecular Approaches. Am. J. Reprod. Immunol. 2025, 93, e70056. [Google Scholar] [CrossRef] [PubMed]
- Bao, Q.; Zheng, Q.; Wang, S.; Tang, W.; Zhang, B. LncRNA HOTAIR regulates cell invasion and migration in endometriosis through miR-519b-3p/PRRG4 pathway. Front. Oncol. 2022, 12, 953055. [Google Scholar] [CrossRef]
- Blanco, L.P.; Salmeri, N.; Temkin, S.M.; Shanmugam, V.K.; Stratton, P. Endometriosis and autoimmunity. Autoimmun. Rev. 2025, 24, 103752. [Google Scholar] [CrossRef] [PubMed]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Gordts, S.; Koninckx, P.; Brosens, I. Pathogenesis of deep endometriosis. Fertil. Steril. 2017, 108, 872–885.e1. [Google Scholar] [CrossRef]
- Guan, Y.; Chen, Y.; Lin, R.; Mo, T.; Li, S.; Cao, Y.; Yin, T.; Diao, L.; Li, Y. Endometriosis: A new perspective on epigenetics and oxidative stress. J. Reprod. Immunol. 2025, 169, 104462. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Götte, M.; Viganò, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The Pathogenesis of Endometriosis: Molecular and Cell Biology Insights. Int. J. Mol. Sci. 2019, 20, 5615. [Google Scholar] [CrossRef]
- Wang, Y.; Nicholes, K.; Shih, I.M. The Origin and Pathogenesis of Endometriosis. Annu. Rev. Pathol. 2020, 15, 71–95. [Google Scholar] [CrossRef]
- Bo, C.; Wang, Y. Angiogenesis signaling in endometriosis: Molecules, diagnosis and treatment (Review). Mol. Med. Rep. 2024, 29, 43. [Google Scholar] [CrossRef]
- Karimi-Zarchi, M.; Dehshiri-Zadeh, N.; Sekhavat, L.; Nosouhi, F. Correlation of CA-125 serum level and clinico-pathological characteristic of patients with endometriosis. Int. J. Reprod. Biomed. 2016, 14, 713–718. [Google Scholar] [CrossRef]
- Neves, D.; Neto, A.C.; Salazar, M.; Fernandes, A.S.; Martinho, M.; Charrua, A.; Rodrigues, A.R.; Gouveia, A.M.; Almeida, H. A narrative review about the intricate crosstalk among endometrium, adipose tissue, and neurons in endometriosis. The multifaceted role of leptin. Obes. Rev. 2025, 26, e13879. [Google Scholar] [CrossRef]
- Abulughod, N.; Valakas, S.; El-Assaad, F. Dietary and Nutritional Interventions for the Management of Endometriosis. Nutrients 2024, 16, 3988. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.H.; Jeong, J.W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, S.K.; Lee, J.R.; Jee, B.C. Management of endometriosis-related infertility: Considerations and treatment options. Clin. Exp. Reprod. Med. 2020, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- García-Gómez, E.; Vázquez-Martínez, E.R.; Reyes-Mayoral, C.; Cruz-Orozco, O.P.; Camacho-Arroyo, I.; Cerbón, M. Regulation of Inflammation Pathways and Inflammasome by Sex Steroid Hormones in Endometriosis. Front. Endocrinol. 2020, 10, 935. [Google Scholar] [CrossRef]
- Rolla, E. Endometriosis: Advances and controversies in classification, pathogenesis, diagnosis, and treatment. F1000Research 2019, 8, F1000-Faculty. [Google Scholar] [CrossRef]
- Rocha, T.P.; Andres, M.P.; Carmona, F.; Baracat, E.C.; Abrão, M.S. Deep Endometriosis: The Involvement of Multiple Pelvic Compartments Is Associated with More Severe Pain Symptoms and Infertility. Reprod. Sci. 2023, 30, 1668–1675. [Google Scholar] [CrossRef]
- Camboni, A.; Marbaix, E. Ectopic Endometrium: The Pathologist’s Perspective. Int. J. Mol. Sci. 2021, 22, 10974. [Google Scholar] [CrossRef]
- Istrate-Ofiţeru, A.M.; Berbecaru, E.I.; Zorilă, G.L.; Roşu, G.C.; Dîră, L.M.; Comănescu, C.M.; Drăguşin, R.C.; Ruican, D.; Nagy, R.D.; Iliescu, D.G.; et al. Specific Local Predictors That Reflect the Tropism of Endometriosis-A Multiple Immunohistochemistry Technique. Int. J. Mol. Sci. 2022, 23, 5614. [Google Scholar] [CrossRef]
- Moraru, L.; Mitranovici, M.I.; Chiorean, D.M.; Moraru, R.; Caravia, L.; Tirón, A.T.; Cotoi, O.S. Adenomyosis and Its Possible Malignancy: A Review of the Literature. Diagnostics 2023, 13, 1883. [Google Scholar] [CrossRef]
- Li, Q.; Chen, G.; Jiang, H.; Dai, H.; Li, D.; Zhu, K.; Zhang, K.; Shen, H.; Xu, H.; Li, S. ITGB3 promotes cisplatin resistance in osteosarcoma tumors. Cancer Med. 2023, 12, 8452–8463. [Google Scholar] [CrossRef]
- Zhu, C.; Kong, Z.; Wang, B.; Cheng, W.; Wu, A.; Meng, X. ITGB3/CD61: A hub modulator and target in the tumor microenvironment. Am. J. Transl. Res. 2019, 11, 7195–7208. [Google Scholar]
- Zhang, L.; Shao, W.; Li, M.; Liu, S. ITCH-Mediated Ubiquitylation of ITGB3 Promotes Cell Proliferation and Invasion of Ectopic Endometrial Stromal Cells in Ovarian Endometriosis. Biomedicines 2023, 11, 2506. [Google Scholar] [CrossRef] [PubMed]
- Shigesi, N.; Kvaskoff, M.; Kirtley, S.; Feng, Q.; Fang, H.; Knight, J.C.; Missmer, S.A.; Rahmioglu, N.; Zondervan, K.T.; Becker, C.M. The association between endometriosis and autoimmune diseases: A systematic review and meta-analysis. Hum. Reprod. Update 2019, 25, 486–503. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Lin, M.; Hu, K.L.; Li, R. Cross-Tissue Regulatory Network Analyses Reveal Novel Susceptibility Genes and Potential Mechanisms for Endometriosis. Biology 2024, 13, 871. [Google Scholar] [CrossRef]
- Wong, F.C.; Kim, C.E.; Garcia-Alonso, L.; Vento-Tormo, R. The human endometrium: Atlases, models, and prospects. Curr. Opin. Genet. Dev. 2025, 92, 102341. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, G. Endometriosis-associated Ovarian Clear Cell Carcinoma: A Special Entity? J. Cancer 2021, 12, 6773–6786. [Google Scholar] [CrossRef]
- Giannini, A.; Massimello, F.; Caretto, M.; Cosimi, G.; Mannella, P.; Luisi, S.; Gadducci, A.; Simoncini, T. Factors in malignant transformation of ovarian endometriosis: A narrative review. Gynecol. Endocrinol. 2024, 40, 2409911. [Google Scholar] [CrossRef]
- Murakami, K.; Kotani, Y.; Nakai, H.; Matsumura, N. Endometriosis-Associated Ovarian Cancer: The Origin and Targeted Therapy. Cancers 2020, 12, 1676. [Google Scholar] [CrossRef]
- Capozzi, V.A.; Scarpelli, E.; dell’Omo, S.; Rolla, M.; Pezzani, A.; Morganelli, G.; Gaiano, M.; Ghi, T.; Berretta, R. Atypical Endometriosis: A Comprehensive Systematic Review of Pathological Patterns and Diagnostic Challenges. Biomedicines 2024, 12, 1209. [Google Scholar] [CrossRef]
- Lei, L.; Xu, X.; Gong, C.; Lin, B.; Li, F. Integrated analysis of genome-wide gene expression and DNA methylation profiles reveals candidate genes in ovary endometriosis. Front. Endocrinol. 2023, 14, 1093683. [Google Scholar] [CrossRef]
- Jung, J.; Kim, N.H.; Park, J.; Lim, D.; Kwon, M.; Gil, W.; Jung, S.; Go, M.; Kim, C.; Cheong, Y.H.; et al. Gremlin-2 is a novel tumor suppressor that negatively regulates ID1 in breast cancer. Breast Cancer Res. 2024, 26, 174. [Google Scholar] [CrossRef]
- Zhai, M.; Yang, W.; Zou, C.; Du, S.; Wu, B.; Wang, C.; Lu, Y.; Zheng, Y. Predictive role of HPGD gene in carcinogenesis and immune environment monitoring in human cervical cancer. Cancer Biomark. Sect. A Dis. Markers 2024, 41, 18758592241296277. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Dai, X.; Huang, C.; Guan, X.; Zhang, C. 17beta-estradiol (E2) Regulates Malignancies and Stemness in Endometrial Carcinoma (EC) via Interacting with ESR1. Reprod. Sci. 2025. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zheng, Y.; Wang, J.; Xu, H.; Zhang, R.; Xie, Z.; Zhang, L.; Zhao, R. Identification of key genes associated with endometriosis and endometrial cancer by bioinformatics analysis. Front. Oncol. 2024, 14, 1387860. [Google Scholar] [CrossRef]
- Anglesio, M.S.; Bashashati, A.; Wang, Y.K.; Senz, J.; Ha, G.; Yang, W.; Aniba, M.R.; Prentice, L.M.; Farahani, H.; Li Chang, H.; et al. Multifocal endometriotic lesions associated with cancer are clonal and carry a high mutation burden. J. Pathol. 2015, 236, 201–209. [Google Scholar] [CrossRef]
- Wilbur, M.A.; Shih, I.M.; Segars, J.H.; Fader, A.N. Cancer Implications for Patients with Endometriosis. Semin. Reprod. Med. 2017, 35, 110–116. [Google Scholar] [CrossRef]
- Lu, Y.; Cuellar-Partida, G.; Painter, J.N.; Nyholt, D.R.; Australian Ovarian Cancer Study; International Endogene Consortium (IEC). Shared genetics underlying epidemiological association between endometriosis and ovarian cancer. Hum. Mol. Genet. 2015, 24, 5955–5964. [Google Scholar] [CrossRef]
- Parra-Herran, C.; Lerner-Ellis, J.; Xu, B.; Khalouei, S.; Bassiouny, D.; Cesari, M.; Ismiil, N.; Nofech-Mozes, S. Molecular-based classification algorithm for endometrial carcinoma categorizes ovarian endometrioid carcinoma into prognostically significant groups. Mod. Pathol. 2017, 30, 1748–1759. [Google Scholar] [CrossRef]
- Chou, Y.C.; Chen, C.H.; Chen, M.J.; Chang, C.W.; Chen, P.H.; Yu, M.H.; Chen, Y.J.; Tsai, E.M.; Yang, P.S.; Lin, S.Y.; et al. Killer cell immunoglobulin-like receptors (KIR) and human leukocyte antigen-C (HLA-C) allorecognition patterns in women with endometriosis. Sci. Rep. 2020, 10, 4897. [Google Scholar] [CrossRef]
- Marin, M.L.C.; Coelho, V.; Visentainer, J.E.L.; Alves, H.V.; Köhler, K.F.; Rached, M.R.; Abrão, M.S.; Kalil, J. Inhibitory KIR2DL2 Gene: Risk for Deep Endometriosis in Euro-descendants. Reprod. Sci. 2021, 28, 291–304. [Google Scholar] [CrossRef]
- Kitawaki, J.; Xu, B.; Ishihara, H.; Fukui, M.; Hasegawa, G.; Nakamura, N.; Mizuno, S.; Ohta, M.; Obayashi, H.; Honjo, H. Association of killer cell immunoglobulin-like receptor genotypes with susceptibility to endometriosis. Am. J. Reprod. Immunol. 2007, 58, 481–486. [Google Scholar] [CrossRef]
- Kula, H.; Balbal, B.; Timur, T.; Yalcın, P.; Yavuz, O.; Kızıldag, S.; Ulukus, E.C.; Posaci, C. NOD1, NOD2, PYDC1, and PYDC2 gene polymorphisms in ovarian endometriosis. Front. Med. 2025, 11, 1495002. [Google Scholar] [CrossRef] [PubMed]
- Badie, A.; Saliminejad, K.; Salahshourifar, I.; Khorram Khorshid, H.R. Interleukin 1 alpha (IL1A) polymorphisms and risk of endometriosis in Iranian population: A case-control study. Gynecol. Endocrinol. 2020, 36, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Hu, M.; Chen, J.M.; Sun, W.; Zhu, M.B. Effects of gene polymorphism and serum levels of IL-2 and IL-6 on endometriosis. Europ. Rev. Med. Pharmacol. Sci. 2020, 24, 4635–4641. [Google Scholar] [CrossRef]
- Zhong, S.; Liang, Y.; Wu, Z.; Wei, L. Association between polymorphisms of cytokine genes and endometriosis: A comprehensive systematic review and meta-analysis. J. Reprod. Immunol. 2023, 158, 103969. [Google Scholar] [CrossRef]
- Zhao, W.; Li, Y.; Zhao, J.; Kang, S. A functional promoter polymorphism in interleukin 12B gene is associated with an increased risk of ovarian endometriosis. Gene 2018, 666, 27–31. [Google Scholar] [CrossRef]
- Zare, M.; Hesampour, F.; Poordast, T.; Valibeigi, M.; Enayatmehri, M.; Ahmadi, S.; Nasri, F.; Gharesi-Fard, B. Association between gene polymorphisms of IL-12, IL-12 receptor and IL-27 and organ involvement in Iranian endometriosis patients. Inter. J. Immun. 2023, 50, 24–33. [Google Scholar] [CrossRef]
- Watrowski, R.; Schuster, E.; Van Gorp, T.; Hofstetter, G.; Fischer, M.B.; Mahner, S.; Polterauer, S.; Zeillinger, R.; Obermayr, E. Association of the Single Nucleotide Polymorphisms rs11556218, rs4778889, rs4072111, and rs1131445 of the Interleukin-16 Gene with Ovarian Cancer. Int. J. Mol. Sci. 2024, 25, 10272. [Google Scholar] [CrossRef]
- Babah, O.A.; Ojewunmi, O.O.; Onwuamah, C.K.; Udenze, I.C.; Osuntoki, A.A.; Afolabi, B.B. Serum concentrations of IL-16 and its genetic polymorphism rs4778889 affect the susceptibility and severity of endometriosis in Nigerian women. BMC Women’s Health 2023, 23, 253. [Google Scholar] [CrossRef]
- Xie, Z.; Ding, X.; Wang, Y.; Zhang, M. The rs2275913 polymorphism of the interleukin-17A gene is associated with the risk of ovarian endometriosis. J. Obstet. Gynaecol. 2023, 43, 2199852. [Google Scholar] [CrossRef]
- Balunathan, N.; Rani, G.U.; Perumal, V.; Kumarasamy, P. Single nucleotide polymorphisms of Interleukin - 4, Interleukin-18, FCRL3 and sPLA2IIa genes and their association in pathogenesis of endometriosis. Mol. Biol. Rep. 2023, 50, 4239–4252. [Google Scholar] [CrossRef]
- Mier-Cabrera, J.; Cruz-Orozco, O.; de la Jara-Díaz, J.; Galicia-Castillo, O.; Buenrostro-Jáuregui, M.; Parra-Carriedo, A.; Hernández-Guerrero, C. Polymorphisms of TNF-alpha (−308), IL-1beta (+3954) and IL1-Ra (VNTR) are associated to severe stage of endometriosis in Mexican women: A case control study. BMC Women’s Health 2022, 22, 356. [Google Scholar] [CrossRef] [PubMed]
- Chekini, Z.; Poursadoughian Yaran, A.; Ansari-Pour, N.; Shahhoseini, M.; Ramazanali, F.; Aflatoonian, R.; Afsharian, P. A novel gene-wide haplotype at the macrophage migration inhibitory factor (MIF) locus is associated with endometrioma. Europ. J. Obst. Gynecol. Reprod. Biol. 2020, 247, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.V.; Machado, D.E.; da Silva, M.C.; de Mello, M.P.; Berardo, P.T.; Medeiros, R.; Perini, J.A. Influence of interleukin-8 polymorphism on endometriosis-related pelvic pain. Hum. Immunol. 2023, 84, 561–566. [Google Scholar] [CrossRef]
- Le, K.N.; Benor, A.; Decherney, A. An update on epigenetic mechanisms in endometriosis. Minerva Obstet. Gynecol. 2024. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yilmaz, B.D.; Sison, C.; Miyazaki, K.; Bernardi, L.; Liu, S.; Kohlmeier, A.; Yin, P.; Milad, M.; Wei, J. Endometriosis. Endocr. Rev. 2019, 40, 1048–1079. [Google Scholar] [CrossRef]
- Raja, M.H.R.; Farooqui, N.; Zuberi, N.; Ashraf, M.; Azhar, A.; Baig, R.; Badar, B.; Rehman, R. Endometriosis, infertility and MicroRNA’s: A review. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102157. [Google Scholar] [CrossRef]
- Hon, J.X.; Wahab, N.A.; Karim, A.K.A.; Mokhtar, N.M.; Mokhtar, M.H. MicroRNAs in Endometriosis: Insights into Inflammation and Progesterone Resistance. Int. J. Mol. Sci. 2023, 24, 15001. [Google Scholar] [CrossRef]
- Azari, Z.D.; Aljubran, F.; Nothnick, W.B. Inflammatory MicroRNAs and the Pathophysiology of Endometriosis and Atherosclerosis: Common Pathways and Future Directions Towards Elucidating the Relationship. Reprod. Sci. 2022, 29, 2089–2104. [Google Scholar] [CrossRef]
- Liao, Z.; Tang, S.; Jiang, P.; Geng, T.; Cope, D.I.; Dunn, T.N.; Guner, J.; Radilla, L.A.; Guan, X.; Monsivais, D. Impaired bone morphogenetic protein (BMP) signaling pathways disrupt decidualization in endometriosis. Commun. Biol. 2024, 7, 227. [Google Scholar] [CrossRef]
- Abbaszadeh, M.; Karimi, M.; Rajaei, S. The landscape of non-coding RNAs in the immunopathogenesis of Endometriosis. Front. Immunol. 2023, 14, 1223828. [Google Scholar] [CrossRef]
- González-Ramos, R.; Van Langendonckt, A.; Defrère, S.; Lousse, J.C.; Colette, S.; Devoto, L.; Donnez, J. Involvement of the nuclear factor-κB pathway in the pathogenesis of endometriosis. Fertil. Steril. 2010, 94, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Zdrojkowski, Ł.; Jasiński, T.; Ferreira-Dias, G.; Pawliński, B.; Domino, M. The Role of NF-κB in Endometrial Diseases in Humans and Animals: A Review. Int. J. Mol. Sci. 2023, 24, 2901. [Google Scholar] [CrossRef] [PubMed]
- Vissers, G.; Giacomozzi, M.; Verdurmen, W.; Peek, R.; Nap, A. The role of fibrosis in endometriosis: A systematic review. Hum. Reprod. Update 2024, 30, 706–750. [Google Scholar] [CrossRef]
- Anchan, M.M.; Kalthur, G.; Datta, R.; Majumdar, K.P.K.; Dutta, R. Unveiling the fibrotic puzzle of endometriosis: An overlooked concern calling for prompt action. F1000Research 2024, 13, 721. [Google Scholar] [CrossRef]
- Almquist, L.D.; Likes, C.E.; Stone, B.; Brown, K.R.; Savaris, R.; Forstein, D.A.; Miller, P.B.; Lessey, B.A. Endometrial BCL6 testing for the prediction of in vitro fertilisation outcomes: A cohort study. Fertil. Steril. 2017, 108, 1063–1069. [Google Scholar] [CrossRef]
- Saadat Varnosfaderani, A.; Kalantari, S.; Ramezanali, F.; Shahhoseini, M.; Amirchaghmaghi, E. Increased Gene Expression of LITAF, TNF-α and BCL6 in Endometrial Tissues of Women with Endometriosis: A Case-Control Study. Cell J. 2024, 26, 243–249. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, S.; Xie, Y.; Tong, Y.; Qi, W.; Wang, Z. Endometrial expression of ERRβ and ERRγ: Prognostic significance and clinical correlations in severe endometriosis. Front. Endocrinol. 2024, 15, 1489097. [Google Scholar] [CrossRef]
- Cheng, C.W.; Licence, D.; Cook, E.; Luo, F.; Arends, M.J.; Smith, S.K.; Print, C.G.; Charnock-Jones, D.S. Activation of mutated K-ras in donor endometrial epithelium and stroma promotes lesion growth in an intact immunocompetent murine model of endometriosis. J. Pathol. 2011, 224, 261–269. [Google Scholar] [CrossRef]
- Maeda, D.; Shih, I.-M. Pathogenesis and the role of ARID1A mutation in endometriosis-related ovarian neoplasms. Adv. Anat. Pathol. 2013, 20, 45–52. [Google Scholar] [CrossRef]
- Steinbuch, S.C.; Lüß, A.M.; Eltrop, S.; Götte, M.; Kiesel, L. Endometriosis-Associated Ovarian Cancer: From Molecular Pathologies to Clinical Relevance. Int. J. Mol. Sci. 2024, 25, 4306. [Google Scholar] [CrossRef]
- Pan, Y.; Pan, C.; Zhang, C. Unraveling the complexity of follicular fluid: Insights into its composition, function, and clinical implications. J. Ovarian Res. 2024, 17, 237. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Hicks, C.; El-Omar, E.; Combes, V.; El-Assaad, F. The Critical Role of Host and Bacterial Extracellular Vesicles in Endometriosis. Biomedicines 2024, 12, 2585. [Google Scholar] [CrossRef] [PubMed]
- Duval, C.; Wyse, B.A.; Tsang, B.K.; Librach, C.L. Extracellular vesicles and their content in the context of polycystic ovarian syndrome and endometriosis: A review. J. Ovarian Res. 2024, 17, 160. [Google Scholar] [CrossRef]
- Nazri, H.M.; Imran, M.; Fischer, R.; Heilig, R.; Manek, S.; Dragovic, R.A.; Kessler, B.M.; Zondervan, K.T.; Tapmeier, T.T.; Becker, C.M. Characterization of exosomes in peritoneal fluid of endometriosis patients. Fertil. Steril. 2020, 113, 364–373.e2. [Google Scholar] [CrossRef]
- Björk, E.; Israelsson, P.; Nagaev, I.; Nagaeva, O.; Lundin, E.; Ottander, U.; Mincheva-Nilsson, L. Endometriotic Tissue-derived Exosomes Downregulate NKG2D-mediated Cytotoxicity and Promote Apoptosis: Mechanisms for Survival of Ectopic Endometrial Tissue in Endometriosis. J. Immunol. 2024, 213, 567–576. [Google Scholar] [CrossRef]
- Ding, D.; Liu, X.; Duan, J.; Guo, S.W. Platelets are an unindicted culprit in the development of endometriosis: Clinical and experimental evidence. Hum. Reprod. 2015, 30, 812–832. [Google Scholar] [CrossRef]
- Bortot, B.; Di Florio, R.; Merighi, S.; Peacock, B.; Lees, R.; Valle, F.; Brucale, M.; Mangogna, A.; Di Lorenzo, G.; Romano, F.; et al. Platelets as key cells in endometriosis patients: Insights from small extracellular vesicles in peritoneal fluid and endometriotic lesions analysis. FASEB J. 2024, 38, e70267. [Google Scholar] [CrossRef]
- Ding, S.; Lin, Q.; Zhu, T.; Li, T.; Zhu, L.; Wang, J.; Zhang, X. Is there a correlation between inflammatory markers and coagulation parameters in women with advanced ovarian endometriosis? BMC Women’s Health 2019, 19, 169. [Google Scholar] [CrossRef]
- Dantzler, M.D.; Miller, T.A.; Dougherty, M.W.; Quevedo, A. The Microbiome Landscape of Adenomyosis: A Systematic Review. Reprod. Sci. 2025, 32, 251–260. [Google Scholar] [CrossRef]
- Guo, W.; Xu, Z.; Hu, S.; Shen, Y. Exploring Microbial Signatures in Endometrial Tissues with Endometriosis. Int. Immunopharmacol. 2025, 148, 114072. [Google Scholar] [CrossRef]
- Qin, R.; Tian, G.; Liu, J.; Cao, L. The gut microbiota and endometriosis: From pathogenesis to diagnosis and treatment. Front. Cell. Infect. Microbiol. 2022, 12, 1069557. [Google Scholar] [CrossRef] [PubMed]
- Escorcia Mora, P.; Valbuena, D.; Diez-Juan, A. The Role of the Gut Microbiota in Female Reproductive and Gynecological Health: Insights into Endometrial Signaling Pathways. Life 2025, 15, 762. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Ding, Q.; Zhang, W.; Kang, M.; Ma, J.; Zhao, L. Gut microbial beta-glucuronidase: A vital regulator in female estrogen metabolism. Gut Microbes 2023, 15, 2236749. [Google Scholar] [CrossRef]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef]
- Wei, W.; Zhang, X.; Tang, H.; Zeng, L.; Wu, R. Microbiota composition and distribution along the female reproductive tract of women with endometriosis. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 15. [Google Scholar] [CrossRef]
- Ata, B.; Yildiz, S.; Turkgeldi, E.; Brocal, V.P.; Dinleyici, E.C.; Moya, A.; Urman, B. The Endobiota Study: Comparison of Vaginal, Cervical and Gut Microbiota Between Women with Stage 3/4 Endometriosis and Healthy Controls. Sci. Rep. 2019, 9, 2204. [Google Scholar] [CrossRef]
- Huang, L.; Liu, B.; Liu, Z.; Feng, W.; Liu, M.; Wang, Y.; Peng, D.; Fu, X.; Zhu, H.; Cui, Z.; et al. Gut Microbiota Exceeds Cervical Microbiota for Early Diagnosis of Endometriosis. Front. Cell. Infect. Microbiol. 2021, 11, 788836. [Google Scholar] [CrossRef]
- Svensson, A.; Brunkwall, L.; Roth, B.; Orho-Melander, M.; Ohlsson, B. Associations Between Endometriosis and Gut Microbiota. Reprod. Sci. 2021, 28, 2367–2377. [Google Scholar] [CrossRef]
- Shan, J.; Ni, Z.; Cheng, W.; Zhou, L.; Zhai, D.; Sun, S.; Yu, C. Gut microbiota imbalance and its correlations with hormone and inflammatory factors in patients with stage 3/4 endometriosis. Arch. Gynecol. Obstet. 2021, 304, 1363–1373. [Google Scholar] [CrossRef]
- Ye, H.; Tian, Y.; Yu, X.; Li, L.; Hou, M. Association Between Pelvic Inflammatory Disease and Risk of Endometriosis: A Systematic Review and Meta-Analysis. J. Women’s Health 2024, 33, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, J.V.; De Sanctis, C.V.; Hajdúch, M.; De Sanctis, J.B. Microbiota and Recurrent Pregnancy Loss (RPL); More than a Simple Connection. Microorganisms 2024, 12, 1641. [Google Scholar] [CrossRef] [PubMed]
- Sobstyl, A.; Chałupnik, A.; Mertowska, P.; Grywalska, E. How Do Microorganisms Influence the Development of Endometriosis? Participation of Genital, Intestinal and Oral Microbiota in Metabolic Regulation and Immunopathogenesis of Endometriosis. Int. J. Mol. Sci. 2023, 24, 10920. [Google Scholar] [CrossRef]
- Leonardi, M.; Hicks, C.; El-Assaad, F.; El-Omar, E.; Condous, G. Endometriosis and the microbiome: A systematic review. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 239–249. [Google Scholar] [CrossRef]
- Jimenez, N.; Norton, T.; Diadala, G.; Bell, E.; Valenti, M.; Farland, L.V.; Mahnert, N.; Herbst-Kralovetz, M.M. Vaginal and rectal microbiome contribute to genital inflammation in chronic pelvic pain. BMC Med. 2024, 22, 283. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, C. Role of the gut microbiota in the pathogenesis of endometriosis: A review. Front. Microbiol. 2024, 15, 1363455. [Google Scholar] [CrossRef]
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate immune pattern recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef]
- Guo, B.; Chen, J.H.; Zhang, J.H.; Fang, Y.; Liu, X.J.; Zhang, J.; Zhu, H.Q.; Zhan, L. Pattern-recognition receptors in endometriosis: A narrative review. Front. Immunol. 2023, 14, 1161606. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, D.; Han, X.; Ren, Y.; Fan, Y.; Zhang, C.; Sun, L.; Ye, T.; Wang, Q.; Ban, Y.; et al. Alarmins and their pivotal role in the pathogenesis of spontaneous abortion: Insights for therapeutic intervention. Eur. J. Med. Res. 2024, 29, 640. [Google Scholar] [CrossRef]
- Chen, F.; Tang, H.; Cai, X.; Lin, J.; Kang, R.; Tang, D.; Liu, J. DAMPs in immunosenescence and cancer. Semin. Cancer Biol. 2024, 106–107, 123–142. [Google Scholar] [CrossRef]
- Kobayashi, H.; Higashiura, Y.; Shigetomi, H.; Kajihara, H. Pathogenesis of endometriosis: The role of initial infection and subsequent sterile inflammation (Review). Mol. Med. Rep. 2014, 9, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Sobstyl, M.; Niedźwiedzka-Rystwej, P.; Grywalska, E.; Korona-Głowniak, I.; Sobstyl, A.; Bednarek, W.; Roliński, J. Toll-Like Receptor 2 Expression as a New Hallmark of Advanced Endometriosis. Cells 2020, 9, 1813. [Google Scholar] [CrossRef] [PubMed]
- Noh, E.J.; Kim, D.J.; Lee, J.Y.; Park, J.H.; Kim, J.S.; Han, J.W.; Kim, B.C.; Kim, C.J.; Lee, S.K. Ureaplasma Urealyticum Infection Contributes to the Development of Pelvic Endometriosis Through Toll-Like Receptor 2. Front. Immunol. 2019, 10, 2373. [Google Scholar] [CrossRef]
- de Azevedo, B.C.; Mansur, F.; Podgaec, S. systematic review of toll-like receptors in endometriosis. Arch. Gynecol. Obstet. 2021, 304, 309–316. [Google Scholar] [CrossRef]
- Almasi, M.Z.; Hosseini, E.; Jafari, R.; Aflatoonian, K.; Aghajanpour, S.; Ramazanali, F.; Moini, A.; Shahhoseini, M.; Afsharian, P.; Aflatoonian, R. Evaluation of Toll-like receptor 3 (TLR3) signaling pathway genes and its genetic polymorphisms in ectopic and eutopic endometrium of women with endometriosis. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102153. [Google Scholar] [CrossRef]
- Allhorn, S.; Böing, C.; Koch, A.A.; Kimmig, R.; Gashaw, I. TLR3 and TLR4 expression in healthy and diseased human endometrium. Reprod. Biol. Endocrinol. 2008, 6, 40. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Inflammasome activation and regulation: Toward a better understanding of complex mechanisms. Cell Discov. 2020, 6, 36. [Google Scholar] [CrossRef]
- Al Mamun, A.; Geng, P.; Wang, S.; Shao, C. Role of Pyroptosis in Endometrial Cancer and Its Therapeutic Regulation. J. Inflamm. Res. 2024, 17, 7037–7056. [Google Scholar] [CrossRef]
- Irandoost, E.; Najibi, S.; Talebbeigi, S.; Nassiri, S. Focus on the role of NLRP3 inflammasome in the pathology of endometriosis: A review on molecular mechanisms and possible medical applications. Naunyn Schmiedeberg’s Arch. Pharmacol. 2023, 396, 621–631. [Google Scholar] [CrossRef]
- Ahn, S.H.; Khalaj, K.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Immune-inflammation gene signatures in endometriosis patients. Fertil. Steril. 2016, 106, 1420–1431.e7. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Pinto, B.; Costa, L.; Felgueira, E.; Rebelo, I. Increased expression of NLRP3 inflammasome components in granulosa cells and follicular fluid interleukin(IL)-1beta and IL-18 levels in fresh IVF/ICSI cycles in women with endometriosis. J. Assist. Reprod. Genet. 2023, 40, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Osuka, S.; Muraoka, A.; Hayashi, S.; Bayasula; Kasahara, Y.; Sonehara, R.; Hariyama, Y.; Shinjo, K.; Tanaka, H.; et al. Effectiveness of NLRP3 Inhibitor as a Non-Hormonal Treatment for ovarian endometriosis. Reprod. Biol. Endocrinol. 2022, 20, 58. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jiang, Z.; Zhang, L.; Tian, W.; Lin, A.; Li, M. Blockage of the NLRP3 inflammasome by MCC950 inhibits migration and invasion in adenomyosis. Reprod. Biomed. Online 2024, 49, 104319. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, Z.; Peng, X.; Cai, D.; Peng, R.; Lin, Y.; Dai, L.; Li, J.; Chen, Y.; Xiao, J.; et al. NLRP3 inflammasome-mediated Pyroptosis induce Notch signal activation in endometriosis angiogenesis. Mol. Cell. Endocrinol. 2023, 574, 111952. [Google Scholar] [CrossRef]
- Zhou, F.; Zhao, F.; Huang, Q.; Lin, X.; Zhang, S.; Dai, Y. NLRP3 activated macrophages promote endometrial stromal cells migration in endometriosis. J. Reprod. Immunol. 2022, 152, 103649. [Google Scholar] [CrossRef]
- Bergqvist, A.; Bruse, C.; Carlberg, M.; Carlström, K. Interleukin 1beta, interleukin-6, and tumor necrosis factor-alpha in endometriotic tissue and in endometrium. Fertil. Steril. 2001, 75, 489–495. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, H.; Xiong, W.; Peng, Y.; Li, X.; Long, X.; Jin, J.; Liang, J.; Weng, R.; Liu, J.; et al. A novel mechanism regulating pyroptosis-induced fibrosis in endometriosis via lnc-MALAT1/miR-141-3p/NLRP3 pathway. Biol. Reprod. 2023, 109, 156–171. [Google Scholar] [CrossRef]
- An, M.; Fu, X.; Meng, X.; Liu, H.; Ma, Y.; Li, Y.; Li, Q.; Chen, J. PI3K/AKT signaling pathway associates with pyroptosis and inflammation in patients with endometriosis. J. Reprod. Immunol. 2024, 162, 104213. [Google Scholar] [CrossRef]
- Hang, Y.; Tan, L.; Chen, Q.; Liu, Q.; Jin, Y. E3 ubiquitin ligase TRIM24 deficiency promotes NLRP3/caspase-1/IL-1β-mediated pyroptosis in endometriosis. Cell Biol. Int. 2021, 45, 1561–1570. [Google Scholar] [CrossRef]
- Han, S.J.; Jung, S.Y.; Wu, S.P.; Hawkins, S.M.; Park, M.J.; Kyo, S.; Lydon, J.P.; Tsai, S.Y.; Tsai, M.J.; DeMayo, F.J.; et al. Estrogen Receptor β Modulates Apoptosis Complexes and the Inflammasome to Drive the Pathogenesis of Endometriosis. Cell 2015, 163, 960–974. [Google Scholar] [CrossRef]
- Choi, J.; Jo, M.; Lee, E.; Kim, S.E.; Lee, D.Y.; Choi, D. Inhibition of the NLRP3 inflammasome by progesterone is attenuated by abnormal autophagy induction in endometriotic cyst stromal cells: Implications for endometriosis. Mol. Hum. Reprod. 2022, 28, gaac007. [Google Scholar] [CrossRef]
- Guo, B.; Zhu, H.; Xiao, C.; Zhang, J.; Liu, X.; Fang, Y.; Wei, B.; Zhang, J.; Cao, Y.; Zhan, L. NLRC5 exerts anti-endometriosis effects through inhibiting ERβ-mediated inflammatory response. BMC Med. 2024, 22, 351. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Yao, S.; Sun, S.; Su, Q.; Li, J.; Wei, B. NLRC5 and autophagy combined as possible predictors in patients with endometriosis. Fertil. Steril. 2018, 110, 949–956. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Liu, X.; Zhang, J.; Wang, Z.; Wang, W.; Fu, L.; Fan, Y.; Sun, S.; Cao, Y.; Zhan, L.; et al. NLRC5 Inhibits Inflammation of Secretory Phase Ectopic Endometrial Stromal Cells by Up-Regulating Autophagy in Ovarian Endometriosis. Front. Pharmacol. 2020, 11, 1281. [Google Scholar] [CrossRef]
- Yeo, S.G.; Won, Y.S.; Kim, S.H.; Park, D.C. Differences in C-type lectin receptors and their adaptor molecules in the peritoneal fluid of patients with endometriosis and gynecologic cancers. Int. J. Med. Sci. 2018, 15, 411–416. [Google Scholar] [CrossRef]
- Izumi, G.; Koga, K.; Takamura, M.; Makabe, T.; Nagai, M.; Urata, Y.; Harada, M.; Hirata, T.; Hirota, Y.; Fujii, T.; et al. Mannose receptor is highly expressed by peritoneal dendritic cells in endometriosis. Fertil. Steril. 2017, 107, 167–173.e2. [Google Scholar] [CrossRef]
- Wei, C.; Mei, J.; Tang, L.; Liu, Y.; Li, D.; Li, M.; Zhu, X. 1-Methyl-tryptophan attenuates regulatory T cells differentiation due to the inhibition of estrogen-IDO1-MRC2 axis in endometriosis. Cell Death Dis. 2016, 7, e2489. [Google Scholar] [CrossRef]
- Sopasi, F.; Spyropoulou, I.; Kourti, M.; Vasileiadis, S.; Tripsianis, G.; Galazios, G.; Koutlaki, N. Oxidative stress and female infertility: The role of follicular fluid soluble receptor of advanced glycation end-products (sRAGE) in women with endometriosis. Hum. Fertil. 2023, 26, 1400–1407. [Google Scholar] [CrossRef]
- Ajona, D.; Cragg, M.S.; Pio, R. The complement system in clinical oncology: Applications, limitations and challenges. Semin. Immunol. 2024, 77, 101921. [Google Scholar] [CrossRef]
- Mastellos, D.C.; Hajishengallis, G.; Lambris, J.D. A guide to complement biology, pathology and therapeutic opportunity. Nat. Rev. Immunol. 2024, 24, 118–141. [Google Scholar] [CrossRef]
- Zeller, J.M.; Henig, I.; Radwanska, E.; Dmowski, W.P. Enhancement of human monocyte and peritoneal macrophage chemiluminescence activities in women with endometriosis. Am. J. Reprod. Immunol. Microbiol. 1987, 13, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Lousse, J.C.; Defrère, S.; Van Langendonckt, A.; Gras, J.; González-Ramos, R.; Colette, S.; Donnez, J. Iron storage is significantly increased in peritoneal macrophages of endometriosis patients and correlates with iron overload in peritoneal fluid. Fertil. Steril. 2009, 91, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Y.; Zhong, Z.; Wei, C.; Liu, Y.; Zhu, X. Peritoneal immune microenvironment of endometriosis: Role and therapeutic perspectives. Front. Immunol. 2023, 14, 1134663. [Google Scholar] [CrossRef]
- Lousse, J.C.; Van Langendonckt, A.; González-Ramos, R.; Defrère, S.; Renkin, E.; Donnez, J. Increased activation of nuclear factor-kappa B (NF-kappaB) in isolated peritoneal macrophages of patients with endometriosis. Fertil. Steril. 2008, 90, 217–220. [Google Scholar] [CrossRef]
- Wu, M.H.; Sun, H.S.; Lin, C.C.; Hsiao, K.Y.; Chuang, P.C.; Pan, H.A.; Tsai, S.J. Distinct mechanisms regulate cyclooxygenase-1 and -2 in peritoneal macrophages of women with and without endometriosis. Mol. Hum. Reprod. 2002, 8, 1103–1110. [Google Scholar] [CrossRef]
- Lai, Z.Z.; Yang, H.L.; Ha, S.Y.; Chang, K.K.; Mei, J.; Zhou, W.J.; Qiu, X.M.; Wang, X.Q.; Zhu, R.; Li, D.J.; et al. Cyclooxygenase-2 in Endometriosis. Int. J. Biol. Sci. 2019, 15, 2783–2797. [Google Scholar] [CrossRef]
- Chan, R.W.S.; Lee, C.L.; Ng, E.H.Y.; Yeung, W.S.B. Co-culture with macrophages enhances the clonogenic and invasion activity of endometriotic stromal cells. Cell Prolif. 2017, 50, e12330. [Google Scholar] [CrossRef]
- Harada, T.; Kaponis, A.; Iwabe, T.; Taniguchi, F.; Makrydimas, G.; Sofikitis, N.; Paschopoulos, M.; Paraskevaidis, E.; Terakawa, N. Apoptosis in human endometrium and endometriosis. Hum. Reprod. Update 2004, 10, 29–38. [Google Scholar] [CrossRef]
- Huang, E.; Wang, X.; Chen, L. Regulated Cell Death in Endometriosis. Biomolecules 2024, 14, 142. [Google Scholar] [CrossRef]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4, 9. [Google Scholar] [CrossRef]
- Vallvé-Juanico, J.; Santamaria, X.; Vo, K.C.; Houshdaran, S.; Giudice, L.C. Macrophages display proinflammatory phenotypes in the eutopic endometrium of women with endometriosis with relevance to an infectious etiology of the disease. Fertil. Steril. 2019, 112, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Hudson, Q.J.; Ashjaei, K.; Perricos, A.; Kuessel, L.; Husslein, H.; Wenzl, R.; Yotova, I. Endometriosis Patients Show an Increased M2 Response in the Peritoneal CD14+low/CD68+low Macrophage Subpopulation Coupled with an Increase in the T-helper 2 and T-regulatory Cells. Reprod. Sci. 2020, 27, 1920–1931. [Google Scholar] [CrossRef] [PubMed]
- Henlon, Y.; Panir, K.; McIntyre, I.; Hogg, C.; Dhami, P.; Cuff, A.O.; Senior, A.; Moolchandani-Adwani, N.; Courtois, E.T.; Horne, A.W.; et al. Single-cell analysis identifies distinct macrophage phenotypes associated with prodisease and proresolving functions in the endometriotic niche. Proc. Natl. Acad. Sci. USA 2024, 121, e2405474121. [Google Scholar] [CrossRef]
- Laganà, A.S.; Salmeri, F.M.; Ban Frangež, H.; Ghezzi, F.; Vrtačnik-Bokal, E.; Granese, R. Evaluation of M1 and M2 macrophages in ovarian endometriomas from women affected by endometriosis at different stages of the disease. Gynecol. Endocrinol. 2020, 36, 441–444. [Google Scholar] [CrossRef]
- Duan, J.; Liu, X.; Wang, H.; Guo, S.W. The M2a macrophage subset may be critically involved in the fibrogenesis of endometriosis in mice. Reprod. Biomed. Online 2018, 37, 254–268. [Google Scholar] [CrossRef]
- Viganò, P.; Ottolina, J.; Bartiromo, L.; Bonavina, G.; Schimberni, M.; Villanacci, R.; Candiani, M. Cellular Components Contributing to Fibrosis in Endometriosis: A Literature Review. J. Minim. Invasive Gynecol. 2020, 27, 287–295. [Google Scholar] [CrossRef]
- Garmendia, J.V.; De Sanctis, J.B. A Brief Analysis of Tissue-Resident NK Cells in Pregnancy and Endometrial Diseases: The Importance of Pharmacologic Modulation. Immuno 2021, 1, 174–193. [Google Scholar] [CrossRef]
- Giuliani, E.; Parkin, K.L.; Lessey, B.A.; Young, S.L.; Fazleabas, A.T. Characterisation of uterine NK cells in women with infertility or recurrent pregnancy loss and associated endometriosis. Am. J. Reprod. Immunol. 2014, 72, 262–269. [Google Scholar] [CrossRef]
- Garmendia, J.V.; De Sanctis, C.V.; Hajdúch, M.; De Sanctis, J.B. Exploring the Immunological Aspects and Treatments of Recurrent Pregnancy Loss and Recurrent Implantation Failure. Int. J. Mol. Sci. 2025, 26, 1295. [Google Scholar] [CrossRef]
- Makoui, M.H.; Fekri, S.; Makoui, R.H.; Ansari, N.; Esmaeilzadeh, A. The Role of Mast Cells in the Development and Advancement of Endometriosis. Am. J. Reprod. Immunol. 2025, 93, e70019. [Google Scholar] [CrossRef]
- Suszczyk, D.; Skiba, W.; Jakubowicz-Gil, J.; Kotarski, J.; Wertel, I. The Role of Myeloid-Derived Suppressor Cells (MDSCs) in the Development and/or Progression of Endometriosis-State of the Art. Cells 2021, 10, 677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; He, Y.; Man, G.C.W.; Ding, Y.; Wang, C.C.; Chung, J.P.W. Myeloid-derived suppressor cells: A new emerging player in endometriosis. Int. Rev. Cell Mol. Biol. 2023, 375, 191–220. [Google Scholar] [CrossRef] [PubMed]
- Satake, E.; Koga, K.; Takamura, M.; Izumi, G.; Elsherbini, M.; Taguchi, A.; Makabe, T.; Takeuchi, A.; Harada, M.; Hirata, T.; et al. The roles of polymorphonuclear myeloid-derived suppressor cells in endometriosis. J. Reprod. Immunol. 2021, 148, 103371. [Google Scholar] [CrossRef]
- Chen, H.; Qin, S.; Lei, A.; Li, X.; Gao, Q.; Dong, J.; Xiao, Q.; Zhou, J. Expansion of monocytic myeloid-derived suppressor cells in endometriosis patients: A pilot study. Int. Immunopharmacol. 2017, 47, 150–158. [Google Scholar] [CrossRef]
- Sun, Y.; Shao, J.; Jiang, F.; Wang, Y.; Yan, Q.; Yu, N.; Zhang, J.; Zhang, J.; Li, M.; He, Y. CD33+ CD14+ CD11b+ HLA-DR- monocytic myeloid-derived suppressor cells recruited and activated by CCR9/CCL25 are crucial for the pathogenic progression of endometriosis. Am. J. Reprod. Immunol. 2019, 81, e13067. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, K.; Xu, Y.; Guo, P.; Hong, B.; Cao, Y.; Wei, Z.; Xue, R.; Wang, C.; Jiang, H. Alteration of Myeloid-Derived Suppressor Cells, Chronic Inflammatory Cytokines, and Exosomal miRNA Contribute to the Peritoneal Immune Disorder of Patients With Endometriosis. Reprod. Sci. 2019, 26, 1130–1138. [Google Scholar] [CrossRef]
- Guo, P.; Bi, K.; Lu, Z.; Wang, K.; Xu, Y.; Wu, H.; Cao, Y.; Jiang, H. CCR5/CCR5 ligand-induced myeloid-derived suppressor cells are related to the progression of endometriosis. Reprod. Biomed. Online 2019, 39, 704–711. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, J.; Man, G.C.W.; Leung, K.T.; Liang, B.; Xiao, B.; Ma, X.; Huang, S.; Huang, H.; Hegde, V.L.; et al. MDSCs drive the process of endometriosis by enhancing angiogenesis and are a new potential therapeutic target. Eur. J. Immunol. 2018, 48, 1059–1073. [Google Scholar] [CrossRef]
- Bosteels, V.; Janssens, S. Striking a balance: New perspectives on homeostatic dendritic cell maturation. Nat. Rev. Immunol. 2025, 25, 125–140. [Google Scholar] [CrossRef]
- Li, W.; Lin, A.; Qi, L.; Lv, X.; Yan, S.; Xue, J.; Mu, N. Immunotherapy: A promising novel endometriosis therapy. Front. Immunol. 2023, 14, 1128301. [Google Scholar] [CrossRef]
- Rahal, D.; Andrade, F.; Nisihara, R. Insights into the role of complement system in the pathophysiology of endometriosis. Immunol. Lett. 2021, 231, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Balduit, A.; Mangogna, A.; Zito, G.; Romano, F.; Ricci, G.; Kishore, U.; Bulla, R. Immunological Basis of the Endometriosis: The Complement System as a Potential Therapeutic Target. Front. Immunol. 2021, 11, 599117. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Toffoli, M.; Zito, G.; Balduit, A.; Pegoraro, S.; Spazzapan, M.; Pascolo, L.; Romano, F.; Di Lorenzo, G.; Mangogna, A.; et al. Proangiogenic properties of complement protein C1q can contribute to endometriosis. Front. Immunol. 2024, 15, 1405597. [Google Scholar] [CrossRef]
- Suryawanshi, S.; Huang, X.; Elishaev, E.; Budiu, R.A.; Zhang, L.; Kim, S.; Donnellan, N.; Mantia-Smaldone, G.; Ma, T.; Tseng, G.; et al. Complement pathway is frequently altered in endometriosis and endometriosis-associated ovarian cancer. Clin. Cancer Res. 2014, 20, 6163–6174. [Google Scholar] [CrossRef]
- Abramiuk, M.; Grywalska, E.; Małkowska, P.; Sierawska, O.; Hrynkiewicz, R.; Niedźwiedzka-Rystwej, P. The Role of the Immune System in the Development of Endometriosis. Cells 2022, 11, 2028. [Google Scholar] [CrossRef]
- Milewski, Ł.; Dziunycz, P.; Barcz, E.; Radomski, D.; Roszkowski, P.I.; Korczak-Kowalska, G.; Kamiński, P.; Malejczyk, J. Increased levels of human neutrophil peptides 1, 2, and 3 in peritoneal fluid of patients with endometriosis: Association with neutrophils, T cells and IL-8. J. Reprod. Immunol. 2011, 91, 64–70. [Google Scholar] [CrossRef]
- Lukács, L.; Kovács, A.R.; Pál, L.; Szűcs, S.; Kövér, Á.; Lampé, R. Phagocyte function of peripheral neutrophil granulocytes and monocytes in endometriosis before and after surgery. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 101796. [Google Scholar] [CrossRef]
- Wilson, T.R.; Kasper, S.; Burns, K.A. An emerging role for neutrophils in the pathogenesis of endometriosis. npj Women’s Health 2025, 3, 9. [Google Scholar] [CrossRef]
- Takamura, M.; Koga, K.; Izumi, G.; Urata, Y.; Nagai, M.; Hasegawa, A.; Harada, M.; Hirata, T.; Hirota, Y.; Wada-Hiraike, O.; et al. Neutrophil depletion reduces endometriotic lesion formation in mice. Am. J. Reprod. Immunol. 2016, 76, 193–198. [Google Scholar] [CrossRef]
- Hogg, C.; Horne, A.W.; Greaves, E. Endometriosis-associated macrophages: Origin, phenotype, and function. Front. Endocrinol. 2020, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Hogg, C.; Panir, K.; Dhami, P.; Rosser, M.; Mack, M.; Soong, D.; Pollard, J.W.; Jenkins, S.J.; Horne, A.W.; Greaves, E. Macrophages inhibit and enhance endometriosis depending on their origin. Proc. Natl. Acad. Sci. USA 2021, 118, e2013776118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Liu, Y.K.; Hu, W.T.; Tang, L.L.; Sheng, Y.R.; Wei, C.Y.; Li, M.Q.; Zhu, X.Y. Elevated heme impairs macrophage phagocytosis in endometriosis. Reproduction 2019, 158, 257–266. [Google Scholar] [CrossRef]
- Gao, X.; Gao, H.; Shao, W.; Wang, J.; Li, M.; Liu, S. The Extracellular Vesicle-Macrophage Regulatory Axis: A Novel Pathogenesis for Endometriosis. Biomolecules 2023, 13, 1376. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Zamora, M.A.; Armengol-Badia, O.; Quintas-Marquès, L.; Carmona, F.; Closa, D. Macrophage Phenotype Induced by Circulating Small Extracellular Vesicles from Women with Endometriosis. Biomolecules 2024, 14, 737. [Google Scholar] [CrossRef]
- Chuang, P.C.; Lin, Y.J.; Wu, M.H.; Wing, L.Y.; Shoji, Y.; Tsai, S.J. Inhibition of CD36-dependent phagocytosis by prostaglandin E2 contributes to the development of endometriosis. Am. J. Pathol. 2010, 176, 850–860. [Google Scholar] [CrossRef]
- Weng, L.C.; Hou, S.H.; Lei, S.T.; Peng, H.Y.; Li, M.Q.; Zhao, D. Estrogen-regulated CD200 inhibits macrophage phagocytosis in endometriosis. J. Reprod. Immunol. 2020, 138, 103090. [Google Scholar] [CrossRef]
- Shiraishi, T.; Ikeda, M.; Watanabe, T.; Negishi, Y.; Ichikawa, G.; Kaseki, H.; Akira, S.; Morita, R.; Suzuki, S. Downregulation of pattern recognition receptors on macrophages involved in aggravation of endometriosis. Am. J. Reprod. Immunol. 2024, 91, e13812. [Google Scholar] [CrossRef]
- Symons, L.K.; Miller, J.E.; Kay, V.R.; Marks, R.M.; Liblik, K.; Koti, M.; Tayade, C. The Immunopathophysiology of Endometriosis. Trends Mol. Med. 2018, 24, 748–762. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Tang, Y.; Xia, Z. Transformation of macrophages into myofibroblasts in fibrosis-related diseases: Emerging biological concepts and potential mechanism. Front. Immunol. 2024, 15, 1474688. [Google Scholar] [CrossRef]
- Tuckerman, E.; Mariee, N.; Prakash, A.; Li, T.C.; Laird, S. Uterine natural killer cells in peri-implantation endometrium from women with repeated implantation failure after IVF. J. Reprod. Immunol. 2010, 87, 60–66. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Ishikawa, N.; Hirata, J.; Imaizumi, E.; Sasa, H.; Nagata, I. Changes of peripheral blood lymphocyte subsets before and after operation of patients with endometriosis. Acta Obstet. Gynecol. Scand. 1993, 72, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Azeze, G.G.; Wu, L.; Alemu, B.K.; Wang, C.C.; Zhang, T. Changes in the number and activity of natural killer cells and its clinical association with endometriosis: Systematic review and meta-analysis. F&S Rev. 2024, 5, 100072. [Google Scholar] [CrossRef]
- Jeung, I.; Cheon, K.; Kim, M.R. Decreased Cytotoxicity of Peripheral and Peritoneal Natural Killer Cell in Endometriosis. Biomed. Res. Int. 2016, 2016, 2916070. [Google Scholar] [CrossRef]
- González-Foruria, I.; Santulli, P.; Chouzenoux, S.; Carmona, F.; Batteux, F.; Chapron, C. Soluble ligands for the NKG2D receptor are released during endometriosis and correlate with disease severity. PLoS ONE 2015, 10, e0119961. [Google Scholar] [CrossRef]
- Kang, Y.J.; Jeung, I.C.; Park, A.; Park, Y.J.; Jung, H.; Kim, T.D.; Lee, H.G.; Choi, I.; Yoon, S.R. An increased level of IL-6 suppresses NK cell activity in peritoneal fluid of patients with endometriosis via regulation of SHP-2 expression. Hum. Reprod. 2014, 29, 2176–2189. [Google Scholar] [CrossRef]
- Guo, S.W.; Du, Y.; Liu, X. Platelet-derived TGF-β1 mediates the down-modulation of NKG2D expression and may be responsible for impaired natural killer (NK) cytotoxicity in women with endometriosis. Hum. Reprod. 2016, 31, 1462–1474. [Google Scholar] [CrossRef]
- Yu, J.J.; Sun, H.T.; Zhang, Z.F.; Shi, R.X.; Liu, L.B.; Shang, W.Q.; Wei, C.Y.; Chang, K.K.; Shao, J.; Wang, M.Y.; et al. IL15 promotes growth and invasion of endometrial stromal cells and inhibits killing activity of NK cells in endometriosis. Reproduction 2016, 152, 151–160. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhou, W.J.; Chang, K.K.; Mei, J.; Huang, L.Q.; Wang, M.Y.; Meng, Y.; Ha, S.Y.; Li, D.J.; Li, M.Q. The crosstalk between endometrial stromal cells and macrophages impairs cytotoxicity of NK cells in endometriosis by secreting IL-10 and TGF-β. Reproduction 2017, 154, 815–825. [Google Scholar] [CrossRef]
- Reis, J.L.; Rosa, N.N.; Ângelo-Dias, M.; Martins, C.; Borrego, L.M.; Lima, J. Natural Killer Cell Receptors and Endometriosis: A Systematic Review. Int. J. Mol. Sci. 2022, 24, 331. [Google Scholar] [CrossRef]
- Saeki, S.; Fukui, A.; Mai, C.; Takeyama, R.; Yamaya, A.; Shibahara, H. Co-expression of activating and inhibitory receptors on peritoneal fluid NK cells in women with endometriosis. J. Reprod. Immunol. 2023, 155, 103765. [Google Scholar] [CrossRef]
- Sugamata, M.; Ihara, T.; Uchiide, I. Increase of activated mast cells in human endometriosis. Am. J. Reprod. Immunol. 2005, 53, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, D.; Kaulfuss, S.; Fuhrmann, U.; Maurer, M.; Zollner, T.M. Mast cells in endometriosis: Guilty or innocent bystanders? Expert Opin. Ther. Targets 2012, 16, 237–241. [Google Scholar] [CrossRef] [PubMed]
- McCallion, A.; Nasirzadeh, Y.; Lingegowda, H.; Miller, J.E.; Khalaj, K.; Ahn, S.; Monsanto, S.P.; Bidarimath, M.; Sisnett, D.J.; Craig, A.W.; et al. Estrogen mediates inflammatory role of mast cells in endometriosis pathophysiology. Front. Immunol. 2022, 13, 961599. [Google Scholar] [CrossRef]
- Xu, X.; Wang, J.; Guo, X.; Chen, Y.; Ding, S.; Zou, G.; Zhu, L.; Li, T.; Zhang, X. GPR30-mediated non-classic estrogen pathway in mast cells participates in endometriosis pain via the production of FGF2. Front. Immunol. 2023, 14, 1106771. [Google Scholar] [CrossRef]
- Schulke, L.; Berbic, M.; Manconi, F.; Tokushige, N.; Markham, R.; Fraser, I.S. Dendritic cell populations in the eutopic and ectopic endometrium of women with endometriosis. Hum. Reprod. 2009, 24, 1695–1703. [Google Scholar] [CrossRef]
- Qiaomei, Z.; Ping, W.; Yanjing, Z.; Jinhua, W.; Shaozhan, C.; Lihong, C. Features of peritoneal dendritic cells in the development of endometriosis. Reprod. Biol. Endocrinol. 2023, 21, 4. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, L.; Xu, Y. HSD11B1 overexpression in dendritic cells and stromal cells relates to endometriosis by inhibiting dendritic cell proliferation and maturation. Gynecol. Endocrinol. 2024, 40, 2411607. [Google Scholar] [CrossRef]
- Maridas, D.E.; Hey-Cunningham, A.J.; Ng, C.H.M.; Markham, R.; Fraser, I.S.; Berbic, M. Peripheral and endometrial dendritic cell populations during the normal cycle and in the presence of endometriosis. J. Endometr. Pelvic Pain Disord. 2014, 6, 67–119. [Google Scholar] [CrossRef]
- Suen, J.L.; Chang, Y.; Shiu, Y.S.; Hsu, C.Y.; Sharma, P.; Chiu, C.C.; Chen, Y.J.; Hour, T.C.; Tsai, E.M. IL-10 from plasmacytoid dendritic cells promotes angiogenesis in the early stage of endometriosis. J. Pathol. 2019, 249, 485–497. [Google Scholar] [CrossRef]
- Knez, J.; Kovačič, B.; Goropevšek, A. The role of regulatory T-cells in the development of endometriosis. Hum. Reprod. 2024, 39, 1367–1380. [Google Scholar] [CrossRef]
- Riccio, L.G.C.; Baracat, E.C.; Chapron, C.; Batteux, F.; Abrão, M.S. The role of the B lymphocytes in endometriosis: A systematic review. J. Reprod. Immunol. 2017, 123, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Kisovar, A.; Becker, C.M.; Granne, I.; Southcombe, J.H. The role of CD8+ T cells in endometriosis: A systematic review. Front. Immunol. 2023, 14, 1225639. [Google Scholar] [CrossRef] [PubMed]
- Chopyak, V.V.; Koval, H.D.; Havrylyuk, A.M.; Lishchuk-Yakymovych, K.A.; Potomkina, H.A.; Kurpisz, M.K. Immunopathogenesis of endometriosis—A novel look at an old problem. Cent. Eur. J. Immunol. 2022, 47, 109–116. [Google Scholar] [CrossRef]
- Hanada, T.; Tsuji, S.; Nakayama, M.; Wakinoue, S.; Kasahara, K.; Kimura, F.; Mori, T.; Ogasawara, K.; Murakami, T. Suppressive regulatory T cells and latent transforming growth factor-β-expressing macrophages are altered in the peritoneal fluid of patients with endometriosis. Reprod. Biol. Endocrinol. 2018, 16, 9. [Google Scholar] [CrossRef]
- Riccio, L.G.C.; Andres, M.P.; Dehó, I.Z.; Fontanari, G.O.; Abrão, M.S. Foxp3+CD39+CD73+ regulatory T-cells are decreased in the peripheral blood of women with deep infiltrating endometriosis. Clinics 2024, 79, 100390. [Google Scholar] [CrossRef]
- Li, M.Q.; Wang, Y.; Chang, K.K.; Meng, Y.H.; Liu, L.B.; Mei, J.; Wang, Y.; Wang, X.Q.; Jin, L.P.; Li, D.J. CD4+Foxp3+ regulatory T cell differentiation mediated by endometrial stromal cell-derived TECK promotes the growth and invasion of endometriotic lesions. Cell Death Dis. 2014, 5, e1436. [Google Scholar] [CrossRef]
- Sisnett, D.J.; Zutautas, K.B.; Miller, J.E.; Lingegowda, H.; Ahn, S.H.; McCallion, A.; Bougie, O.; Lessey, B.A.; Tayade, C. The Dysregulated IL-23/TH17 Axis in Endometriosis Pathophysiology. J. Immunol. 2024, 212, 1428–1441. [Google Scholar] [CrossRef]
- Shi, J.; Xu, Q.; Yu, S.; Zhang, T. Perturbations of the endometrial immune microenvironment in endometriosis and adenomyosis: Their impact on reproduction and pregnancy. Semin. Immunopathol. 2025, 47, 16. [Google Scholar] [CrossRef]
- Olkowska-Truchanowicz, J.; Białoszewska, A.; Zwierzchowska, A.; Sztokfisz-Ignasiak, A.; Janiuk, I.; Dąbrowski, F.; Korczak-Kowalska, G.; Barcz, E.; Bocian, K.; Malejczyk, J. Peritoneal Fluid from Patients with Ovarian Endometriosis Displays Immunosuppressive Potential and Stimulates Th2 Response. Int. J. Mol. Sci. 2021, 22, 8134. [Google Scholar] [CrossRef]
- Reis, J.L.; Rosa, N.N.; Martins, C.; Ângelo-Dias, M.; Borrego, L.M.; Lima, J. The Role of NK and T Cells in Endometriosis. Int. J. Mol. Sci. 2024, 25, 10141. [Google Scholar] [CrossRef]
- Schmitz, T.; Hoffmann, V.; Olliges, E.; Bobinger, A.; Popovici, R.; Nößner, E.; Meissner, K. Reduced frequency of perforin-positive CD8+ T cells in menstrual effluent of endometriosis patients. J. Reprod. Immunol. 2021, 148, 103424. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, R.; Moini, A.; Hosseini, R.; Fatehnejad, M.; Yekaninejad, M.S.; Javidan, M.; Changaei, M.; Feizisani, F.; Rajaei, S. A higher number of exhausted local PD1+, but not TIM3+, NK cells in advanced endometriosis. Heliyon 2023, 10, e23294. [Google Scholar] [CrossRef] [PubMed]
- Abramiuk, M.; Bębnowska, D.; Hrynkiewicz, R.; Polak, P.N.G.; Kotarski, J.; Roliński, J.; Grywalska, E. CLTA-4 Expression is Associated with the Maintenance of Chronic Inflammation in Endometriosis and Infertility. Cells 2021, 10, 487. [Google Scholar] [CrossRef]
- Podgaec, S.; Abrao, M.S.; Días, J.A., Jr.; Rizzo, L.V.; de Oliveira, R.M.; Baracat, E.C. Endometriosis: An inflammatory disease with a Th2 immune response component. Hum. Reprod. 2007, 22, 1373–1379. [Google Scholar] [CrossRef]
- Santoso, B.; Sa’adi, A.; Dwiningsih, S.R.; Tunjungseto, A.; Widyanugraha, M.Y.A.; Mufid, A.F.; Rahmawati, N.Y.; Ahsan, F. Soluble immune checkpoints CTLA-4, HLA-G, PD-1, and PD-L1 are associated with endometriosis-related infertility. Am. J. Reprod. Immunol. 2020, 84, e13296. [Google Scholar] [CrossRef]
- Chen, P.; Wang, D.B.; Liang, Y.M. Evaluation of estrogen in endometriosis patients: Regulation of GATA-3 in endometrial cells and effects on Th2 cytokines. J. Obstet. Gynaecol. Res. 2016, 42, 669–677. [Google Scholar] [CrossRef]
- Lin, K.R.; Li, P.X.; Zhu, X.H.; Mao, X.F.; Peng, J.L.; Chen, X.P.; SiTu, C.Y.; Zhang, L.F.; Luo, W.; Han, Y.B.; et al. Peripheral immune characteristics and subset disorder in reproductive females with endometriosis. Front. Immunol. 2024, 15, 1431175. [Google Scholar] [CrossRef]
- Delbandi, A.A.; Mahmoudi, M.; Shervin, A.; Farhangnia, P.; Mohammadi, T.; Zarnani, A.H. Increased circulating T helper 17 (TH17) cells and endometrial tissue IL-17-producing cells in patients with endometriosis compared with non-endometriotic subjects. Reprod. Biol. 2025, 25, 101019, Advance online publication. [Google Scholar] [CrossRef]
- Kitaya, K. B Cell Lineage in the Human Endometrium: Physiological and Pathological Implications. Cells 2025, 14, 648. [Google Scholar] [CrossRef]
- Slawek, A.; Lorek, D.; Kedzierska, A.E.; Kubik, P.; Pajak, J.; Chrobak, A.; Chelmonska-Soyta, A. Peripheral blood subpopulations of Bregs producing IL-35 in women with endometriosis. Am. J. Reprod. Immunol. 2023, 89, e13675. [Google Scholar] [CrossRef]
- Ren, Y.; Zhu, D.; Han, X.; Zhang, Q.; Chen, B.; Zhou, P.; Wei, Z.; Zhang, Z.; Cao, Y.; Zou, H. HMGB1: A double-edged sword and therapeutic target in the female reproductive system. Front. Immunol. 2023, 14, 1238785. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, X.; Liu, J. High mobility group box 1 promotes endometriosis under hypoxia by regulating inflammation and autophagy in vitro and in vivo. Int. Immunopharmacol. 2024, 127, 111397. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Guo, R.; Na, X.; Jiang, S.; Liang, J.; Guo, C.; Fang, Y.; Na, Z.; Li, D. Hypoxia and the endometrium: An indispensable role for HIF-1α as therapeutic strategies. Redox Biol. 2024, 73, 103205. [Google Scholar] [CrossRef]
- Huang, Y.; Li, R.; Hu, R.; Yao, J.; Yang, Y. PEG2-Induced Pyroptosis Regulates the Expression of HMGB1 and Promotes hEM15A Migration in Endometriosis. Int. J. Mol. Sci. 2022, 23, 11707. [Google Scholar] [CrossRef]
- Jana, B.; Andronowska, A.; Całka, J.; Mówińska, A. Biosynthetic pathway for leukotrienes is stimulated by lipopolysaccharide and cytokines in pig endometrial stromal cells. Sci. Rep. 2025, 15, 2806. [Google Scholar] [CrossRef]
- Ihara, T.; Uchiide, I.; Sugamata, M. Light and electron microscopic evaluation of antileukotriene therapy for experimental rat endometriosis. Fertil. Steril. 2004, 81 (Suppl. S1), 819–823. [Google Scholar] [CrossRef]
- Kiykac Altinbas, S.; Tapisiz, O.L.; Cavkaytar, S.; Simsek, G.; Oguztuzun, S.; Goktolga, U. Is montelukast effective in regression of endometrial implants in an experimentally induced endometriosis model in rats? Europ. J. Obstet. Gynecol. Reprod. Biol. 2015, 184, 7–12. [Google Scholar] [CrossRef]
- Wu, R.; Zhou, W.; Chen, S.; Shi, Y.; Su, L.; Zhu, M.; Chen, Q.; Chen, Q. Lipoxin A4 suppresses the development of endometriosis in an ALX receptor-dependent manner via the p38 MAPK pathway. Br. J. Pharmacol. 2014, 171, 4927–4940. [Google Scholar] [CrossRef]
- Huang, Z.X.; He, X.R.; Ding, X.Y.; Chen, J.H.; Lei, Y.H.; Bai, J.B.; Lin, D.C.; Hong, Y.H.; Lan, J.F.; Chen, Q.H. Lipoxin A4 depresses inflammation and promotes autophagy via AhR/mTOR/AKT pathway to suppress endometriosis. Am. J. Reprod. Immunol. 2023, 89, e13659. [Google Scholar] [CrossRef]
- Dmitrieva, N.; Suess, G.; Shirley, R. Resolvins RvD1 and 17(R)-RvD1 alleviate signs of inflammation in a rat model of endometriosis. Fertil. Steril. 2014, 102, 1191–1196. [Google Scholar] [CrossRef]
- Gu, Z.; Lamont, G.J.; Lamont, R.J.; Uriarte, S.M.; Wang, H.; Scott, D.A. Resolvin D1, resolvin D2 and maresin 1 activate the GSK3β anti-inflammatory axis in TLR4-engaged human monocytes. Innate Immun. 2016, 622, 186–195. [Google Scholar] [CrossRef] [PubMed]
- de Fáveri, C.; Fermino, P.M.P.; Piovezan, A.P.; Volpato, L.K. The Inflammatory Role of Pro-Resolving Mediators in Endometriosis: An Integrative Review. Int. J. Mol. Sci. 2021, 22, 4370. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Castillo, M.; Ortega, Á.; Cudris-Torres, L.; Duran, P.; Rojas, M.; Manzano, A.; Garrido, B.; Salazar, J.; Silva, A.; Rojas-Gomez, D.M.; et al. Specialized Pro-Resolving Lipid Mediators: The Future of Chronic Pain Therapy? Int. J. Mol. Sci. 2021, 22, 10370. [Google Scholar] [CrossRef]
- Collie, B.; Troisi, J.; Lombardi, M.; Symes, S.; Richards, S. The Current Applications of Metabolomics in Understanding Endometriosis: A Systematic Review. Metabolites 2025, 15, 50. [Google Scholar] [CrossRef]
- Wilson, R.B. Hypoxia, cytokines and stromal recruitment: Parallels between pathophysiology of encapsulating peritoneal sclerosis, endometriosis and peritoneal metastasis. Pleura Peritoneum 2018, 3, 20180103. [Google Scholar] [CrossRef]
- Ni, C.; Li, D. Ferroptosis and oxidative stress in endometriosis: A systematic review of the literature. Medicine 2024, 103, e37421. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, X.; Yang, L.; He, Q.; Huang, D.; Tan, X. Ferroptosis and tumor immunity: In perspective of the major cell components in the tumor microenvironment. Eur. J. Pharmacol. 2023, 961, 176124. [Google Scholar] [CrossRef]
- Kondera-Anasz, Z.; Sikora, J.; Mielczarek-Palacz, A.; Jońca, M. Concentrations of interleukin (IL)-1alpha, IL-1 soluble receptor type II (IL-1 sRII) and IL-1 receptor antagonist (IL-1 Ra) in the peritoneal fluid and serum of infertile women with endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 123, 198–203. [Google Scholar] [CrossRef]
- Sikora, J.; Ferrero, S.; Mielczarek-Palacz, A.; Kondera-Anasz, Z. The Delicate Balance between the Good and the Bad IL-1 Proinflammatory Effects in Endometriosis. Curr. Med. Chem. 2018, 25, 2105–2121. [Google Scholar] [CrossRef]
- Malvezzi, H.; Hernandes, C.; Piccinato, C.A.; Podgaec, S. Interleukin in endometriosis-associated infertility-pelvic pain: Systematic review and meta-analysis. Reproduction 2019, 158, 1–12. [Google Scholar] [CrossRef]
- Werdel, R.; Mabie, A.; Evans, T.L.; Coté, R.D.; Schlundt, A.; Doehrman, P.; Dilsaver, D.; Coté, J.J. Serum Levels of Interleukins in Endometriosis Patients: A Systematic Review and Meta-analysis. J. Minim. Invasive Gynecol. 2024, 31, 387–396.e11. [Google Scholar] [CrossRef] [PubMed]
- Koumantakis, E.; Matalliotakis, I.; Neonaki, M.; Froudarakis, G.; Georgoulias, V. Soluble serum interleukin-2 receptor, interleukin-6 and interleukin-1a in patients with endometriosis and in controls. Arch. Gynecol. Obstet. 1994, 255, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Guo, S.W.; Ogawa, K.; Fujishita, A.; Mori, T. The role of innate and adaptive immunity in endometriosis. J. Reprod. Immunol. 2024, 163, 104242. [Google Scholar] [CrossRef]
- Xia, T.; Zeng, K.; Peng, Q.; Wu, X.; Lei, X. Clinical significance of serum Th1/Th2 cytokines in patients with endometriosis. Women Health 2023, 63, 73–82. [Google Scholar] [CrossRef]
- Voltolini Velho, R.; Halben, N.; Chekerov, R.; Keye, J.; Plendl, J.; Sehouli, J.; Mechsner, S. Functional changes of immune cells: Signal of immune tolerance of the ectopic lesions in endometriosis? Reprod. Biomed. Online 2021, 43, 319–328. [Google Scholar] [CrossRef]
- Mao, X.D.; Hu, C.Y.; Zhu, M.C.; Ou, H.L.; Qian, Y.L. Immunological microenvironment alterations in follicles of women with proven severe endometriosis undergoing in vitro fertilization. Mol. Biol. Rep. 2019, 46, 4675–4684. [Google Scholar] [CrossRef]
- Oală, I.E.; Mitranovici, M.I.; Chiorean, D.M.; Irimia, T.; Crișan, A.I.; Melinte, I.M.; Cotruș, T.; Tudorache, V.; Moraru, L.; Moraru, R.; et al. Endometriosis and the Role of Pro-Inflammatory and Anti-Inflammatory Cytokines in Pathophysiology: A Narrative Review of the Literature. Diagnostics 2024, 14, 312. [Google Scholar] [CrossRef]
- Ghodsi, M.; Hojati, V.; Attaranzadeh, A.; Saifi, B. Evaluation of IL-3, IL-5, and IL-6 concentration in the follicular fluid of women with endometriosis: A cross-sectional study. Int. J. Reprod. Biomed. 2022, 20, 213–220. [Google Scholar] [CrossRef]
- Pellicer, A.; Albert, C.; Mercader, A.; Bonilla-Musoles, F.; Remohí, J.; Simón, C. The follicular and endocrine environment in women with endometriosis: Local and systemic cytokine production. Fertil. Steril. 1998, 70, 425–431. [Google Scholar] [CrossRef]
- Monsanto, S.P.; Edwards, A.K.; Zhou, J.; Nagarkatti, P.; Nagarkatti, M.; Young, S.L.; Lessey, B.A.; Tayade, C. Surgical removal of endometriotic lesions alters local and systemic proinflammatory cytokines in endometriosis patients. Fertil. Steril. 2016, 105, 968–977.e5. [Google Scholar] [CrossRef]
- Bellelis, P.; Frediani Barbeiro, D.; Gueuvoghlanian-Silva, B.Y.; Kalil, J.; Abrão, M.S.; Podgaec, S. Interleukin-15 and Interleukin-7 are the Major Cytokines to Maintain Endometriosis. Gynecol. Obstet. Investig. 2019, 84, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Akoum, A.; Lawson, C.; McColl, S.; Villeneuve, M. Ectopic endometrial cells express high concentrations of interleukin (IL)-8 in vivo regardless of the menstrual cycle phase and respond to oestradiol by up-regulating IL-1-induced IL-8 expression in vitro. Mol. Hum. Reprod. 2001, 7, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Sikora, J.; Smycz-Kubańska, M.; Mielczarek-Palacz, A.; Kondera-Anasz, Z. Abnormal peritoneal regulation of chemokine activation-The role of IL-8 in pathogenesis of endometriosis. Am. J. Reprod. Immunol. 2017, 77, e12622. [Google Scholar] [CrossRef] [PubMed]
- Punnonen, J.; Teisala, K.; Ranta, H.; Bennett, B.; Punnonen, R. Increased levels of interleukin-6 and interleukin-10 in the peritoneal fluid of patients with endometriosis. Am. J. Obstet. Gynecol. 1996, 174, 1522–1526. [Google Scholar] [CrossRef]
- Mazzeo, D.; Viganó, P.; Di Blasio, A.M.; Sinigaglia, F.; Vignali, M.; Panina-Bordignon, P. Interleukin-12 and its free p40 subunit regulate immune recognition of endometrial cells: Potential role in endometriosis. J. Clin. Endocrinol. Metab. 1998, 83, 911–916. [Google Scholar] [CrossRef]
- Rahmawati, N.Y.; Ahsan, F.; Santoso, B.; Mufid, A.F.; Sa’adi, A.; Dwiningsih, S.R.; Tunjungseto, A.; Widyanugraha, M.Y.A. IL-8 and IL-12p70 are associated with pelvic pain among infertile women with endometriosis. Pain Med. 2023, 24, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Chegini, N.; Roberts, M.; Ripps, B. Differential expression of interleukins (IL)-13 and IL-15 in ectopic and eutopic endometrium of women with endometriosis and normal fertile women. Am. J. Reprod. Immunol. 2003, 49, 75–83. [Google Scholar] [CrossRef]
- Bailey, A.P.; Hill, A.S.; Beste, M.T.; Cook, C.D.; Sarda, V.; Laufer, M.R.; Isaacson, K.B.; Griffith, L.G.; Missmer, S.A. Comparison of cytokines in the peritoneal fluid and conditioned medium of adolescents and adults with and without endometriosis. Am. J. Reprod. Immunol. 2021, 85, e13347. [Google Scholar] [CrossRef]
- Koga, K.; Osuga, Y.; Yoshino, O.; Hirota, Y.; Yano, T.; Tsutsumi, O.; Taketani, Y. Elevated interleukin-16 levels in the peritoneal fluid of women with endometriosis may be a mechanism for inflammatory reactions associated with endometriosis. Fertil. Steril. 2005, 83, 878–882. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, W.; Zhou, Y.; Xi, S.; Xu, X.; Du, X.; Zheng, X.; Hu, W.; Sun, R.; Tian, Z.; et al. Pyroptotic T cell-derived active IL-16 has a driving function in ovarian endometriosis development. Cell Rep. Med. 2024, 5, 101476. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, H.; Lin, J.; Qian, Y.; Deng, L. Peritoneal fluid concentrations of interleukin-17 correlate with the severity of endometriosis and infertility of this disorder. BJOG Int. J. Obstet. Gynaecol. 2005, 112, 1153–1155. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Edwards, A.K.; Singhm, S.S.; Young, S.L.; Lessey, B.A.; Tayade, C. IL-17A Contributes to the Pathogenesis of Endometriosis by Triggering Proinflammatory Cytokines and Angiogenic Growth Factors. J. Immunol. 2015, 195, 2591–2600. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.L.; Zheng, Z.M.; Chen, M.; Shen, H.H.; Li, M.Q.; Shao, J. IL-17: An important pathogenic factor in endometriosis. Int. J. Med. Sci. 2022, 19, 769–778. [Google Scholar] [CrossRef]
- Arici, A.; Matalliotakis, I.; Goumenou, A.; Koumantakis, G.; Vassiliadis, S.; Mahutte, N.G. Altered expression of interleukin-18 in the peritoneal fluid of women with endometriosis. Fertil. Steril. 2003, 80, 889–894. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Chen, G.Y.; Liu, Y.; Huang, H.J.; Song, Y.F. Relationship between resistin and IL-23 levels in follicular fluid in infertile patients with endometriosis undergoing IVF-ET. Adv. Clin. Exp. Med. 2017, 26, 1431–1435. [Google Scholar] [CrossRef]
- Bungum, H.F.; Nygaard, U.; Vestergaard, C.; Martensen, P.M.; Knudsen, U.B. Increased IL-25 levels in the peritoneal fluid of patients with endometriosis. J. Reprod. Immunol. 2016, 114, 6–9. [Google Scholar] [CrossRef]
- O, D.; Waelkens, E.; Vanhie, A.; Peterse, D.; Fassbender, A.; D’Hooghe, T. The Use of Antibody Arrays in the Discovery of New Plasma Biomarkers for Endometriosis. Reprod. Sci. 2020, 27, 751–762. [Google Scholar] [CrossRef]
- Lee, M.Y.; Kim, S.H.; Oh, Y.S.; Heo, S.H.; Kim, K.H.; Chae, H.D.; Kim, C.H.; Kang, B.M. Role of interleukin-32 in the pathogenesis of endometriosis: In vitro, human and transgenic mouse data. Hum. Reprod. 2018, 33, 807–816. [Google Scholar] [CrossRef]
- Choi, Y.S.; Kim, S.; Oh, Y.S.; Cho, S.; Hoon Kim, S. Elevated serum interleukin-32 levels in patients with endometriosis: A cross-sectional study. Am. J. Reprod. Immunol. 2019, 82, e13149. [Google Scholar] [CrossRef]
- Lin, K.; Ma, J.; Peng, Y.; Sun, M.; Xu, K.; Wu, R.; Lin, J. Autocrine Production of Interleukin-34 Promotes the Development of Endometriosis through CSF1R/JAK3/STAT6 signaling. Sci. Rep. 2019, 9, 16781. [Google Scholar] [CrossRef]
- Zhang, C.; Peng, Z.; Ban, D.; Zhang, Y. Upregulation of Interleukin 35 in Patients With Endometriosis Stimulates Cell Proliferation. Reprod. Sci. 2018, 25, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Smycz-Kubanska, M.; Wendlocha, D.; Witek, A.; Mielczarek-Palacz, A. The role of selected cytokines from the interleukin-1 family in the peritoneal fluid of women with endometriosis. Ginekol. Pol. 2025, 96, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Xavier, P.; Belo, L.; Beires, J.; Rebelo, I.; Martinez-de-Oliveira, J.; Lunet, N.; Barros, H. Serum levels of VEGF and TNF-alpha and their association with C-reactive protein in patients with endometriosis. Arch. Gynecol. Obst. 2006, 273, 227–231. [Google Scholar] [CrossRef]
- Iwabe, T.; Harada, T.; Terakawa, N. Role of cytokines in endometriosis-associated infertility. Gynecol. Obst. Investig. 2002, 53 (Suppl. S1), 19–25. [Google Scholar] [CrossRef]
- Ullah, A.; Wang, M.J.; Wang, Y.X.; Shen, B. CXC chemokines influence immune surveillance in immunological disorders: Polycystic ovary syndrome and endometriosis. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166704. [Google Scholar] [CrossRef]
- Shimoya, K.; Zhang, Q.; Temma-Asano, K.; Hayashi, S.; Kimura, T.; Murata, Y. Fractalkine in the peritoneal fluid of women with endometriosis. Int. J. Gynaecol. Obst. 2005, 91, 36–41. [Google Scholar] [CrossRef]
- Smycz-Kubańska, M.; Kondera-Anasz, Z.; Sikora, J.; Wendlocha, D.; Królewska-Daszczyńska, P.; Englisz, A.; Janusz, A.; Janusz, J.; Mielczarek-Palacz, A. The Role of Selected Chemokines in the Peritoneal Fluid of Women with Endometriosis—Participation in the Pathogenesis of the Disease. Processes 2021, 9, 2229. [Google Scholar] [CrossRef]
- Arici, A.; Oral, E.; Attar, E.; Tazuke, S.I.; Olive, D.L. Monocyte chemotactic protein-1 concentration in peritoneal fluid of women with endometriosis and its modulation of expression in mesothelial cells. Fertil. Steril. 1997, 67, 1065–1072. [Google Scholar] [CrossRef]
- Nirgianakis, K.; McKinnon, B.; Ma, L.; Imboden, S.; Bersinger, N.; Mueller, M.D. Peritoneal fluid biomarkers in patients with endometriosis: A cross-sectional study. Horm. Mol. Biol. Clin. Investig. 2020, 42, 113–122. [Google Scholar] [CrossRef]
- Heidari, S.; Kolahdouz-Mohammadi, R.; Khodaverdi, S.; Tajik, N.; Delbandi, A.A. Expression levels of MCP-1, HGF, and IGF-1 in endometriotic patients compared with non-endometriotic controls. BMC Women’s Health 2021, 21, 422. [Google Scholar] [CrossRef]
- Han, M.T.; Cheng, W.; Zhu, R.; Wu, H.H.; Ding, J.; Zhao, N.N.; Li, H.; Wang, F.X. The cytokine profiles in follicular fluid and reproductive outcomes in women with endometriosis. Am. J. Reprod. Immunol. 2023, 89, e13633. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mu, L. Association between macrophage migration inhibitory factor in the endometrium and estrogen in endometriosis. Exp. Ther. Med. 2015, 10, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Elbaradie, S.M.Y.; Bakry, M.S.; Bosilah, A.H. Serum macrophage migration inhibition factor for diagnosing endometriosis and its severity: Case-control study. BMC Women’s Health 2020, 20, 189. [Google Scholar] [CrossRef]
- Fukaya, T.; Sugawara, J.; Yoshida, H.; Yajima, A. The role of macrophage colony stimulating factor in the peritoneal fluid in infertile patients with endometriosis. Tohoku J. Exp. Med. 1994, 172, 221–226. [Google Scholar] [CrossRef]
- Budrys, N.M.; Nair, H.B.; Liu, Y.G.; Kirma, N.B.; Binkley, P.A.; Kumar, S.; Schenken, R.S.; Tekmal, R.R. Increased expression of macrophage colony-stimulating factor and its receptor in patients with endometriosis. Fertil. Steril. 2012, 97, 1129–1135.e1. [Google Scholar] [CrossRef]
- Shi, J.; Tan, X.; Feng, G.; Zhuo, Y.; Jiang, Z.; Banda, S.; Wang, L.; Zheng, W.; Chen, L.; Yu, D.; et al. Research advances in drug therapy of endometriosis. Front. Pharmacol. 2023, 14, 1199010. [Google Scholar] [CrossRef]
- Nisolle, M.; Casanas-Roux, F.; Anaf, V.; Mine, J.M.; Donnez, J. Morphometric study of the stromal vascularisation in peritoneal endometriosis. Fertil. Steril. 1993, 59, 681–684. [Google Scholar] [CrossRef]
- McLaren, J.; Prentice, A.; Charnock-Jones, D.S.; Millican, S.A.; Müller, K.H.; Sharkey, A.M.; Smith, S.K. Vascular endothelial growth factor is produced by peritoneal fluid macrophages in endometriosis and is regulated by ovarian steroids. J. Clin. Investig. 1996, 98, 482–489. [Google Scholar] [CrossRef]
- Lee, S.R.; Kim, S.H.; Lee, Y.J.; Hong, S.H.; Chae, H.D.; Kim, C.H.; Kang, B.M.; Choi, Y.M. Expression of epidermal growth factor, fibroblast growth factor-2, and platelet-derived growth factor-A in the eutopic endometrium of women with endometriosis. J. Obstet. Gynaecol. Res. 2007, 33, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Szaflik, T.; Romanowicz, H.; Bryś, M.; Forma, E.; Szyłło, K. Analysis of VEGF, IGF1/2 and the Long Noncoding RNA (lncRNA) H19 Expression in Polish Women with Endometriosis. Int. J. Mol. Sci. 2024, 25, 5271. [Google Scholar] [CrossRef]
- Pantelis, A.; Machairiotis, N.; Lapatsanis, D.P. The Formidable yet Unresolved Interplay between Endometriosis and Obesity. Sci. World J. 2021, 2021, 6653677. [Google Scholar] [CrossRef] [PubMed]
- Oosterlynck, D.J.; Meuleman, C.; Waer, M.; Koninckx, P.R. Transforming growth factor-beta activity is increased in peritoneal fluid from women with endometriosis. Obstet. Gynecol. 1994, 83, 287–292. [Google Scholar] [PubMed]
- Abdoli, M.; Hoseini, S.M.; Sandoghsaz, R.S.; Javaheri, A.; Montazeri, F.; Moshtaghioun, S.M. Endometriotic lesions and their recurrence: A Study on the mediators of immunoregulatory (TGF-β/miR-20a) and stemness (NANOG/miR-145). J. Reprod. Immunol. 2024, 166, 104336. [Google Scholar] [CrossRef]
- Suen, J.L.; Chang, Y.; Chiu, P.R.; Hsieh, T.H.; His, E.; Chen, Y.C.; Chen, Y.F.; Tsai, E.M. Serum level of IL-10 is increased in patients with endometriosis, and IL-10 promotes the growth of lesions in a murine model. Am. J. Pathol. 2014, 184, 464–471. [Google Scholar] [CrossRef]
- Santulli, P.; Borghese, B.; Chouzenoux, S.; Streuli, I.; Borderie, D.; de Ziegler, D.; Weill, B.; Chapron, C.; Batteux, F. Interleukin-19 and interleukin-22 serum levels are decreased in patients with ovarian endometrioma. Fertil. Steril. 2013, 99, 219–226.e2. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, Y.; Liu, L.B.; Chang, K.K.; Li, H.; Li, M.Q.; Shao, J. IL-22 in the endometriotic milieu promotes the proliferation of endometrial stromal cells via stimulating the secretion of CCL2 and IL-8. Int. J. Clin. Exp. Pathol. 2013, 6, 2011–2020. [Google Scholar]
- Liu, S.; Zhao, R.; Zang, Y.; Huang, P.; Zhang, Q.; Fan, X.; Bai, J.; Zheng, X.; Zhao, S.; Kuai, D.; et al. Interleukin-22 promotes endometrial carcinoma cell proliferation and cycle progression via ERK1/2 and p38 activation. Mol. Cell. Biochem. 2025, 480, 3147–3160. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, L.; Shao, J.; Wang, Y.; Jin, L.P.; Li, D.J.; Li, M.Q. IL-22 enhances the invasiveness of endometrial stromal cells of adenomyosis in an autocrine manner. Int. J. Clin. Exp. Pathol. 2014, 7, 5762–5771. [Google Scholar]
- Jiang, J.; Jiang, Z.; Xue, M. Serum and peritoneal fluid levels of interleukin-6 and interleukin-37 as biomarkers for endometriosis. Gynecol. Endocrinol. 2019, 35, 571–575. [Google Scholar] [CrossRef]
- Hsu, C.C.; Yang, B.C.; Wu, M.H.; Huang, K.E. Enhanced interleukin-4 expression in patients with endometriosis. Fertil. Steril. 1997, 67, 1059–1064. [Google Scholar] [CrossRef]
- OuYang, Z.; Hirota, Y.; Osuga, Y.; Hamasaki, K.; Hasegawa, A.; Tajima, T.; Hirata, T.; Koga, K.; Yoshino, O.; Harada, M.; et al. Interleukin-4 stimulates proliferation of endometriotic stromal cells. Am. J. Pathol. 2008, 173, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Krygere, L.; Jukna, P.; Jariene, K.; Drejeriene, E. Diagnostic Potential of Cytokine Biomarkers in En dometriosis: Challenges and Insights. Biomedicines 2024, 12, 2867. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, H.; Jin, D.; Zhang, Y. Serum miR-17, IL-4, and IL-6 levels for diagnosis of endometriosis. Medicine 2018, 97, e10853. [Google Scholar] [CrossRef]
- Sadat Sandoghsaz, R.; Montazeri, F.; Shafienia, H.; Mehdi Kalantar, S.; Javaheri, A.; Samadi, M. Expression of miR-21 &IL-4 in endometriosis. Hum. Immunol. 2024, 85, 110746. [Google Scholar] [CrossRef]
- Wang, X.M.; Ma, Z.Y.; Song, N. Inflammatory cytokines IL-6, IL-10, IL-13, TNF-α and peritoneal fluid flora were associated with infertility in patients with endometriosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2513–2518. [Google Scholar] [CrossRef]
- Qiu, X.M.; Lai, Z.Z.; Ha, S.Y.; Yang, H.L.; Liu, L.B.; Wang, Y.; Shi, J.W.; Ruan, L.Y.; Ye, J.F.; Wu, J.N.; et al. IL-2 and IL-27 synergistically promote growth and invasion of endometriotic stromal cells by maintaining the balance of IFN-γ and IL-10 in endometriosis. Reproduction 2020, 159, 251–260. [Google Scholar] [CrossRef]
- Miller, J.E.; Monsanto, S.P.; Ahn, S.H.; Khalaj, K.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Interleukin-33 modulates inflammation in endometriosis. Sci. Rep. 2017, 7, 17903. [Google Scholar] [CrossRef]
- Miller, J.E.; Lingegowda, H.; Symons, L.K.; Bougie, O.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. IL-33 activates group 2 innate lymphoid cell expansion and modulates endometriosis. JCI Insight 2021, 6, e149699. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Tian, Q.; Li, S.; Zhou, X.; Sun, Q.; Wang, Y.; Xiao, Y.; Li, M.; Chang, K.; Yi, X. The IL-33-ST2 axis plays a vital role in endometriosis via promoting epithelial-mesenchymal transition by phosphorylating β-catenin. Cell Commun. Signal. 2024, 22, 318. [Google Scholar] [CrossRef]
- Xiao, F.; Liu, X.; Guo, S.W. Interleukin-33 Derived from Endometriotic Lesions Promotes Fibrogenesis through Inducing the Production of Profibrotic Cytokines by Regulatory T Cells. Biomedicines 2022, 10, 2893. [Google Scholar] [CrossRef]
- Sharpe-Timms, K.L.; Bruno, P.L.; Penney, L.L.; Bickel, J.T. Immunohistochemical localization of granulocyte-macrophage colony-stimulating factor in matched endometriosis and endometrial tissues. Am. J. Obstet. Gynecol. 1994, 171, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Propst, A.M.; Quade, B.J.; Nowak, R.A.; Stewart, E.A. Granulocyte macrophage colony-stimulating factor in adenomyosis and autologous endometrium. J. Soc. Gynecol. Investig. 2002, 9, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Toullec, L.; Batteux, F.; Santulli, P.; Chouzenoux, S.; Jeljeli, M.; Belmondo, T.; Hue, S.; Chapron, C. High Levels of Anti-GM-CSF Antibodies in Deep Infiltrating Endometriosis. Reprod. Sci. 2020, 27, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Q.; Ren, Y.F.; Li, B.B.; Wei, C.; Yu, B. The mysterious association between adiponectin and endometriosis. Front. Pharmacol. 2024, 15, 1396616. [Google Scholar] [CrossRef]
- Matarese, G.; Alviggi, C.; Sanna, V.; Howard, J.K.; Lord, G.M.; Carravetta, C.; Fontana, S.; Lechler, R.I.; Bloom, S.R.; De Placido, G. Increased leptin levels in serum and peritoneal fluid of patients with pelvic endometriosis. J. Clin. Endocrinol. Metab. 2000, 85, 2483–2487. [Google Scholar] [CrossRef]
- Hong, J.; Yi, K.W. What is the link between endometriosis and adiposity? Obstet. Gynecol. Sci. 2022, 65, 227–233. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, Y.; Zhao, Y.; Chang, X.H.; Zhu, H.L. Serum and peritoneal fluid leptin levels in endometriosis: A systematic review and meta-analysis. Gynecol. Endocrinol. 2021, 37, 689–693. [Google Scholar] [CrossRef]
- Wójtowicz, M.; Zdun, D.; Owczarek, A.J.; Skrzypulec-Plinta, V.; Olszanecka-Glinianowicz, M. Evaluation of adipokines concentrations in plasma, peritoneal, and endometrioma fluids in women operated on for ovarian endometriosis. Front. Endocrinol. 2023, 14, 1218980. [Google Scholar] [CrossRef]
- Kim, T.H.; Bae, N.; Kim, T.; Hsu, A.L.; Hunter, M.I.; Shin, J.H.; Jeong, J.W. Leptin Stimulates Endometriosis Development in Mouse Models. Biomedicines 2022, 10, 2160. [Google Scholar] [CrossRef]
- Kalaitzopoulos, D.R.; Lempesis, I.G.; Samartzis, N.; Kolovos, G.; Dedes, I.; Daniilidis, A.; Nirgianakis, K.; Leeners, B.; Goulis, D.G.; Samartzis, E.P. Leptin concentrations in endometriosis: A systematic review and meta-analysis. J. Reprod. Immunol. 2021, 146, 103338. [Google Scholar] [CrossRef]
- Wu, M.H.; Chen, K.F.; Lin, S.C.; Lgu, C.W.; Tsai, S.J. Aberrant expression of leptin in human endometriotic stromal cells is induced by elevated levels of hypoxia inducible factor-1alpha. Am. J. Pathol. 2007, 170, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Zyguła, A.; Sankiewicz, A.; Sakowicz, A.; Dobrzyńska, E.; Dakowicz, A.; Mańka, G.; Kiecka, M.; Spaczynski, R.; Piekarski, P.; Banaszewska, B.; et al. Is the leptin/BMI ratio a reliable biomarker for endometriosis? Front. Endocrinol. 2024, 15, 1359182. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Osuga, Y.; Harada, M.; Hirata, T.; Koga, K.; Morimoto, C.; Hirota, Y.; Yoshino, O.; Yano, T.; Taketani, Y. Serum adiponectin concentrations are decreased in women with endometriosis. Hum. Reprod. 2005, 20, 3510–3513. [Google Scholar] [CrossRef]
- Zhao, Z.; Wu, Y.; Zhang, H.; Wang, X.; Tian, X.; Wang, Y.; Qiu, Z.; Zou, L.; Tang, Z.; Huang, M. Association of leptin and adiponectin levels with endometriosis: A systematic review and meta-analysis. Gynecol. Endocrinol. 2021, 37, 591–599. [Google Scholar] [CrossRef]
- Yi, K.W.; Shin, J.H.; Park, H.T.; Kim, T.; Kim, S.H.; Hur, J.Y. Resistin concentration is increased in the peritoneal fluid of women with endometriosis. Am. J. Reprod. Immunol. 2010, 64, 318–323. [Google Scholar] [CrossRef]
- Oh, Y.K.; Ha, Y.R.; Yi, K.W.; Park, H.T.; Shin, J.H.; Kim, T.; Hur, J.Y. Increased expression of resistin in ectopic endometrial tissue of women with endometriosis. Am. J. Reprod. Immunol. 2017, 78, e12726. [Google Scholar] [CrossRef]
- Lee, J.C.; Kim, S.H.; Oh, Y.S.; Kim, J.H.; Lee, S.R.; Chae, H.D. Increased Expression of Retinol-Binding Protein 4 in Ovarian Endometrioma and Its Possible Role in the Pathogenesis of Endometriosis. Int. J. Mol. Sci. 2021, 22, 5827. [Google Scholar] [CrossRef]
- Lv, S.J.; Sun, J.N.; Gan, L.; Sun, J. Identification of molecular subtypes and immune infiltration in endometriosis: A novel bioinformatics analysis and In vitro validation. Front. Immunol. 2023, 14, 1130738. [Google Scholar] [CrossRef]
- Krasnyi, A.M.; Sadekova, A.A.; Smolnova, T.Y.; Chursin, V.V.; Buralkina, N.A.; Chuprynin, V.D.; Yarotskaya, E.; Pavlovich, S.V.; Sukhikh, G.T. The Levels of Ghrelin, Glucagon, Visfatin and Glp-1 Are Decreased in the Peritoneal Fluid of Women with Endometriosis along with the Increased Expression of the CD10 Protease by the Macrophages. Int. J. Mol. Sci. 2022, 23, 10361. [Google Scholar] [CrossRef]
- Morotti, M.; Vincent, K.; Becker, C.M. Mechanisms of pain in endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 209, 8–13. [Google Scholar] [CrossRef]
- Zheng, P.; Zhang, W.; Leng, J.; Lang, J. Research on central sensitisation of endometriosis-associated pain: A systematic review of the literature. J. Pain Res. 2019, 12, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Monnin, N.; Fattet, A.J.; Koscinski, I. Endometriosis: Update of Pathophysiology, (Epi) Genetic and Environmental Involvement. Biomedicines 2023, 11, 978. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Lv, W. A laparoscopic surgery for deep infiltrating endometriosis and the review of literature. Clin. Exp. Obstet. Gynecol. 2016, 43, 616–618. [Google Scholar] [CrossRef]
- Gentles, A.; Goodwin, E.; Bedaiwy, Y.; Marshall, N.; Yong, P.J. Nociplastic Pain in Endometriosis: A Scoping Review. J. Clin. Med. 2024, 13, 7521. [Google Scholar] [CrossRef]
- Godin, S.K.; Wagner, J.; Huang, P.; Bree, D. The role of peripheral nerve signaling in endometriosis. FASEB BioAdvances 2021, 3, 802–813. [Google Scholar] [CrossRef]
- Astruc, A.; Roux, L.; Robin, F.; Sall, N.R.; Dion, L.; Lavoué, V.; Legendre, G.; Leveque, J.; Bessede, T.; Bertrand, M.; et al. Advanced Insights into Human Uterine Innervation: Implications for Endometriosis and Pelvic Pain. J. Clin. Med. 2024, 13, 1433. [Google Scholar] [CrossRef]
- Fattori, V.; Zaninelli, T.H.; Rasquel-Oliveira, F.S.; Heintz, O.K.; Jain, A.; Sun, L.; Seshan, M.L.; Peterse, D.; Lindholm, A.E.; Anchan, R.M.; et al. Nociceptor-to-macrophage communication through CGRP/RAMP1 signaling drives endometriosis-associated pain and lesion growth in mice. Sci. Transl. Med. 2024, 16, eadk8230. [Google Scholar] [CrossRef]
- Lingegowda, H.; Williams, B.J.; Spiess, K.G.; Sisnett, D.J.; Lomax, A.E.; Koti, M.; Tayadeet, C. Role of the endocannabinoid system in the pathophysiology of endometriosis and therapeutic implications. J. Cannabis Res. 2022, 4, 54. [Google Scholar] [CrossRef]
- Clayton, P.; Subah, S.; Venkatesh, R.; Hill, M.; Bogoda, N. Palmitoylethanolamide: A Potential Alternative to Cannabidiol. J. Diet. Supp. 2023, 20, 505–530. [Google Scholar] [CrossRef]
- Farooqi, T.; Bhuyan, D.J.; Low, M.; Sinclair, J.; Leonardi, M.; Armour, M. Cannabis and Endometriosis: The Roles of the Gut Microbiota and the Endocannabinoid System. J. Clin. Med. 2023, 12, 7071. [Google Scholar] [CrossRef]
- Vercellini, P.; Fedele, L.; Aimi, G.; Pietropaolo, G.; Consonni, D.; Crosignani, P.G. Association between endometriosis stage, lesion type, patient characteristics and severity of pelvic pain symptoms: A multivariate analysis of over 1000 patients. Hum. Reprod. 2007, 22, 266–271. [Google Scholar] [CrossRef] [PubMed]
- As-Sanie, S.; Kim, J.; Schmidt-Wilcke, T.; Sundgren, P.C.; Clauw, D.J.; Napadow, V.; Harris, R.E. Functional Connectivity is Associated With Altered Brain Chemistry in Women with Endometriosis-Associated Chronic Pelvic Pain. J. Pain 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Eippert, F.; Tracey, I.; Chen, A.; De, E.; Argoff, C. Pain and the PAG: Learning from painful mistakes. Nat. Neurosci. 2014, 17, 1438–1439. [Google Scholar] [CrossRef]
- Tran, L.V.; Tokushige, N.; Berbic, M.; Markham, R.; Fraser, I.S. Macrophages and nerve fibres in peritoneal endometriosis. Hum. Reprod. 2009, 24, 835–841. [Google Scholar] [CrossRef]
- Wei, Y.; Liang, Y.; Lin, H.; Dai, Y.; Yao, S. Autonomic nervous system and inflammation interaction in endometriosis-associated pain. J. Neuroinflamm. 2020, 17, 80. [Google Scholar] [CrossRef]
- Wu, J.; Xie, H.; Yao, S.; Liang, Y. Macrophage and nerve interaction in endometriosis. J. Neuroinflamm. 2017, 14, 53. [Google Scholar] [CrossRef]
- Castro, J.; Maddern, J.; Erickson, A.; Harrington, A.M.; Brierley, S.M. Peripheral and central neuroplasticity in a mouse model of endometriosis. J. Neurochem. 2024, 168, 3777–3800. [Google Scholar] [CrossRef]
- Machairiotis, N.; Vasilakaki, S.; Thomakos, N. Inflammatory Mediators and Pain in Endometriosis: A Systematic Review. Biomedicines 2021, 9, 54. [Google Scholar] [CrossRef]
- Maddern, J.; Grundy, L.; Castro, J.; Brierley, S.M. Pain in Endometriosis. Front. Cell. Neurosci. 2020, 14, 590823. [Google Scholar] [CrossRef]
- Greaves, E.; Temp, J.; Esnal-Zufiurre, A.; Mechsner, S.; Horne, A.W.; Saunders, P.T. Estradiol is a critical mediator of macrophage-nerve cross talk in peritoneal endometriosis. Am. J. Pathol. 2015, 185, 2286–2297. [Google Scholar] [CrossRef]
- Falcone, T.; Flyckt, R. Clinical Management of Endometriosis. Obstet. Gynecol. 2018, 131, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Bajaj, P.; Madsen, H.; Arendt-Nielsen, L. Endometriosis is associated with central sensitization: A psychophysical controlled study. J. Pain 2003, 4, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Guo, X.; Zhu, L.; Wang, J.; Li, T.; Yu, Q.; Zhang, X. Macrophage-derived netrin-1 contributes to endometriosis-associated pain. Ann. Transl. Med. 2021, 9, 29. [Google Scholar] [CrossRef]
- Forster, R.; Sarginson, A.; Velichkova, A.; Hogg, C.; Dorning, A.; Horne, A.W.; Saunders, P.T.K.; Greaves, E. Macrophage-derived insulin-like growth factor-1 is a key neurotrophic and nerve-sensitizing factor in pain associated with endometriosis. FASEB J. 2019, 33, 11210–11222. [Google Scholar] [CrossRef]
- Hosseinirad, H.; Rahman, M.S.; Jeong, J.W. Targeting TET3 in macrophages provides a concept strategy for the treatment of endometriosis. J. Clin. Investig. 2024, 134, e185421. [Google Scholar] [CrossRef]
- Li, J.; Wu, Z.; Li, N.; Wang, J.; Huang, M.; Zhu, L.; Wan, G.; Zhang, Z. Exploring macrophage and nerve interaction in endometriosis-associated pain: The inductive role of IL-33. Inflamm. Res. 2025, 74, 42. [Google Scholar] [CrossRef]
- Midavaine, É.; Moraes, B.C.; Benitez, J.; Rodriguez, S.R.; Braz, J.M.; Kochhar, N.P.; Eckalbar, W.L.; Tian, L.; Domingos, A.I.; Pintar, J.E.; et al. Meningeal regulatory T cells inhibit nociception in female mice. Science 2025, 388, 96–104. [Google Scholar] [CrossRef]
- Nielsen, N.M.; Jørgensen, K.T.; Pedersen, B.V.; Rostgaard, K.; Frisch, M. The co-occurrence of endometriosis with multiple sclerosis, systemic lupus erythematosus and Sjogren syndrome. Hum. Reprod. 2011, 26, 1555–1559. [Google Scholar] [CrossRef]
- Matalliotaki, C.; Matalliotakis, M.; Zervou, M.I.; Trivli, A.; Matalliotakis, I.; Mavromatidis, G.; Spandidos, D.A.; Albertsen, H.M.; Chettier, R.; Ward, K.; et al. Co-existence of endometriosis with 13 non-gynecological co-morbidities: Mutation analysis by whole exome sequencing. Mol. Med. Rep. 2018, 18, 5053–5057. [Google Scholar] [CrossRef]
- Hamouda, R.K.; Arzoun, H.; Sahib, I.; Escudero Mendez, L.; Srinivasan, M.; Shoukrie, S.I.; Dhanoa, R.K.; Selvaraj, R.; Malla, J.; Selvamani, T.Y.; et al. The Comorbidity of Endometriosis and Systemic Lupus Erythematosus: A Systematic Review. Cureus 2023, 15, e42362. [Google Scholar] [CrossRef]
- Días, J.A., Jr.; de Oliveira, R.M.; Abrao, M.S. Antinuclear antibodies and endometriosis. Int. J. Gynaecol. Obstet. 2006, 93, 262–263. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, S.; Uldall Torp, N.M.; Andersen, S.; Andersen, S.L. Endometriosis, polycystic ovary syndrome, and the thyroid: A review. Endocr. Connect. 2024, 13, e230431. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.P.; Holt, V.L.; Lee, J.H.; Jiang, H.; Rust, P.F. Levels of antibodies to transferrin and alpha 2-HS glycoprotein in women with and without endometriosis. Am. J. Reprod. Immunol. 1998, 40, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Dotan, A.; Kanduc, D.; Muller, S.; Makatsariya, A.; Shoenfeld, Y. Molecular mimicry between SARS-CoV-2 and the female reproductive system. Am. J. Reprod. Immunol. 2021, 86, e13494. [Google Scholar] [CrossRef]
- Garvey, M. Endometriosis: Future Biological Perspectives for Diagnosis and Treatment. Int. J. Mol. Sci. 2024, 25, 12242. [Google Scholar] [CrossRef]
- Zhang, W.; Li, K.; Jian, A.; Zhang, G.; Zhang, X. Prospects for potential therapy targeting immune associated factors in endometriosis (Review). Mol. Med. Rep. 2025, 31, 57. [Google Scholar] [CrossRef]
- Kato, T.; Yasuda, K.; Matsushita, K.; Ishii, K.J.; Hirota, S.; Yoshimoto, T.; Shibahara, H. Interleukin-1/-33 Signaling Pathways as Therapeutic Targets for Endometriosis. Front. Immunol. 2019, 10, 2021. [Google Scholar] [CrossRef]
- Fedotcheva, T.A.; Fedotcheva, N.I.; Shimanovsky, N.L. Progesterone as an Anti-Inflammatory Drug and Immunomodulator: New Aspects in Hormonal Regulation of the Inflammation. Biomolecules 2022, 12, 1299. [Google Scholar] [CrossRef]
- Chang, C.Y.; Chiang, A.J.; Yan, M.J.; Lai, M.T.; Su, Y.Y.; Huang, H.Y.; Chang, C.Y.; Li, Y.H.; Li, P.F.; Chen, C.M.; et al. Ribosome biogenesis serves as a therapeutic target for treating endometriosis and the associated complications. Biomedicines 2022, 10, 185. [Google Scholar] [CrossRef]
- Hamid, A.M.; Madkour, W.A.; Moawad, A.; Elzaher, M.A.; Roberts, M.P. Does cabergoline help in decreasing endometrioma size compared to LHRH agonist? A prospective randomized study. Arch. Gynecol. Obstet. 2014, 290, 677–682. [Google Scholar] [CrossRef]
- DiVasta, A.D.; Stamoulis, C.; Gallagher, J.S.; Laufer, M.R.; Anchan, R.; Hornstein, M.D. Nonhormonal therapy for endometriosis: A randomized, placebo-controlled, pilot study of cabergoline versus norethindrone acetate. F&S Rep. 2021, 2, 454–461. [Google Scholar] [CrossRef]
- Wyatt, J.; Fernando, S.M.; Powell, S.G.; Hill, C.J.; Arshad, I.; Probert, C.; Ahmed, S.; Hapangama, D.K. The role of iron in the pathogenesis of endometriosis: A systematic review. Hum. Reprod. Open 2023, 2023, hoad033. [Google Scholar] [CrossRef] [PubMed]
- Tewary, S.; Lucas, E.S.; Fujihara, R.; Kimani, P.K.; Polanco, A.; Brighton, P.J.; Muter, J.; Fishwick, K.J.; Da Costa, M.J.M.D.; Ewington, L.J.; et al. Impact of sitagliptin on endometrial mesenchymal stem-like progenitor cells: A randomised, double-blind placebo-controlled feasibility trial. eBioMedicine 2020, 51, 102597. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lv, X.; Jiang, M.; Jin, Z. Sitagliptin ameliorates hypoxia-induced damages in endometrial stromal cells: An implication in endometriosis. Bioengineered 2022, 13, 800–809. [Google Scholar] [CrossRef]
- Grzymajlo, K.; El Hafny-Rahbi, B.; Kieda, C. Tumour suppressor PTEN activity is differentially inducible by myo-inositol phosphates. J. Cell. Mol. Med. 2023, 27, 879–890. [Google Scholar] [CrossRef]
- Dera-Szymanowska, A.; Chmaj-Wierzchowska, K.; Horst, N.; Stryjakowska, K.; Wirlstein, P.; Andrusiewicz, M.; Florek, E.; Beltrano, J.; Szymanowski, K.; Wolun-Cholewa, M. Immunomodulation inhibits the development of endometriosis in rats. J. Physiol. Pharmacol. 2020, 71, 145–153. [Google Scholar] [CrossRef]
- Chu, X.; Hou, M.; Li, Y.; Zhang, Q.; Wang, S.; Ma, J. Extracellular vesicles in endometriosis: Role and potential. Front. Endocrinol. 2024, 15, 1365327. [Google Scholar] [CrossRef]
- Meczekalski, B.; Nowicka, A.; Bochynska, S.; Szczesnowicz, A.; Bala, G.; Szeliga, A. Kisspeptin and Endometriosis-Is There a Link? J. Clin. Med. 2024, 13, 7683. [Google Scholar] [CrossRef]
- Chen, Y.; Li, T. Unveiling the Mechanisms of Pain in Endometriosis: Comprehensive Analysis of Inflammatory Sensitization and Therapeutic Potential. Int. J. Mol. Sci. 2025, 26, 1770. [Google Scholar] [CrossRef]
- Genovese, T.; Siracusa, R.; D’Amico, R.; Cordaro, M.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Trovato Salinaro, A.; Raffone, E.; Impellizzeri, D.; et al. Regulation of Inflammatory and Proliferative Pathways by Fotemustine and Dexamethasone in Endometriosis. Int. J. Mol. Sci. 2021, 22, 5998. [Google Scholar] [CrossRef]
- Maksym, R.B.; Hoffmann-Młodzianowska, M.; Skibińska, M.; Rabijewski, M.; Mackiewicz, A.; Kieda, C. Immunology and Immunotherapy of Endometriosis. J. Clin. Med. 2021, 10, 5879. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Kim, S.K.; Kim, S.R.; Kim, D.; Kim, K.W.; Hong, I.S.; Lee, H.Y. Noncanonical functions of glucocorticoids: A novel role for glucocorticoids in performing multiple beneficial functions in endometrial stem cells. Cell Death Dis. 2021, 12, 612. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.F.; Li, H.; Zeng, L.; Yang, H.; Hu, Y.; Qu, Y.; Chen, W.; Sun, Y.; Zhang, W.; Zeng, X.; et al. Use of statins and risks of ovarian, uterine, and cervical diseases: A cohort study in the UK Biobank. Eur. J. Clin. Pharmacol. 2024, 80, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Wang, Q.; Xu, D.; Sun, Y.; Xu, W.; Wang, B.; Yang, Z.; Hao, L. Atorvastatin exerts dual effects of lesion regression and ovarian protection in the prevention and treatment of endometriosis. Eur. J. Pharmacol. 2024, 964, 176261. [Google Scholar] [CrossRef]
- Dillon, G.A.; Stanhewicz, A.E.; Serviente, C.; Flores, V.A.; Stachenfeld, N.; Alexander, L.M. Seven days of statin treatment improves nitric-oxide mediated endothelial-dependent cutaneous microvascular function in women with endometriosis. Microvasc. Res. 2022, 144, 104421. [Google Scholar] [CrossRef]
- Perelló, M.; González-Foruria, I.; Castillo, P.; Martínez-Florensa, M.; Lozano, F.; Balasch, J.; Carmona, F. Oral Administration of Pentoxifylline Reduces Endometriosis-Like Lesions in a Nude Mouse Model. Reprod. Sci. 2017, 24, 911–918. [Google Scholar] [CrossRef]
- Creus, M.; Fábregues, F.; Carmona, F.; del Pino, M.; Manau, D.; Balasch, J. Combined laparoscopic surgery and pentoxifylline therapy for treatment of endometriosis-associated infertility: A preliminary trial. Hum. Reprod. 2008, 23, 1910–1916. [Google Scholar] [CrossRef]
- Kamencic, H.; Thiel, J.A. Pentoxifylline after conservative surgery for endometriosis: A randomized, controlled trial. J. Minim. Invasive Gynecol. 2008, 15, 62–66. [Google Scholar] [CrossRef]
- Alborzi, S.; Ghotbi, S.; Parsanezhad, M.E.; Dehbashi, S.; Alborzi, S.; Alborzi, M. Pentoxifylline therapy after laparoscopic surgery for different stages of endometriosis: A prospective, double-blind, randomized, placebo-controlled study. J. Minim. Invasive Gynecol. 2007, 14, 54–58. [Google Scholar] [CrossRef]
- Grammatis, A.L.; Georgiou, E.X.; Becker, C.M. Pentoxifylline for the treatment of endometriosis-associated pain and infertility. Cochrane Database Syst. Rev. 2021, 8, CD007677. [Google Scholar] [CrossRef]
- Clemenza, S.; Sorbi, F.; Noci, I.; Capezzuoli, T.; Turrini, I.; Carriero, C.; Buffi, N.; Fambrini, M.; Petraglia, F. From pathogenesis to clinical practice: Emerging medical treatments for endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 51, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Lebovic, D.I.; Kir, M.; Casey, C.L. Peroxisome proliferator-activated receptor-gamma induces regression of endometrial explants in a rat model of endometriosis. Fertil. Steril. 2004, 82 (Suppl. S3), 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Lebovic, D.I.; Mwenda, J.M.; Chai, D.C.; Mueller, M.D.; Santi, A.; Fisseha, S.; D’Hooghe, T. PPAR-gamma receptor ligand induces regression of endometrial explants in baboons: A prospective, randomized, placebo- and drug-controlled study. Fertil. Steril. 2007, 88 (Suppl. S4), 1108–1119. [Google Scholar] [CrossRef]
- Kim, C.H.; Lee, Y.J.; Kim, J.B.; Lee, K.H.; Kwon, S.K.; Ahn, J.W.; Kim, S.H.; Chae, H.D.; Kang, B.M. Effect of Pioglitazone on Production of Regulated upon Activation Normal T-cell Expressed and Secreted (RANTES) and IVF Outcomes in Infertile Women with Endometriosis. Dev. Reprod. 2013, 17, 207–213. [Google Scholar] [CrossRef]
- Ren, X.U.; Wang, Y.; Xu, G.; Dai, L. Effect of rapamycin on endometriosis in mice. Exp. Therap. Med. 2016, 12, 101–106. [Google Scholar] [CrossRef]
- Fan, J.; Chen, C.; Zhong, Y. A cohort study on IVF outcomes in infertile endometriosis patients: The effects of rapamycin treatment. Reprod. Biomed. Online 2024, 48, 103319. [Google Scholar] [CrossRef]
- Palmer, S.S.; Altan, M.; Denis, D.; Tos, E.G.; Gotteland, J.P.; Osteen, K.G.; Bruner-Tran, K.L.; Nataraja, S.G. Bentamapimod (JNK Inhibitor AS602801) Induces Regression of Endometriotic Lesions in Animal Models. Reprod. Sci. 2016, 23, 11–23. [Google Scholar] [CrossRef]
- Hussein, M.; Chai, D.C.; Kyama, C.M.; Mwenda, J.M.; Palmer, S.S.; Gotteland, J.P.; D’Hooghe, T.M. c-Jun NH2-terminal kinase inhibitor bentamapimod reduces induced endometriosis in baboons: An assessor-blind placebo-controlled randomized study. Fertil. Steril. 2016, 105, 815–824.e5. [Google Scholar] [CrossRef]
- Feng, Y.; Dong, H.; Zheng, L. Ligustrazine inhibits inflammatory response of human endometrial stromal cells through the STAT3/IGF2BP1/RELA axis. Pharm. Biol. 2023, 61, 666–673. [Google Scholar] [CrossRef]
- Jiang, T.; Chen, Y.; Gu, X.; Miao, M.; Hu, D.; Zhou, H.; Chen, J.; Teichmann, A.T.; Yang, Y. Review of the Potential Therapeutic Effects and Molecular Mechanisms of Resveratrol on Endometriosis. Int. J. Women’s Health 2023, 15, 741–763. [Google Scholar] [CrossRef]
- Bruner-Tran, K.L.; Osteen, K.G.; Taylor, H.S.; Sokalska, A.; Haines, K.; Duleba, A.J. Resveratrol inhibits development of experimental endometriosis in vivo and reduces endometrial stromal cell invasiveness in vitro. Biol. Reprod. 2011, 84, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Kodarahmian, M.; Amidi, F.; Moini, A.; Kashani, L.; Shabani Nashtaei, M.; Pazhohan, A.; Bahramrezai, M.; Berenjian, S.; Sobhani, A. The modulating effects of Resveratrol on the expression of MMP-2 and MMP-9 in endometriosis women: A randomized exploratory trial. Gynecol. Endocrinol. 2019, 35, 719–726. [Google Scholar] [CrossRef]
- Khodarahmian, M.; Amidi, F.; Moini, A.; Kashani, L.; Salahi, E.; Danaii-Mehrabad, S.; Nashtaei, M.S.; Mojtahedi, M.F.; Esfandyari, S.; Sobhani, A. A randomized exploratory trial to assess the effects of resveratrol on VEGF and TNF-α 2 expression in endometriosis women. J. Reprod. Immunol. 2021, 143, 103248. [Google Scholar] [CrossRef] [PubMed]
- Podgrajsek, R.; Ban Frangez, H.; Stimpfel, M. Molecular Mechanism of Resveratrol and Its Therapeutic Potential on Female Infertility. Int. J. Mol. Sci. 2024, 25, 3613. [Google Scholar] [CrossRef] [PubMed]
- Rostami, S.; Alyasin, A.; Saedi, M.; Nekoonam, S.; Khodarahmian, M.; Moeini, A.; Amidi, F. Astaxanthin ameliorates inflammation, oxidative stress, and reproductive outcomes in endometriosis patients undergoing assisted reproduction: A randomized, triple-blind placebo-controlled clinical trial. Front. Endocrinol. 2023, 14, 1144323. [Google Scholar] [CrossRef]
- Kamal, D.A.M.; Salamt, N.; Yusuf, A.N.M.; Kashim, M.I.A.M.; Mokhtar, M.H. Potential Health Benefits of Curcumin on Female Reproductive Disorders: A Review. Nutrients 2021, 13, 3126. [Google Scholar] [CrossRef]
- Chowdhury, I.; Banerjee, S.; Driss, A.; Xu, W.; Mehrabi, S.; Nezhat, C.; Sidell, N.; Taylor, R.N.; Thompson, W.E. Curcumin attenuates proangiogenic and proinflammatory factors in human eutopic endometrial stromal cells through the NF-κB signaling pathway. J. Cell. Physiol. 2019, 234, 6298–6312. [Google Scholar] [CrossRef]
- Jannatifar, R.; Asa, E.; Cheraghi, E.; Verdi, A. Nanomicelle curcumin improves oxidative stress, inflammatory markers, and assisted reproductive techniques outcomes in endometriosis cases: A randomized clinical trial. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2025. [Google Scholar] [CrossRef]
- Hipólito-Reis, M.; Neto, A.C.; Neves, D. Impact of curcumin, quercetin, or resveratrol on the pathophysiology of endometriosis: A systematic review. Phytother. Res. 2022, 36, 2416–2433. [Google Scholar] [CrossRef]
- Jian, X.; Shi, C.; Luo, W.; Zhou, L.; Jiang, L.; Liu, K. Therapeutic effects and molecular mechanisms of quercetin in gynecological disorders. Biomed. Pharmacother. 2024, 173, 116418. [Google Scholar] [CrossRef]
- Park, S.; Lim, W.; Bazer, F.W.; Whang, K.Y.; Song, G. Quercetin inhibits proliferation of endometriosis regulating cyclin D1 and its target microRNAs in vitro and in vivo. J. Nutr. Biochem. 2019, 63, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Włodarczyk, M.; Ciebiera, M.; Nowicka, G.; Łoziński, T.; Ali, M.; Al-Hendy, A. Epigallocatechin Gallate for the Treatment of Benign and Malignant Gynecological Diseases-Focus on Epigenetic Mechanisms. Nutrients 2024, 16, 559. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.W.; Liang, B.; Gao, Y.; Zhang, R.; Tan, Z.; Zhang, T.; Chung, P.W.J.; Chan, T.H.; Wang, C.C. An In-Silico, In-Vitro and In-Vivo Combined Approach to Identify NMNATs as Potential Protein Targets of ProEGCG for Treatment of Endometriosis. Front. Pharmacol. 2021, 12, 714790. [Google Scholar] [CrossRef]
- Stochino Loi, E.; Pontis, A.; Cofelice, V.; Pirarba, S.; Fais, M.F.; Daniilidis, A.; Melis, I.; Paoletti, A.M.; Angioni, S. Effect of ultramicronized-palmitoylethanolamide and co-micronized palmitoylethanolamide/polydatin on chronic pelvic pain and quality of life in endometriosis patients: An open-label pilot study. Int. J. Women’s Health 2019, 11, 443–449. [Google Scholar] [CrossRef]
- Indraccolo, U.; Indraccolo, S.R.; Mignini, F. Micronized palmitoylethanolamide/trans-polydatin treatment of endometriosis-related pain: A meta-analysis. Ann. Ist. Super. Sanita 2017, 53, 125–134. [Google Scholar] [CrossRef]
- Genovese, T.; Cordaro, M.; Siracusa, R.; Impellizzeri, D.; Caudullo, S.; Raffone, E.; Macrí, F.; Interdonato, L.; Gugliandolo, E.; Interlandi, C.; et al. Molecular and Biochemical Mechanism of Cannabidiol in the Management of the Inflammatory and Oxidative Processes Associated with Endometriosis. Int. J. Mol. Sci. 2022, 23, 5427. [Google Scholar] [CrossRef]
- Sinclair, J.; Collett, L.; Abbott, J.; Pate, D.W.; Sarris, J.; Armour, M. Effects of cannabis ingestion on endometriosis-associated pelvic pain and related symptoms. PLoS ONE 2021, 16, e0258940. [Google Scholar] [CrossRef]
- Mistry, M.; Simpson, P.; Morris, E.; Fritz, A.K.; Karavadra, B.; Lennox, C.; Prosser-Snelling, E. Cannabidiol for the Management of Endometriosis and Chronic Pelvic Pain. J. Minim. Invasive Gynecol. 2022, 29, 169–176. [Google Scholar] [CrossRef]
- Dimitrov, N.V.; Meyer, C.J.; Perloff, M.; Ruppenthal, M.M.; Phillipich, M.J.; Gilliland, D.; Malone, W.; Minn, F.L. Alteration of retinol-binding-protein concentrations by the synthetic retinoid fenretinide in healthy human subjects. Am. J. Clin. Nutr. 1990, 51, 1082–1087. [Google Scholar] [CrossRef]
- Pavone, M.E.; Malpani, S.S.; Dyson, M.; Kim, J.J.; Bulun, S.E. Fenretinide: A Potential Treatment for Endometriosis. Reprod. Sci. 2016, 23, 1139–1147. [Google Scholar] [CrossRef]
- Garić, D.; Dumut, D.C.; Shah, J.; De Sanctis, J.B.; Radzioch, D. The role of essential fatty acids in cystic fibrosis and normalizing effect of fenretinide. Cell. Mol. Life Sci. 2020, 77, 4255–4267. [Google Scholar] [CrossRef] [PubMed]
- Aristarco, V.; Serrano, D.; Maisonneuve, P.; Guerrieri-Gonzaga, A.; Lazzeroni, M.; Feroce, I.; Macis, D.; Cavadini, E.; Albertazzi, E.; Jemos, C.; et al. Fenretinide in Young Women at Genetic or Familial Risk of Breast Cancer: A Placebo-Controlled Biomarker Trial. Cancer Prev. Res. 2024, 17, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinskaya, M.; Denisova, A.; Tkachenko, N.; Ivashenko, T.; Bespalova, O.; Tolibova, G.; Tral, T. Vitamin D significance in pathogenesis of endometriosis. Gynecol. Endocrinol. 2021, 37 (Suppl. S1), 40–43. [Google Scholar] [CrossRef] [PubMed]
- Kalaitzopoulos, D.R.; Samartzis, N.; Daniilidis, A.; Leeners, B.; Makieva, S.; Nirgianakis, K.; Dedes, I.; Metzler, J.M.; Imesch, P.; Lempesis, I.G. Effects of vitamin D supplementation in endometriosis: A systematic review. Reprod. Biol. Endocrinol. 2022, 20, 176. [Google Scholar] [CrossRef]
- Xie, B.; Liao, M.; Huang, Y.; Hang, F.; Ma, N.; Hu, Q.; Wang, J.; Jin, Y.; Qin, A. Association between vitamin D and endometriosis among American women: National Health and Nutrition Examination Survey. PLoS ONE 2024, 19, e0296190. [Google Scholar] [CrossRef]
- Quattrone, F.; Sanchez, A.M.; Pannese, M.; Hemmerle, T.; Viganò, P.; Candiani, M.; Petraglia, F.; Neri, D.; Panina-Bordignon, P. The Targeted Delivery of Interleukin 4 Inhibits Development of Endometriotic Lesions in a Mouse Model. Reprod. Sci. 2015, 22, 1143–1152. [Google Scholar] [CrossRef]
- Somigliana, E.; Viganò, P.; Rossi, G.; Carinelli, S.; Vignali, M.; Panina-Bordignon, P. Endometrial ability to implant in ectopic sites can be prevented by interleukin-12 in a murine model of endometriosis. Hum. Reprod. 1999, 14, 2944–2950. [Google Scholar] [CrossRef]
- Itoh, H.; Sashihara, T.; Hosono, A.; Kaminogawa, S.; Uchida, M. Interleukin-12 inhibits development of ectopic endometriotic tissues in peritoneal cavity via activation of NK cells in a murine endometriosis model. Cytotechnology 2011, 63, 133–141. [Google Scholar] [CrossRef]
- Altintas, D.; Kokcu, A.; Tosun, M.; Cetinkaya, M.B.; Kandemir, B. Efficacy of recombinant human interferon alpha-2b on experimental endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 139, 95–99. [Google Scholar] [CrossRef]
- Dicitore, A.; Castiglioni, S.; Saronni, D.; Gentilini, D.; Borghi, M.O.; Stabile, S.; Vignali, M.; Di Blasio, A.M.; Persani, L.; Vitale, G. Effects of human recombinant type I IFNs (IFN-α2b and IFN-β1a) on growth and migration of primary endometrial stromal cells from women with deeply infiltrating endometriosis: A preliminary study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 230, 192–198. [Google Scholar] [CrossRef]
- Acién, P.; Quereda, F.; Campos, A.; Gomez-Torres, M.J.; Velasco, I.; Gutierrez, M. Use of intraperitoneal interferon alpha-2b therapy after conservative surgery for endometriosis and postoperative medical treatment with depot gonadotropin-releasing hormone analog: A randomized clinical trial. Fertil. Steril. 2002, 78, 705–711. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xiong, T.; Guo, F.; Du, Z.; Fan, Y.; Sun, H.; Feng, Z.; Zhang, G. Interleukin-37b inhibits the growth of murine endometriosis-like lesions by regulating proliferation, invasion, angiogenesis and inflammation. Mol. Hum. Reprod. 2020, 26, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liao, Z.; Ye, M.; Jiang, J. Recombinant human IL-37 inhibited endometriosis development in a mouse model through increasing Th1/Th2 ratio by inducing the maturation of dendritic cells. Reprod. Biol. Endocrinol. 2021, 19, 128. [Google Scholar] [CrossRef]
- Önalan, G.; Tohma, Y.A.; Zeyneloğlu, H.B. Effect of Etanercept on the Success of Assisted Reproductive Technology in Patients with Endometrioma. Gynecol. Obstet. Investig. 2018, 83, 358–364. [Google Scholar] [CrossRef]
- Liu, M.; Li, Y.; Yuan, Y.; Jiang, M.; Yin, P.; Yang, D. Peri-implantation treatment with TNF-α inhibitor for endometriosis and/or adenomyosis women undergoing frozen-thawed embryo transfer: A retrospective cohort study. J. Reprod. Immunol. 2025, 167, 104415. [Google Scholar] [CrossRef]
- Lu, D.; Song, H.; Shi, G. Anti-TNF-α treatment for pelvic pain associated with endometriosis. Cochrane Database Syst. Rev. 2013, 2013, CD008088. [Google Scholar] [CrossRef]
- Sullender, R.T.; Agarwal, R.K.; Jacobs, M.B.; Wessels, J.M.; Foster, W.G.; Agarwal, S.K. Pilot Study of IL-1 Antagonist Anakinra for Treatment of Endometriosis. Int. J. Women’s Health 2024, 16, 1583–1593. [Google Scholar] [CrossRef]
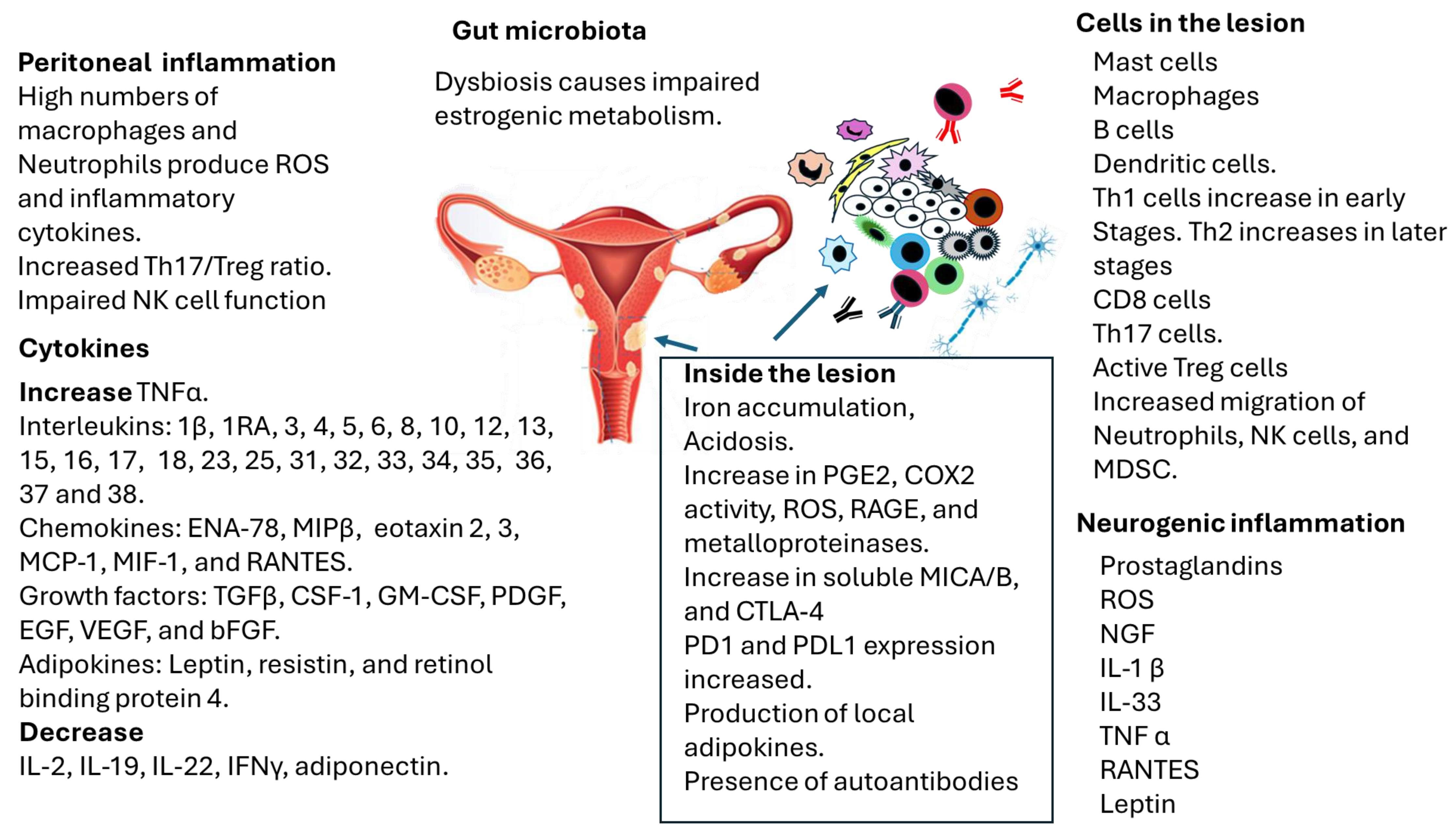
| Lesion | Clinical Characteristics | Possible Coexisting Medical Conditions |
|---|---|---|
| Pelvic Superficial peritoneal [9,13] Focal (adenomyoma) or diffuse (adenomyosis) lesions within the myometrium [9,13] Ovarian endometriosis [9,13] Deep endometriosis Lesions > 5 mm [13,15] Common in the rectovaginal septum It may involve the large intestine, bladder, ureters, and appendix [13,15] | Pelvic pain Dysmenorrhea [9,13] Dysuria [9,13] Dyschezia [9,13] Dyspareunia [13] | Pelvic Cystitis/painful bladder [9,13] Irritable bowel syndrome [13] Anal bleeding [13] Chronic endometritis [9,13] Inflammatory bowel disease [9,10,13] Polycystic ovary syndrome [9,10,13] Ovarian cancer [9,13] Uterine Fibroids (leiomyomata) [9,13] Vulvodynia [13] Possible link with recurrent vaginal infections [9,13] |
| Extra pelvic Thoracic [9,13,15] Diaphragm, lungs, pleura, pericardium Liver and spleen [9,13,15] Abdominal wall [9,13,15] Lymph nodes [9,10,13] Brain [9,13,15] Kidney [9,13,15] | Infertility Implantation failure [2,4,9,13] Spontaneous miscarriage [4,9,13] Alterations of tubal structures [4,9,13] Diminished ovarian reserve [9,13] Placenta previa [9] Premature delivery [9] | Extra pelvic Fatigue [2,4,9,13] Fibromyalgia [13] Joint disorders [13] Migraine [2,9,13] Systemic [2,4,9,13] Systemic autoimmune diseases (Lupus, Sjögren’s syndrome, Rheumatoid arthritis) [2,13,16,34] Tissue-specific autoimmune diseases (Thyroiditis, Crohn’s disease, Addison’s disease) [2,13,16,34], Immune-related conditions (allergies, chronic inflammation) Thyroid disorders [13,16,34] Mental health conditions (depression, anxiety) [2,9,13] |
| Cytokine | Role in Endometriosis | Reference |
|---|---|---|
| Proinflammatory cytokines | ||
| IL-1β | Increased levels in the follicular fluid of endometriosis patients. | [26,250,251] |
| IL-1 RA | Increased levels in the follicular fluid of endometriosis patients. It binds active IL-1β, reducing receptor binding and biological activity. | [252,253] |
| IL-2 | Decreased levels in endometriotic lesions. Increase in soluble CD25 in peritoneal fluid. | [254,255,256,257] |
| Increased levels were reported in patients with severe endometriosis. | [258] | |
| IL-3 | Increased levels were reported in endometriotic lesions. | [259,260,261] |
| IL-5 | Increased levels were reported in the follicular fluid of endometriosis patients. | [260,261] |
| IL-6 | Increased levels were reported in the follicular fluid of endometriosis patients. It is a proinflammatory cytokine. | [252,253,261] |
| IL-7 | Higher in the eutopic as compared to the ectopic tissue of endometriosis patients. | [262] |
| Associated with IL-15 in maintaining endometriosis. | [263] | |
| IL-8 | Increased levels were reported in the follicular fluid of endometriosis patients. | [264,265,266] |
| IL-12p40 | Increased levels were reported in the follicular fluid of endometriosis patients. | [267,268] |
| IL-15 | Higher levels were reported in the peritoneal fluid of women with endometriosis. | [263,269,270] |
| IL-16 | Increased levels were reported in the peritoneal fluid of patients with advanced stage endometriosis (III/IV). | [271,272] |
| IL-17 | Elevated levels of IL-17 have been observed in patients during the early stages of the disease. IL-17 promotes the proliferation, invasion, and implantation of endometriotic cells. Additionally, increased IL-17 levels have been linked to higher levels of IL-8, VEGF, CSF-1, and GM-CSF. | [273,274,275] |
| IL-18 | Increased levels have been reported in the peritoneal fluid of endometriosis patients. Affects endometrial receptivity. | [276] |
| IL-23 | Increased levels are observed in the follicular fluid of endometriosis patients, stages III–IV > stages I–II. Involved in IL-17 production and inflammation. | [277] |
| IL-25 | Increased levels were reported in the peritoneal fluid of patients with endometriosis. | [278] |
| IL-31 | Increased levels were reported in the plasma of endometriosis patients | [279] |
| IL-32 | Increased concentrations of IL-32 were reported in the peritoneal fluid of patients with endometriosis. An increase in IL-32 is correlated with elevated levels of IL-8 and CCL2 and enhanced cell proliferation. | [280,281] |
| IL-34 | Increased serum levels in patients with endometriosis. Autocrine production promotes endometriosis. | [282] |
| IL-35 | Enhanced levels are reported in ectopic endometrium. Suppresses immune response, increasing the growth of endometrial cells. | [283] |
| IL-36α, β, γ and Receptor | Increased levels are reported in the peritoneal fluid of endometriosis patients. | [284] |
| IFNγ | Decreased levels are reported in the peritoneal fluid of endometriosis patients. It induces macrophage activation (M1) and enhances the proinflammatory response. | [178,256,257] |
| TNFα | High levels of TNFα are found in patients with endometriosis, particularly at their highest in those with severe endometriosis. | [285,286] |
| CXCL chemokines | In women with endometriosis or polycystic ovary syndrome, the levels of CXCL1-8, 10, 12, 13, and 16 are increased, while CXCL9 and 14 are decreased. | [287] |
| FRACTAL-KINE | Decreased levels were reported in the peritoneal fluid of patients with endometriosis. | [288] |
| Increased levels were reported in patients with endometriosis. | [289] | |
| MCP-1 (CCL2) | Increased follicular fluid levels in endometriosis correlate with severity and attract neutrophils, NK cells, and lymphocytes linked to RANTES and IL-8. | [290,291] |
| Association with hepatocyte growth factor and insulin-like growth factor-1. | [292] | |
| MCP-2/3/4 | Increased levels are reported in patients with endometriosis. | [286] |
| MIP-1α | A decrease in cytokine levels was reported in patients with endometriosis. Increased levels were reported. | [289] |
| MIP-1β | Increased levels were reported in patients with endometriosis. | [289] |
| eotaxin 2/3 | Increased levels were reported in patients with endometriosis. | [289] |
| ENA-78 | Increased levels were reported in patients with endometriosis. | [289] |
| RANTES (CCL5) | Increased levels were reported in patients with endometriosis. | [286,289] |
| MIF-1 | Increased levels were reported in patients with endometriosis. The levels are associated with 17β-estradiol. MIF-1 is a proinflammatory cytokine. | [293,294,295] |
| CSF-1 | Increased levels were reported in patients with severe endometriosis. | [296,297,298] |
| PDGF | Increased levels were reported in the peritoneal fluid of patients with endometriosis. | [298,299,300] |
| VEGF | Increased levels were reported in the peritoneal fluid of patients with endometriosis. Involved in increased vascularization. | [298,299,300,301,302] |
| bFGF | Increased levels were reported in the peritoneal fluid of patients with endometriosis. | [298,299,300,301,303] |
| Anti-inflammatory cytokines | ||
| TGFβ | Patients with severe endometriosis exhibited increased levels of TGFβ, which play a role in the fibrosis observed in these individuals. | [77,304,305] |
| IL-10 | Increased levels were found in the follicular fluid of patients with endometriosis, produced by various cells. | [266,306] |
| IL-19 | A decrease in serum levels of IL-19 has been observed in patients with endometriosis. | [307] |
| IL-22 | A decrease in serum levels was reported in patients with endometriosis. | [308] |
| IL-22 is implicated in endometrial cell invasion in humans and mice and carcinoma cell proliferation. | [309,310] | |
| IL-37 | Increased levels are reported in the peritoneal fluid of endometriosis patients. Involved in anti-inflammatory response in vitro and animal models. | [253,284,311] |
| IL-38 | Increased levels are reported in the peritoneal fluid of endometriosis patients. Involved in anti-inflammatory response. | [284] |
| Mixed effects proinflammatory and antinflammatory | ||
| IL-4 | Increased levels in the follicular fluid of endometriosis patients. | [312,313,314,315,316] |
| IL-13 | Differential expression in ectopic and eutopic endometrium in endometriosis patients. High levels of the cytokine have been associated with infertility. | [314,317] |
| IL-27 | IL-2 + IL-27 are involved in the growth of human endometrial cells in vitro. Its role in endometriosis is still controversial. | [318] |
| IL-33 | Increased serum levels were reported in the peritoneal fluid of patients with deep endometriosis, which could induce an anti-inflammatory response. | [319,320] |
| It is involved in epithelial–mesenchymal transition. | [321,322] | |
| EGF | Increased levels were reported in the peritoneal fluid of endometriosis patients involved in endometrial invasion. | [298,299,300,301,303] |
| GM-CSF | Increased levels were reported in patients with severe endometriosis. It is controversial since it may have local anti-inflammatory effects. | [29,323,324] |
| Autoantibodies against GM-CSF are present in the serum of patients with deep endometriosis. | [325] | |
| Adipokine | Characteristics | References |
|---|---|---|
| Leptin | Elevated leptin levels have been observed in serum and peritoneal fluid in patients with endometriosis. | [327,329,330] |
| Researchers found a positive association between leptin levels and endometriosis in the mouse model. | [21,331] | |
| Controversial results have been reported in humans. | [332] | |
| Elevated local leptin levels in endometriosis lesions are associated with increased transcription factor HIF-1α. | [333] | |
| Endometriosis may be related to dysfunctional adipose tissue, which affects metabolism, browning, body weight regulation, and pain pathways. | [21,334] | |
| Adiponectin | Low circulating adiponectin levels in women are associated with endometriosis. | [335,336] |
| Resistin | Increased concentrations have been reported in women with endometriosis. | [337,338] |
| Research suggests a potential correlation between resistin and IL-23 levels. | [277] | |
| Retinol binding protein 4 (RTB4) | Increased plasma levels of RBP4 have been reported in patients with endometriosis. | [339] |
| RTB4 may play a role in the infiltration of immune cells in human endometriosis. | [340] | |
| Visfatin/NAMPT and resistin | The three adipokines may be secreted locally within the human endometrioma as part of an inflammatory response, regardless of the stage of endometriosis. | [341] |
| Ghrelin, GLP-1, visfatin, GLP-1. | A reduction in ghrelin, GLP-1, glucagon, and visfatin levels in the peritoneal fluid of women with endometriosis may contribute to lesion development by proinflammatory macrophages. | [341] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garmendia, J.V.; De Sanctis, C.V.; Hajdúch, M.; De Sanctis, J.B. Endometriosis: An Immunologist’s Perspective. Int. J. Mol. Sci. 2025, 26, 5193. https://doi.org/10.3390/ijms26115193
Garmendia JV, De Sanctis CV, Hajdúch M, De Sanctis JB. Endometriosis: An Immunologist’s Perspective. International Journal of Molecular Sciences. 2025; 26(11):5193. https://doi.org/10.3390/ijms26115193
Chicago/Turabian StyleGarmendia, Jenny Valentina, Claudia Valentina De Sanctis, Marian Hajdúch, and Juan Bautista De Sanctis. 2025. "Endometriosis: An Immunologist’s Perspective" International Journal of Molecular Sciences 26, no. 11: 5193. https://doi.org/10.3390/ijms26115193
APA StyleGarmendia, J. V., De Sanctis, C. V., Hajdúch, M., & De Sanctis, J. B. (2025). Endometriosis: An Immunologist’s Perspective. International Journal of Molecular Sciences, 26(11), 5193. https://doi.org/10.3390/ijms26115193