
Ferric-Chelate Reductase FRO3 Is Involved in Iron Homeostasis in Table Grape and Enhanced Plant Tolerance to Iron-Deficient Conditions

Abstract
1. Introduction
2. Results
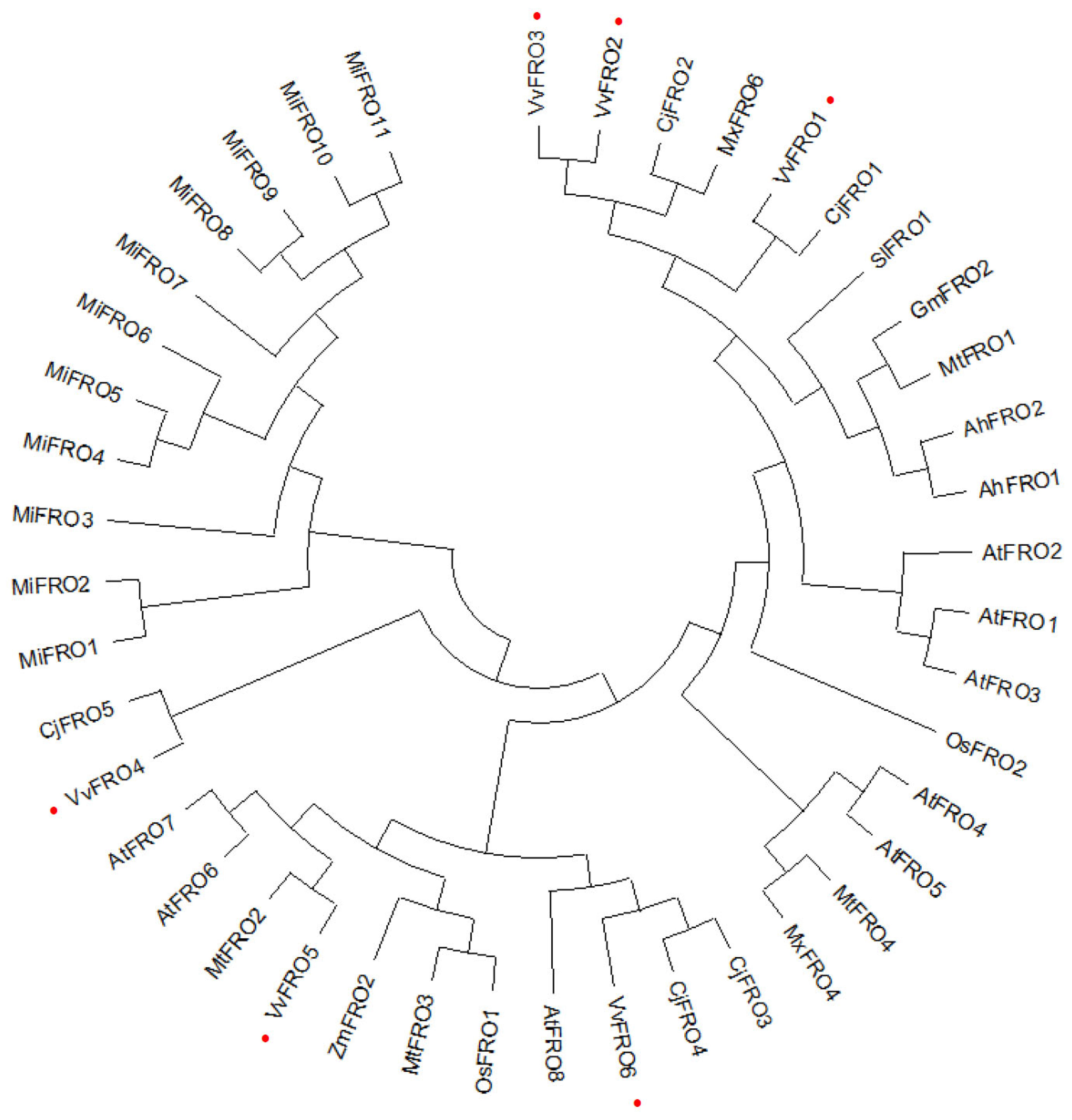
2.1. Identification and Isolation of VvFRO Family Genes in Table Grape
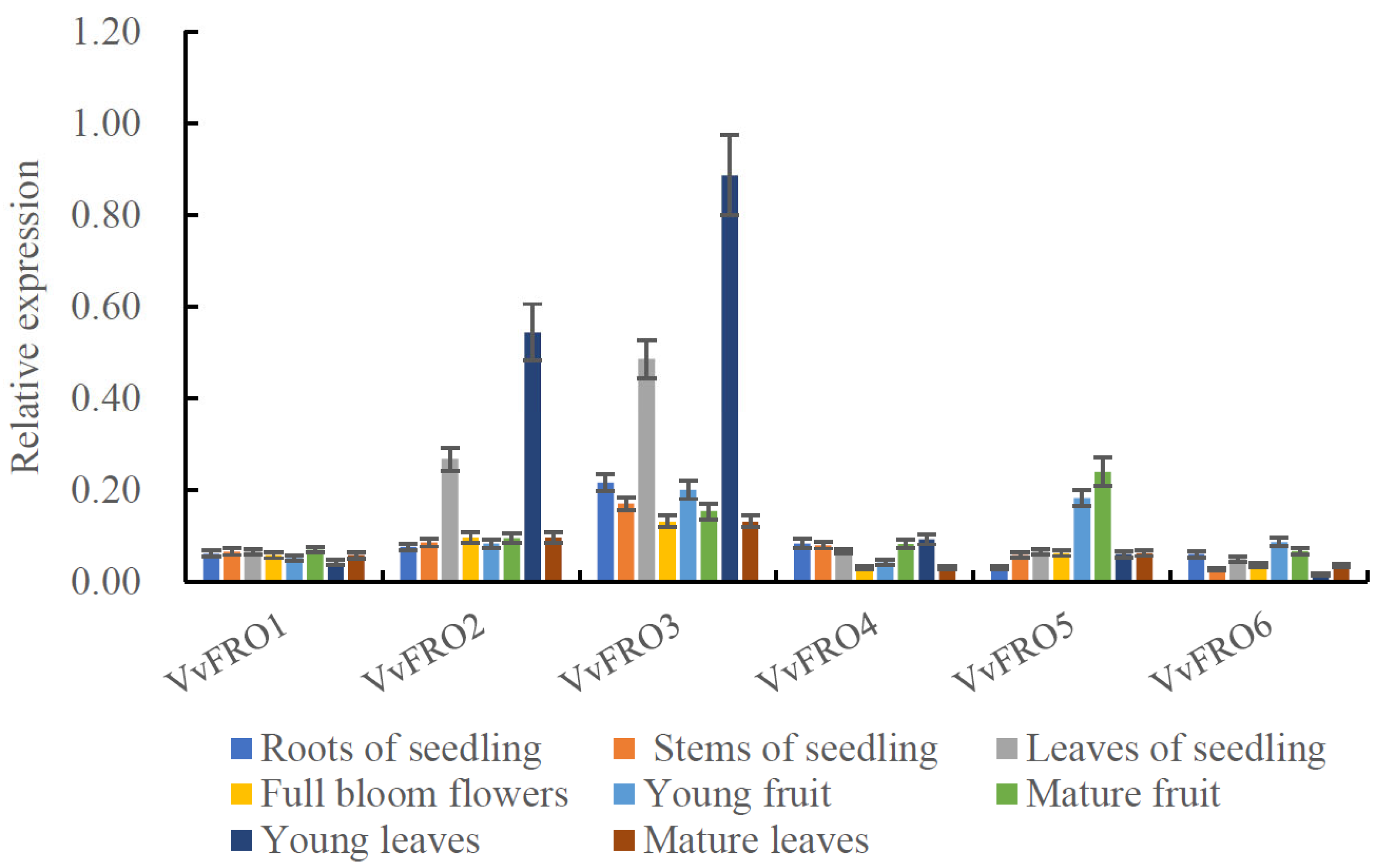
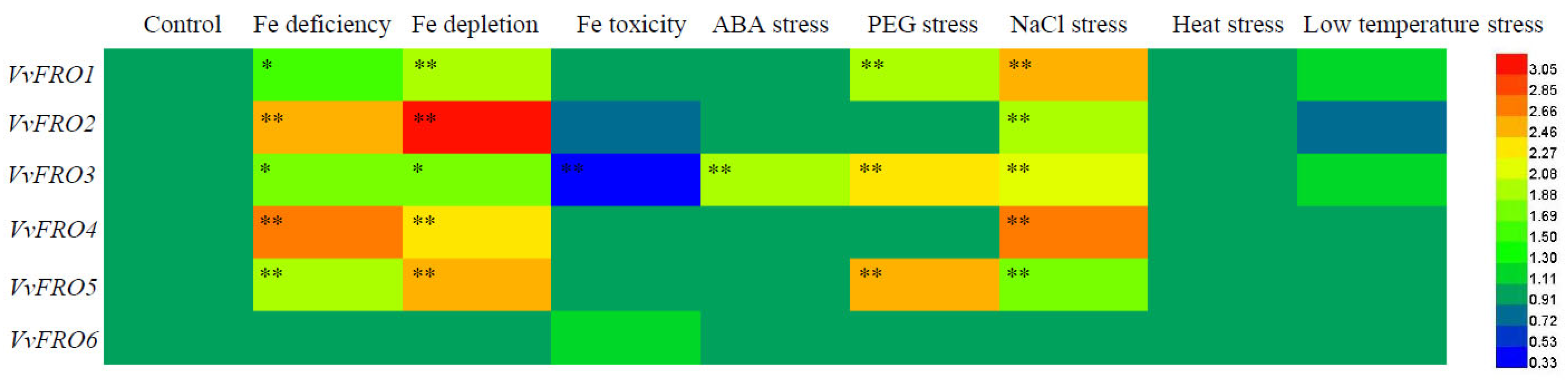
2.2. Expression Profiles of VvFRO Genes
2.3. VvFRO3 Rescued the Impaired Growth of Arabidopsis fro2 Mutant
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. Physiological Analysis
4.3. Isolation and Cloning of VvFRO Family Genes from Table Grape
4.4. Phylogenetic Tree Analysis
4.5. Quantitative Real Time PCR (qRT-PCR)
4.6. Overexpression of VvFRO3 in Arabidopsis fro2 Mutant
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lill, R. Function and biogenesis of iron-sulphur proteins. Nature 2009, 460, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Kermeur, N.; Pédrot, M.; Cabello-Hurtado, F. Iron availability and homeostasis in plants: A review of responses, adaptive mechanisms, and signaling. Methods Mol. Biol. 2023, 2642, 49–81. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Touraine, B.; Briat, J.F.; Gaymard, F.; Rouhier, N. The iron-sulfur cluster assembly machineries in plants: Current knowledge and open questions. Front. Plant Sci. 2013, 4, 259. [Google Scholar] [CrossRef]
- Song, Z.Z.; Lin, S.Z.; Fu, J.Y.; Chen, Y.H.; Zhang, H.X.; Li, J.Z.; Liang, M.X. Heterologous expression of ISU1 gene from Fragaria vesca enhances plant tolerance to Fe depletion in Arabidopsis. Plant Physiol. Biochem. 2022, 184, 65–74. [Google Scholar] [CrossRef]
- Tagliavini, M.; Rombolà, A.D. Iron deficiency and chlorosis in orchard and vineyard ecosystems. Eur. J. Agron. 2001, 15, 72–92. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Montejano-Ramírez, V.; Valencia-Cantero, E. Cross-talk between iron deficiency response and defense establishment in plants. Int. J. Mol. Sci. 2023, 24, 6263. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Sun, W.; Wang, T. The adaptive mechanism of plants to iron deficiency via iron uptake, transport, and homeostasis. Int. J. Mol. Sci. 2019, 20, 2424. [Google Scholar] [CrossRef]
- Fourcroy, P.; Tissot, N.; Gaymard, F.; Briat, J.F.; Dubos, C. Facilitated Fe nutrition by phenolic compounds excreted by the Arabidopsis ABCG37/PDR9 transporter requires the IRT1/FRO2 high-affinity root Fe2+ transport system. Mol. Plant 2016, 9, 485. [Google Scholar] [CrossRef]
- Romheld, V.; Marschener, H. Evidence for a specific uptake system for iron phytosiderophores in roots of grasses. Plant Physiol. 1986, 80, 175–180. [Google Scholar] [CrossRef]
- Muhammad, I.; Jin, X.Q.; Shalmani, A.; Ali, M.; Yi, S.; Gan, P.F.; Li, W.Q.; Liu, W.T.; Chen, K.M. Comparative in silico analysis of ferric reduction oxidase (FRO) genes expression patterns in response to abiotic stresses, metal and hormone applications. Molecules 2018, 23, 1163. [Google Scholar] [CrossRef] [PubMed]
- Wu, H. Molecular and biochemical characterization of the Fe (III) chelate reductase gene family in Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Connolly, E.L.; Campbell, N.H.; Grotz, N.; Prichard, C.L.; Guerinot, M.L. Overexpression of the FRO2 ferric chelate reductase confers tolerance to growth on low iron and uncovers posttranscriptional control. Plant Physiol. 2003, 133, 1102–1110. [Google Scholar] [CrossRef]
- Li, L.; Ye, L.X.; Kong, Q.H.; Shou, H.X. A vacuolar membrane ferric-chelate reductase, OsFRO1, alleviates Fe toxicity in rice (Oryza sativa L.). Front. Plant Sci. 2019, 10, 700. [Google Scholar] [CrossRef]
- Yao, X.L.; Chen, S.C.; He, Y.R. Cloning and analysis of ferric reduction oxidase family genes in Ziyangxiangcheng. In Proceedings of the 2014 Academic Annual Conference of the Chinese Society for Horticultural Science, Wuhan, China, 17–21 March 2014; Summary collection of papers. The Chinese Society for Horticultural Science, Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences: Beijing, China, 2014. (In Chinese). [Google Scholar]
- Zhou, Z.Y. Isolation and functional analysis of ferric reductase gene MxFRO4 and MxFRO6 in Malus xiaojinensis. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2021. (In Chinese). [Google Scholar]
- Ren, C.; Luo, G.; Li, X.; Yao, A.; Liu, W.; Zhang, L.; Wang, Y.; Li, W.; Han, D. MxFRO4 confers iron and salt tolerance through up-regulating antioxidant capacity associated with the ROS scavenging. J Plant Physiol. 2023, 285, 154001. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, J.; Huang, P.; Shao, B.; Li, W.; Liu, W.; Wang, Y.; Xie, L.; Han, M.; Han, D. Overexpression of MxFRO6, a FRO gene from Malus xiaojinensis, increases iron and salt tolerance in Arabidopsis thaliana. In Vitro Cell. Dev. Biol.-Plant 2022, 58, 189–199. [Google Scholar] [CrossRef]
- Han, G.D.; Sun, F.P.; Ren, Z.Y.; Cao, J.W.; Gao, A.P.; Huang, J.F.; Song, Z.Z. Identification and expression pattern analysis of ferric reduction oxidase encoding genes in mango. Chin. J. Trop. Crops 2024, 45, 225–233. (In Chinese) [Google Scholar]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrabde, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef]
- Song, Z.Z.; Wang, X.; Li, M.Y.; Ning, Y.Z.; Shi, S.P.; Yang, G.R.; Zhang, H.X.; Tang, M.L.; Peng, B. Isolation, heterologous expression, and functional determination of an iron regulated transporter (IRT) gene involved in Fe2+ transport and tolerance to Fe2+ deficiency in Vitis vinifera. Plant Growth Regul. 2024, 156, 65. [Google Scholar] [CrossRef]
- Song, Z.Z.; Yang, Y.; Xu, J.L.; Ma, R.J.; YU, M.L. Physiological and transcriptional responses in the iron- sulphur cluster assembly pathway under abiotic stress in peach (Prunus persica L.) seedlings. Plant Cell Tiss. Org. 2014, 117, 419–430. [Google Scholar] [CrossRef]
- Gao, Y.C.; Yu, C.Y.; Zhang, K.; Zhang, H.X.; Zhang, S.Y.; Song, Z.Z. Identification and characterization of the strawberry KT/HAK/KUP transporter gene family in response to K+ deficiency. Acta Physiol. Plant. 2021, 43, 1. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, X.M.; Lin, S.Z.; Wang, J.P.; Tang, M.L.; Huang, J.F.; Gao, T.P.; Zhang, H.X.; Song, Z.Z. Heterologous expression of the MiHAK14 homologue from Mangifera indica enhances plant tolerance to K+ deficiency and salinity stress in Arabidopsis. Plant Growth Regul. 2022, 98, 39–49. [Google Scholar] [CrossRef]
- Peng, B.; Ran, J.G.; Li, Y.Y.; Tang, M.L.; Xiao, H.L.; Shi, S.P.; Ning, Y.Z.; Dark, A.; Guan, X.Q.; Song, Z.Z. Site-directed mutagenesis of VvCYP76F14 (cytochrome P450) unveils its potential for selection in wine grape varieties linked to the development of wine bouquet. J. Agric. Food Chem. 2024, 72, 3683–3694. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.Z.; Tang, M.L.; Xiao, H.L.; Xu, H.H.; Shi, M.; Dark, A.; Xie, Z.Q.; Peng, B. Unraveling the trisubstrate-triproduct reaction mechanisms of wine grape VvCYP76F14 to improve wine bouquet. Food Chem. 2025, 474, 143077. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Genbank No. | Accession ID | Chromosome Location | CDS (bp) | Intron |
|---|---|---|---|---|---|
| VvFRO1 | PQ893568 | GSVIVT01028874001 | chr16:17911251..17919834 reverse | 2184 | 8 |
| VvFRO2 | PQ893569 | GSVIVT01028873001 | chr16:17920090..17930273 reverse | 3537 | 13 |
| VvFRO3 | PQ893570 | GSVIVT01026993001 | chr15:18731174..18734997 reverse | 2082 | 7 |
| VvFRO4 | PQ893571 | GSVIVT01026991001 | chr15:18754824..18759433 reverse | 2145 | 7 |
| VvFRO5 | PQ893572 | GSVIVT01023105001 | chr12:22531017..22539454 forward | 2208 | 8 |
| VvFRO6 | PQ893573 | GSVIVT01007662001 | chr17:10745402..10748530 forward | 2112 | 7 |
| Items | Treatment | Wild Type | fro2 Mutant | #1 fro2/35S::VvFRO3 |
|---|---|---|---|---|
| Fresh weight (g) | Control | 2.63 ± 0.27 a(a) | 1.22 ± 0.13 a(b) | 2.53 ± 0.32 a(a) |
| Fe deficiency | 0.81 ± 0.06 b(a) | 0.38 ± 0.03 b(b) | 0.76 ± 0.08 b(a) | |
| Fe depletion | 0.44 ± 0.03 c(a) | 0.20 ± 0.01 c(b) | 0.37 ± 0.04 c(a) | |
| Dry weight (g) | Control | 0.22 ± 0.02 a(a) | 0.11 ± 0.01 a(b) | 0.21 ± 0.02 a(a) |
| Fe deficiency | 0.07 ± 0.007 b(a) | 0.02 ± 0.003 b(b) | 0.06 ± 0.007 b(a) | |
| Fe depletion | 0.03 ± 0.004 c(a) | 0.01 ± 0.001 bc(b) | 0.03 ± 0.02 c(a) | |
| Primary root length (cm) | Control | 10.54 ± 1.15 a(a) | 5.78 ± 0.61 a(b) | 10.13 ± 1.22 a(a) |
| Fe deficiency | 6.69 ± 0.54 b(a) | 4.25 ± 0.52 b(b) | 6.64 ± 0.56 b(a) | |
| Fe depletion | 4.30 ± 0.41 c(a) | 2.68 ± 0.34 c(b) | 4.32 ± 0.37 c(a) | |
| Lateral root numbers | Control | 24 ± 4 a(a) | 15 ± 3 a(b) | 22 ± 3 a(a) |
| Fe deficiency | 17 ± 3 b(a) | 8 ± 1 b(b) | 17 ± 2 b(b) | |
| Fe depletion | 9 ± 2 c(a) | 4 ± 2 c(b) | 11 ± 2 c(a) | |
| Leaf total chlorophyll (g·kg−1 FW) | Control | 1.67 ± 0.19 a(a) | 1.02 ± 0.022 a(b) | 1.56 ± 0.023 a(a) |
| Fe deficiency | 0.53 ± 0.52 b(a) | 0.38 ± 0.045 b(b) | 0.57 ± 0.061 b(a) | |
| Fe depletion | 0.40 ± 0.043 bc(a) | 0.28 ± 0.034 bc(b) | 0.39 ± 0.046 c(a) | |
| ACO activity [U·(mg protein)−1] | Control | 0.89 ± 0.091 a(a) | 0.53 ± 0.055 a(b) | 0.91 ± 0.098 a(a) |
| Fe deficiency | 0.58 ± 0.063 b(a) | 0.32 ± 0.027 b(b) | 0.57 ± 0.062 b(a) | |
| Fe depletion | 0.39 ± 0.028 bc(a) | 0.21 ± 0.022 c(b) | 0.42 ± 0.031 bc(a) | |
| NiR activity [U·(mg protein)−1] | Control | 1.48 ± 0.16 a(a) | 0.82 ± 0.095 a(b) | 1.39 ± 0.15 a(a) |
| Fe deficiency | 0.83 ± 0.092 b(a) | 0.48 ± 0.051 b(b) | 0.87 ± 0.094 b(a) | |
| Fe depletion | 0.62 ± 0.073 bc(a) | 0.36 ± 0.025 bc(b) | 0.64 ± 0.059 bc(a) | |
| SDH activity [U·(mg protein)−1] | Control | 21.45 ± 2.26 a(a) | 13.72 ± 1.67 a(b) | 20.69 ± 3.04 a(a) |
| Fe deficiency | 15.77 ± 1.62 b(a) | 9.83 ± 0.87 b(b) | 15.36 ± 1.49 b(a) | |
| Fe depletion | 10.68 ± 1.47 c(a) | 6.79 ± 0.85 c(b) | 9.99 ± 1.23 c(a) | |
| Fe content (g·kg−1 DW) | Control | 13.33 ± 1.45 a(a) | 7.89 ± 0.65 a(b) | 12.03 ± 1.51 a(a) |
| Fe deficiency | 9.62 ± 0.91 b(a) | 5.01 ± 0.63 b(b) | 8.98 ± 0.91 b(a) | |
| Fe depletion | 4.77 ± 0.42 c(a) | 2.62 ± 0.02 c(b) | 4.58 ± 0.05 c(a) |
| Gene | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| VvFRO1 | TGGAAGCAAAGGAAGCTCTTA | GAGGAGGTTGGCACCTTGTT |
| VvFRO2 | GCCGTCATCCATCTCCCATC | CTCTGACCAGAGGCCGAAAG |
| VvFRO3 | GCTCCTTCAACTCCGAGCAA | GGGCTGCTTATAGACTTATTATC |
| VvFRO4 | TGCTTGCCTTGATGATCTTCCT | GCCGATCTTTTGGGCTGCTTT |
| VvFRO5 | CCTTTTGGAGTGGTCTCTGCT | GGCGGAGAAGAACTGATCCC |
| VvFRO6 | CCAAATCATTGAAGCGGCAGT | ATCATTGCAGTCCCAAGCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Wang, C.; Cui, Y.; Shi, M.; Tang, M.; Song, Z. Ferric-Chelate Reductase FRO3 Is Involved in Iron Homeostasis in Table Grape and Enhanced Plant Tolerance to Iron-Deficient Conditions. Int. J. Mol. Sci. 2025, 26, 5172. https://doi.org/10.3390/ijms26115172
Wang J, Wang C, Cui Y, Shi M, Tang M, Song Z. Ferric-Chelate Reductase FRO3 Is Involved in Iron Homeostasis in Table Grape and Enhanced Plant Tolerance to Iron-Deficient Conditions. International Journal of Molecular Sciences. 2025; 26(11):5172. https://doi.org/10.3390/ijms26115172
Chicago/Turabian StyleWang, Jianping, Chenxiao Wang, Yutong Cui, Matthew Shi, Meiling Tang, and Zhizhong Song. 2025. "Ferric-Chelate Reductase FRO3 Is Involved in Iron Homeostasis in Table Grape and Enhanced Plant Tolerance to Iron-Deficient Conditions" International Journal of Molecular Sciences 26, no. 11: 5172. https://doi.org/10.3390/ijms26115172
APA StyleWang, J., Wang, C., Cui, Y., Shi, M., Tang, M., & Song, Z. (2025). Ferric-Chelate Reductase FRO3 Is Involved in Iron Homeostasis in Table Grape and Enhanced Plant Tolerance to Iron-Deficient Conditions. International Journal of Molecular Sciences, 26(11), 5172. https://doi.org/10.3390/ijms26115172