

Differences in the Temporal Kinetics of the Metabolic Responses to Salinity Between the Salt-Tolerant Thellungiella salsuginea and the Salt-Sensitive Arabidopsis thaliana Reveal New Insights in Salt Tolerance Mechanisms

Abstract
1. Introduction
2. Results
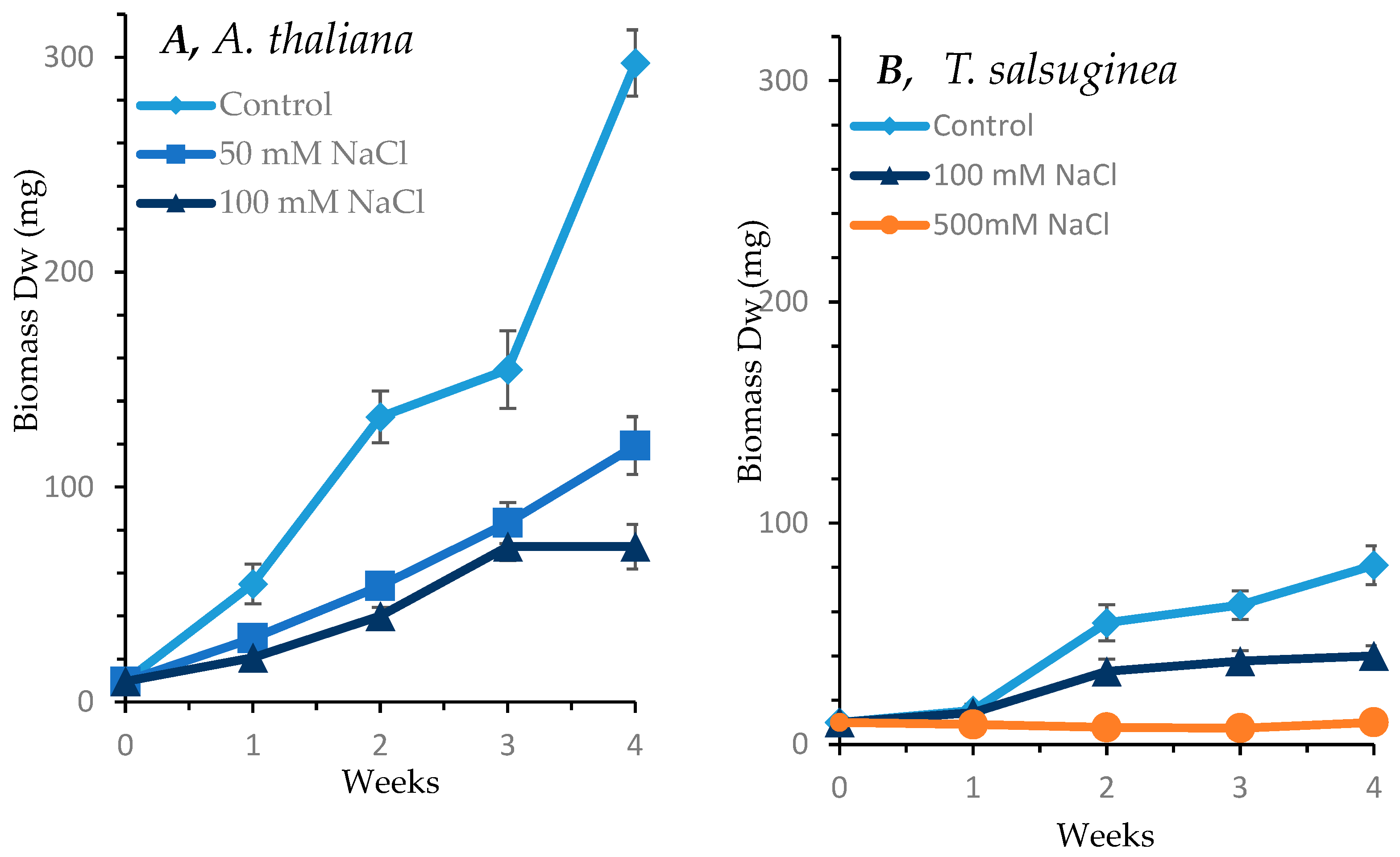
2.1. Effect of Salinity on Growth of A. Thaliana and T. salsuginea
2.2. Effect of Salinity on Photosynthesis in A. thaliana and T. salsuginea
2.3. Effect of Salinity on Ion Uptake: Na+ and K+
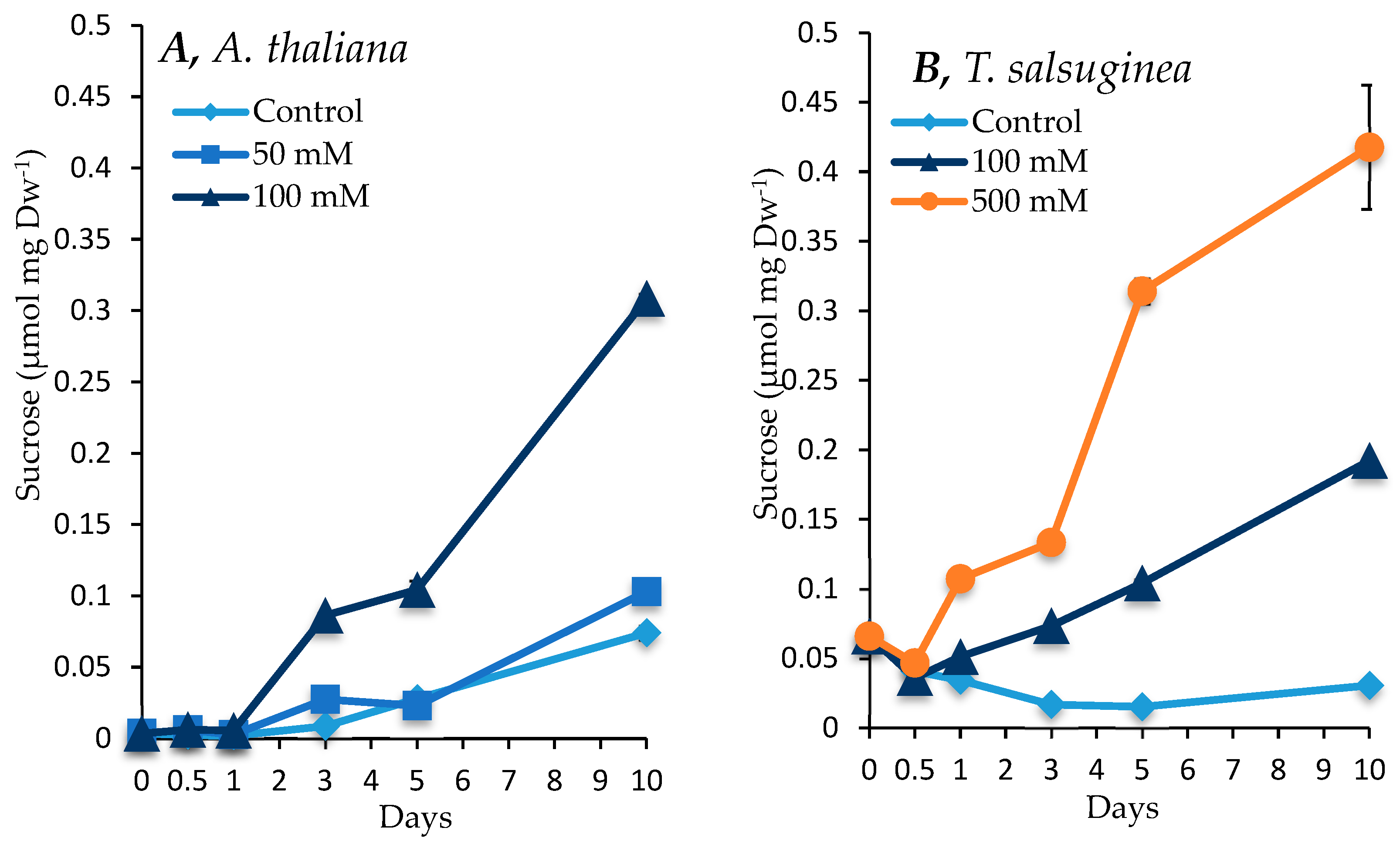
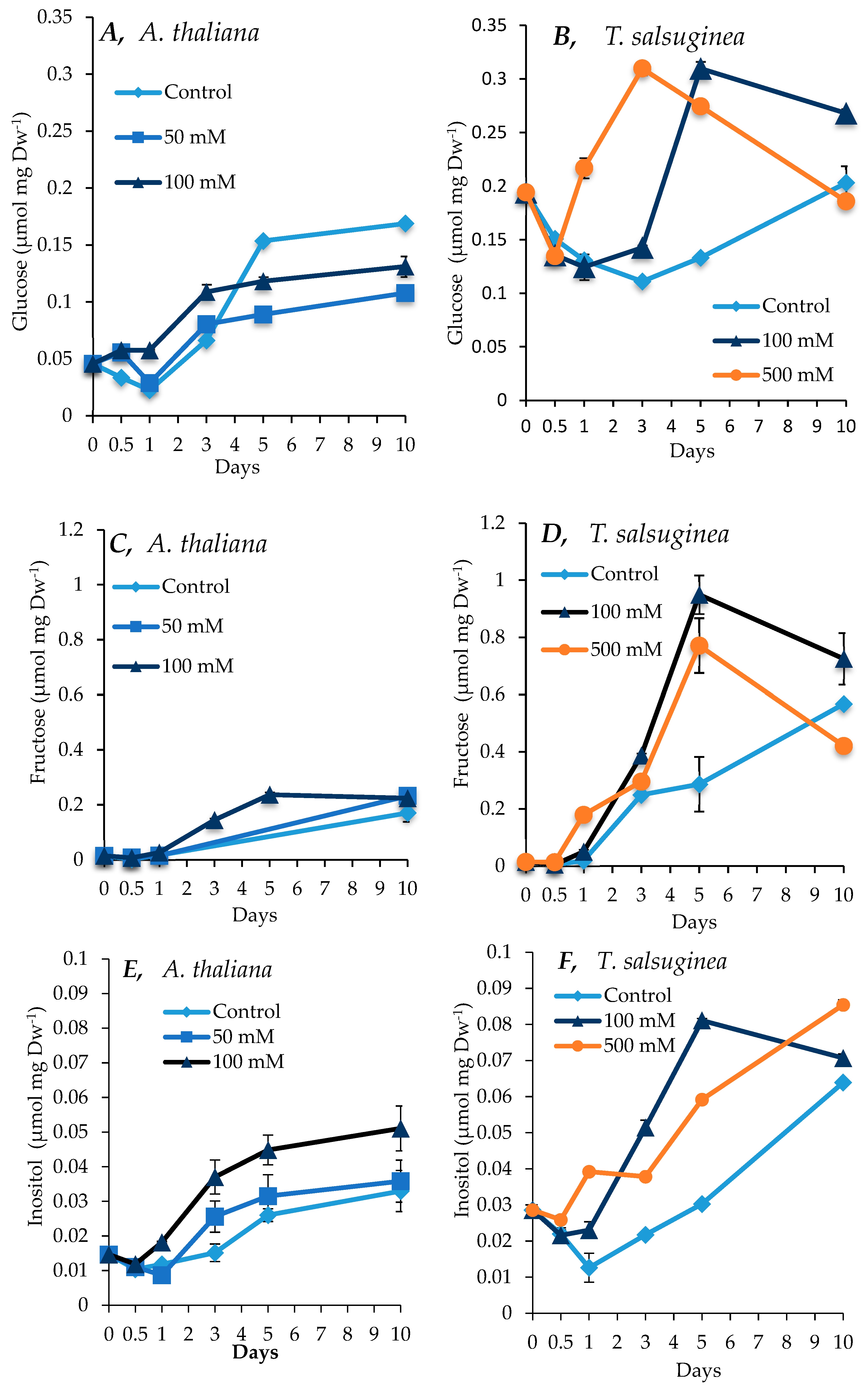
2.4. Effect of Salinity on Accumulation of Sugars and Sugar Alcohols
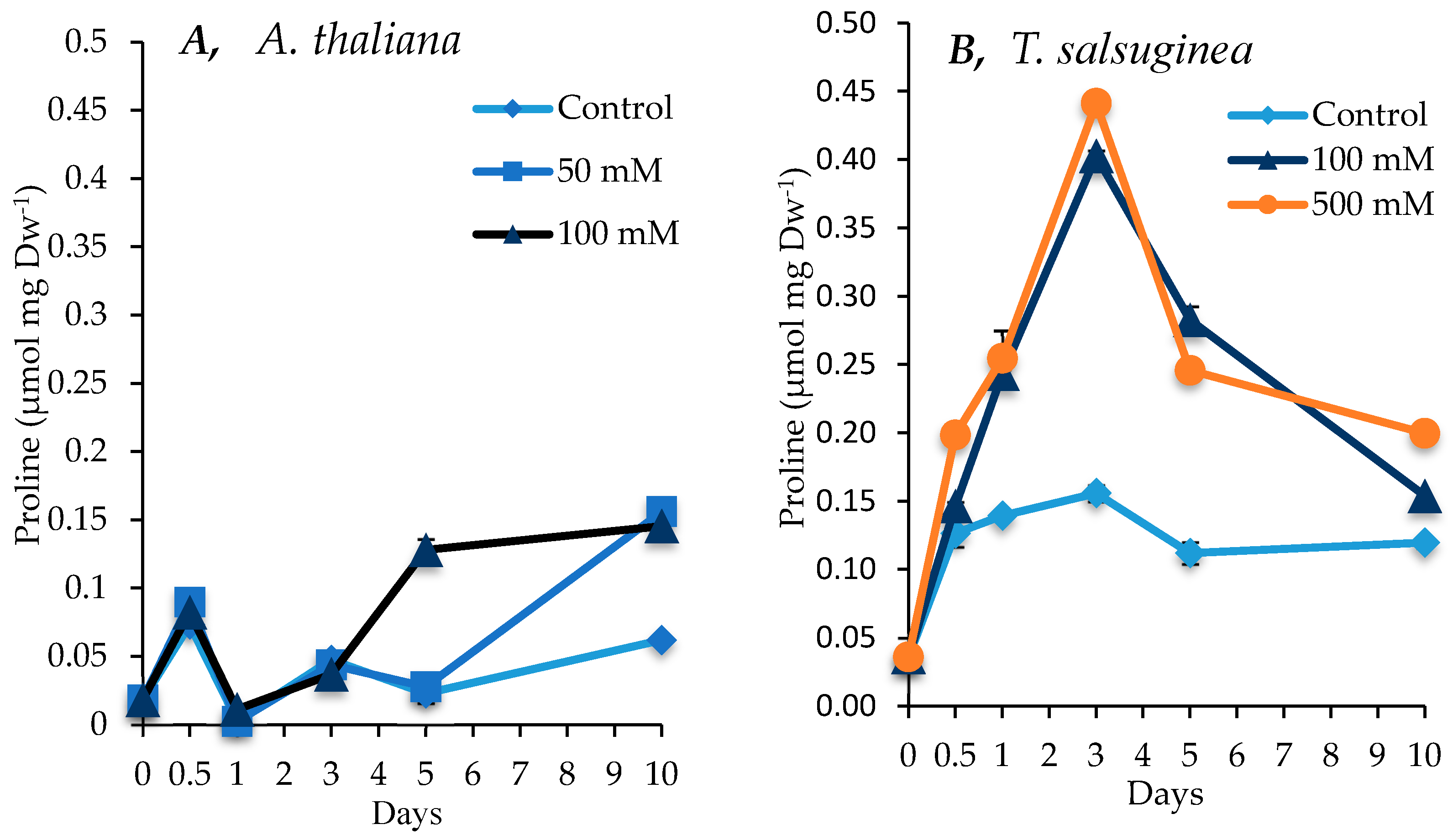
2.5. Effect of Salinity on Proline Accumulation
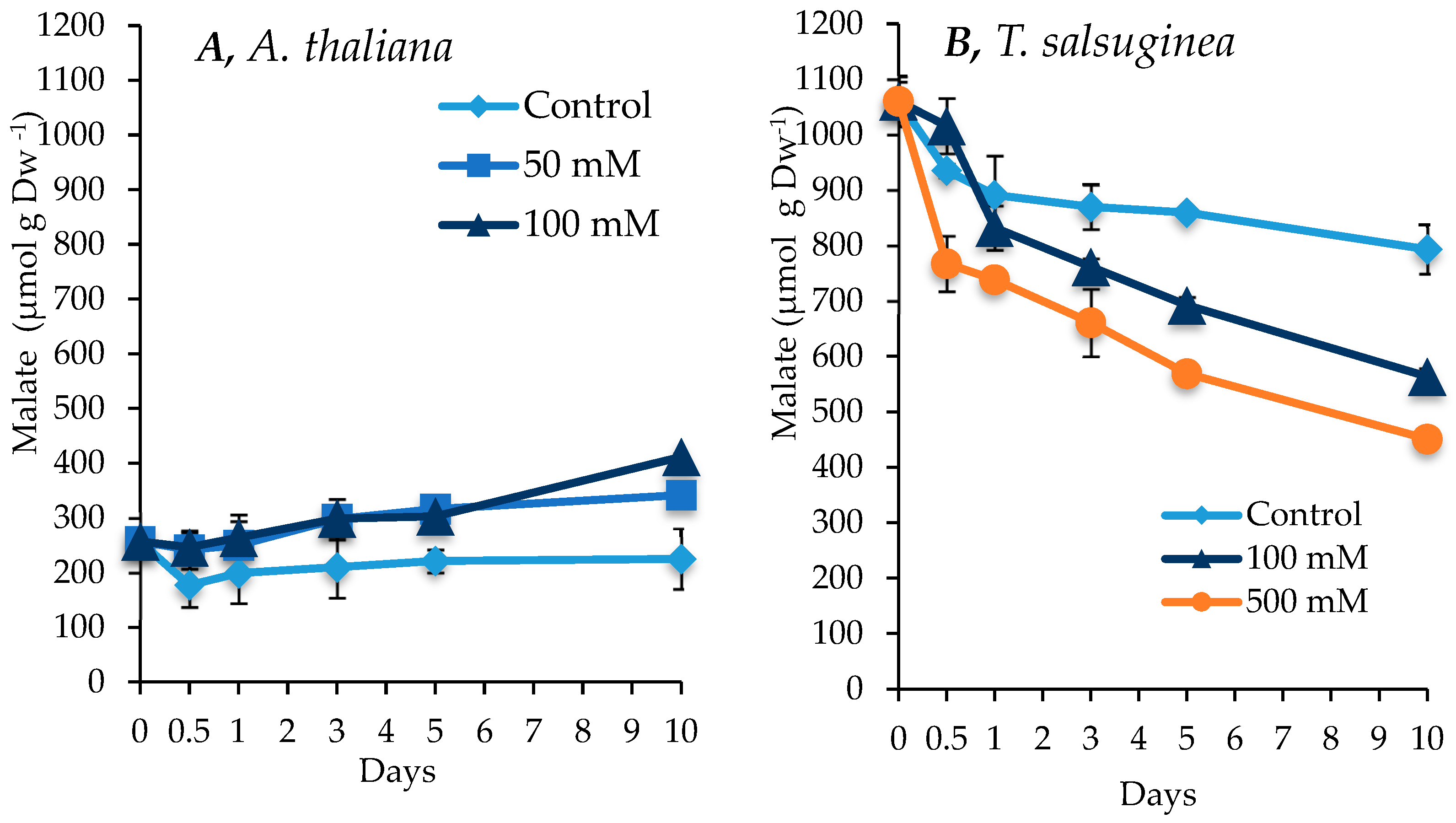
2.6. Effect of Salinity on Malate Accumulation
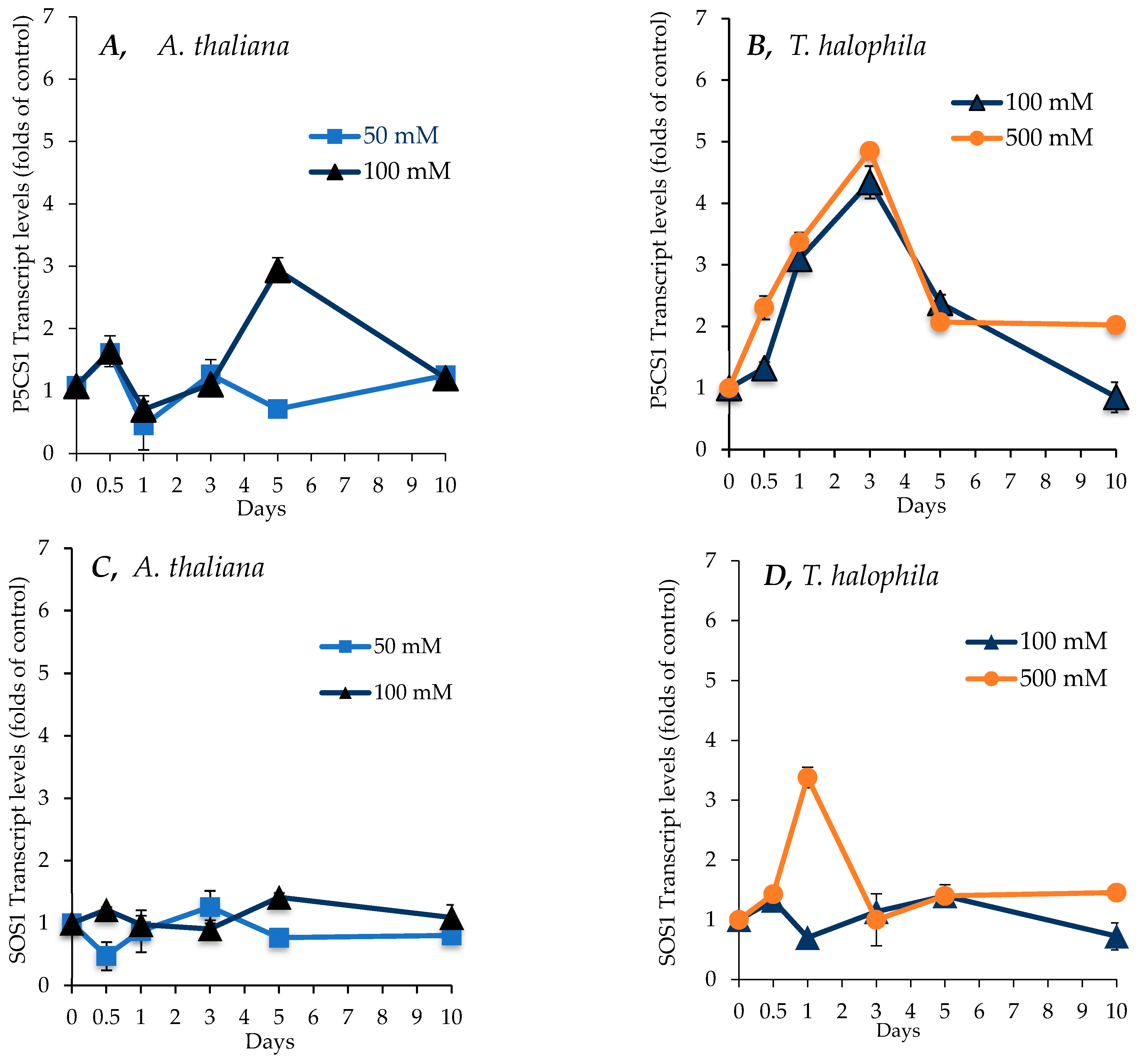
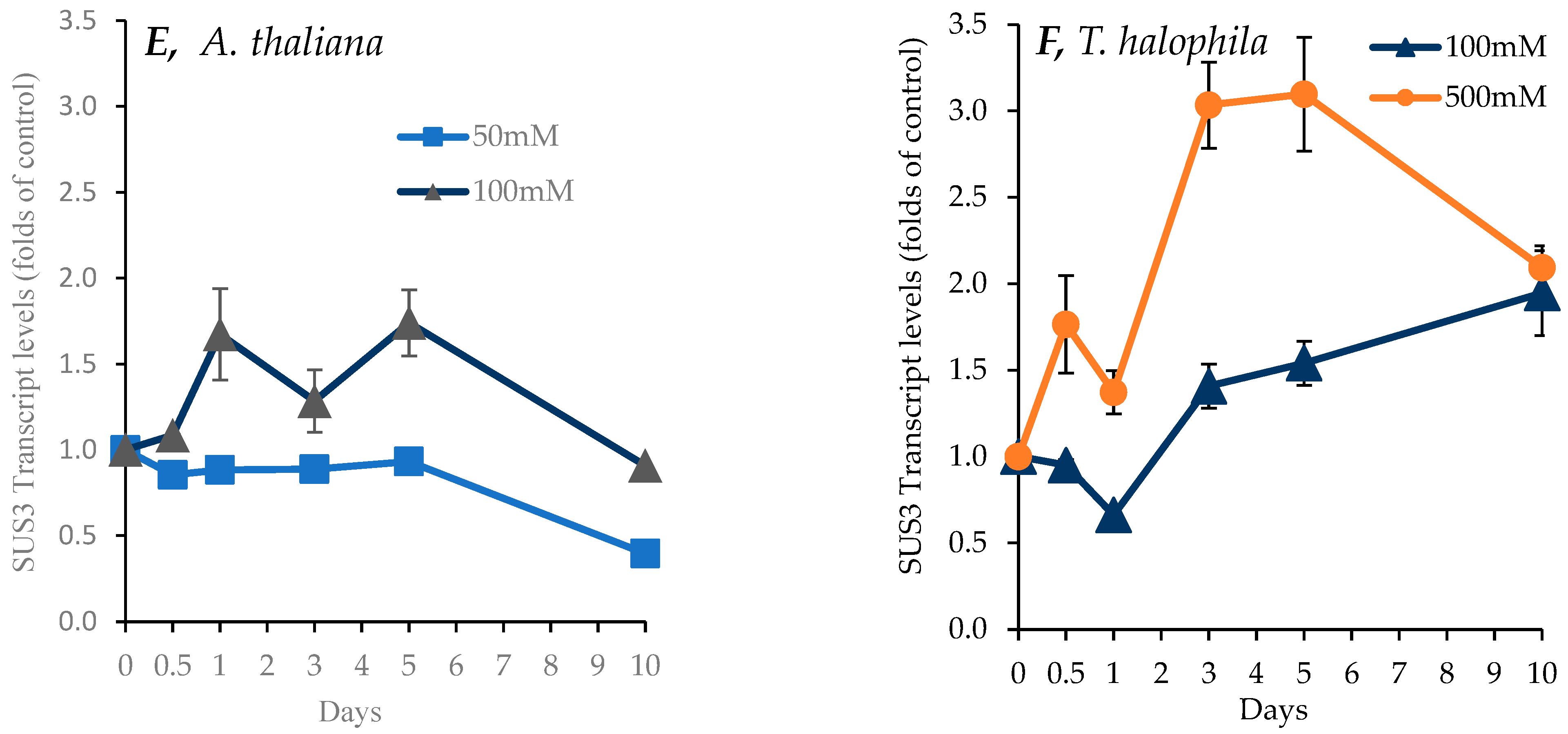
2.7. Effect of Salinity on Transcript Levels for P5CS1, SOS1, and SUS1 Genes in Shoots of A. thaliana and T. salsuginea
3. Discussion
3.1. Impact of Salinity on Growth and Photosynthetic Capacities
3.2. Ion Selectivity: Na+ and K+ Content Under Salt Stress
3.3. Impact of Salinity on Metabolite Accumulation
4. Materials and Methods
4.1. Plant Growth and Salt Treatment
4.2. Determination of Growth Rate
4.3. Determination of Sodium and Potassium Contents
4.4. Determination of Proline Content
4.5. Analyses of Sugars and Sugar Alcohols
4.6. Determination of Malate Content
4.7. RNA Extraction and qRT-PCR
4.8. Δ1-Pyrroline-5-Carboxylate Synthase 1 (P5CS1), Amplicon Size: 80 bp
4.9. Salt Overly Sensitive 1 (SOS1), Amplicon Size: 130 bp
4.10. Sucrose Synthase 3 (SUS3), Amplicon Size: 188 bp
4.11. Ubiquitin 10 (UBQ10), Amplicon Size: 58 bp
4.12. Sampling and Statistical Analysis
5. Conclusions
5.1. Potassium
5.2. Osmoregulation
5.3. Organic Acids
5.4. The Kinetics of the Salt Response
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, Y.P.; Funk, C.; Erban, A.; Kopka, J.; Köhl, K.I.; Zuther, E.; Hincha, D.K. Salt stress responses in a geographically diverse collection of Eutrema/Thellungiella spp. accessions. Funct. Plant Biol. 2016, 43, 590–606. [Google Scholar] [CrossRef] [PubMed]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 29, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Mostafa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A Review on Plant Responses to Salt Stress and Their Mechanisms of Salt Resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef]
- Dong, S.Y.; Zhang, J.; Beckles, D.M. A pivotal role for starch in the reconfiguration of 14C-partitioning and allocation in Arabidopsis thaliana under short-term abiotic stress. Sci. Rep. 2018, 8, 931. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–469. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Singh, P.; Choudhary, K.K.; Chaudhary, N.; Gupta, S.; Sahu, M.; Tejaswini, B.; Sarkar, S. Salt stress resilience in plants mediated through osmolyte accumulation and its crosstalk mechanism with phytohormones. Front. Plant Sci. 2022, 13, 1006617. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Delauney, A.; Verma, D. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar] [CrossRef]
- Louis, P.; Galinski, E.A. Characterization of genes for the biosynthesis of the compatible solute ectoine from Marinococcus halophilus and osmoregulated expression in Escherichia coli. Microbiology 1997, 143, 1141–1149. [Google Scholar] [CrossRef]
- Raven, J.A. Regulation of pH and generation of osmolarity in vascular plants: A cost-benefit analysis in relation to efficiency of use of energy, nitrogen and water. New Phytol. 1985, 101, 25–77. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Argamasilla, R.; Gomez-Cadenas, A. Commom and divergent physiological, hormonal and metabolic responses of Arabidopsis thaliana and Thellungiella halophila to water and salt stress. J. Plant Physiol. 2010, 167, 1342–1350. [Google Scholar] [CrossRef]
- Widodo; Patterson, J.H.; Newbigin, E.; Tester, M.; Bacic, A.; Roessner, U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J. Exp. Bot. 2009, 60, 4089–4103. [Google Scholar] [CrossRef]
- Niu, X.; Bressan, R.; Pardo, J.M. Ion homeostasis in NaCl stress environment. Plant Physiol. 1995, 109, 735–742. [Google Scholar] [CrossRef]
- Zhu, J.K.; Liu, J.; Xion, L. genetic analysis of salt tolerance in Arabidopsis: Evidence for a critical role of potassium nutrition. Plant Cell 1998, 10, 1181–1191. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt-tolerance in non-halophytes. Ann. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Su, Y.; Luo, W.; Lin, W.; Ma, L.; Kabir, M. Model of cation transportation mediated by high-affinity potassium transporters (HKTs) in higher plants. Biol. Proced. Online 2015, 17, 1. [Google Scholar] [CrossRef]
- Oh, D.-H.; Leidi, E.; Zhang, Q.; Hwang, S.; Li, Y.; Quintero, F.; Jiang, X.; D’Urzo, M.; Lee, S.; Zhao, Y.; et al. Loss of halophytism by interference with SOS1 expression. Plant Physiol. 2009, 151, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.-H.; Lee, S.Y.; Bressan, R.A.; Yun, D.-J.; Bohnert, H.J. Intracellular consequences of SOS1 deficiency during salt stress. J. Exp. Bot. 2010, 61, 1205–1213. [Google Scholar] [CrossRef]
- Volkov, V.; Wang, B.; Dominy, P.J.; Fricke, W.; Amtmann, A. Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, possesses effective mechanisms to discriminate between potassium and sodium. Plant Cell Environ. 2004, 27, 1–14. [Google Scholar] [CrossRef]
- Ali, Z.; Park, H.C.; Ali, A.; Oh, D.-H.; Aman, R.; Kropornika, A.; Hong, H.; Choi, W.; Chung, W.S.; Kim, W.-Y.; et al. TsHKT1;2, a HKT1 homolog from the extremophile Arabidopsis relative Thellungiella salsuginea, shows K+ specificity in the presence of NaCl. Plant Physiol. 2012, 158, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.-Q.; Guo, H.; Wang, S.-M.; Zhao, B.; Zhang, J.-L.; Ma, Q.; Yin, H.-J.; Bao, A.-K. The Photosynthesis, Na+/K+ Homeostasis and Osmotic Adjustment of Atriplex canescens in Response to Salinity. Front. Plant Sci. 2016, 7, 848. [Google Scholar] [CrossRef]
- Ghars, M.A.; Parre, E.; Debez, A.; Bordenave, M.; Richard, L.; Leport, L.; Bouchereau, A.; Savouré, A.; Abdelly, C. Comparative salt tolerance analysis between Arabidopsis thaliana and Thellungiella halophila, with special emphasis on K+/Na+ selectivity and proline accumulation. J. Plant Physiol. 2008, 165, 588–599. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Ozturk, M.; Unal, B.T.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Loewus, F.A.; Murthy, P.P.N. Myo-Inositol metabolism in plants. Plant Sci. 2000, 105, 1–19. [Google Scholar] [CrossRef]
- Gil, R.; Boscaiu, M.; Lull, C.; Bautista, I.; Vicente, A.L.A.O. Are soluble carbohydrates ecologically relevant for salt tolerance in halophytes? Funct. Plant Biol. 2013, 40, 805–818. [Google Scholar] [CrossRef]
- Scheibe, R. Malate valves to balance cellular energy supply. Physiol. Plant. 2004, 120, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Lance, C.; Rustin, P. The central role of malate in plant metabolism. Physiol. Veg. 1984, 22, 625–641. [Google Scholar]
- Sanchez, D.H.; Siahpoosh, M.R.; Roessner, U.; Kopka, M.U.a.J. Plant metabolomics reaveals conserved and divergent metabolic responses to salinity. Physiol. Plant. 2008, 132, 209–219. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Vries, R.S.M.; de Boer, A.H. Effect of Salt Stress on Growth, Na+ Accumulation and Proline Metabolism in Potato (Solanum tuberosum) Cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Zhang, Z.; Wang, J.Y.; Oh, D.H.; Dassanayake, M.; Liu, B.; Huang, Q.; Sun, H.X.; Xia, R.; Wu, Y.; et al. Insights into salt tol-erance from the genome of Thellungiella salsuginea. Proc. Natl. Acad. Sci. USA 2012, 109, 12219–12224. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Potential Biochemical Indicators of Salinity Tolerance in Plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Singroha, G.; Kumar, S.; Gupta, O.P.; Singh, G.P.; Sharma, P. Uncovering the Epigenetic Marks Involved in Mediating Salt Stress Tolerance in Plants. Front. Genet. 2022, 13, 811732. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rashid, M.M.; Vaishnav, A.; Verma, R.K.; Sharma, P.; Suprasanna, P.; Gaur, R.K. Epigenetic regulation of salinity stress responses in cereals. Mol. Biol. Rep. 2022, 49, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Inan, G.; Zhang, Q.; Li, P.; Wang, Z.; Cao, Z.; Zhang, H.; Zhang, C.; Quist, T.M.; Goodwin, S.M.; Zhu, J.; et al. Salt Cress, a halophyte and cryophyte Arabidopsis relative model system and its applicability to molecular genetic analyses of growth and development of extremophiles. Plant Physiol. 2004, 135, 1718–1737. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Hohorst, H.J. L-Malate estimation with Malate dehydrogenase and NAD. In Methods in Enzymatic Analysis; Bergmeyer, H.V., Ed.; Chemie: Weinheim, Germany, 1970; Volume 8, pp. 1544–1548. [Google Scholar]
- Taybi, T.; Cushman, J. Signaling Events Leading to Crassulacean Acid Metabolism Induction in the Common Ice Plant. Plant Physiol. 1999, 121, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Kant, S.; Kant, P.; Raveh, S.; Barak, S. Evidence that differential gene expression between the halophyte, Thellungiella halophila, and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline. Plant Cell Environ. 2006, 29, 1220–1234. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite/Gene | Correlation (%) | |
|---|---|---|
| A. thaliana | T. salsuginea | |
| Proline/P5CS1 | 66 | 87 |
| Sucrose/SUS3 | −32 | 89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Taybi, T. Differences in the Temporal Kinetics of the Metabolic Responses to Salinity Between the Salt-Tolerant Thellungiella salsuginea and the Salt-Sensitive Arabidopsis thaliana Reveal New Insights in Salt Tolerance Mechanisms. Int. J. Mol. Sci. 2025, 26, 5141. https://doi.org/10.3390/ijms26115141
Sharma A, Taybi T. Differences in the Temporal Kinetics of the Metabolic Responses to Salinity Between the Salt-Tolerant Thellungiella salsuginea and the Salt-Sensitive Arabidopsis thaliana Reveal New Insights in Salt Tolerance Mechanisms. International Journal of Molecular Sciences. 2025; 26(11):5141. https://doi.org/10.3390/ijms26115141
Chicago/Turabian StyleSharma, Aayush, and Tahar Taybi. 2025. "Differences in the Temporal Kinetics of the Metabolic Responses to Salinity Between the Salt-Tolerant Thellungiella salsuginea and the Salt-Sensitive Arabidopsis thaliana Reveal New Insights in Salt Tolerance Mechanisms" International Journal of Molecular Sciences 26, no. 11: 5141. https://doi.org/10.3390/ijms26115141
APA StyleSharma, A., & Taybi, T. (2025). Differences in the Temporal Kinetics of the Metabolic Responses to Salinity Between the Salt-Tolerant Thellungiella salsuginea and the Salt-Sensitive Arabidopsis thaliana Reveal New Insights in Salt Tolerance Mechanisms. International Journal of Molecular Sciences, 26(11), 5141. https://doi.org/10.3390/ijms26115141