
Investigating the Role of Gut Microbiota in the Pathogenesis and Progression of Rheumatoid Arthritis in a Collagen-Induced Arthritis Mouse Model

 , , ,
, , ,
Abstract
1. Introduction
2. Results
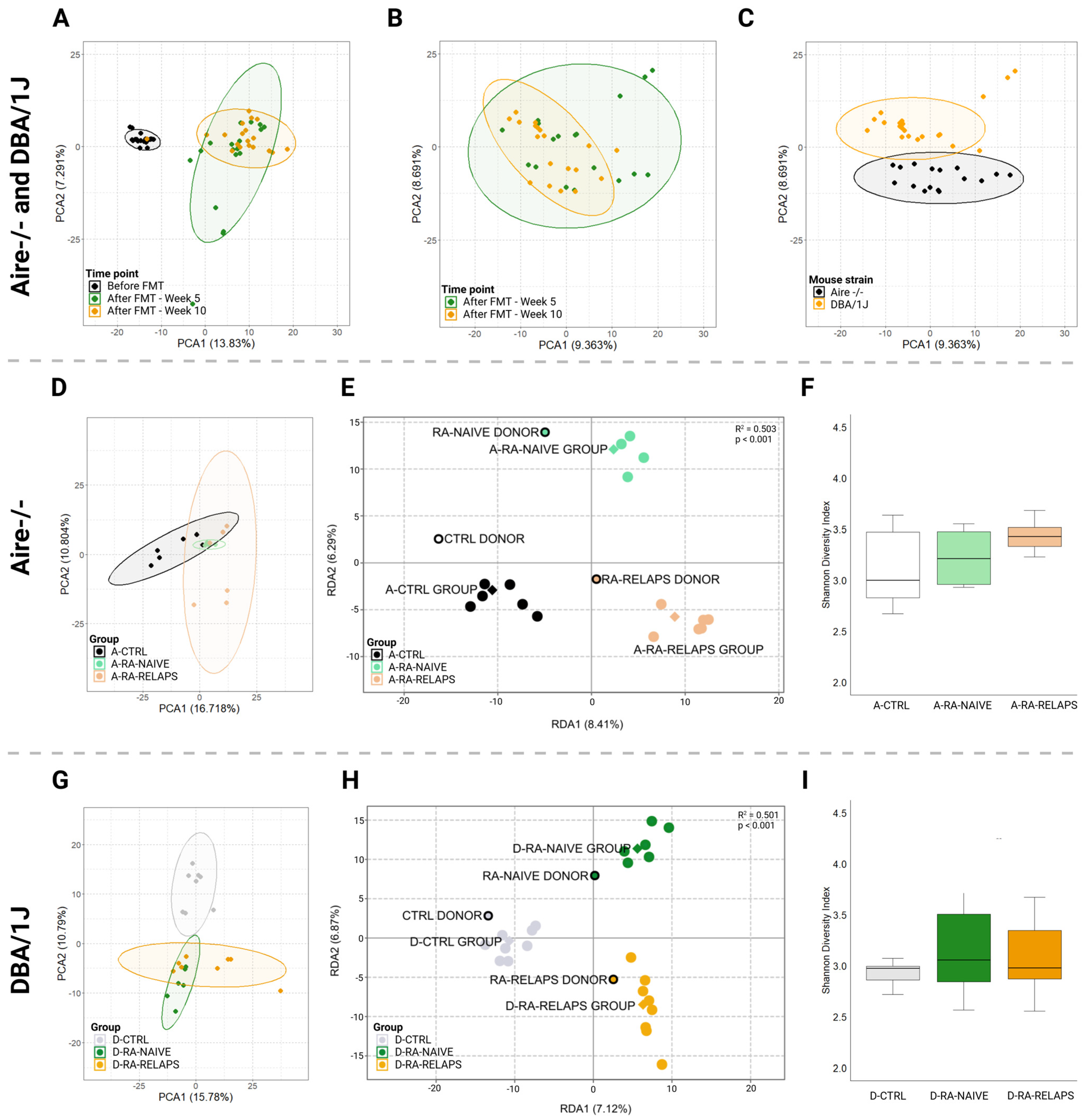
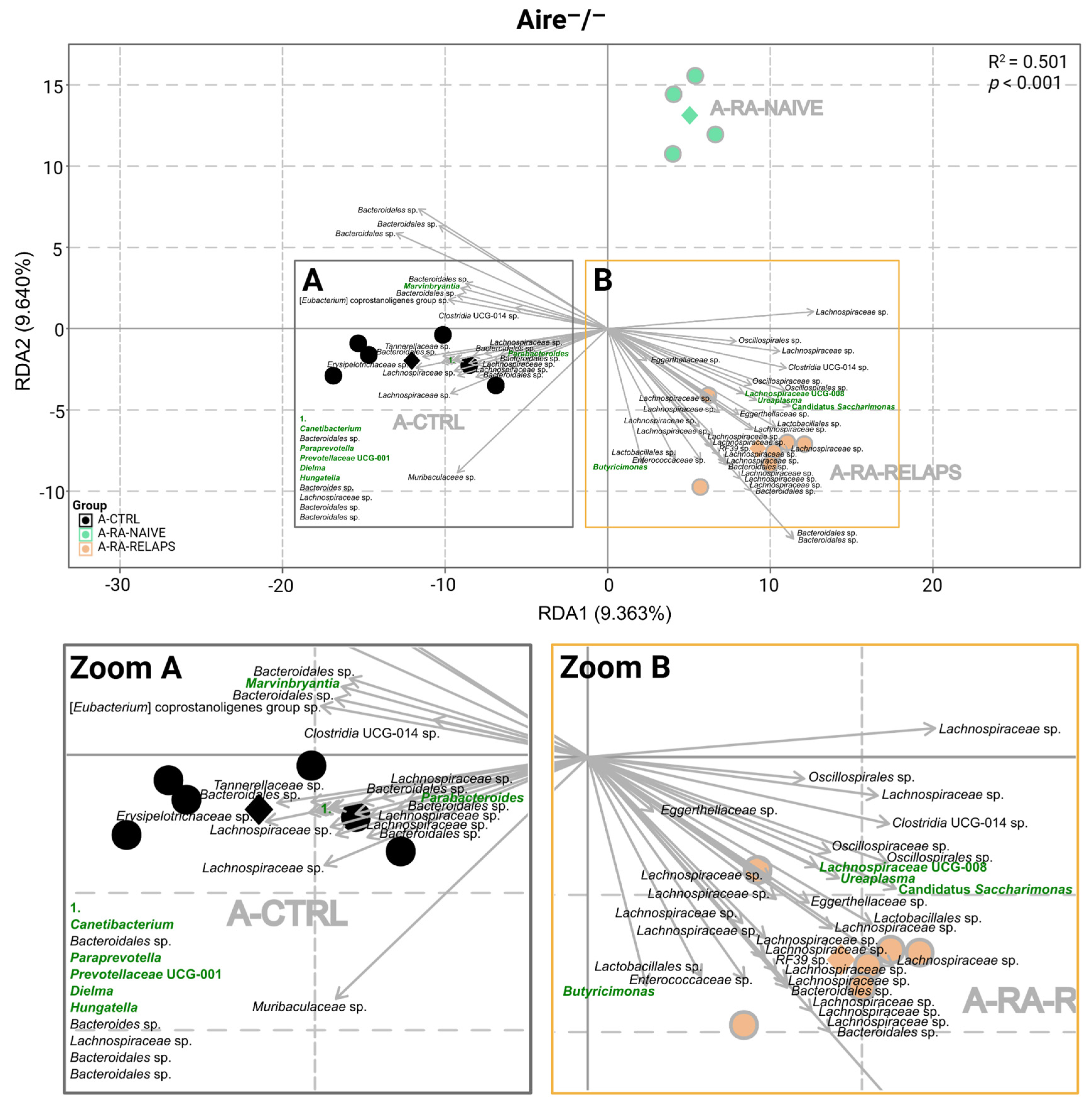
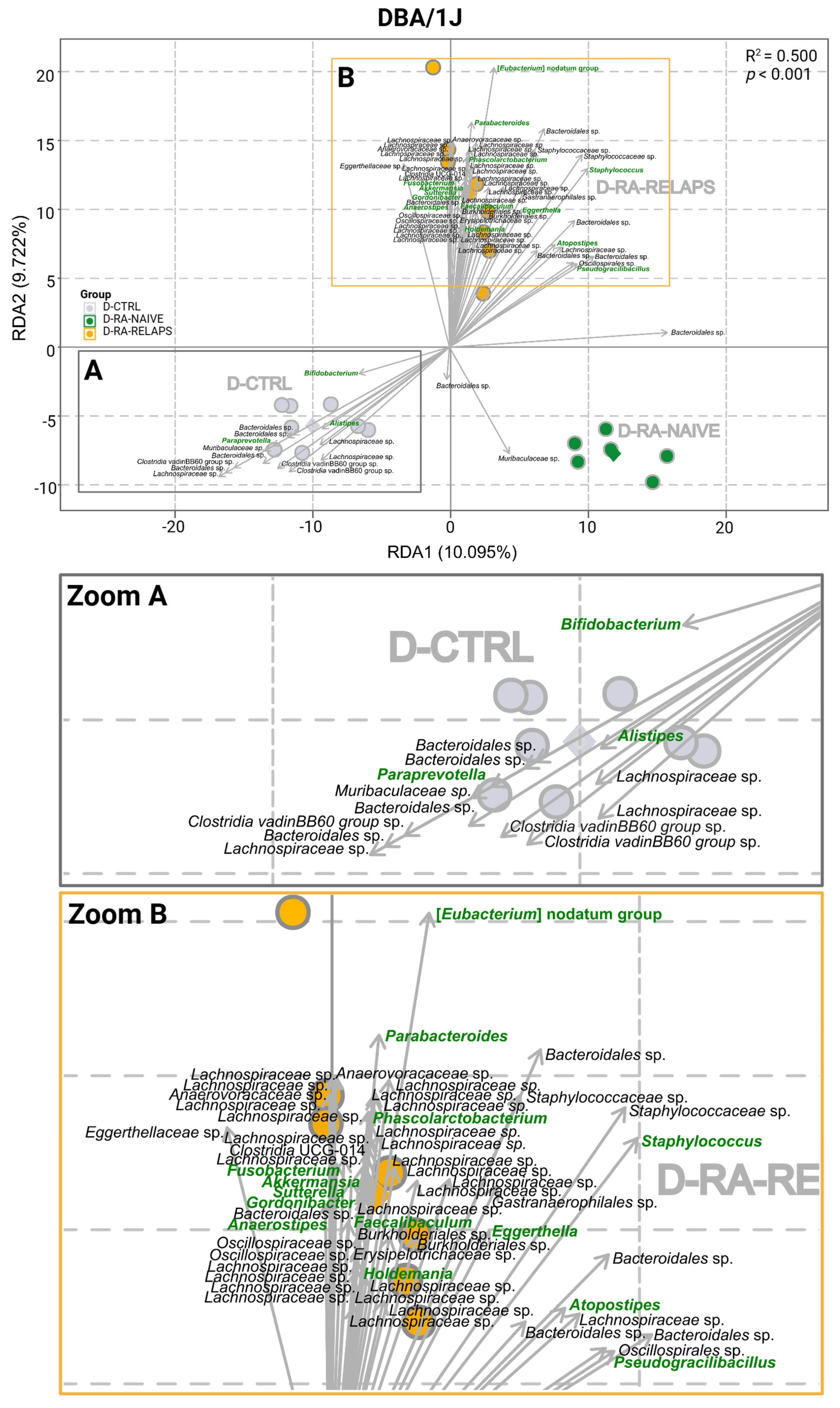
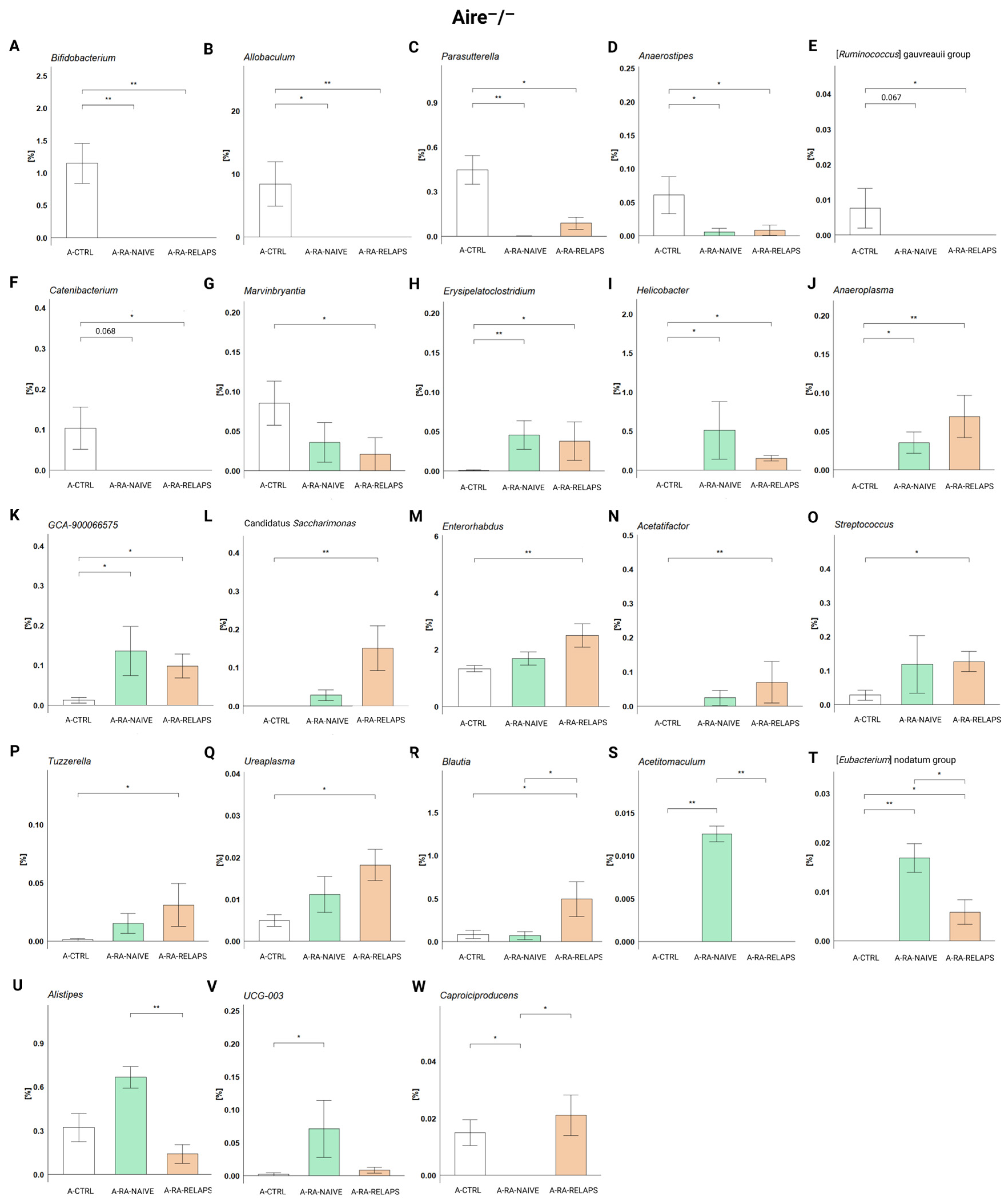
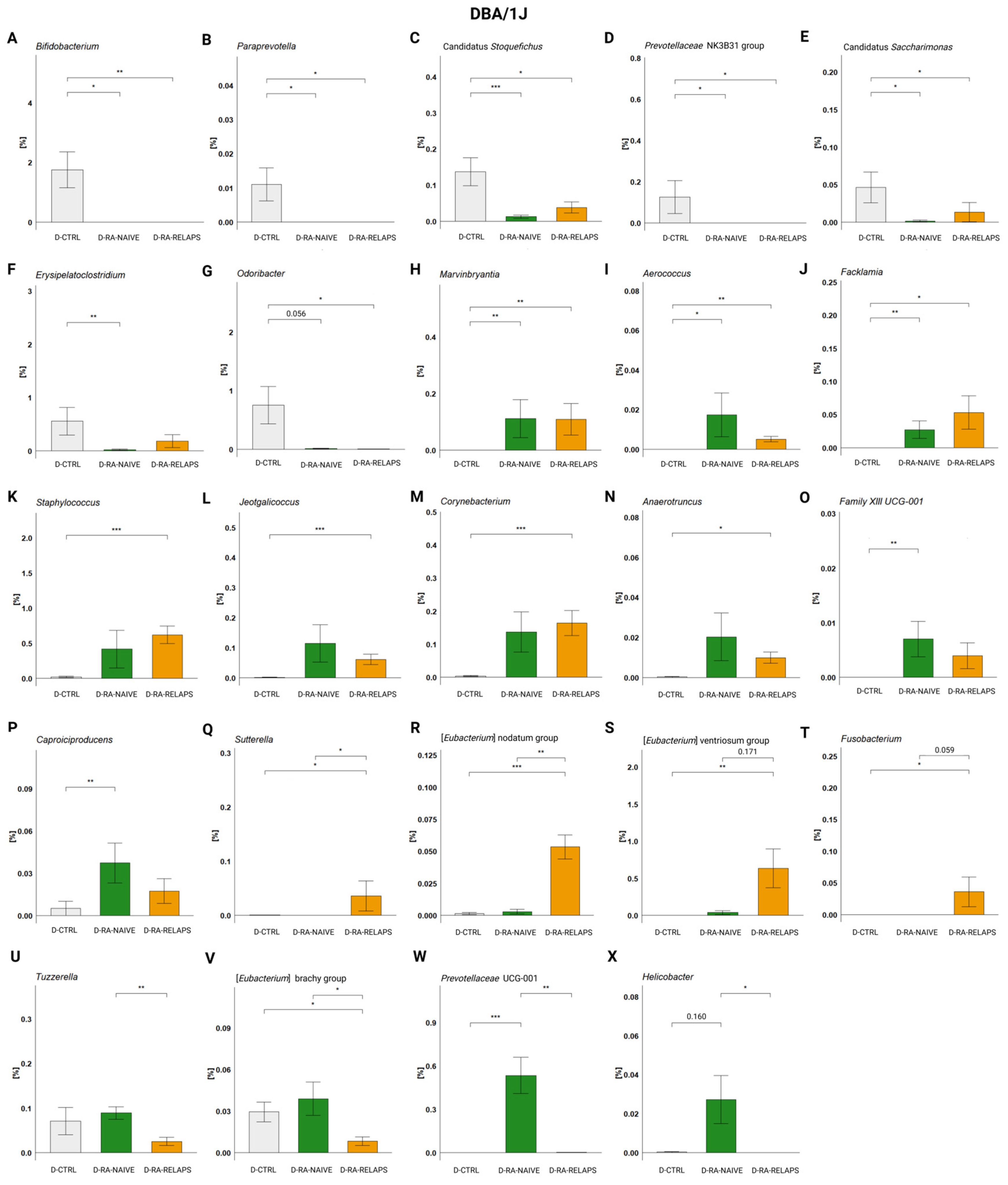
2.1. Gut Microbiota Differences
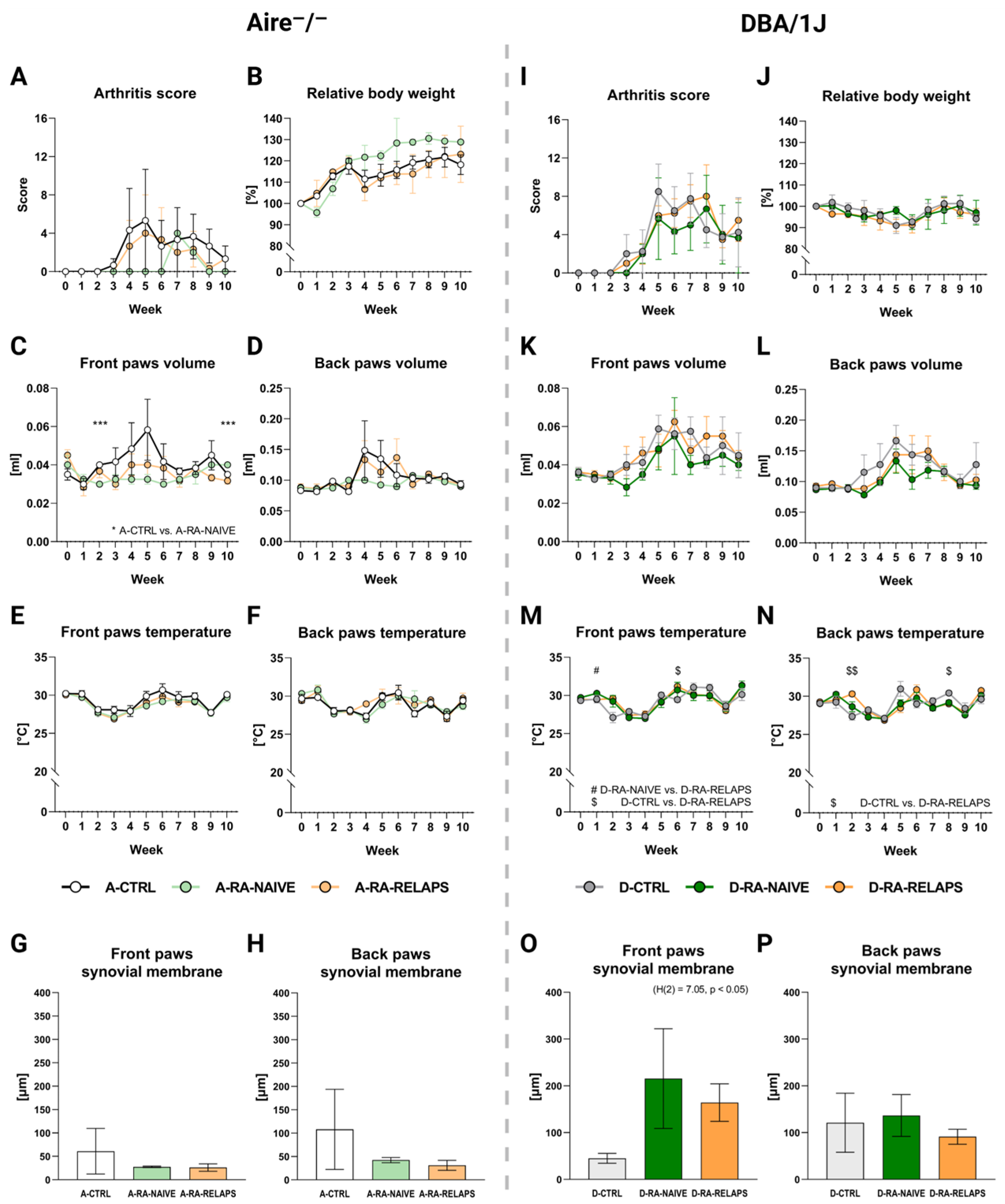
2.2. Severity of Arthritis
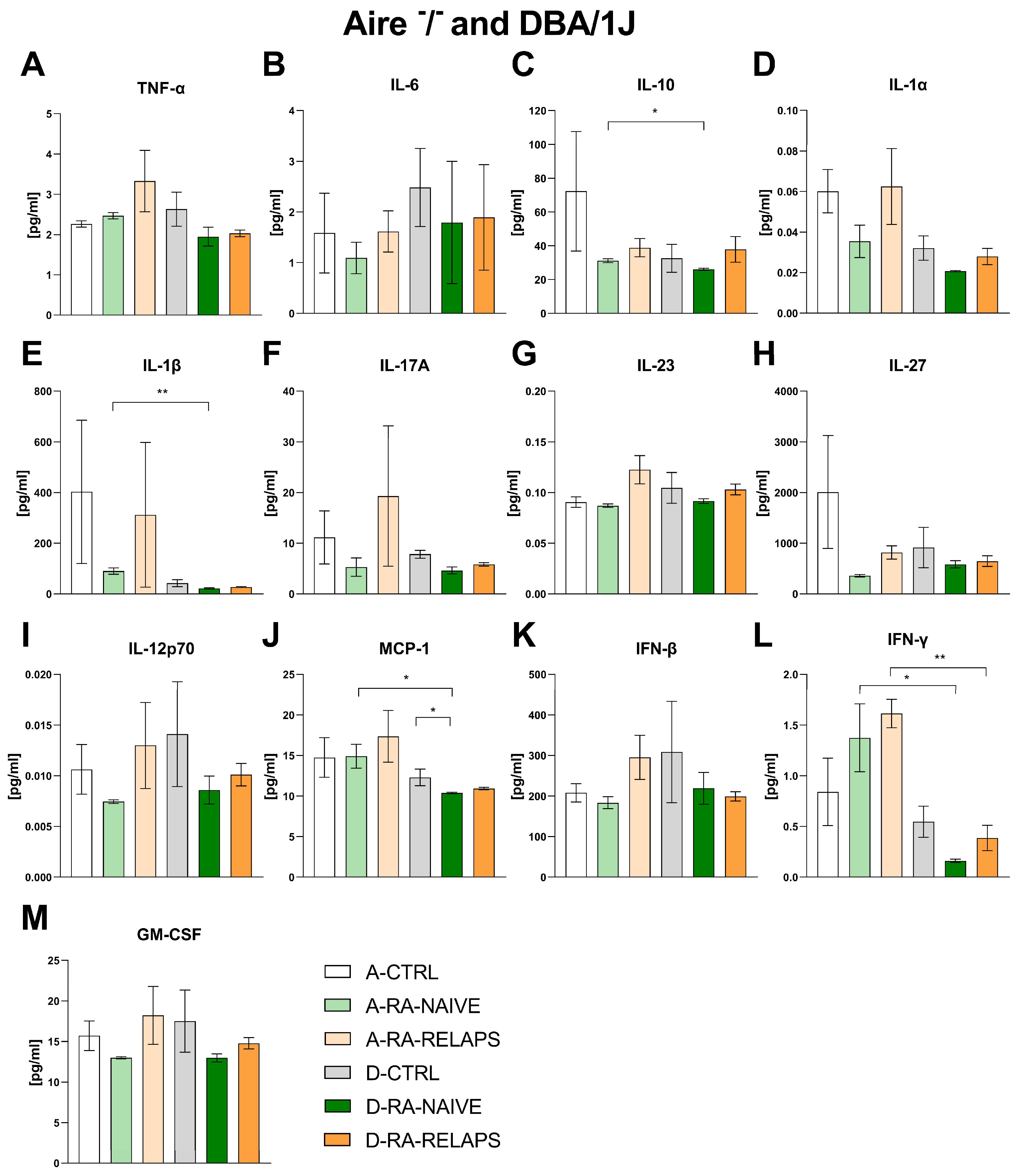
2.3. Inflammatory Cytokines
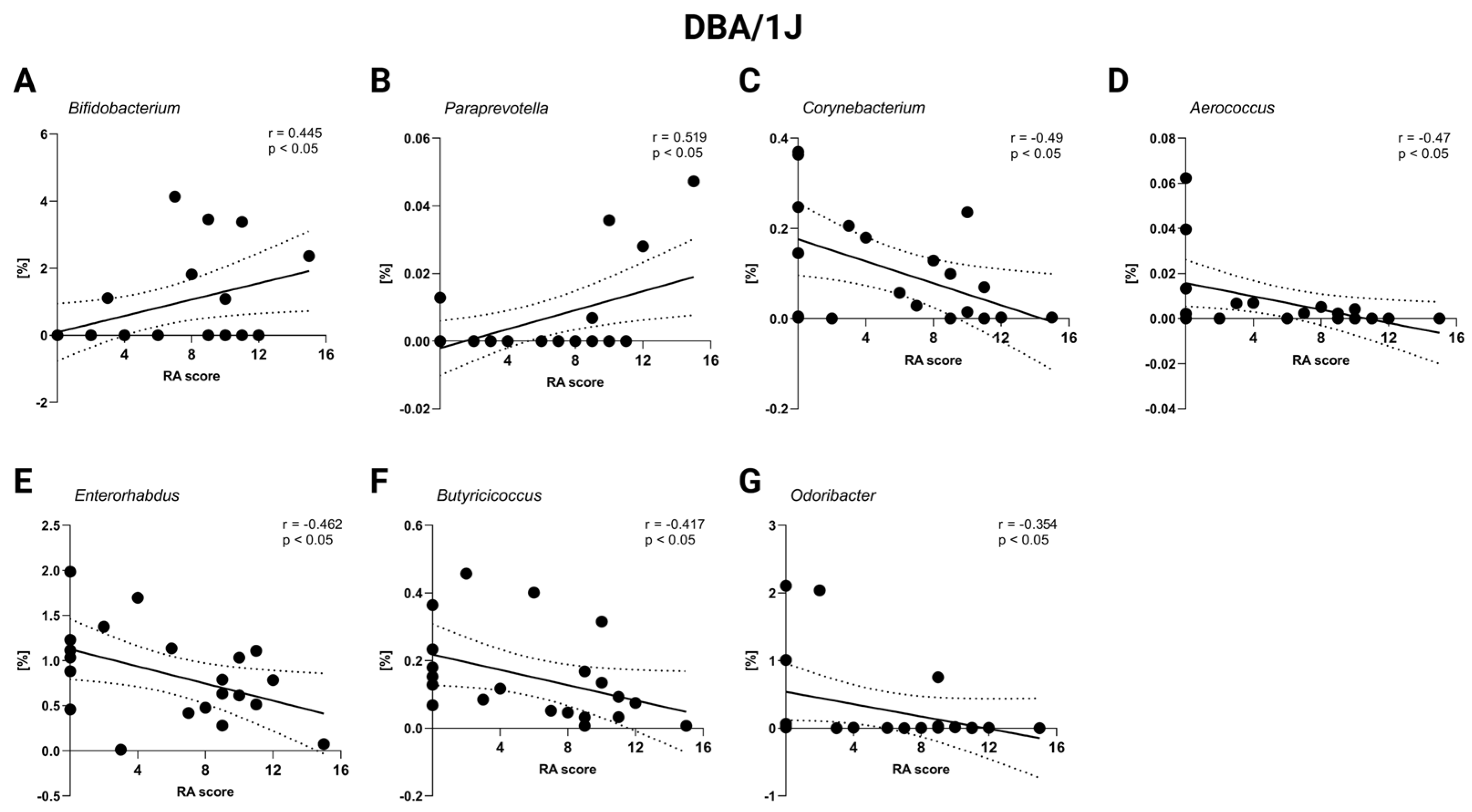
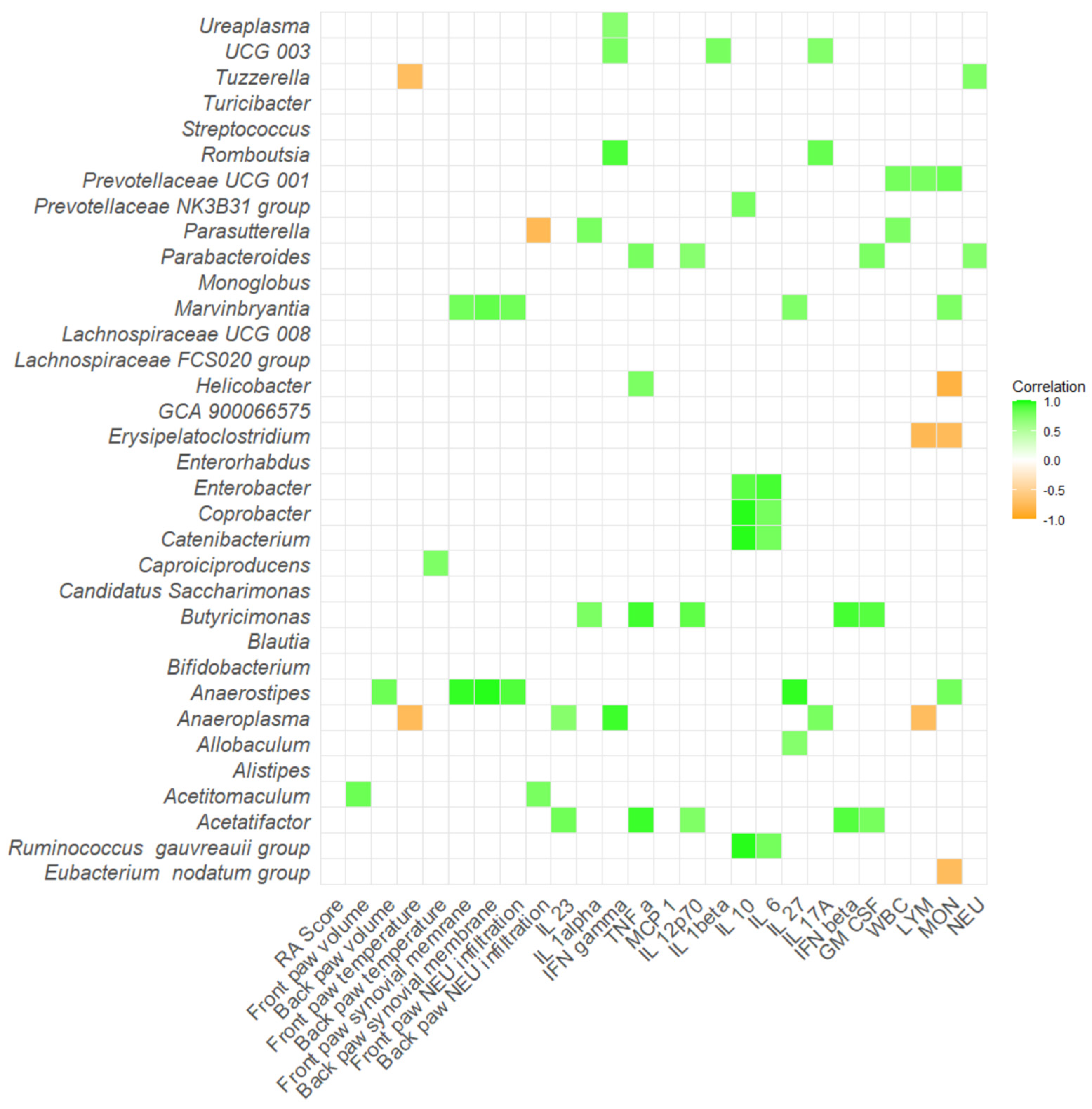
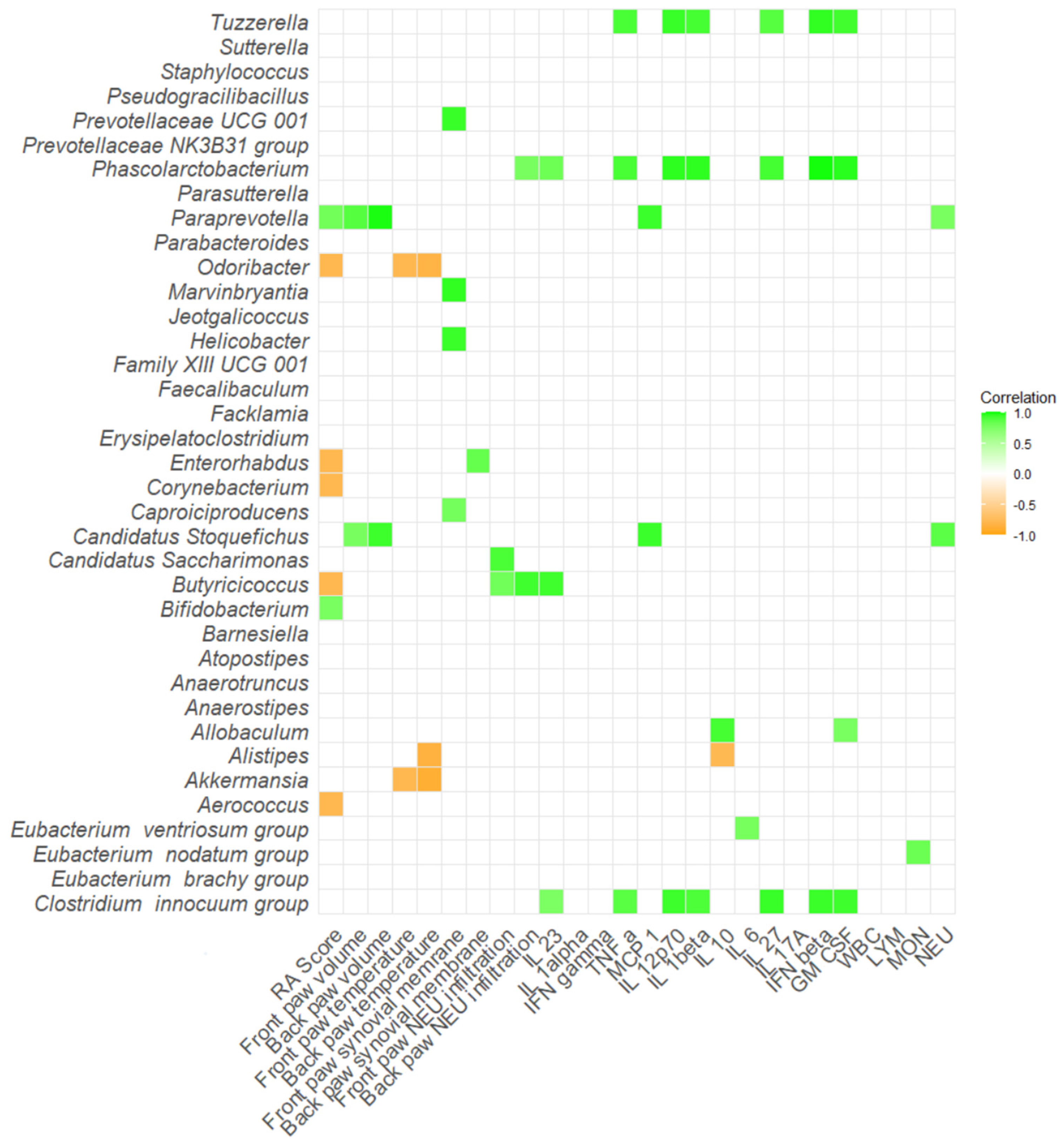
2.4. Correlation Severity of Arthritis and Gut Microbiota
3. Discussion
4. Materials and Methods
4.1. Donor Sample Collection
4.2. Recipient Housing Conditions
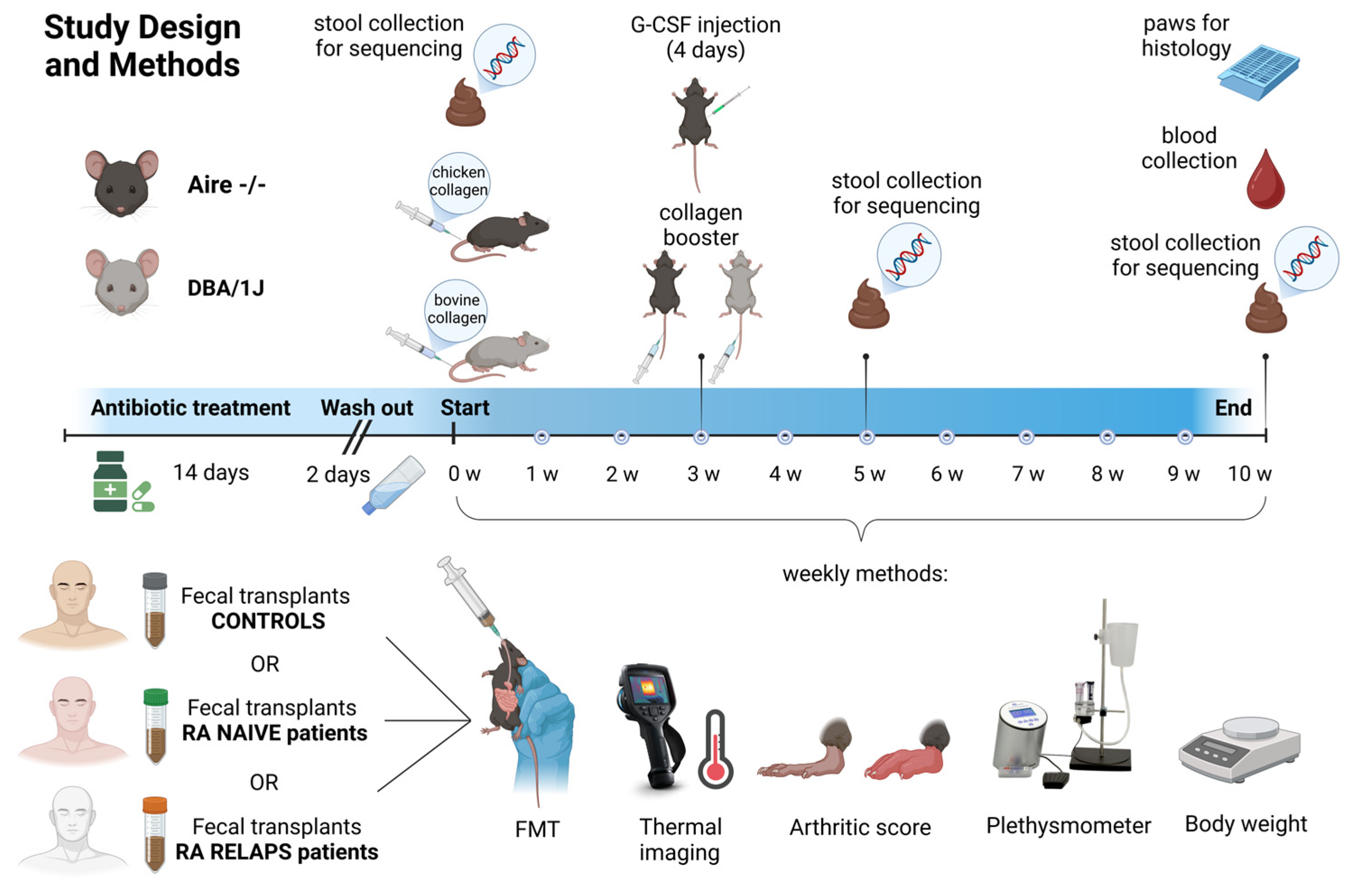
4.3. Experimental Groups and Study Design
4.4. Fecal Filtrate Preparation and Fecal Microbiota Transplantation
4.5. Recipient Sample Collection
4.6. Library Preparation and Sequencing
4.7. Bioinformatic Analysis
4.8. Severity of Arthritis and Paw Swelling
4.9. Paw Temperature
4.10. Histology
4.11. Inflammatory Cytokines Concentration
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid Arthritis. Nat. Rev. Dis. Primers 2018, 4, 18001. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Cytokines in the Pathogenesis of Rheumatoid Arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Romão, V.C.; Fonseca, J.E. Etiology and Risk Factors for Rheumatoid Arthritis: A State-of-the-Art Review. Front. Med. 2021, 8, 689698. [Google Scholar] [CrossRef] [PubMed]
- Esberg, A.; Johansson, L.; Johansson, I.; Dahlqvist, S.R. Oral Microbiota Identifies Patients in Early Onset Rheumatoid Arthritis. Microorganisms 2021, 9, 1657. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Mannaa, M.; Kim, Y.; Kim, J.; Kim, G.-T.; Seo, Y.-S. Comparative Analysis of Fecal Microbiota Composition Between Rheumatoid Arthritis and Osteoarthritis Patients. Genes 2019, 10, 748. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The Oral and Gut Microbiomes Are Perturbed in Rheumatoid Arthritis and Partly Normalized after Treatment. Nat. Med. 2015, 21, 895–905. [Google Scholar] [CrossRef]
- Bingham, C.O.; Moni, M. Periodontal Disease and Rheumatoid Arthritis: The Evidence Accumulates for Complex Pathobiologic Interactions. Curr. Opin. Rheumatol. 2013, 25, 345–353. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Romero-Figueroa, M.d.S.; Ramírez-Durán, N.; Montiel-Jarquín, A.J.; Horta-Baas, G. Gut-Joint Axis: Gut Dysbiosis Can Contribute to the Onset of Rheumatoid Arthritis via Multiple Pathways. Front. Cell. Infect. Microbiol. 2023, 13, 1092118. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Hooper, L.V.; Macpherson, A.J. Immune Adaptations That Maintain Homeostasis with the Intestinal Microbiota. Nat. Rev. Immunol. 2010, 10, 159–169. [Google Scholar] [CrossRef] [PubMed]
- van der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-Chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef]
- Bodkhe, R.; Balakrishnan, B.; Taneja, V. The Role of Microbiome in Rheumatoid Arthritis Treatment. Ther. Adv. Musculoskelet. Dis. 2019, 11, 1759720X19844632. [Google Scholar] [CrossRef]
- Zhao, T.; Wei, Y.; Zhu, Y.; Xie, Z.; Hai, Q.; Li, Z.; Qin, D. Gut Microbiota and Rheumatoid Arthritis: From Pathogenesis to Novel Therapeutic Opportunities. Front. Immunol. 2022, 13, 1007165. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An Expansion of Rare Lineage Intestinal Microbes Characterizes Rheumatoid Arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef]
- Chiang, H.-I.; Li, J.-R.; Liu, C.-C.; Liu, P.-Y.; Chen, H.-H.; Chen, Y.-M.; Lan, J.-L.; Chen, D.-Y. An Association of Gut Microbiota with Different Phenotypes in Chinese Patients with Rheumatoid Arthritis. J. Clin. Med. 2019, 8, 1770. [Google Scholar] [CrossRef]
- Drago, L. Prevotella Copri and Microbiota in Rheumatoid Arthritis: Fully Convincing Evidence? J. Clin. Med. 2019, 8, 1837. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.; Kandalgaonkar, M.R.; Golonka, R.M.; Yeoh, B.S.; Vijay-Kumar, M.; Saha, P. Crosstalk between Gut Microbiota and Host Immunity: Impact on Inflammation and Immunotherapy. Biomedicines 2023, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Tong, Y.; Wu, L.; Niu, H.; Li, Y.; Su, L.C.; Wu, Y.; Bozec, A.; Zaiss, M.M.; Qing, P.; et al. Alteration of Gut Microbiota in Individuals at High-Risk for Rheumatoid Arthritis Associated With Disturbed Metabolome and the Initiation of Arthritis Through the Triggering of Mucosal Immunity Imbalance. Arthritis Rheumatol. 2023, 75, 1736–1748. [Google Scholar] [CrossRef]
- Liptak, R.; Gromova, B.; Maronek, M.; Gardlik, R. Reverse Phenotype Transfer via Fecal Microbial Transplantation in Inflammatory Bowel Disease. Med. Hypotheses 2019, 122, 41–44. [Google Scholar] [CrossRef]
- Hvas, C.L.; Dahl Jørgensen, S.M.; Jørgensen, S.P.; Storgaard, M.; Lemming, L.; Hansen, M.M.; Erikstrup, C.; Dahlerup, J.F. Fecal Microbiota Transplantation Is Superior to Fidaxomicin for Treatment of Recurrent Clostridium Difficile Infection. Gastroenterology 2019, 156, 1324–1332.e3. [Google Scholar] [CrossRef] [PubMed]
- Belvoncikova, P.; Maronek, M.; Gardlik, R. Gut Dysbiosis and Fecal Microbiota Transplantation in Autoimmune Diseases. Int. J. Mol. Sci. 2022, 23, 10729. [Google Scholar] [CrossRef]
- Zeng, J.; Peng, L.; Zheng, W.; Huang, F.; Zhang, N.; Wu, D.; Yang, Y. Fecal Microbiota Transplantation for Rheumatoid Arthritis: A Case Report. Clin. Case Rep. 2021, 9, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-Induced Arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef]
- Schuh, B.M.; Macáková, K.; Feješ, A.; Groß, T.; Belvončíková, P.; Janko, J.; Juskanič, D.; Hollý, S.; Borbélyová, V.; Šteňová, E.; et al. Sex Differences in Long-Term Effects of Collagen-Induced Arthritis in Middle-Aged Mice. Front. Physiol. 2023, 14, 1195604. [Google Scholar] [CrossRef] [PubMed]
- He, M.; van Wijk, E.; Berger, R.; Wang, M.; Strassburg, K.; Schoeman, J.C.; Vreeken, R.J.; van Wietmarschen, H.; Harms, A.C.; Kobayashi, M.; et al. Collagen Induced Arthritis in DBA/1J Mice Associates with Oxylipin Changes in Plasma. Mediat. Inflamm. 2015, 2015, 543541. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Qin, L.; Cheng, W.; Lai, Y.; Yue, Y.; Ren, P.; Pan, X.; Zhang, P. Study in Treatment of Collagen-Induced Arthritis in DBA/1 Mice Model by Genistein. Curr. Pharm. Des. 2016, 22, 6975–6981. [Google Scholar] [CrossRef]
- Anderson, M.S.; Venanzi, E.S.; Klein, L.; Chen, Z.; Berzins, S.P.; Turley, S.J.; von Boehmer, H.; Bronson, R.; Dierich, A.; Benoist, C.; et al. Projection of an Immunological Self Shadow within the Thymus by the Aire Protein. Science 2002, 298, 1395–1401. [Google Scholar] [CrossRef]
- Miyoshi, M.; Liu, S. Collagen-Induced Arthritis Models. Methods Mol. Biol. 2024, 2766, 3–7. [Google Scholar] [CrossRef]
- Chen, F.Y.; Lee, A.; Ge, S.; Nathan, S.; Knox, S.M.; McNamara, N.A. Aire-Deficient Mice Provide a Model of Corneal and Lacrimal Gland Neuropathy in Sjögren’s Syndrome. PLoS ONE 2017, 12, e0184916. [Google Scholar] [CrossRef]
- Kisand, K.; Peterson, P.; Laan, M. Lymphopenia-Induced Proliferation in Aire-Deficient Mice Helps to Explain Their Autoimmunity and Differences from Human Patients. Front. Immunol. 2014, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qian, H.; Horai, R.; Chan, C.-C.; Caspi, R.R. Mouse Models of Experimental Autoimmune Uveitis: Comparative Analysis of Adjuvant-Induced vs Spontaneous Models of Uveitis. Curr. Mol. Med. 2015, 15, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Hardtke-Wolenski, M.; Taubert, R.; Noyan, F.; Sievers, M.; Dywicki, J.; Schlue, J.; Falk, C.S.; Ardesjö Lundgren, B.; Scott, H.S.; Pich, A.; et al. Autoimmune Hepatitis in a Murine Autoimmune Polyendocrine Syndrome Type 1 Model Is Directed against Multiple Autoantigens. Hepatology 2015, 61, 1295–1305. [Google Scholar] [CrossRef]
- García-Lozano, J.-R.; Torres-Agrela, B.; Montes-Cano, M.-A.; Ortiz-Fernández, L.; Conde-Jaldón, M.; Teruel, M.; García, A.; Núñez-Roldán, A.; Martín, J.; González-Escribano, M.-F. Association of the AIRE Gene with Susceptibility to Rheumatoid Arthritis in a European Population: A Case Control Study. Arthritis Res. Ther. 2013, 15, R11. [Google Scholar] [CrossRef]
- Terao, C.; Yamada, R.; Ohmura, K.; Takahashi, M.; Kawaguchi, T.; Kochi, Y.; Human Disease Genomics Working Group; RA Clinical and Genetic Study Consortium; Okada, Y.; Nakamura, Y.; et al. The Human AIRE Gene at Chromosome 21q22 Is a Genetic Determinant for the Predisposition to Rheumatoid Arthritis in Japanese Population. Hum. Mol. Genet. 2011, 20, 2680–2685. [Google Scholar] [CrossRef]
- Huo, F.-F.; Zou, X.-Y.; Zhang, Y.; Lu, Y.-P.; Zhao, M.-W.; Yu, X.-Y.; Cao, F.-G.; Yang, W. Aire Attenuate Collagen-Induced Arthritis by Suppressing T Follicular Helper Cells through ICOSL. Int. Immunopharmacol. 2025, 144, 113732. [Google Scholar] [CrossRef]
- Redmond, P.N.E. Protocol for the Successful Induction of Collagen-Induced Arthritis (CIA) and Collagen Antibody-Induced Arthritis (CAIA) in Mice. Chondrex INC 2017, 98052, 1–6. [Google Scholar]
- Fukui, S.; Gutch, S.; Fukui, S.; Cherpokova, D.; Aymonnier, K.; Sheehy, C.E.; Chu, L.; Wagner, D.D. The Prominent Role of Hematopoietic Peptidyl Arginine Deiminase 4 in Arthritis: Collagen- and Granulocyte Colony-Stimulating Factor-Induced Arthritis Model in C57BL/6 Mice. Arthritis Rheumatol. 2022, 74, 1139–1146. [Google Scholar] [CrossRef]
- Wang, H.; Cai, Y.; Wu, W.; Zhang, M.; Dai, Y.; Wang, Q. Exploring the Role of Gut Microbiome in Autoimmune Diseases: A Comprehensive Review. Autoimmun. Rev. 2024, 23, 103654. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, Q.; Lin, P.; Xu, R.; He, D.; Ji, W.; Bian, Y.; Shen, Y.; Li, Q.; Liu, C.; et al. Characteristics of Gut Microbiota in Patients With Rheumatoid Arthritis in Shanghai, China. Front. Cell. Infect. Microbiol. 2019, 9, 369. [Google Scholar] [CrossRef]
- Rooney, C.M.; Mankia, K.; Mitra, S.; Moura, I.B.; Emery, P.; Wilcox, M.H. Perturbations of the Gut Microbiome in Anti-CCP Positive Individuals at Risk of Developing Rheumatoid Arthritis. Rheumatology 2021, 60, 3380–3387. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chen, Z.; Cai, J. Fecal Microbiota Transplantation: Emerging Applications in Autoimmune Diseases. J. Autoimmun. 2023, 141, 103038. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, T.; Marole, T.A.; Thomashoff, U.L.; Thantsha, M.S.; Buys, E.M. Bifidobacterium Species Viability in Dairy-Based Probiotic Foods: Challenges and Innovative Approaches for Accurate Viability Determination and Monitoring of Probiotic Functionality. Front. Microbiol. 2024, 15, 1327010. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, J.; Xiao, L.; Lu, A.; Zhang, G. Alterations of Gut Microbiome in Rheumatoid Arthritis. Osteoarthr. Cartil. 2017, 25, S287–S288. [Google Scholar] [CrossRef]
- Marietta, E.V.; Murray, J.A.; Luckey, D.H.; Jeraldo, P.R.; Lamba, A.; Patel, R.; Luthra, H.S.; Mangalam, A.; Taneja, V. Suppression of Inflammatory Arthritis by Human Gut-Derived Prevotella Histicola in Humanized Mice. Arthritis Rheumatol. 2016, 68, 2878–2888. [Google Scholar] [CrossRef]
- Koch, A.E.; Kunkel, S.L.; Harlow, L.A.; Johnson, B.; Evanoff, H.L.; Haines, G.K.; Burdick, M.D.; Pope, R.M.; Strieter, R.M. Enhanced Production of Monocyte Chemoattractant Protein-1 in Rheumatoid Arthritis. J. Clin. Investig. 1992, 90, 772–779. [Google Scholar] [CrossRef]
- Dürholz, K.; Hofmann, J.; Iljazovic, A.; Häger, J.; Lucas, S.; Sarter, K.; Strowig, T.; Bang, H.; Rech, J.; Schett, G.; et al. Dietary Short-Term Fiber Interventions in Arthritis Patients Increase Systemic SCFA Levels and Regulate Inflammation. Nutrients 2020, 12, 3207. [Google Scholar] [CrossRef]
- Ichimura, A.; Hasegawa, S.; Kasubuchi, M.; Kimura, I. Free Fatty Acid Receptors as Therapeutic Targets for the Treatment of Diabetes. Front. Pharmacol. 2014, 5, 236. [Google Scholar] [CrossRef]
- Fan, Z.; Yang, B.; Ross, R.P.; Stanton, C.; Shi, G.; Zhao, J.; Zhang, H.; Chen, W. Protective Effects of Bifidobacterium Adolescentis on Collagen-Induced Arthritis in Rats Depend on Timing of Administration. Food Funct. 2020, 11, 4499–4511. [Google Scholar] [CrossRef]
- Jeong, Y.; Jhun, J.; Lee, S.-Y.; Na, H.S.; Choi, J.; Cho, K.-H.; Lee, S.Y.; Lee, A.R.; Park, S.-J.; You, H.J.; et al. Therapeutic Potential of a Novel Bifidobacterium Identified Through Microbiome Profiling of RA Patients With Different RF Levels. Front. Immunol. 2021, 12, 736196. [Google Scholar] [CrossRef]
- Yang, Y.; Hong, Q.; Zhang, X.; Liu, Z. Rheumatoid Arthritis and the Intestinal Microbiome: Probiotics as a Potential Therapy. Front. Immunol. 2024, 15, 1331486. [Google Scholar] [CrossRef]
- Liu, X.; Zeng, B.; Zhang, J.; Li, W.; Mou, F.; Wang, H.; Zou, Q.; Zhong, B.; Wu, L.; Wei, H.; et al. Role of the Gut Microbiome in Modulating Arthritis Progression in Mice. Sci. Rep. 2016, 6, 30594. [Google Scholar] [CrossRef]
- Kondo, N.; Kuroda, T.; Kobayashi, D. Cytokine Networks in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 10922. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takahashi, N.; Kato, T.; Matsuda, Y.; Yokoji, M.; Yamada, M.; Nakajima, T.; Kondo, N.; Endo, N.; Yamamoto, R.; et al. Aggravation of Collagen-Induced Arthritis by Orally Administered Porphyromonas Gingivalis through Modulation of the Gut Microbiota and Gut Immune System. Sci. Rep. 2017, 7, 6955. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yin, M.; Cao, D.; Zhang, X.; Zhu, Y.; Wei, Y.; Li, Y.; Wen, C.; Zhou, J. Disruption of Gut Microbiota and Associated Fecal Metabolites in Collagen-Induced Arthritis Mice During the Early Stage. J. Inflamm. Res. 2025, 18, 1703–1717. [Google Scholar] [CrossRef]
- Hidalgo-Villeda, F.; Million, M.; Defoort, C.; Vannier, T.; Svilar, L.; Lagier, M.; Wagner, C.; Arroyo-Portilla, C.; Chasson, L.; Luciani, C.; et al. Prolonged Dysbiosis and Altered Immunity under Nutritional Intervention in a Physiological Mouse Model of Severe Acute Malnutrition. iScience 2023, 26, 106910. [Google Scholar] [CrossRef]
- Gomez, A.; Luckey, D.; Yeoman, C.J.; Marietta, E.V.; Miller, M.E.B.; Murray, J.A.; White, B.A.; Taneja, V. Loss of Sex and Age Driven Differences in the Gut Microbiome Characterize Arthritis-Susceptible *0401 Mice but Not Arthritis-Resistant *0402 Mice. PLoS ONE 2012, 7, e36095. [Google Scholar] [CrossRef]
- Zhuang, J.; Zhuang, Z.; Chen, B.; Yang, Y.; Chen, H.; Guan, G. Odoribacter Splanchnicus-Derived Extracellular Vesicles Alleviate Inflammatory Bowel Disease by Modulating Gastrointestinal Inflammation and Intestinal Barrier Function via the NLRP3 Inflammasome Suppression. Mol. Med. 2025, 31, 56. [Google Scholar] [CrossRef] [PubMed]
- Bakinowska, E.; Stańska, W.; Kiełbowski, K.; Szwedkowicz, A.; Boboryko, D.; Pawlik, A. Gut Dysbiosis and Dietary Interventions in Rheumatoid Arthritis—A Narrative Review. Nutrients 2024, 16, 3215. [Google Scholar] [CrossRef]
- Lan, W.; Lu, Q.; Ma, W.; Jiang, Z.; Chen, Y.; Wang, Z.; Yao, X.; Tang, F. Investigating the Causal Relationship between the Gut Microbiome and Rheumatoid Arthritis: Mediating Effects of Immune Cells. J. Transl. Med. 2025, 23, 187. [Google Scholar] [CrossRef]
- Jouhault, Q.; Cherqaoui, B.; Jobart-Malfait, A.; Glatigny, S.; Lauraine, M.; Hulot, A.; Morelle, G.; Hagege, B.; Ermoza, K.; El Marjou, A.; et al. Interleukin 27 Is a Novel Cytokine with Anti-Inflammatory Effects against Spondyloarthritis through the Suppression of Th17 Responses. Front. Immunol. 2022, 13, 1072420. [Google Scholar] [CrossRef]
- Duncan, S.H.; Conti, E.; Ricci, L.; Walker, A.W. Links between Diet, Intestinal Anaerobes, Microbial Metabolites and Health. Biomedicines 2023, 11, 1338. [Google Scholar] [CrossRef]
- Laperrousaz, B.; Levast, B.; Fontaine, M.; Nancey, S.; Dechelotte, P.; Doré, J.; Lehert, P. Safety Comparison of Single-Donor and Pooled Fecal Microbiota Transfer Product Preparation in Ulcerative Colitis: Systematic Review and Meta-Analysis. BMC Gastroenterol. 2024, 24, 402. [Google Scholar] [CrossRef] [PubMed]
- Levast, B.; Fontaine, M.; Nancey, S.; Dechelotte, P.; Doré, J.; Lehert, P. Single-Donor and Pooling Strategies for Fecal Microbiota Transfer Product Preparation in Ulcerative Colitis: A Systematic Review and Meta-Analysis. Clin. Transl. Gastroenterol. 2023, 14, e00568. [Google Scholar] [CrossRef]
- Reygner, J.; Delannoy, J.; Barba-Goudiaby, M.-T.; Gasc, C.; Levast, B.; Gaschet, E.; Ferraris, L.; Paul, S.; Kapel, N.; Waligora-Dupriet, A.-J.; et al. Reduction of Product Composition Variability Using Pooled Microbiome Ecosystem Therapy and Consequence in Two Infectious Murine Models. Appl. Environ. Microbiol. 2024, 90, e00016-24. [Google Scholar] [CrossRef] [PubMed]
- Sidiropoulos, D.N.; Al-Ghalith, G.A.; Shields-Cutler, R.R.; Ward, T.L.; Johnson, A.J.; Vangay, P.; Knights, D.; Kashyap, P.C.; Xian, Y.; Ramer-Tait, A.E.; et al. Wild Primate Microbiomes Prevent Weight Gain in Germ-Free Mice. Anim. Microbiome 2020, 2, 16. [Google Scholar] [CrossRef]
- Kay, J.; Upchurch, K.S. ACR/EULAR 2010 Rheumatoid Arthritis Classification Criteria. Rheumatology 2012, 51, vi5–vi9. [Google Scholar] [CrossRef] [PubMed]
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 March 2025).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.S. Using DECIPHER v2.0 to Analyze Big Biological Sequence Data in R. R J. 2016, 8, 352. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RA-NAIVE | RA-RELAPS | p-Value | |
|---|---|---|---|
| Age | 55.67 ± 8.45 | 59.25 ± 7.59 | ns |
| Duration of RA | 0.75 ± 0.75 | 4.67 ± 0.17 | <0.01 |
| DAS28 | 4.83 ± 0.78 | 4.99 ± 0.44 | ns |
| Anti-CCP [U/mL] | 21.80 ± 18.17 | 121.70 ± 49.17 | ns |
| RF [IU/mL] | 11.00 ± 1.00 | 15.00 ± 2.08 | ns |
| CRP [mg/L] | 29.95 ± 27.03 | 25.63 ± 22.24 | ns |
| ESR [mm/h.] | 34.50 ± 24.10 | 55.67 ± 15.51 | ns |
| LYM [x 10⁹/l] | 1.840 ± 0.20 | 1.62 ± 0.39 | ns |
| LEU [× 10⁹/l] | 7.62 ± 0.91 | 5.56 ± 0.97 | ns |
| NEU [× 10⁹/l] | 5.17 ± 0.89 | 3.31 ± 0.92 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belvončíková, P.; Macáková, K.; Tóthová, N.; Babál, P.; Tarabčáková, L.; Gardlík, R. Investigating the Role of Gut Microbiota in the Pathogenesis and Progression of Rheumatoid Arthritis in a Collagen-Induced Arthritis Mouse Model. Int. J. Mol. Sci. 2025, 26, 5099. https://doi.org/10.3390/ijms26115099
Belvončíková P, Macáková K, Tóthová N, Babál P, Tarabčáková L, Gardlík R. Investigating the Role of Gut Microbiota in the Pathogenesis and Progression of Rheumatoid Arthritis in a Collagen-Induced Arthritis Mouse Model. International Journal of Molecular Sciences. 2025; 26(11):5099. https://doi.org/10.3390/ijms26115099
Chicago/Turabian StyleBelvončíková, Paulína, Kristína Macáková, Nikola Tóthová, Pavel Babál, Lenka Tarabčáková, and Roman Gardlík. 2025. "Investigating the Role of Gut Microbiota in the Pathogenesis and Progression of Rheumatoid Arthritis in a Collagen-Induced Arthritis Mouse Model" International Journal of Molecular Sciences 26, no. 11: 5099. https://doi.org/10.3390/ijms26115099
APA StyleBelvončíková, P., Macáková, K., Tóthová, N., Babál, P., Tarabčáková, L., & Gardlík, R. (2025). Investigating the Role of Gut Microbiota in the Pathogenesis and Progression of Rheumatoid Arthritis in a Collagen-Induced Arthritis Mouse Model. International Journal of Molecular Sciences, 26(11), 5099. https://doi.org/10.3390/ijms26115099