
Exploring the Multi-Faceted Effects of Berberine in Ameliorating Diastolic Dysfunction in Rats with Heart Failure with Preserved Ejection Fraction

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. BBR Ameliorates Cardiac Diastolic Dysfunction, Hypertrophy, and Extracardiac Comorbidities in HFHS Diet-Fed Rats
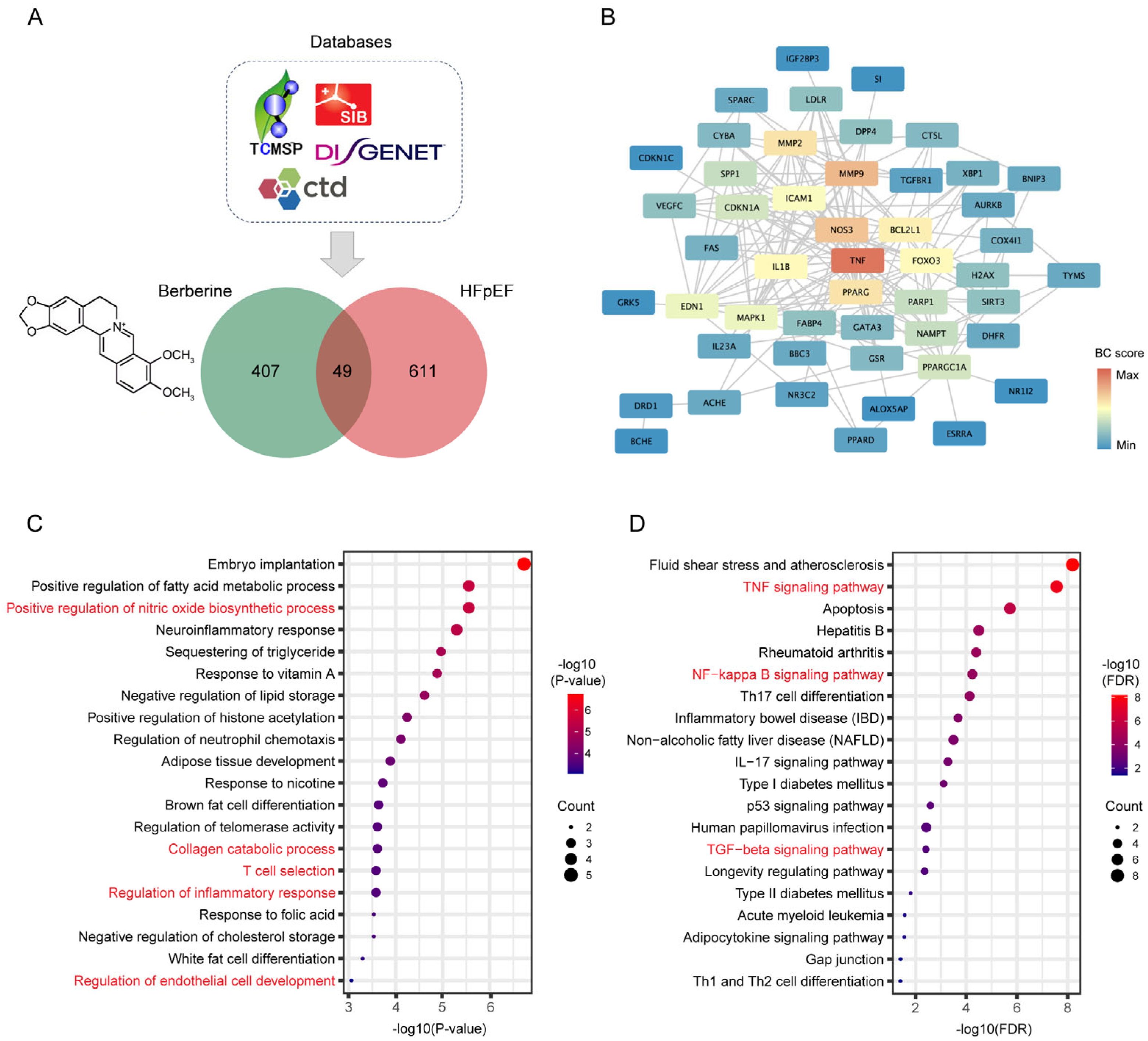
2.2. Network Pharmacology Predicates the Potential Mechanisms of BBR in the Treatment of HFpEF
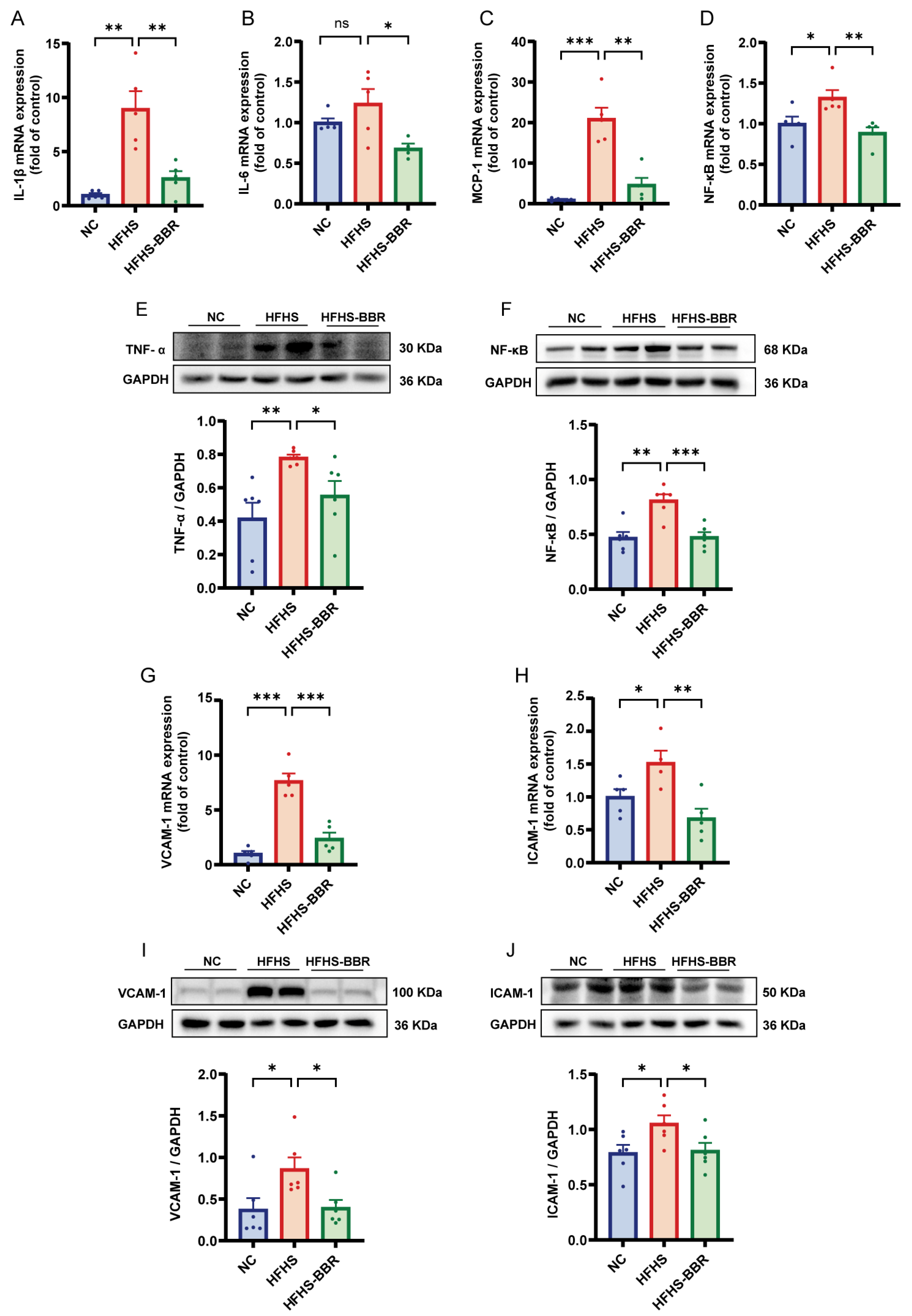
2.3. BBR Alleviates Myocardial Inflammation and Microvascular Inflammation
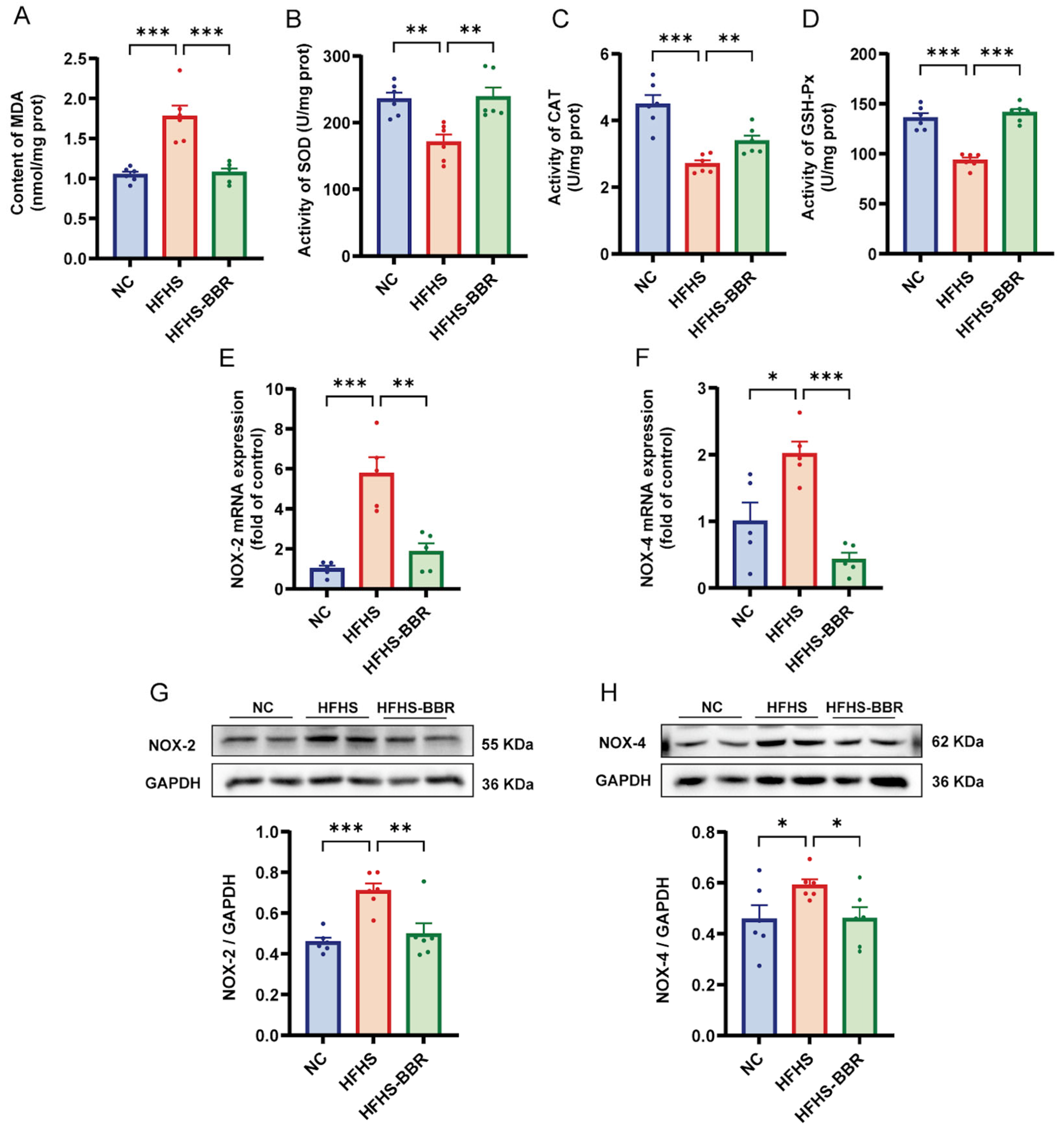
2.4. BBR Suppresses Myocardial Oxidative Stress
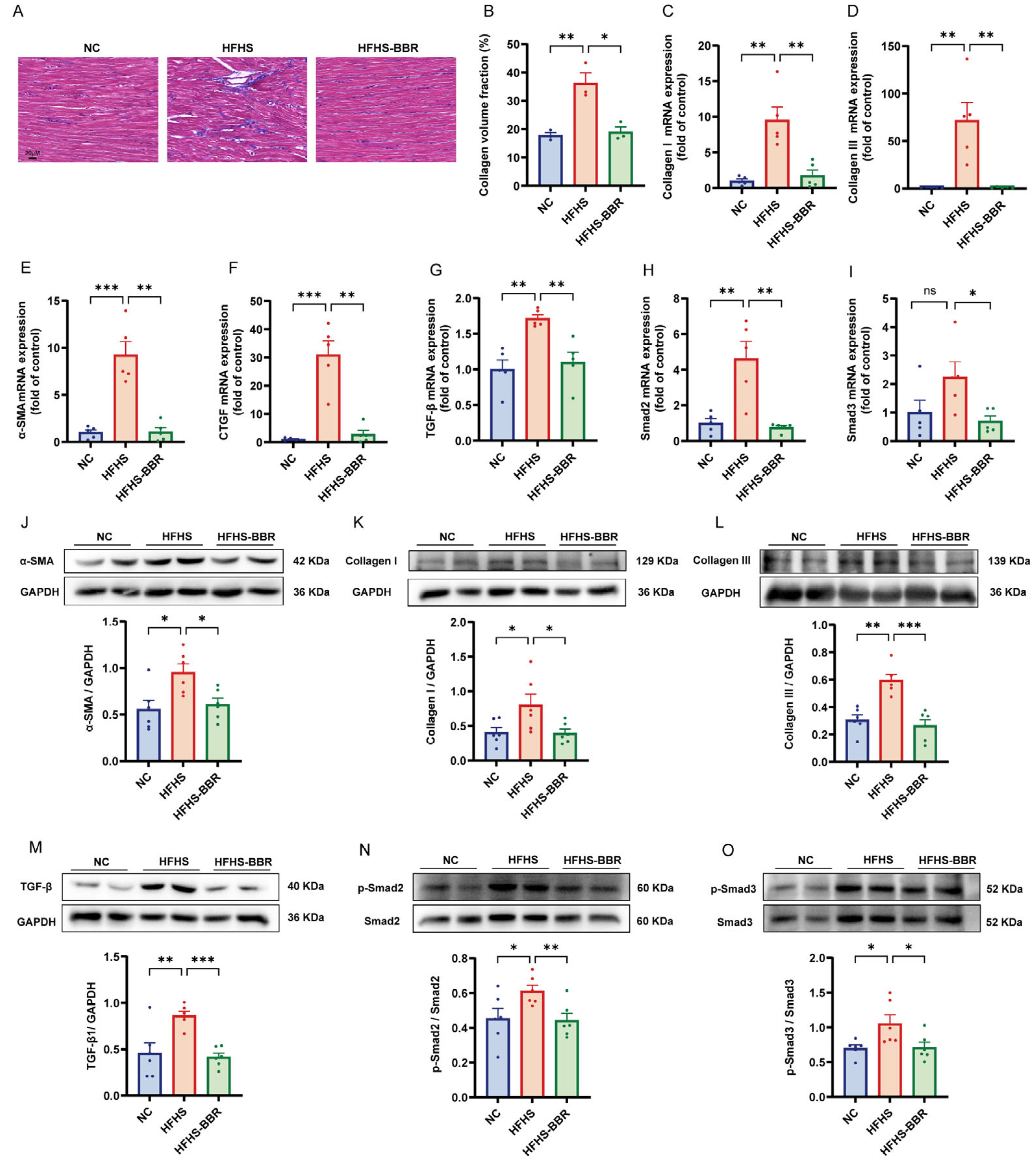
2.5. BBR Diminishes Myocardial Fibrosis and Inhibits TGF-β/Smads Pathway
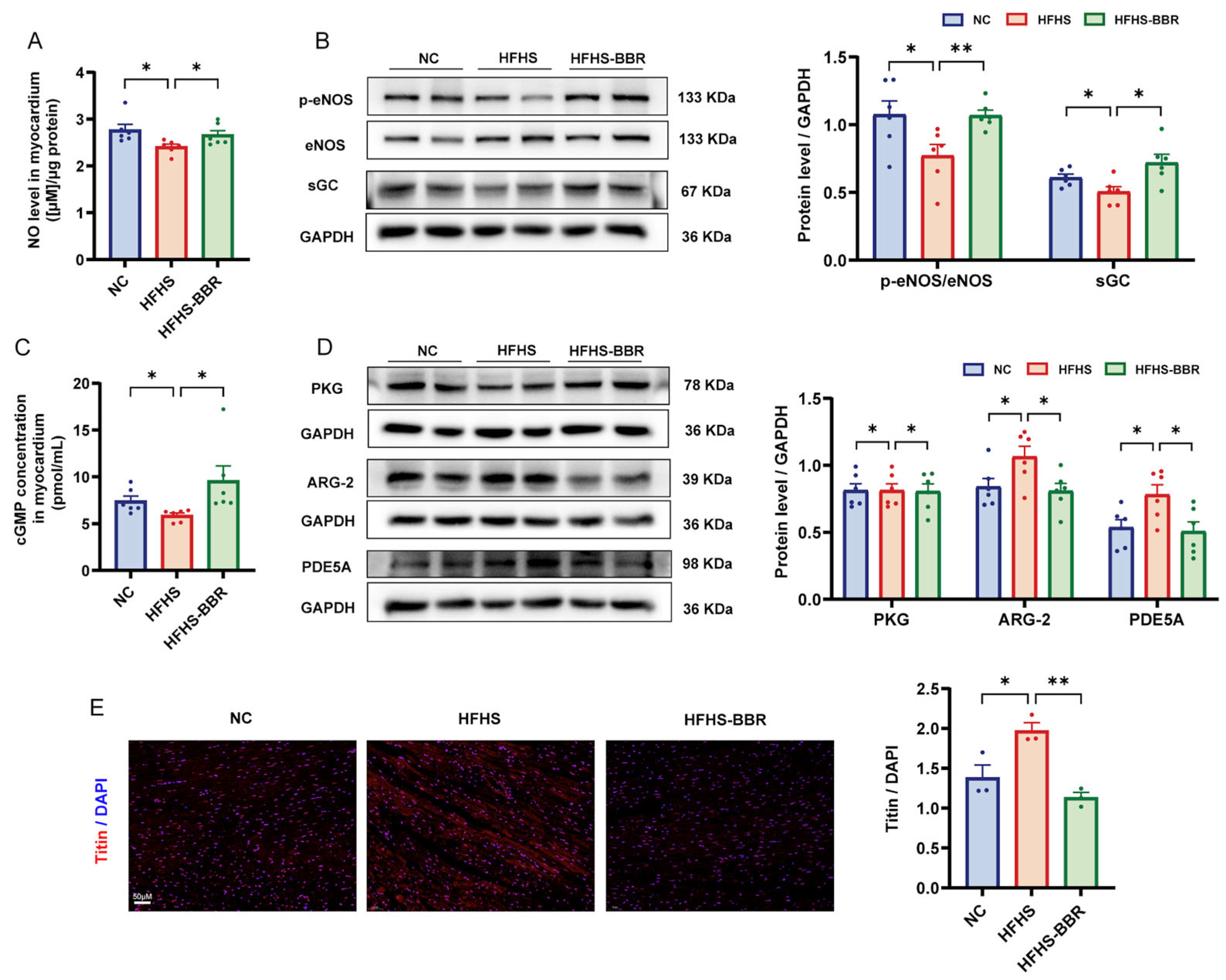
2.6. BBR Restores the Impaired NO/cGMP/PKG Pathway and Reduces Cardiomyocytes Stiffness
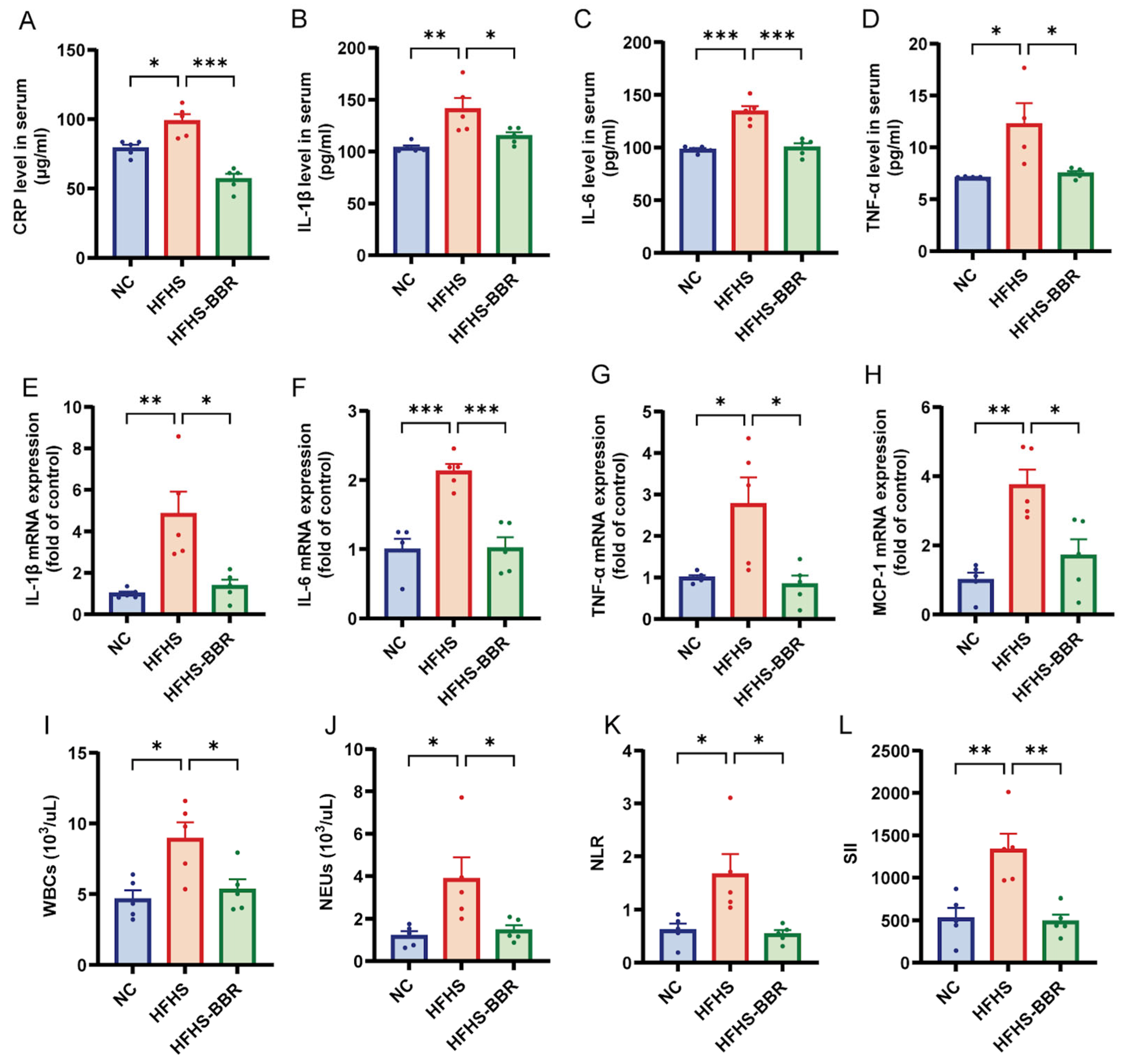
2.7. BBR Mitigates Systemic Inflammation in HFHS Diet-Fed Rats
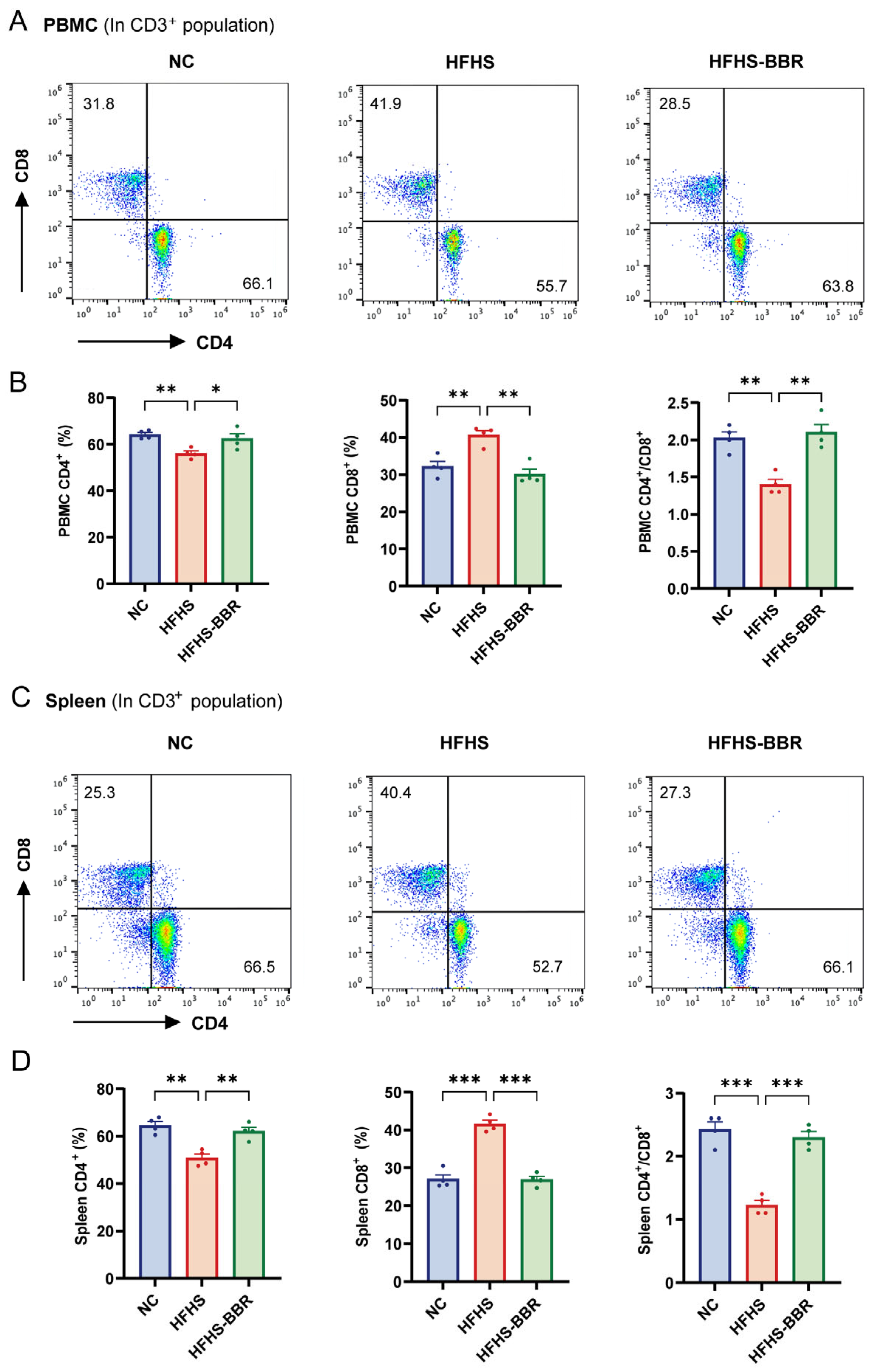
2.8. BBR Correctes Immune Dysregulation in HFHS Diet-Fed Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Echocardiographic Analysis
4.3. Oral Glucose Tolerance Test (OGTT)
4.4. Blood Pressure Measurements
4.5. Network Pharmacology
4.6. Biochemical Analyses
4.7. Hemogram Analysis
4.8. Histopathological Analysis
4.9. Immunofluorescence Analysis
4.10. RNA Isolation and qRT-PCR
4.11. Western Blotting
4.12. Flow Cytometry
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borlaug, B.A.; Sharma, K.; Shah, S.J.; Ho, J.E. Heart Failure with Preserved Ejection Fraction: JACC Scientific Statement. J. Am. Coll. Cardiol. 2023, 81, 1810–1834. [Google Scholar] [CrossRef] [PubMed]
- Wintrich, J.; Kindermann, I.; Ukena, C.; Selejan, S.; Werner, C.; Maack, C.; Laufs, U.; Tschöpe, C.; Anker, S.D.; Lam, C.S.P.; et al. Therapeutic Approaches in Heart Failure with Preserved Ejection Fraction: Past, Present, and Future. Clin. Res. Cardiol. 2020, 109, 1079–1098. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.J.; Borlaug, B.A.; Kitzman, D.W.; McCulloch, A.D.; Blaxall, B.C.; Agarwal, R.; Chirinos, J.A.; Collins, S.; Deo, R.C.; Gladwin, M.T.; et al. Research Priorities for Heart Failure with Preserved Ejection Fraction: National Heart, Lung, and Blood Institute Working Group Summary. Circulation 2020, 141, 1001–1026. [Google Scholar] [CrossRef]
- Dunlay, S.M.; Roger, V.L.; Redfield, M.M. Epidemiology of Heart Failure with Preserved Ejection Fraction. Nat. Rev. Cardiol. 2017, 14, 591–602. [Google Scholar] [CrossRef]
- Solomon, S.D.; McMurray, J.J.V.; Claggett, B.; de Boer, R.A.; DeMets, D.; Hernandez, A.F.; Inzucchi, S.E.; Kosiborod, M.N.; Lam, C.S.P.; Martinez, F.; et al. Dapagliflozin in Heart Failure with Mildly Reduced or Preserved Ejection Fraction. N. Engl. J. Med. 2022, 387, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Borlaug, B.A.; Kitzman, D.W.; Davies, M.J.; Rasmussen, S.; Barros, E.; Butler, J.; Einfeldt, M.N.; Hovingh, G.K.; Møller, D.V.; Petrie, M.C.; et al. Semaglutide in HFpEF across Obesity Class and by Body Weight Reduction: A Prespecified Analysis of the STEP-HFpEF Trial. Nat. Med. 2023, 29, 2358–2365. [Google Scholar] [CrossRef]
- Gallo, G.; Volpe, M. Potential Mechanisms of the Protective Effects of the Cardiometabolic Drugs Type-2 Sodium-Glucose Transporter Inhibitors and Glucagon-like Peptide-1 Receptor Agonists in Heart Failure. Int. J. Mol. Sci. 2024, 25, 2484. [Google Scholar] [CrossRef] [PubMed]
- Belli, M.; Barone, L.; Bellia, A.; Sergi, D.; Lecis, D.; Prandi, F.R.; Milite, M.; Galluccio, C.; Muscoli, S.; Romeo, F.; et al. Treatment of HFpEF beyond the SGLT2-Is: Does the Addition of GLP-1 RA Improve Cardiometabolic Risk and Outcomes in Diabetic Patients? Int. J. Mol. Sci. 2022, 23, 14598. [Google Scholar] [CrossRef]
- Kolijn, D.; Pabel, S.; Tian, Y.; Lódi, M.; Herwig, M.; Carrizzo, A.; Zhazykbayeva, S.; Kovács, Á.; Fülöp, G.Á.; Falcão-Pires, I.; et al. Empagliflozin Improves Endothelial and Cardiomyocyte Function in Human Heart Failure with Preserved Ejection Fraction via Reduced Pro-Inflammatory-Oxidative Pathways and Protein Kinase Gα Oxidation. Cardiovasc. Res. 2021, 117, 495–507. [Google Scholar] [CrossRef]
- Bonfioli, G.B.; Rodella, L.; Metra, M.; Vizzardi, E. GLP-1 Receptor Agonists as Promising Anti-Inflammatory Agents in Heart Failure with Preserved Ejection Fraction. Heart Fail. Rev. 2025, 30, 131–136. [Google Scholar] [CrossRef]
- Habtemariam, S. Berberine Pharmacology and the Gut Microbiota: A Hidden Therapeutic Link. Pharmacol. Res. 2020, 155, 104722. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.-J.; Vernieri, C.; Foiani, M.; Jiang, J.-D. Berberine in the Treatment of Metabolism-Related Chronic Diseases: A Drug Cloud (dCloud) Effect to Target Multifactorial Disorders. Pharmacol. Ther. 2020, 209, 107496. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Sureda, A.; Jafari, S.; Memariani, Z.; Tewari, D.; Barrea, L.; Hassan, S.T.S.; Šmejkal, K.; Malaník, M.; Sychrová, A.; et al. Berberine in Cardiovascular and Metabolic Diseases: From Mechanisms to Therapeutics. Theranostics 2019, 9, 1923–1951. [Google Scholar] [CrossRef]
- Zeng, X.-H.; Zeng, X.-J.; Li, Y.-Y. Efficacy and Safety of Berberine for Congestive Heart Failure Secondary to Ischemic or Idiopathic Dilated Cardiomyopathy. Am. J. Cardiol. 2003, 92, 173–176. [Google Scholar] [CrossRef]
- Wang, S.; Lu, K.; Lin, L.; Li, G.; Han, Y.; Lin, Z.; Chu, Q.; Wu, K.; Liu, P.; Zhou, G.; et al. Exploring the Mechanism of Berberine Treatment for Atherosclerosis Combined with Non-Alcoholic Fatty Liver Disease Based on Bioinformatic and Experimental Study. PLoS ONE 2024, 19, e0314961. [Google Scholar] [CrossRef]
- Abudureyimu, M.; Yang, M.; Wang, X.; Luo, X.; Ge, J.; Peng, H.; Zhang, Y.; Ren, J. Berberine Alleviates Myocardial Diastolic Dysfunction by Modulating Drp1-Mediated Mitochondrial Fission and Ca2+ Homeostasis in a Murine Model of HFpEF. Front. Med. 2023, 17, 1219–1235. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Yang, Y.; Jiang, S.; Liu, C.; Han, Y.; Jiang, J.; Wang, Y. Benefits of Puerarin on Metabolic Syndrome and Its Associated Cardiovascular Diseases in Rats Fed a High-Fat/High-Sucrose Diet. Nutrients 2024, 16, 1273. [Google Scholar] [CrossRef]
- Liu, H.; Huang, Y.; Zhao, Y.; Kang, G.-J.; Feng, F.; Wang, X.; Liu, M.; Shi, G.; Revelo, X.; Bernlohr, D.; et al. Inflammatory Macrophage Interleukin-1β Mediates High-Fat Diet-Induced Heart Failure with Preserved Ejection Fraction. JACC Basic Transl. Sci. 2023, 8, 174–185. [Google Scholar] [CrossRef]
- Li, C.; Jiang, S.; Wang, H.; Wang, Y.; Han, Y.; Jiang, J. Berberine Exerts Protective Effects on Cardiac Senescence by Regulating the Klotho/SIRT1 Signaling Pathway. Biomed. Pharmacother. 2022, 151, 113097. [Google Scholar] [CrossRef]
- Zhao, Y.; Shao, W.; Zhu, Q.; Zhang, R.; Sun, T.; Wang, B.; Hu, X. Association between Systemic Immune-Inflammation Index and Metabolic Syndrome and Its Components: Results from the National Health and Nutrition Examination Survey 2011–2016. J. Transl. Med. 2023, 21, 691. [Google Scholar] [CrossRef]
- James, A.S.; Eteng, O.E.; Dosumu, O.A.; Moses, C.A.; Ogbonna, C.U.; Adeleye, O.A.; Ugwor, E.I.; Omilo, B.C.; Fabunmi, R.F.; Olakitan, A.M.; et al. Morin Augmented Myocardial eNOS/cGMP/PKG Signaling Pathway and Abated Oxidative and Inflammo-Apoptotic Responses in Diethyl Phthalate and Bisphenol-S Co-Exposed Male Albino Rats. Inflammation 2023, 46, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Van Heerebeek, L.; Hamdani, N.; Falcão-Pires, I.; Leite-Moreira, A.F.; Begieneman, M.P.V.; Bronzwaer, J.G.F.; Van Der Velden, J.; Stienen, G.J.M.; Laarman, G.J.; Somsen, A.; et al. Low Myocardial Protein Kinase G Activity in Heart Failure With Preserved Ejection Fraction. Circulation 2012, 126, 830–839. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, K.; Liu, M.; Su, J.; Qin, X.; Wang, X.; Zhang, J.; Li, S.; Fan, G. An Herbal Preparation Ameliorates Heart Failure with Preserved Ejection Fraction by Alleviating Microvascular Endothelial Inflammation and Activating NO-cGMP-PKG Pathway. Phytomedicine 2021, 91, 153633. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory Links Between High Fat Diets and Diseases. Front. Immunol. 2018, 9, 2649. [Google Scholar] [CrossRef]
- Kiran, S.; Rakib, A.; Kodidela, S.; Kumar, S.; Singh, U.P. High-Fat Diet-Induced Dysregulation of Immune Cells Correlates with Macrophage Phenotypes and Chronic Inflammation in Adipose Tissue. Cells 2022, 11, 1327. [Google Scholar] [CrossRef]
- Paulus, W.J.; Tschöpe, C. A Novel Paradigm for Heart Failure with Preserved Ejection Fraction: Comorbidities Drive Myocardial Dysfunction and Remodeling through Coronary Microvascular Endothelial Inflammation. J. Am. Coll. Cardiol. 2013, 62, 263–271. [Google Scholar] [CrossRef]
- Jeong, E.-M.; Chung, J.; Liu, H.; Go, Y.; Gladstein, S.; Farzaneh-Far, A.; Lewandowski, E.D.; Dudley, S.C. Role of Mitochondrial Oxidative Stress in Glucose Tolerance, Insulin Resistance, and Cardiac Diastolic Dysfunction. J. Am. Heart Assoc. 2016, 5, e003046. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cheng, C.-F.; Li, Z.-F.; Huang, X.-J.; Cai, S.-Q.; Ye, S.-Y.; Zhao, L.-J.; Xiong, Y.; Chen, D.-F.; Liu, H.-L.; et al. Berberine Blocks Inflammasome Activation and Alleviates Diabetic Cardiomyopathy via the miR-18a-3p/Gsdmd Pathway. Int. J. Mol. Med. 2023, 51, 49. [Google Scholar] [CrossRef]
- Zhong, C.; Xie, Y.; Wang, H.; Chen, W.; Yang, Z.; Zhang, L.; Deng, Q.; Cheng, T.; Li, M.; Ju, J.; et al. Berberine Inhibits NLRP3 Inflammasome Activation by Regulating mTOR/mtROS Axis to Alleviate Diabetic Cardiomyopathy. Eur. J. Pharmacol. 2024, 964, 176253. [Google Scholar] [CrossRef]
- Wang, Y.; Liao, J.; Luo, Y.; Li, M.; Su, X.; Yu, B.; Teng, J.; Wang, H.; Lv, X. Berberine Alleviates Doxorubicin-Induced Myocardial Injury and Fibrosis by Eliminating Oxidative Stress and Mitochondrial Damage via Promoting Nrf-2 Pathway Activation. Int. J. Mol. Sci. 2023, 24, 3257. [Google Scholar] [CrossRef]
- Jia, X.; Shao, W.; Tian, S. Berberine Alleviates Myocardial Ischemia-Reperfusion Injury by Inhibiting Inflammatory Response and Oxidative Stress: The Key Function of miR-26b-5p-Mediated PTGS2/MAPK Signal Transduction. Pharm. Biol. 2022, 60, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Sun, G. Blocking RIPK2 Function Alleviates Myocardial Ischemia/Reperfusion Injury by Regulating the AKT and NF-κB Pathways. Immunol. Investig. 2023, 52, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qian, J.; Wang, L.; Li, J.; Zhao, Y.; Han, J.; Khan, Z.; Chen, X.; Wang, J.; Liang, G. Kaempferol Attenuates Hyperglycemia-Induced Cardiac Injuries by Inhibiting Inflammatory Responses and Oxidative Stress. Endocrine 2018, 60, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Teuber, J.P.; Essandoh, K.; Hummel, S.L.; Madamanchi, N.R.; Brody, M.J. NADPH Oxidases in Diastolic Dysfunction and Heart Failure with Preserved Ejection Fraction. Antioxidants 2022, 11, 1822. [Google Scholar] [CrossRef]
- Simmonds, S.J.; Cuijpers, I.; Heymans, S.; Jones, E.A.V. Cellular and Molecular Differences between HFpEF and HFrEF: A Step Ahead in an Improved Pathological Understanding. Cells 2020, 9, 242. [Google Scholar] [CrossRef]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative Stress and Reactive Oxygen Species in Endothelial Dysfunction Associated with Cardiovascular and Metabolic Diseases. Vasc. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef]
- Wu, M.; Guo, Y.; Wu, Y.; Xu, K.; Lin, L. Protective Effects of Sacubitril/Valsartan on Cardiac Fibrosis and Function in Rats with Experimental Myocardial Infarction Involves Inhibition of Collagen Synthesis by Myocardial Fibroblasts Through Downregulating TGF-Β1/Smads Pathway. Front. Pharmacol. 2021, 12, 696472. [Google Scholar] [CrossRef]
- Duca, F.; Kammerlander, A.A.; Zotter-Tufaro, C.; Aschauer, S.; Schwaiger, M.L.; Marzluf, B.A.; Bonderman, D.; Mascherbauer, J. Interstitial Fibrosis, Functional Status, and Outcomes in Heart Failure with Preserved Ejection Fraction: Insights From a Prospective Cardiac Magnetic Resonance Imaging Study. Circ. Cardiovasc. Imaging 2016, 9, e005277. [Google Scholar] [CrossRef]
- Franssen, C.; Chen, S.; Unger, A.; Korkmaz, H.I.; De Keulenaer, G.W.; Tschöpe, C.; Leite-Moreira, A.F.; Musters, R.; Niessen, H.W.M.; Linke, W.A.; et al. Myocardial Microvascular Inflammatory Endothelial Activation in Heart Failure with Preserved Ejection Fraction. JACC Heart Fail. 2016, 4, 312–324. [Google Scholar] [CrossRef]
- Cai, Z.; Wu, C.; Xu, Y.; Cai, J.; Zhao, M.; Zu, L. The NO-cGMP-PKG Axis in HFpEF: From Pathological Mechanisms to Potential Therapies. Aging Dis. 2023, 14, 46. [Google Scholar] [CrossRef]
- Sanders-van Wijk, S.; Tromp, J.; Beussink-Nelson, L.; Hage, C.; Svedlund, S.; Saraste, A.; Swat, S.A.; Sanchez, C.; Njoroge, J.; Tan, R.-S.; et al. Proteomic Evaluation of the Comorbidity-Inflammation Paradigm in Heart Failure with Preserved Ejection Fraction: Results From the PROMIS-HFpEF Study. Circulation 2020, 142, 2029–2044. [Google Scholar] [CrossRef] [PubMed]
- Vahedi-Mazdabadi, Y.; Shahinfar, H.; Toushih, M.; Shidfar, F. Effects of Berberine and Barberry on Selected Inflammatory Biomarkers in Adults: A Systematic Review and Dose-Response Meta-Analysis of Randomized Clinical Trials. Phytother. Res. 2023, 37, 5541–5557. [Google Scholar] [CrossRef]
- Engin, A. Reappraisal of Adipose Tissue Inflammation in Obesity. Adv. Exp. Med. Biol. 2024, 1460, 297–327. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, S.; Nagai, Y.; Shutta, R.; Masuda, D.; Yamashita, S.; Seo, M.; Yamada, T.; Nakagawa, A.; Yasumura, Y.; Nakagawa, Y.; et al. Combination of Neutrophil-to-Lymphocyte and Platelet-to-Lymphocyte Ratios as a Novel Predictor of Cardiac Death in Patients with Acute Decompensated Heart Failure with Preserved Left Ventricular Ejection Fraction: A Multicenter Study. J. Am. Heart Assoc. 2023, 12, e026326. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Narisawa, M.; Li, P.; Piao, L.; Li, Y.; Yang, G.; Cheng, X.W. Adaptive Immune Disorders in Hypertension and Heart Failure: Focusing on T-Cell Subset Activation and Clinical Implications. J. Hypertens. 2020, 38, 1878–1889. [Google Scholar] [CrossRef]
- Xiaojing, C.; Yanfang, L.; Yanqing, G.; Fangfang, C. Thymopentin Improves Cardiac Function in Older Patients with Chronic Heart Failure. Anatol. J. Cardiol. 2017, 17, 24–30. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Du, X.; Ma, H.; Yao, J. The Anti-Cancer Mechanisms of Berberine: A Review. Cancer Manag. Res. 2020, 12, 695–702. [Google Scholar] [CrossRef]
- Ehteshamfar, S.-M.; Akhbari, M.; Afshari, J.T.; Seyedi, M.; Nikfar, B.; Shapouri-Moghaddam, A.; Ghanbarzadeh, E.; Momtazi-Borojeni, A.A. Anti-Inflammatory and Immune-Modulatory Impacts of Berberine on Activation of Autoreactive T Cells in Autoimmune Inflammation. J. Cell Mol. Med. 2020, 24, 13573–13588. [Google Scholar] [CrossRef]
- Surendar, J.; Frohberger, S.J.; Karunakaran, I.; Schmitt, V.; Stamminger, W.; Neumann, A.-L.; Wilhelm, C.; Hoerauf, A.; Hübner, M.P. Adiponectin Limits IFN-γ and IL-17 Producing CD4 T Cells in Obesity by Restraining Cell Intrinsic Glycolysis. Front. Immunol. 2019, 10, 2555. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Y.; Geng, J.; Liu, C.; Jiang, S.; Han, Y.; Jiang, J.; Wang, Y. Exploring the Multi-Faceted Effects of Berberine in Ameliorating Diastolic Dysfunction in Rats with Heart Failure with Preserved Ejection Fraction. Int. J. Mol. Sci. 2025, 26, 4847. https://doi.org/10.3390/ijms26104847
Mu Y, Geng J, Liu C, Jiang S, Han Y, Jiang J, Wang Y. Exploring the Multi-Faceted Effects of Berberine in Ameliorating Diastolic Dysfunction in Rats with Heart Failure with Preserved Ejection Fraction. International Journal of Molecular Sciences. 2025; 26(10):4847. https://doi.org/10.3390/ijms26104847
Chicago/Turabian StyleMu, Yu, Jing Geng, Chilu Liu, Shuang Jiang, Yanxing Han, Jiandong Jiang, and Yuhong Wang. 2025. "Exploring the Multi-Faceted Effects of Berberine in Ameliorating Diastolic Dysfunction in Rats with Heart Failure with Preserved Ejection Fraction" International Journal of Molecular Sciences 26, no. 10: 4847. https://doi.org/10.3390/ijms26104847
APA StyleMu, Y., Geng, J., Liu, C., Jiang, S., Han, Y., Jiang, J., & Wang, Y. (2025). Exploring the Multi-Faceted Effects of Berberine in Ameliorating Diastolic Dysfunction in Rats with Heart Failure with Preserved Ejection Fraction. International Journal of Molecular Sciences, 26(10), 4847. https://doi.org/10.3390/ijms26104847