
Genome-Wide Identification and Expression Analysis of HSP70 Gene Family Under High-Temperature Stress in Lettuce (Lactuca sativa L.)


Abstract
1. Introduction
2. Results
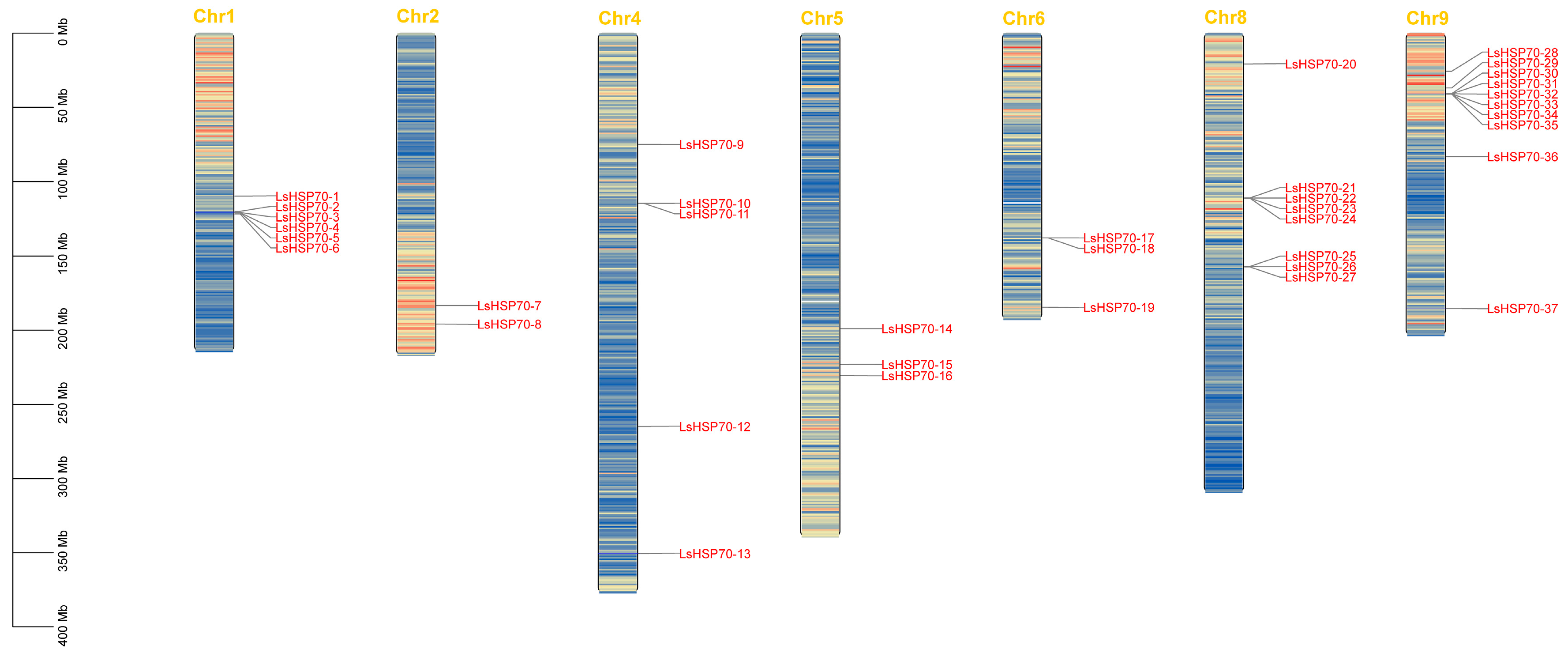
2.1. Identification, Chromosomal Localization and Physicochemical Properties of HSP70 Family Members in Lettuce
2.2. Phylogenetic Relationship Analysis of LsHSP70
2.3. Gene Structure and Motif Localization Analysis of LsHSP70
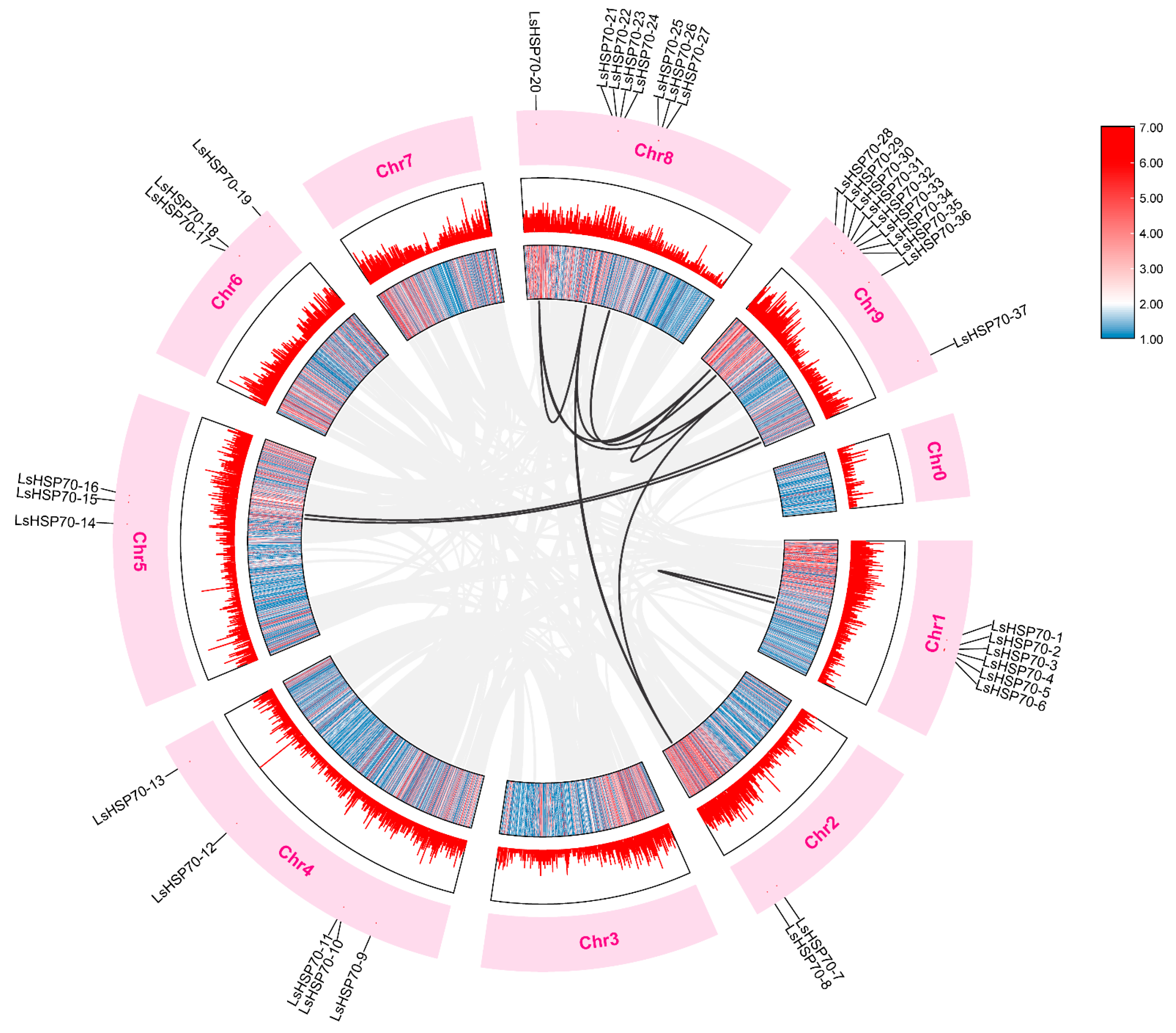
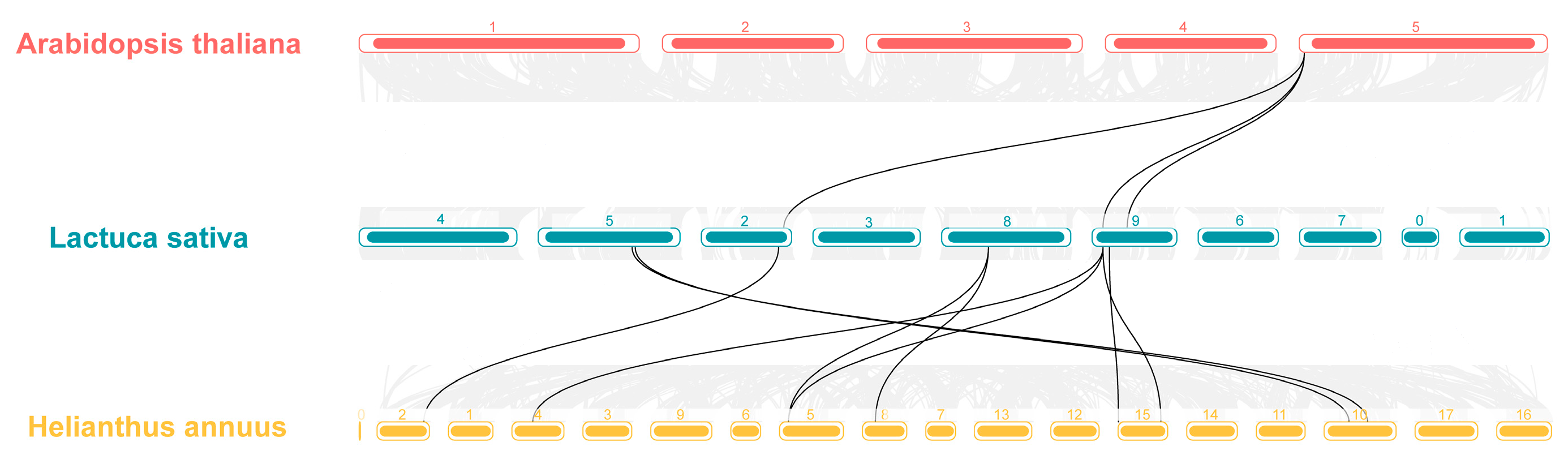
2.4. Covariance Analysis of LsHSP70
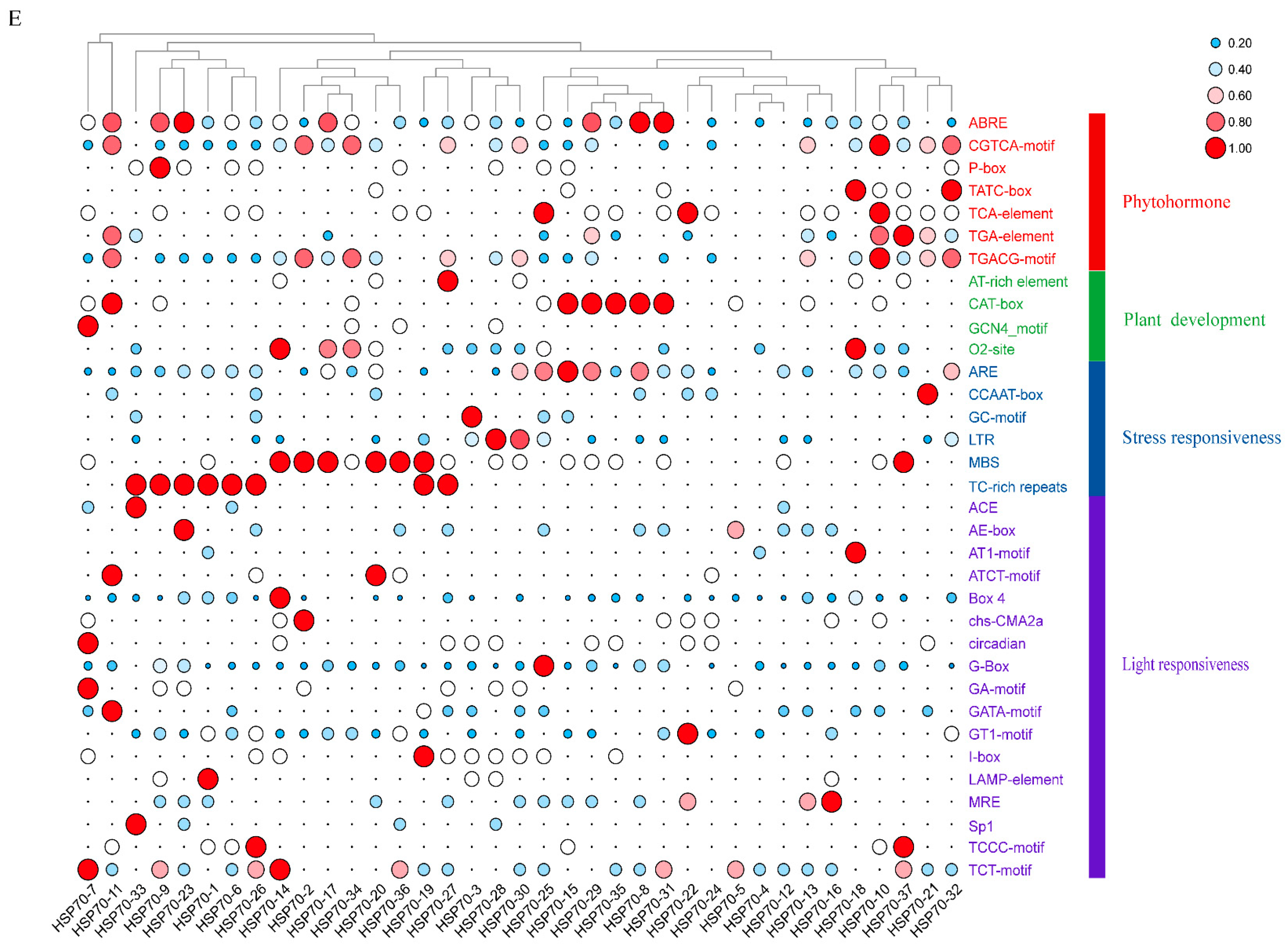
2.5. Cis-Acting Element Analysis of LsHSP70
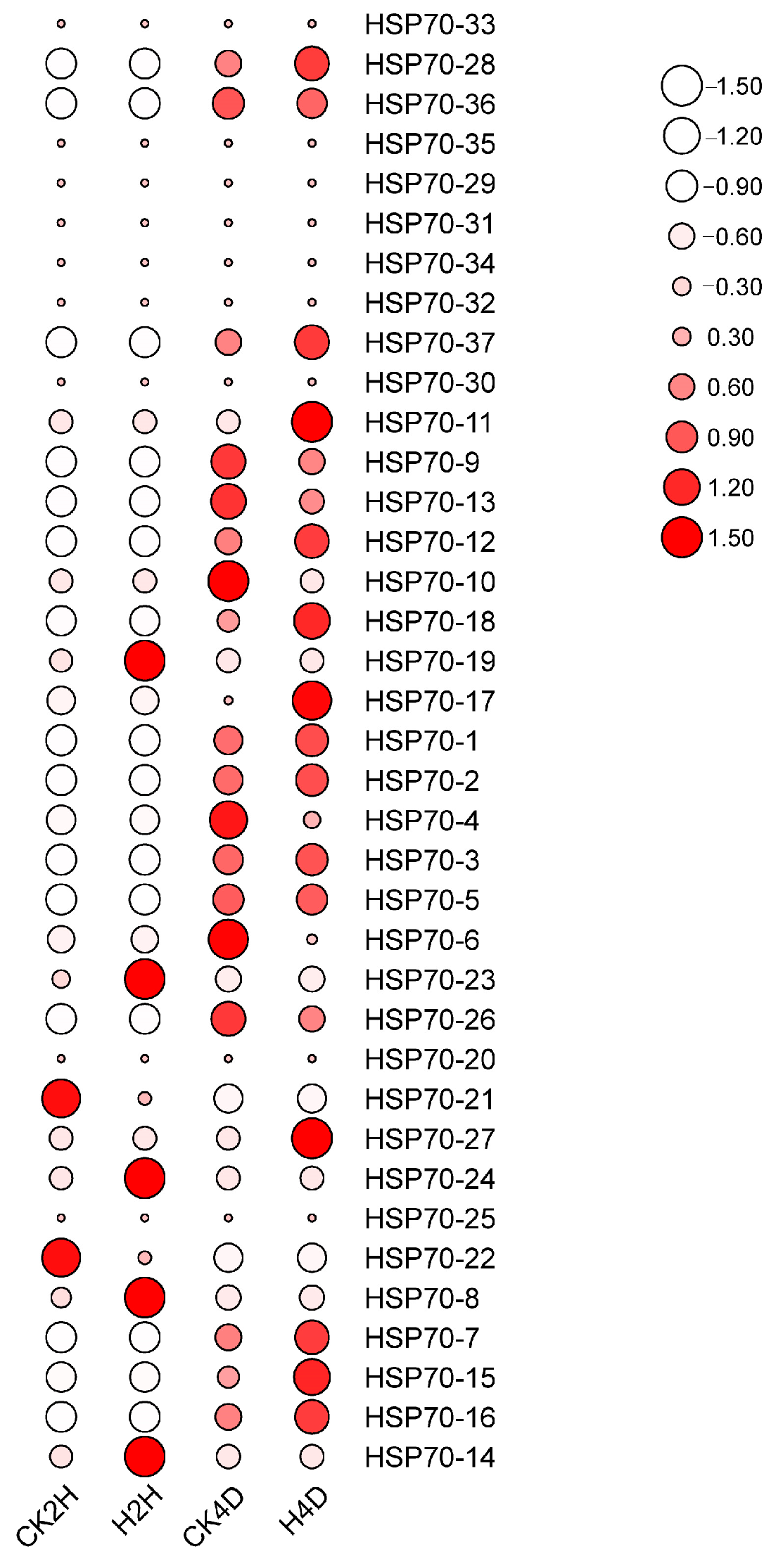
2.6. Analysis of the Expression Pattern of LsHSP70 Under High-Temperature Stress
2.7. Role of LsHSP70-19 Under High-Temperature Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Identification and Physicochemical Properties of HSP70 Family Members in Lettuce
4.3. Chromosomal Localization of LsHSP70 and Construction of Phylogenetic Tree
4.4. Gene Structure and Motif Analysis of LsHSP70
4.5. Analysis of Covariance
4.6. Analysis of Promoter Action Elements of LsHSP70 Gene
4.7. Expression Analysis of LsHSP70-19 Under High-Temperature Stress
4.8. Construction of VIGS Gene Silencing and Overexpression Vector
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, T.Z.; Liu, M.J.; Wu, Y.; Tian, Y.F.; Han, Y.Y.; Liu, C.J.; Hao, J.H.; Fan, S.X. Genome-Wide Identification and Expression Analysis of MAPK Gene Family in Lettuce (Lactuca sativa L.) and Functional Analysis of LsMAPK4 in High—Temperature-Induced Bolting. Int. J. Mol. Sci. 2022, 23, 11129. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, J.H.; Lu, J.P.; Zhai, Y.F.; Wang, H.; Gong, Z.H.; Wang, S.B.; Lu, M.H. Genome-wide analysis of the CaHsp20 gene family in pepper: Comprehensive sequence and expression profile analysis under heat stress. Front. Plant Sci. 2015, 6, 806. [Google Scholar] [CrossRef]
- Johnson, J.L.; Brown, C. Plasticity of the Hsp90 chaperone machine in divergent eukaryotic organisms. Cell Stress Chaperones 2009, 14, 83–94. [Google Scholar] [CrossRef]
- Wu, J.P. Cloning and functional identification of chrysanthemum heat shock protein gene CmHSP90.5. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2020. [Google Scholar]
- Song, Z.; Pan, F.; Lou, X.; Wang, D.; Yang, C.; Zhang, B.; Zhang, H. Genome-wide identification and characterization of Hsp70 gene family in Nicotiana tabacum. Mol. Biol. Rep. 2019, 46, 1941–1954. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, K.M.; DeLuca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat-shock cognate protein. Nature 1990, 346, 623–628. [Google Scholar] [CrossRef]
- Liu, J.; Pang, X.; Cheng, Y.; Yin, Y.; Zhang, Q.; Su, W.; Hu, B.; Guo, Q.; Ha, S.; Zhang, J.; et al. The Hsp70 Gene Family in Solanum tuberosum: Genome-Wide Identification, Phylogeny, and Expression Patterns. Sci. Rep. 2018, 8, 16628. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Zhai, Y.F.; Gong, Z.H.; Lu, M.H. Genome-wide analysis of the Hsp70 family genes in pepper (Capsicum annuum L.) and functional identification of CaHsp70-2 involvement in heat stress. Plant Sci. 2016, 252, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Sung, D.Y.; Guy, C.L. Physiological and molecular assessment of altered expression of Hsc70-1 in Arabidopsis. Evidence for pleiotropic consequences. Plant Physiol. 2003, 132, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.K.; Hong, C.B. Over-expression of tobacco NtHSP70-1 contributes to drought-stress tolerance in plants. Plant Cell Rep. 2006, 25, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.H.; Guo, J.; Ding, K.; Wang, S.J.; Zhang, H.; Dai, X.W.; Chen, Y.Y.; Govers, F.; Huang, L.L.; Kang, Z.S. Characterization of a wheat HSP70 gene and its expression in response to stripe rust infection and abiotic stresses. Mol. Biol. Rep. 2011, 38, 301–307. [Google Scholar] [CrossRef]
- Kim, S.R.; An, G. Rice chloroplast-localized heat shock protein 70, OsHsp70CP1, is essential for chloroplast development under high-temperature conditions. J. Plant Physiol. 2013, 170, 854–863. [Google Scholar] [CrossRef]
- Xinna, L.; Haiyang, C.; Shenchang, L.; Lijun, W. Genome-Wide Identification of the Hsp70 Gene Family in Grape and Their Expression Profile during Abiotic Stress. Horticulturae 2022, 8, 743. [Google Scholar] [CrossRef]
- Mengru, Y.; Rongqian, H.; Aiping, S.; Zhiyong, G.; Fadi, C.; Jiafu, J. Genome-Wide Identification and Expression Analysis of HSP70 Gene Family in Chrysanthemum lavandulifolium under Heat Stress. Horticulturae 2023, 9, 238. [Google Scholar] [CrossRef]
- Lin, B.L.; Wang, J.S.; Liu, H.C.; Chen, R.W.; Meyer, Y.; Barakat, A.; Delseny, M. Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 201–208. [Google Scholar] [CrossRef]
- Guy, C.L.; Li, Q.B. The organization and evolution of the spinach stress 70 molecular chaperone gene family. Plant Cell 1998, 10, 539–556. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Gho, H.J.; Nguyen, M.X.; Kim, S.R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genomics 2013, 13, 391–402. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, H.K.; Dong, Q.L.; Zhang, Y.Y.; Wang, Y.M.; Li, H.Y.; Xing, G.J.; Li, Q.Y.; Dong, Y.S. Genome-wide analysis and expression profiling under heat and drought treatments of HSP70 gene family in soybean (Glycine max L.). Front. Plant Sci. 2015, 6, 773. [Google Scholar] [CrossRef]
- Ni, Z.Y.; Liu, N.; Yu, Y.H.; Bi, C.X.; Chen, Q.J.; Qu, Y.Y. The cotton 70-kDa heat shock protein GhHSP70-26 plays a positive role in the drought stress response. Environ. Exp. Bot. 2021, 191, 104628. [Google Scholar] [CrossRef]
- Yang, X.; Wei, S.; Liu, B.; Guo, D.; Zheng, B.; Feng, L.; Liu, Y.; Tomás-Barberán, F.A.; Luo, L.; Huang, D. A novel integrated non-targeted metabolomic analysis reveals significant metabolite variations between different lettuce (Lactuca sativa L.) varieties. Hortic. Res. 2018, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Davoudi, M.; Chen, J.; Lou, Q. Genome-Wide Identification and Expression Analysis of Heat Shock Protein 70 (HSP70) Gene Family in Pumpkin (Cucurbita moschata L.) Rootstock under Drought Stress Suggested the Potential Role of these Chaperones in Stress Tolerance. Int. J. Mol. Sci. 2022, 23, 1918. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Atif, R.M.; Qayyum, A.; Du, X.; Hinze, L.; Azhar, M.T. Genome-wide identification and characterization of HSP70 gene family in four species of cotton. Genomics 2020, 112, 4442–4453. [Google Scholar] [CrossRef]
- Wang, X.R.; Wang, C.; Ban, F.X.; Zhu, D.T.; Liu, S.S.; Wang, X.W. Genome-wide identification and characterization of HSP gene superfamily in whitefly (Bemisia tabaci) and expression profiling analysis under temperature stress. Insect Sci. 2019, 26, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Zhang, Z.R.; Di, Z.X.; Qi, X.Y.; Zhang, Y.; Wang, C.; Lu, Y.Z.; Zheng, J. Identification of HSP70 gene family in Sorbus pohuashanensis and its expression analysis in response to abiotic stress. Genom. Appl. Biol. 2022, 41, 1973–1984. [Google Scholar] [CrossRef]
- Zhang, Y.T. Evolution of HSP70 gene family and functional analysis of GhHSP70-20 in Gossypium raimondii. Master’s Thesis, Zhejiang Agricultural and Forestry University, Hangzhou, China, 2014. [Google Scholar]
- Zhou, Z.; Xiao, L.; Zhao, J.; Hu, Z.; Zhou, Y.; Liu, S.; Wu, H.; Zhou, Y. Comprehensive Genomic Analysis and Expression Profile of Hsp70 Gene Family Related to Abiotic and Biotic Stress in Cucumber. Horticulturae 2023, 9, 1057. [Google Scholar] [CrossRef]
- Parra, G.; Bradnam, K.; Rose, A.B.; Korf, I. Comparative and functional analysis of intron-mediated enhancement signals reveals conserved features among plants. Nucleic Acids Res. 2011, 39, 5328–5337. [Google Scholar] [CrossRef] [PubMed]
- Magadum, S.; Banerjee, U.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Tanna, B. Halophytes: Potential Resources for Salt Stress Tolerance Genes and Promoters. Front. Plant Sci. 2017, 8, 829. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.F.; Yu, S.W.; Shu, L.B.; Wu, J.H.; Wu, A.Z.; Luo, L.J. Expression profile analysis of 9 heat shock protein genes throughout the life cycle and under abiotic stress in rice. Chin. Sci. Bull. 2012, 57, 336–347. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus ID | Chromosomal Location | Length (aa) | MW (kDa) | pI | Subcellular Localization | |||
|---|---|---|---|---|---|---|---|---|---|
| Chr | Chr-Start | Chr-End | Direction | ||||||
| LsHSP70-1 | Lsat_1_v5_gn_1_87340 | 1 | 109,889,259 | 109,895,753 | - | 645 | 72 | 5.18 | Endoplasmic reticulum |
| LsHSP70-2 | Lsat_1_v5_gn_1_91700 | 1 | 120,103,665 | 120,109,556 | + | 617 | 69 | 8.11 | Endoplasmic reticulum |
| LsHSP70-4 | Lsat_1_v5_gn_1_93321 | 1 | 121,003,737 | 121,009,958 | + | 617 | 68 | 7.84 | Endoplasmic reticulum |
| LsHSP70-3 | Lsat_1_v5_gn_1_91141 | 1 | 120,849,862 | 120,855,133 | + | 617 | 69 | 6.51 | Mitochondrion |
| LsHSP70-5 | Lsat_1_v5_gn_1_91241 | 1 | 121,066,030 | 121,067,978 | - | 617 | 69 | 7.53 | Endoplasmic reticulum |
| LsHSP70-6 | Lsat_1_v5_gn_1_91381 | 1 | 121,849,518 | 121,855,607 | + | 617 | 69 | 8.48 | Endoplasmic reticulum |
| LsHSP70-8 | Lsat_1_v5_gn_2_116640 | 2 | 195,818,062 | 195,820,849 | + | 648 | 71 | 5.07 | Nucleus |
| LsHSP70-7 | Lsat_1_v5_gn_2_105720 | 2 | 183,329,797 | 183,331,788 | + | 626 | 70 | 5.30 | Chloroplast |
| LsHSP70-11 | Lsat_1_v5_gn_4_72600 | 4 | 114,729,310 | 114,731,461 | + | 614 | 69 | 8.01 | Mitochondrion |
| LsHSP70-9 | Lsat_1_v5_gn_4_50420 | 4 | 74,953,550 | 74,956,176 | + | 614 | 69 | 8.32 | Chloroplast |
| LsHSP70-13 | Lsat_1_v5_gn_4_173180 | 4 | 350,529,997 | 350,533,516 | + | 632 | 71 | 6.21 | Mitochondrion |
| LsHSP70-12 | Lsat_1_v5_gn_4_137301 | 4 | 264,836,010 | 264,839,010 | - | 636 | 70 | 8.28 | Endoplasmic reticulum Mitochondrion |
| LsHSP70-10 | Lsat_1_v5_gn_4_72580 | 4 | 114,717,746 | 114,719,920 | + | 616 | 69 | 8.20 | Mitochondrion |
| LsHSP70-15 | Lsat_1_v5_gn_5_104601 | 5 | 223,025,473 | 223,028,553 | - | 701 | 75 | 5.14 | Chloroplast |
| LsHSP70-16 | Lsat_1_v5_gn_5_110901 | 5 | 230,527,527 | 230,530,603 | - | 680 | 73 | 5.78 | Mitochondrion |
| LsHSP70-14 | Lsat_1_v5_gn_5_89440 | 5 | 198,889,223 | 198,891,499 | + | 649 | 71 | 5.18 | Nucleus |
| LsHSP70-18 | Lsat_1_v5_gn_6_83520 | 6 | 137,995,823 | 137,998,823 | - | 661 | 73 | 5.08 | Endoplasmic reticulum Nucleus |
| LsHSP70-19 | Lsat_1_v5_gn_6_112621 | 6 | 184,461,293 | 184,464,461 | - | 690 | 74 | 5.11 | Chloroplast |
| LsHSP70-17 | Lsat_1_v5_gn_6_83581 | 6 | 137,948,656 | 137,951,646 | + | 623 | 69 | 5.84 | Endoplasmic reticulum Nucleus |
| LsHSP70-23 | Lsat_1_v5_gn_8_78021 | 8 | 111,243,941 | 111,246,426 | + | 649 | 71 | 5.13 | Nucleus |
| LsHSP70-26 | Lsat_1_v5_gn_8_103521 | 8 | 156,999,462 | 157,001,748 | - | 610 | 68 | 6.47 | Chloroplast |
| LsHSP70-20 | Lsat_1_v5_gn_8_15681 | 8 | 20,742,298 | 20,744,707 | + | 651 | 72 | 5.58 | Nucleus |
| LsHSP70-21 | Lsat_1_v5_gn_8_78141 | 8 | 111,049,782 | 111,052,054 | + | 646 | 71 | 5.12 | Chloroplast |
| LsHSP70-27 | Lsat_1_v5_gn_8_103901 | 8 | 157,310,376 | 157,312,452 | - | 613 | 68 | 8.35 | Mitochondrion |
| LsHSP70-24 | Lsat_1_v5_gn_8_78000 | 8 | 111,278,374 | 111,281,033 | - | 648 | 71 | 5.15 | Endoplasmic reticulum Mitochondrion |
| LsHSP70-25 | Lsat_1_v5_gn_8_104441 | 8 | 156,970,950 | 156,973,039 | + | 614 | 68 | 8.47 | Mitochondrion |
| LsHSP70-22 | Lsat_1_v5_gn_8_78100 | 8 | 111,121,876 | 111,124,251 | - | 646 | 71 | 5.12 | Chloroplast |
| LsHSP70-33 | Lsat_1_v5_gn_9_36681 | 9 | 40,914,392 | 40,916,855 | - | 598 | 66 | 8.24 | Chloroplast |
| LsHSP70-28 | Lsat_1_v5_gn_9_22920 | 9 | 25,664,742 | 25,667,641 | + | 647 | 71 | 5.11 | Chloroplast |
| LsHSP70-36 | Lsat_1_v5_gn_9_65381 | 9 | 83,179,845 | 83,182,256 | + | 647 | 71 | 5.2 | Chloroplast |
| LsHSP70-35 | Lsat_1_v5_gn_9_36981 | 9 | 41,045,215 | 41,047,623 | - | 610 | 68 | 8.02 | Chloroplast |
| LsHSP70-31 | Lsat_1_v5_gn_9_36560 | 9 | 40,874,942 | 40,877,359 | + | 635 | 71 | 8.55 | Endoplasmic reticulum Mitochondrion |
| LsHSP70-29 | Lsat_1_v5_gn_9_34760 | 9 | 36,912,224 | 36,914,256 | + | 613 | 68 | 5.08 | Mitochondrion Nucleus |
| LsHSP70-34 | Lsat_1_v5_gn_9_36861 | 9 | 40,973,006 | 40,975,340 | - | 610 | 68 | 5.56 | Chloroplast |
| LsHSP70-32 | Lsat_1_v5_gn_9_36600 | 9 | 40,892,961 | 40,895,271 | + | 608 | 67 | 5.73 | Chloroplast Nucleus |
| LsHSP70-37 | Lsat_1_v5_gn_9_113681 | 9 | 185,253,863 | 185,258,114 | - | 682 | 73 | 5.89 | Mitochondrion |
| LsHSP70-30 | Lsat_1_v5_gn_9_36480 | 9 | 40,736,150 | 40,738,239 | + | 610 | 68 | 6.20 | Chloroplast Nucleus |
| Treatment | Fresh Weight (g) | Plant Height (cm) | Root Length (cm) | Shoot Weight (g) | Root Weight (g) |
|---|---|---|---|---|---|
| CK | 14.49 ± 1.05 b | 17.97 ± 2.40 a | 21.6 ± 1.67 b | 9.89 ± 0.84 b | 2.34 ± 0.50 a |
| Overexpression | 19.5 ± 1.22 a | 21.67 ± 2.00 a | 27.53 ± 2.21 a | 17.31 ± 0.67 a | 2.32 ± 0.92 a |
| Silence | 10.44 ± 0.91 c | 19.63 ± 0.78 a | 27.67 ± 3.21 a | 7.26 ± 0.12 c | 1.66 ± 0.36 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Sun, W.; Duan, Y.; Xu, Y.; Wang, H.; Hao, J.; Han, Y.; Liu, C. Genome-Wide Identification and Expression Analysis of HSP70 Gene Family Under High-Temperature Stress in Lettuce (Lactuca sativa L.). Int. J. Mol. Sci. 2025, 26, 102. https://doi.org/10.3390/ijms26010102
Wang Q, Sun W, Duan Y, Xu Y, Wang H, Hao J, Han Y, Liu C. Genome-Wide Identification and Expression Analysis of HSP70 Gene Family Under High-Temperature Stress in Lettuce (Lactuca sativa L.). International Journal of Molecular Sciences. 2025; 26(1):102. https://doi.org/10.3390/ijms26010102
Chicago/Turabian StyleWang, Qian, Wenjing Sun, Yipei Duan, Yikun Xu, Huiyu Wang, Jinghong Hao, Yingyan Han, and Chaojie Liu. 2025. "Genome-Wide Identification and Expression Analysis of HSP70 Gene Family Under High-Temperature Stress in Lettuce (Lactuca sativa L.)" International Journal of Molecular Sciences 26, no. 1: 102. https://doi.org/10.3390/ijms26010102
APA StyleWang, Q., Sun, W., Duan, Y., Xu, Y., Wang, H., Hao, J., Han, Y., & Liu, C. (2025). Genome-Wide Identification and Expression Analysis of HSP70 Gene Family Under High-Temperature Stress in Lettuce (Lactuca sativa L.). International Journal of Molecular Sciences, 26(1), 102. https://doi.org/10.3390/ijms26010102