
Maternal Diet High in Linoleic Acid Alters Renal Branching Morphogenesis and mTOR/AKT Signalling Genes in Rat Fetal Kidneys
 ,
,  , ,
, ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Effect of Maternal HLA Diet on Renal Genes Involved in Branching Morphogenesis
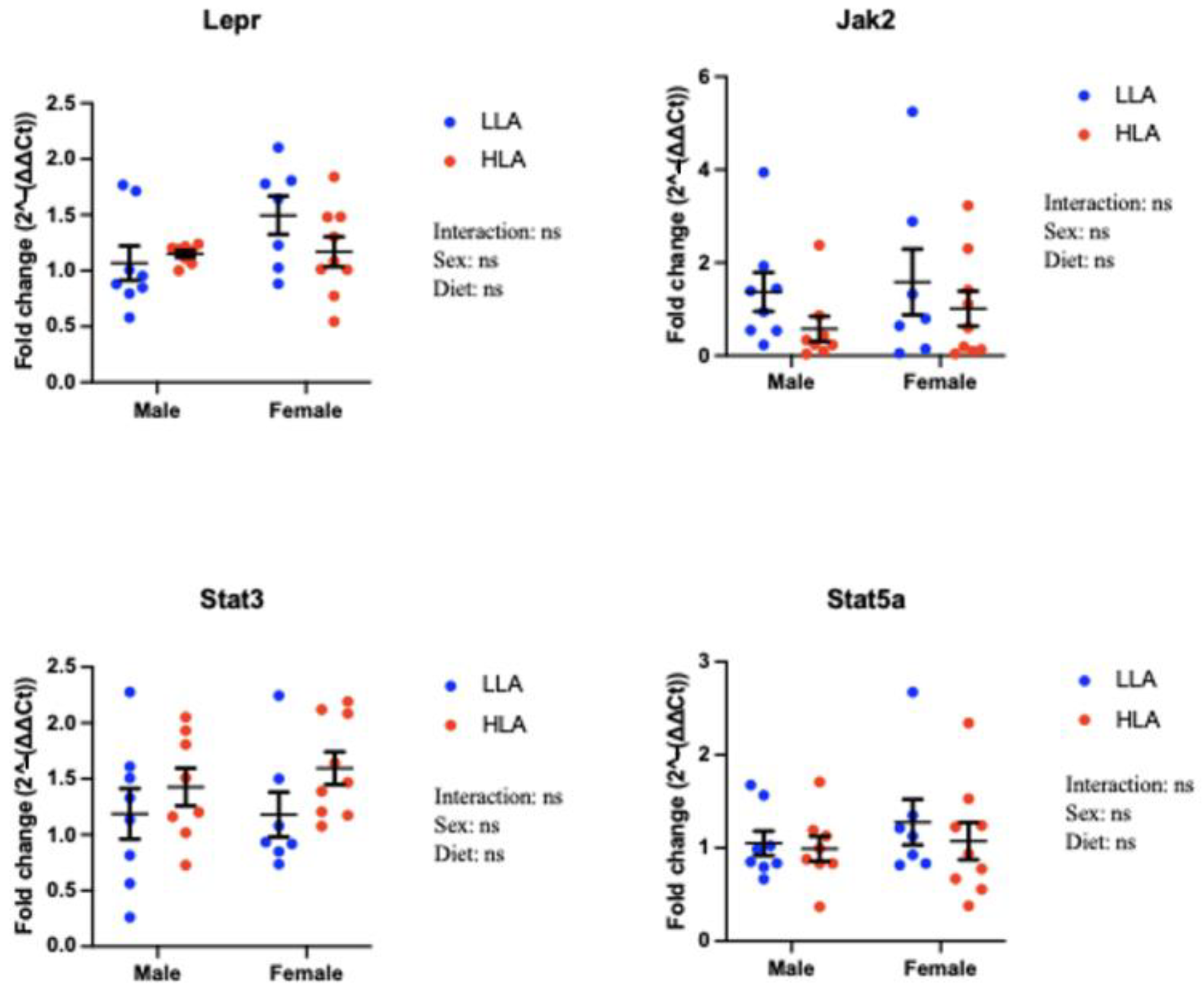
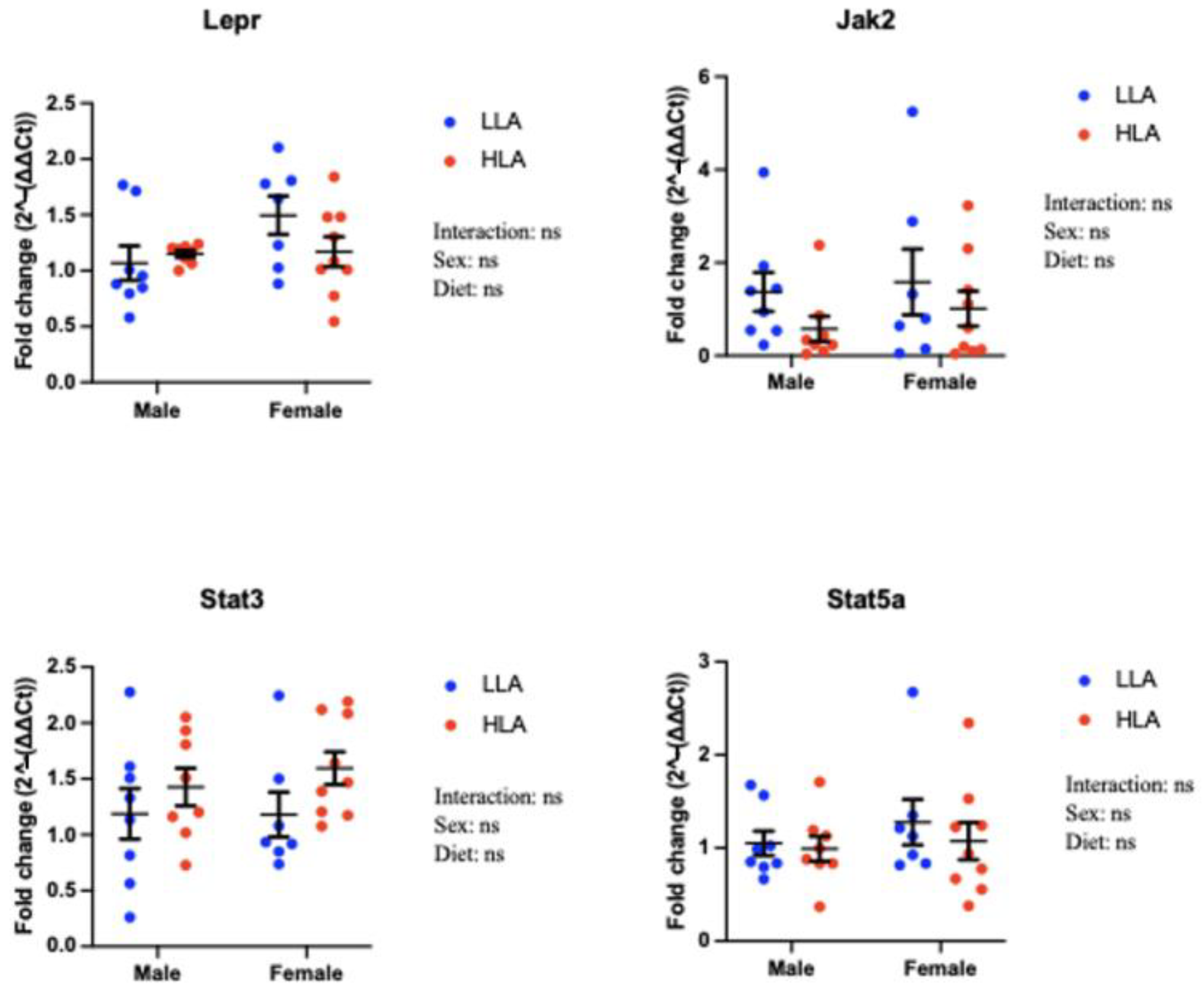
2.2. Effect of Maternal HLA Diet on Renal Genes Involved in Leptin Signalling in Embryonic Offspring
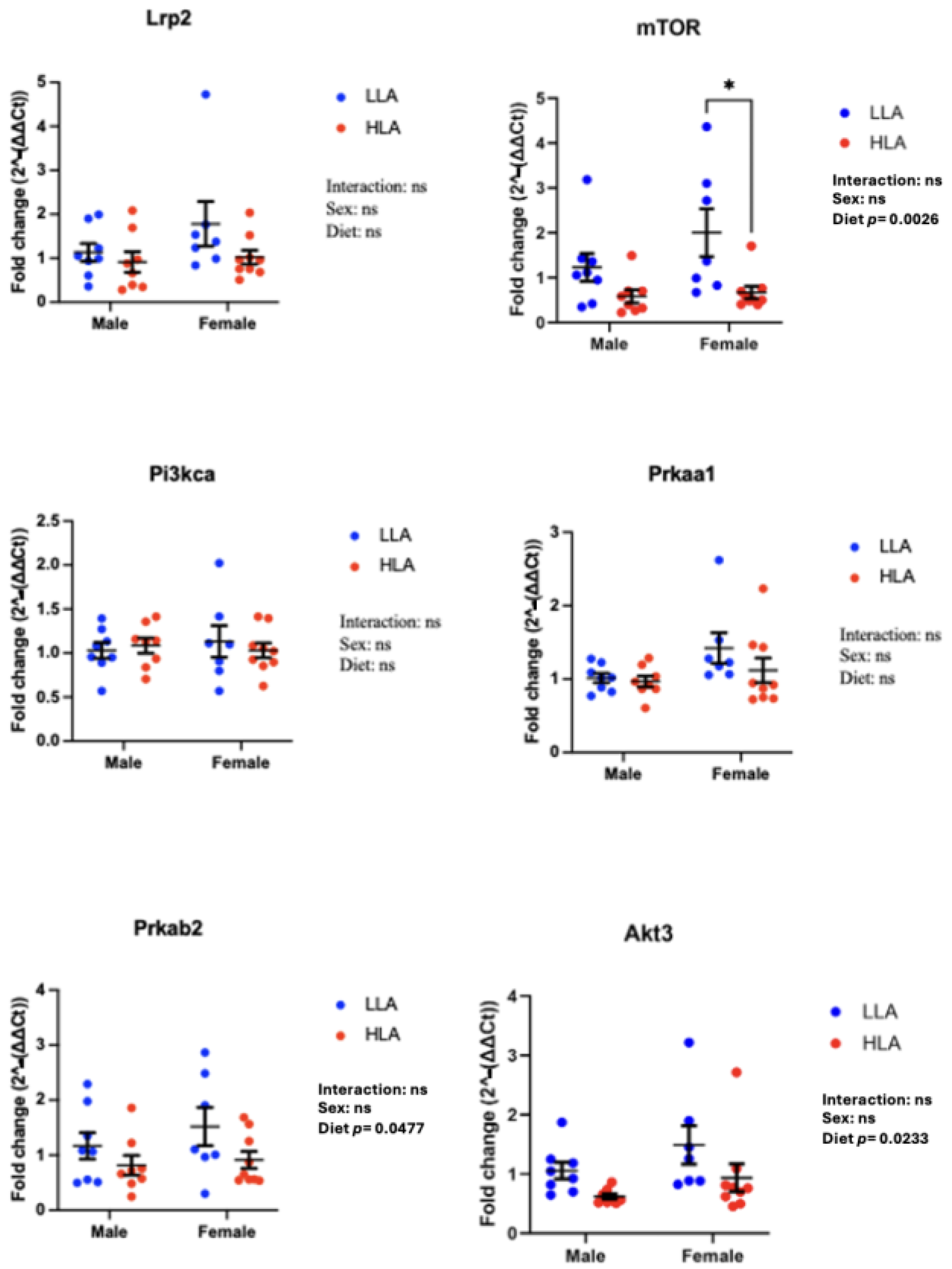
2.3. Effect of Maternal HLA Diet on Renal Genes Involved in Megalin Signalling in Embryonic Offspring
2.4. Effect of Maternal HLA Diet on Angiogenesis Genes in Embryonic Kidneys
3. Discussion
4. Materials and Methods
4.1. Experimental Animal Model and Diet
4.2. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Linoleic acid and the pathogenesis of obesity. Prostaglandins Other Lipid Mediat. 2016, 125, 90–99. [Google Scholar] [CrossRef]
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Australia’s nutrition transition 1961–2009: A focus on fats. Br. J. Nutr. 2015, 114, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, C.E.; Ringel, A.; Feldstein, A.E.; Taha, A.Y.; MacIntosh, B.A.; Hibbeln, J.R.; Majchrzak-Hong, S.F.; Faurot, K.R.; Rapoport, S.I.; Cheon, Y.; et al. Lowering dietary linoleic acid reduces bioactive oxidized linoleic acid metabolites in humans. Prostaglandins Leukot. Essent. Fat. Acids 2012, 87, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Ailhaud, G.; Massiera, F.; Weill, P.; Legrand, P.; Alessandri, J.M.; Guesnet, P. Temporal changes in dietary fats: Role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog. Lipid Res. 2006, 45, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.; Álvarez, D.; Muñoz, Y.; Crisosto, N.; Valenzuela, R.; Maliqueo, M. Linoleic and arachidonic fatty acids and their potential relationship with inflammation, pregnancy, and fetal development. Curr. Med. Chem. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Cuffe, J.S.M.; Holland, O.J.; Bulmer, A.C.; Hill, M.; Perkins, A.V.; Muhlhausler, B.S.; McAinch, A.J.; Hryciw, D.H. Elevated maternal linoleic acid reduces circulating leptin concentrations, cholesterol levels and male fetal survival in a rat model. J. Physiol. 2019, 597, 3349–3361. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Holland, O.J.; Kent, N.L.; Perkins, A.V.; McAinch, A.J.; Cuffe, J.S.M.; Hryciw, D.H. Maternal High Linoleic Acid Alters Placental Fatty Acid Composition. Nutrients 2020, 12, 2183. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Sleep, S.; Helman, T.; Holland, O.; Cuffe, J.S.M.; Perkins, A.V.; McAinch, A.J.; Headrick, J.P.; Hryciw, D.H. Maternal diet high in linoleic acid alters offspring fatty acids and cardiovascular function in a rat model. Br. J. Nutr. 2022, 127, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Sleep, S.L.; Holland, O.J.; Vidimce, J.; Bulmer, A.C.; Cuffe, J.S.M.; Perkins, A.V.; McAinch, A.J.; Hryciw, D.H. Maternal Diet High in Linoleic Acid Alters Offspring Lipids and Hepatic Regulators of Lipid Metabolism in an Adolescent Rat Model. Int. J. Mol. Sci. 2024, 25, 1129. [Google Scholar] [CrossRef]
- Shrestha, N.; Vidimce, J.; Holland, O.J.; Cuffe, J.S.M.; Beck, B.R.; Perkins, A.V.; McAinch, A.J.; Hryciw, D.H. Maternal and Postnatal High Linoleic Acid Diet Impacts Lipid Metabolism in Adult Rat Offspring in a Sex-Specific Manner. Int. J. Mol. Sci. 2021, 22, 2946. [Google Scholar] [CrossRef] [PubMed]
- Attig, L.; Larcher, T.; Gertler, A.; Abdennebi-Najar, L.; Djaine, J. Postnatal leptin is necessary for maturation of numerous organs in newborn rats. Organogenesis 2011, 7, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Attig, L.; Solomon, G.; Ferezou, J.; Abdennebi-Najar, L.; Taouis, M.; Gertler, A.; Djiane, J. Early postnatal leptin blockage leads to a long-term leptin resistance and susceptibility to diet-induced obesity in rats. Int. J. Obes. 2008, 32, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Moritz, K.M.; Dodic, M.; Wintour, E.M. Kidney development and the fetal programming of adult disease. Bioessays 2003, 25, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Cuffe, J.S.M.; Briffa, J.F.; Rosser, S.; Siebel, A.L.; Romano, T.; Hryciw, D.H.; Wlodek, M.E.; Moritz, K.M. Uteroplacental insufficiency in rats induces renal apoptosis and delays nephrogenesis completion. Acta Physiol. 2018, 222, e12982. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.R.; Cullen-McEwen, L.A.; Kett, M.M.; Boon, W.M.; Dowling, J.; Bertram, J.F.; Moritz, K.M. Prenatal corticosterone exposure results in altered AT1/AT2, nephron deficit and hypertension in the rat offspring. J. Physiol. 2007, 579, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.P.; Denton, K.M.; Cullen-McEwen, L.; Bertram, J.F.; Moritz, K.M. Prenatal exposure to alcohol reduces nephron number and raises blood pressure in progeny. J. Am. Soc. Nephrol. 2010, 21, 1891–1902. [Google Scholar] [CrossRef] [PubMed]
- Short, K.M.; Smyth, I.M. Branching morphogenesis as a driver of renal development. Anat. Rec. 2020, 303, 2578–2587. [Google Scholar] [CrossRef]
- Briffa, J.F.; McAinch, A.J.; Romano, T.; Wlodek, M.E.; Hryciw, D.H. Leptin in pregnancy and development: A contributor to adulthood disease? Am. J. Physiol. Endocrinol. Metab. 2015, 308, E335–E350. [Google Scholar] [CrossRef]
- Ahima, R.S.; Osei, S.Y. Leptin signaling. Physiol. Behav. 2004, 81, 223–241. [Google Scholar] [CrossRef]
- Briffa, J.F.; Grinfeld, E.; Poronnik, P.; McAinch, A.J.; Hryciw, D.H. Uptake of leptin and albumin via separate pathways in proximal tubule cells. Int. J. Biochem. Cell Biol. 2016, 79, 194–198. [Google Scholar] [CrossRef] [PubMed]
- van Abeelen, A.F.; de Rooij, S.R.; Osmond, C.; Painter, R.C.; Veenendaal, M.V.; Bossuyt, P.M.; Elias, S.G.; Grobbee, D.E.; van der Schouw, Y.T.; Barker, D.J.; et al. The sex-specific effects of famine on the association between placental size and later hypertension. Placenta 2011, 32, 694–698. [Google Scholar] [CrossRef]
- Cheong, J.N.; Cuffe, J.S.; Jefferies, A.J.; Anevska, K.; Moritz, K.M.; Wlodek, M.E. Sex-Specific Metabolic Outcomes in Offspring of Female Rats Born Small or Exposed to Stress During Pregnancy. Endocrinology 2016, 157, 4104–4120. [Google Scholar] [CrossRef]
- Thomas, K.N.; Zimmel, K.N.; Roach, A.N.; Basel, A.; Mehta, N.A.; Bedi, Y.S.; Golding, M.C. Maternal background alters the penetrance of growth phenotypes and sex-specific placental adaptation of offspring sired by alcohol-exposed males. FASEB J. 2021, 35, e22035. [Google Scholar] [CrossRef]
- Shrestha, N.; Melvin, S.D.; McKeating, D.R.; Holland, O.J.; Cuffe, J.S.M.; Perkins, A.V.; McAinch, A.J.; Hryciw, D.H. Sex-Specific Differences in Lysine, 3-Hydroxybutyric Acid and Acetic Acid in Offspring Exposed to Maternal and Postnatal High Linoleic Acid Diet, Independent of Diet. Int. J. Mol. Sci. 2021, 22, 10223. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cerezales, S.; Ramos-Ibeas, P.; Rizos, D.; Lonergan, P.; Bermejo-Alvarez, P.; Gutiérrez-Adán, A. Early sex-dependent differences in response to environmental stress. Reproduction 2018, 155, R39–R51. [Google Scholar] [CrossRef]
- Bazer, F.W.; Spencer, T.E.; Wu, G.; Cudd, T.A.; Meininger, C.J. Maternal Nutrition and Fetal Development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef]
- Vega, Q.C.; Worby, C.A.; Lechner, M.S.; Dixon, J.E.; Dressler, G.R. Glial cell line-derived neurotrophic factor activates the receptor tyrosine kinase RET and promotes kidney morphogenesis. Proc. Natl. Acad. Sci. USA 1996, 93, 10657–10661. [Google Scholar] [CrossRef]
- Walker, K.A.; Bertram, J.F. Kidney Development: Core Curriculum 2011. Am. J. Kidney Dis. 2011, 57, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Henry, T.; Abdallah, M.; Abdel-Hakeem, A.; Desai, M.; Nast, C.; Mansano, R.Z.; Toss, M.; Magee, T. 458: Reduced nephrogenesis in low birth offspring is regulated by GDNF signaling dysregulation through the MAPK/ERK signaling pathway. Am. J. Obstet. Gynecol. 2008, 199, S137. [Google Scholar] [CrossRef]
- Abdel-Hakeem, A.K.; Henry, T.Q.; Magee, T.R.; Desai, M.; Ross, M.G.; Mansano, R.Z.; Torday, J.S.; Nast, C.C. Mechanisms of impaired nephrogenesis with fetal growth restriction: Altered renal transcription and growth factor expression. Am. J. Obstet. Gynecol. 2008, 199, 252.e1–252.e7. [Google Scholar] [CrossRef] [PubMed]
- Costantini, F. GDNF/Ret signaling and renal branching morphogenesis. Organogenesis 2010, 6, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Wood-Bradley, R.J.; Henry, S.L.; Barrand, S.; Giot, A.; Eipper, L.; Bertram, J.F.; Cullen-McEwen, L.A.; Armitage, J.A. Analysis of structure and gene expression in developing kidneys of male and female rats exposed to low protein diets in utero. Anat. Rec. 2020, 303, 2657–2667. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.E.; Howie, S.E.M. The role of megalin (LRP-2/Gp330) during development. Dev. Biol. 2006, 296, 279–297. [Google Scholar] [CrossRef]
- Chai, O.-H.; Song, C.-H.; Park, S.-K.; Kim, W.; Cho, E.-S. Molecular regulation of kidney development. Anat. Cell Biol. 2013, 46, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Marwarha, G.; Dasari, B.; Prabhakara, J.P.R.; Schommer, J.; Ghribi, O. β-Amyloid regulates leptin expression and tau phosphorylation through the mTORC1 signaling pathway. J. Neurochem. 2010, 115, 373–384. [Google Scholar] [CrossRef]
- Fantus, D.; Rogers, N.M.; Grahammer, F.; Huber, T.B.; Thomson, A.W. Roles of mTOR complexes in the kidney: Implications for renal disease and transplantation. Nat. Rev. Nephrol. 2016, 12, 587–609. [Google Scholar] [CrossRef] [PubMed]
- Carney, E. Regulation of nephrogenesis. Nat. Rev. Nephrol. 2018, 14, 536. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Kun-Liang, G. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Thoreen, C.C.; Chantranupong, L.; Keys, H.R.; Wang, T.; Gray, N.S.; Sabatini, D.M. A unifying model for mTORC1-mediated regulation of mRNA translation. Nature 2012, 485, 109–113. [Google Scholar] [CrossRef]
- Gleixner, E.M.; Canaud, G.; Hermle, T.; Guida, M.C.; Kretz, O.; Helmstädter, M.; Huber, T.B.; Eimer, S.; Terzi, F.; Simons, M. V-ATPase/mTOR signaling regulates megalin-mediated apical endocytosis. Cell Rep. 2014, 8, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, E.C.; Cripps, R.L.; Duncan, J.S.; Barrett, P.; Mercer, J.G.; Herwig, A.; Ozanne, S.E. Developmental changes in hypothalamic leptin receptor: Relationship with the postnatal leptin surge and energy balance neuropeptides in the postnatal rat. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2009, 296, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Delahaye, F.; Breton, C.; Risold, P.-Y.; Enache, M.; Dutriez-Casteloot, I.; Laborie, C.; Lesage, J.; Vieau, D. Maternal Perinatal Undernutrition Drastically Reduces Postnatal Leptin Surge and Affects the Development of Arcuate Nucleus Proopiomelanocortin Neurons in Neonatal Male Rat Pups. Endocrinology 2008, 149, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Rosario, F.J.; Jansson, N.; Kanai, Y.; Prasad, P.D.; Powell, T.L.; Jansson, T. Maternal Protein Restriction in the Rat Inhibits Placental Insulin, mTOR, and STAT3 Signaling and Down-Regulates Placental Amino Acid Transporters. Endocrinology 2011, 152, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, A.; Maymó, J.; Dueñas, J.L.; Goberna, R.; Calvo, J.C.; Varone, C.; Sánchez-Margalet, V. Leptin prevents apoptosis of trophoblastic cells by activation of MAPK pathway. Arch. Biochem. Biophys. 2008, 477, 390–395. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. Br. J. Pharmacol. 2020, 177, 3617–3624. [Google Scholar] [CrossRef] [PubMed]
- Choque, B.; Catheline, D.; Delplanque, B.; Guesnet, P.; Legrand, P. Dietary linoleic acid requirements in the presence of α-linolenic acid are lower than the historical 2% of energy intake value, study in rats. Br. J. Nutr. 2015, 113, 1056–1068. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McClelland, C.; Holland, O.J.; Shrestha, N.; Jukes, C.L.; Brandon, A.E.; Cuffe, J.S.M.; Perkins, A.V.; McAinch, A.J.; Hryciw, D.H. Maternal Diet High in Linoleic Acid Alters Renal Branching Morphogenesis and mTOR/AKT Signalling Genes in Rat Fetal Kidneys. Int. J. Mol. Sci. 2024, 25, 4688. https://doi.org/10.3390/ijms25094688
McClelland C, Holland OJ, Shrestha N, Jukes CL, Brandon AE, Cuffe JSM, Perkins AV, McAinch AJ, Hryciw DH. Maternal Diet High in Linoleic Acid Alters Renal Branching Morphogenesis and mTOR/AKT Signalling Genes in Rat Fetal Kidneys. International Journal of Molecular Sciences. 2024; 25(9):4688. https://doi.org/10.3390/ijms25094688
Chicago/Turabian StyleMcClelland, Connie, Olivia J. Holland, Nirajan Shrestha, Claire L. Jukes, Anna E. Brandon, James S. M. Cuffe, Anthony V. Perkins, Andrew J. McAinch, and Deanne H. Hryciw. 2024. "Maternal Diet High in Linoleic Acid Alters Renal Branching Morphogenesis and mTOR/AKT Signalling Genes in Rat Fetal Kidneys" International Journal of Molecular Sciences 25, no. 9: 4688. https://doi.org/10.3390/ijms25094688
APA StyleMcClelland, C., Holland, O. J., Shrestha, N., Jukes, C. L., Brandon, A. E., Cuffe, J. S. M., Perkins, A. V., McAinch, A. J., & Hryciw, D. H. (2024). Maternal Diet High in Linoleic Acid Alters Renal Branching Morphogenesis and mTOR/AKT Signalling Genes in Rat Fetal Kidneys. International Journal of Molecular Sciences, 25(9), 4688. https://doi.org/10.3390/ijms25094688