

[Cu(NN1)2]ClO4, a Copper (I) Complex as an Antimicrobial Agent for the Treatment of Piscirickettsiosis in Atlantic Salmon
 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Synthesis of Cu (I) Coordination Complex
2.2. Antibacterial Activity of [Cu(NN1)2]ClO4
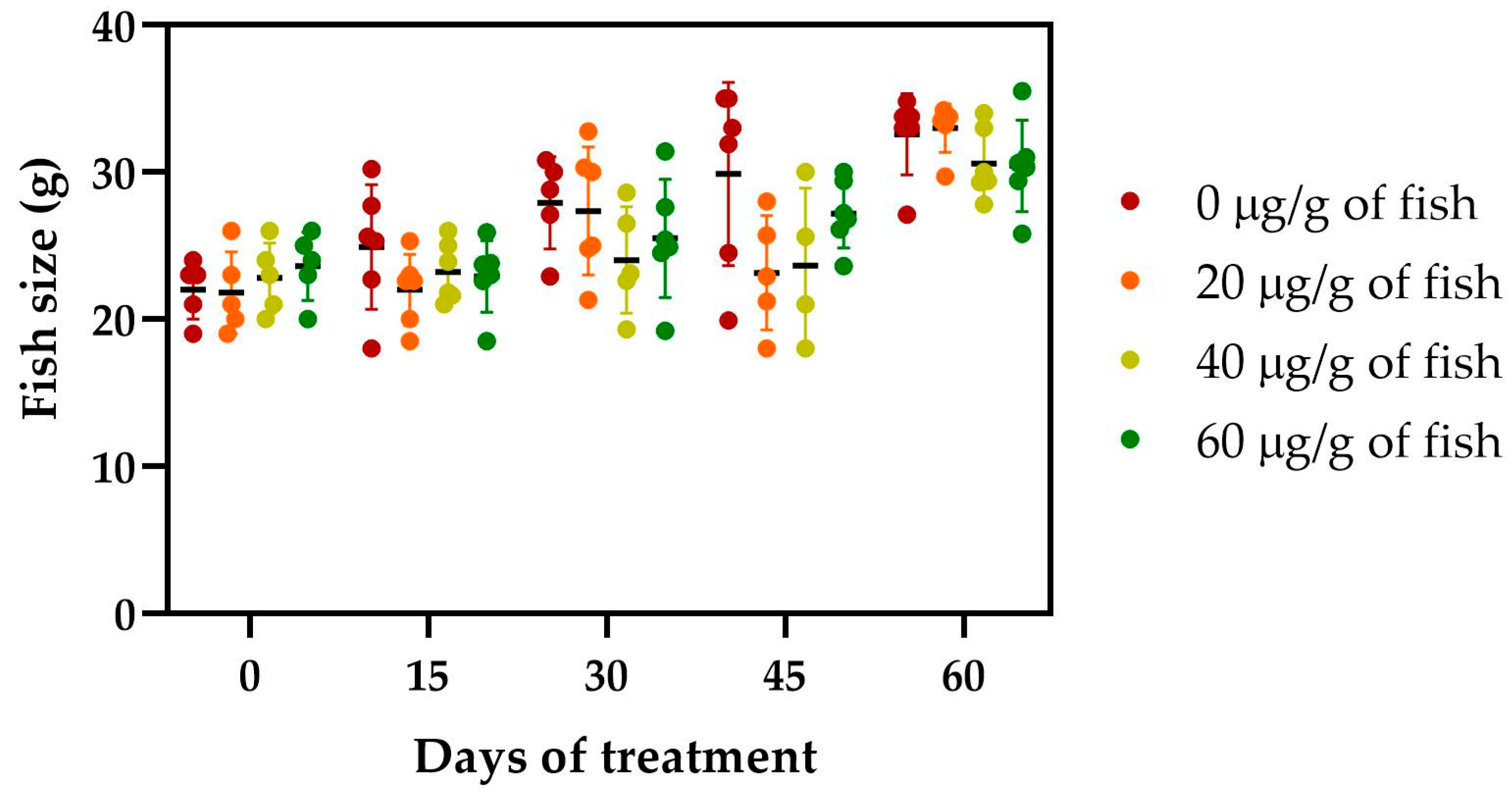
2.3. Effect of [Cu(NN1)2]ClO4 on the Growth of Atlantic Salmon
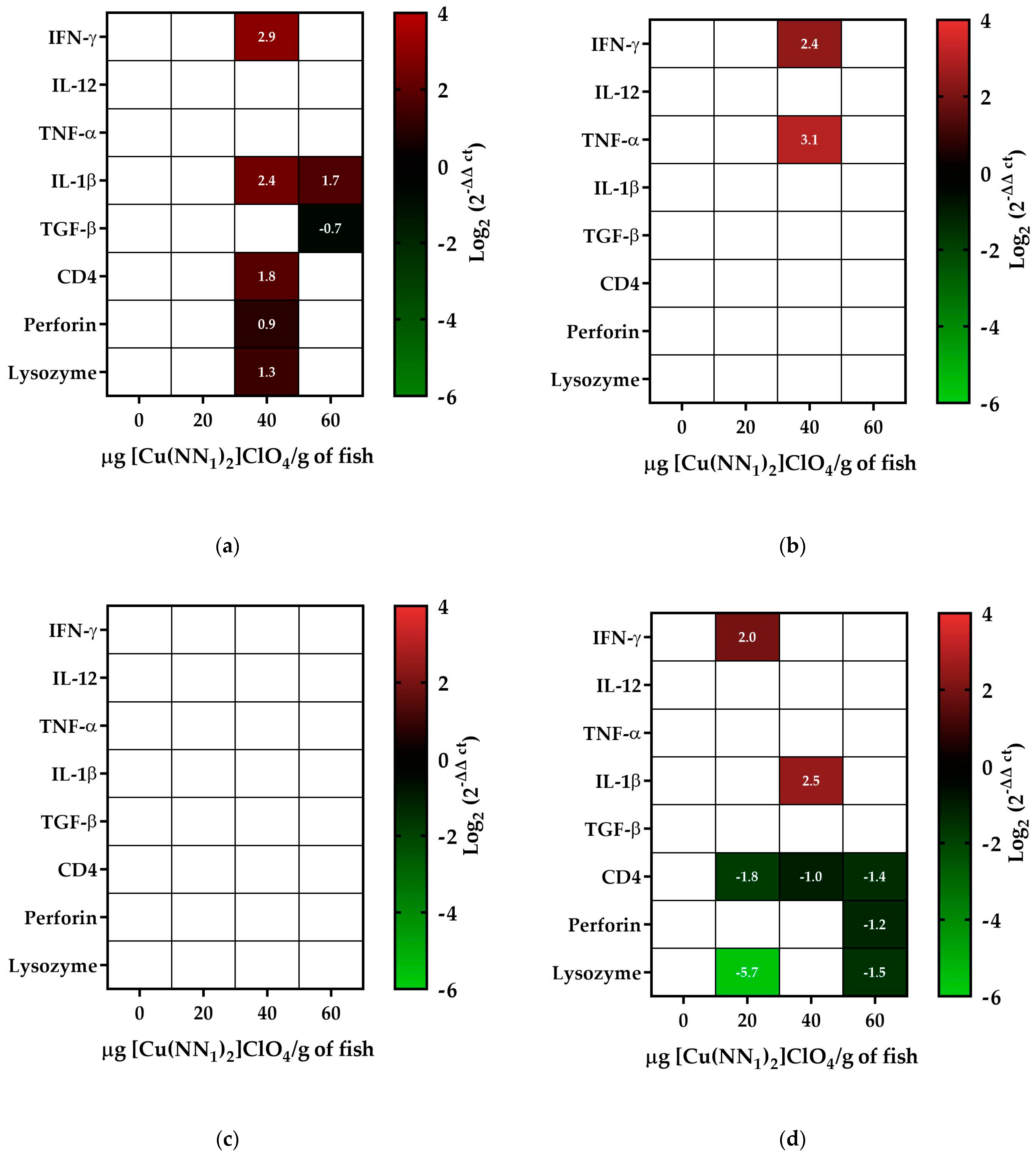
2.4. Evaluation of the Administration of [Cu(NN1)2]ClO4 on Immune Status of Atlantic Salmon
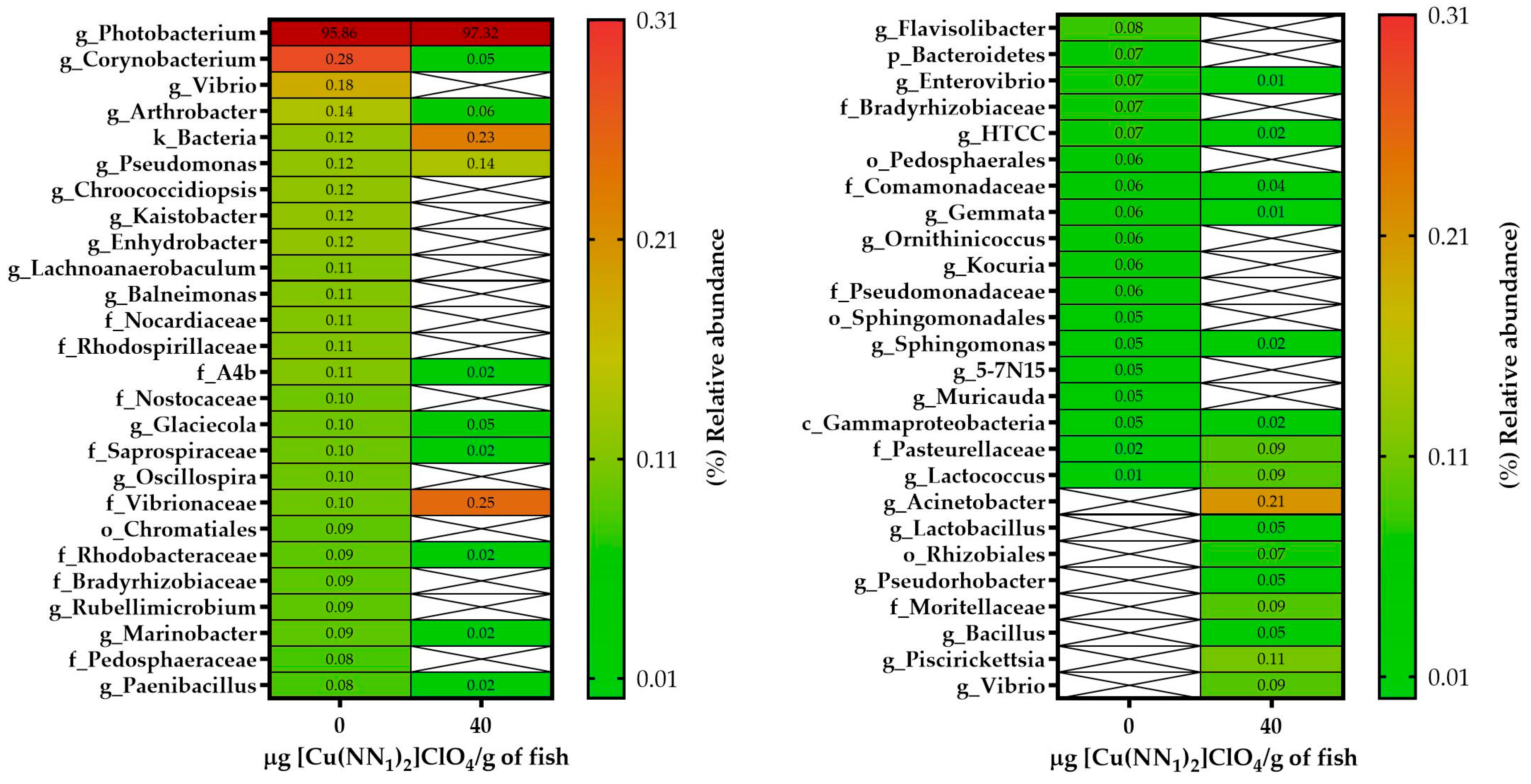
2.5. Effect of the Administration of [Cu(NN1)2]ClO4 on the Composition of the Intestinal Microbiota of Atlantic Salmon
2.6. Evaluation of the Protective Effect of [Cu(NN1)2]ClO4 against a Challenge with P. salmonis
3. Discussion
4. Materials and Methods
4.1. Synthesis of Cu (I) Coordination Complex
4.2. Fish and Maintenance
4.3. Antibacterial Activity
4.4. Evaluation of the Immune Response and Modulation of the Intestinal Microbiota
4.5. Growth of P. salmonis in Cell Culture
4.6. Evaluation of the Protective Effect of [Cu(NN1)2]ClO4
4.7. RNA Extraction
4.8. DNA Extraction
4.9. Real-Time PCR
4.10. 16 S Ribosomal Sequencing
4.11. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- SERNAPESCA. Informe Sanitario Con Información Sanitaria de Agua Dulce y Mar Año 2022; Informe Sanitario; Ministerio de Economía, Fomento y Turismo: Valparaíso, Chile, 2023. [Google Scholar]
- Mohapatra, S.; Chakraborty, T.; Kumar, V.; Deboeck, G.; Mohanta, K.N. Aquaculture and Stress Management: A Review of Probiotic Intervention. J. Anim. Physiol. Anim. Nutr. 2013, 97, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Maisey, K.; Montero, R.; Christodoulides, M. Vaccines for Piscirickettsiosis (Salmonid Rickettsial Septicaemia, SRS): The Chile Perspective. Expert. Rev. Vaccines 2017, 16, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Rozas, M.; Enríquez, R. Piscirickettsiosis and Piscirickettsia salmonis in Fish: A Review. J. Fish. Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef]
- Yañez, A.J.; Valenzuela, K.; Silva, H.; Retamales, J.; Romero, A.; Enriquez, R.; Figueroa, J.; Claude, A.; Gonzalez, J.; Avendaño-Herrera, R.; et al. Broth Medium for the Successful Culture of the Fish Pathogen Piscirickettsia salmonis. Dis. Aquat. Organ. 2012, 97, 197–205. [Google Scholar] [CrossRef]
- Rojas, V.; Galanti, N.; Bols, N.C.; Jiménez, V.; Paredes, R.; Marshall, S.H. Piscirickettsia salmonis Induces Apoptosis in Macrophages and Monocyte-like Cells from Rainbow Trout. J. Cell Biochem. 2010, 110, 468–476. [Google Scholar] [CrossRef]
- Gómez, F.A.; Tobar, J.A.; Henríquez, V.; Sola, M.; Altamirano, C.; Marshall, S.H. Evidence of the Presence of a Functional Dot/Icm Type IV-B Secretion System in the Fish Bacterial Pathogen Piscirickettsia salmonis. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rozas-Serri, M.; Peña, A.; Arriagada, G.; Enríquez, R.; Maldonado, L. Comparison of Gene Expression in Post-Smolt Atlantic Salmon Challenged by LF-89-like and EM-90-like Piscirickettsia salmonis Isolates Reveals Differences in the Immune Response Associated with Pathogenicity. J. Fish. Dis. 2018, 41, 539–552. [Google Scholar] [CrossRef]
- Rozas-Serri, M.; Peña, A.; Maldonado, L. Transcriptomic Profiles of Post-Smolt Atlantic Salmon Challenged with Piscirickettsia salmonis Reveal a Strategy to Evade the Adaptive Immune Response and Modify Cell-Autonomous Immunity. Dev. Comp. Immunol. 2018, 81, 348–362. [Google Scholar] [CrossRef]
- Almendras, F.E.; Fuentealba, I.C. Salmonid Rickettsial Septicemia Caused by Piscirickettsia salmonis: A Review. Dis. Aquat. Organ. 1997, 29, 137–144. [Google Scholar] [CrossRef]
- Cvitanich, J.D. The Isolation of a Rickettsia-like Organism Causing Disease and Mortality in Chilean Salmonids and Its Confirmation by Koch’s Postulate. J. Fish. Dis. 1991, 14, 2–45. [Google Scholar] [CrossRef]
- Figueroa, C.; Torrealba, D.; Morales-Lange, B.; Mercado, L.; Dixon, B.; Conejeros, P.; Silva, G.; Soto, C.; Gallardo, J.A. Commercial Vaccines Do Not Confer Protection against Two Genogroups of Piscirickettsia salmonis, LF-89 and EM-90, in Atlantic Salmon. Biology 2022, 11, 993. [Google Scholar] [CrossRef]
- SERNAPESCA. Informe Sobre Uso de Antimicrobianos En La Salmonicultura Nacional; Informe Sanitario; Ministerio de Economía, Fomento y Turismo: Valparaíso, Chile, 2023. [Google Scholar]
- Henríquez, P.; Kaiser, M.; Bohle, H.; Bustos, P.; Mancilla, M. Comprehensive Antibiotic Susceptibility Profiling of Chilean Piscirickettsia salmonis Field Isolates. J. Fish. Dis. 2016, 39, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, J.; Hernandez, N.; Osses, A.; Castillo, A.; Cancino, A.; Grothusen, H.; Navas, E.; Henriquez, P.; Bohle, H.; Bustamante, F.; et al. Prevalence, Geographic Distribution and Phenotypic Differences of Piscirickettsia salmonis EM-90-like Isolates. J. Fish. Dis. 2017, 40, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Habibullah-Al-Mamun, M.; Nagano, I.; Masunaga, S.; Kitazawa, D.; Matsuda, H. Antibiotics, Antibiotic-Resistant Bacteria, and Resistance Genes in Aquaculture: Risks, Current Concern, and Future Thinking. Environ. Sci. Pollut. Res. 2022, 29, 11054–11075. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, R.A.; Fuentes, M.; Montes, R.M.; Soto, D.; León-Muñoz, J. Environmental Issues in Chilean Salmon Farming: A Review. Rev. Aquac. 2019, 11, 375–402. [Google Scholar] [CrossRef]
- Donati, V.L.; Madsen, L.; Middelboe, M.; Strube, M.L.; Dalsgaard, I. The Gut Microbiota of Healthy and Flavobacterium psychrophilum-Infected Rainbow Trout Fry Is Shaped by Antibiotics and Phage Therapies. Front. Microbiol. 2022, 13, 771296. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Jarau, M.; MacInnes, J.; Stevenson, R.; Lumsden, J. Impact of Infection with Flavobacterium psychrophilum and Antimicrobial Treatment on the Intestinal Microbiota of Rainbow Trout. Pathogens 2023, 12, 454. [Google Scholar] [CrossRef]
- Xavier, R.; Severino, R.; Silva, S.M. Signatures of Dysbiosis in Fish Microbiomes in the Context of Aquaculture. Rev. Aquac. 2023, 16, 706–731. [Google Scholar] [CrossRef]
- Hu, Y.; Shen, Y.; Wu, X.; Tu, X.; Wang, G.X. Synthesis and Biological Evaluation of Coumarin Derivatives Containing Imidazole Skeleton as Potential Antibacterial Agents. Eur. J. Med. Chem. 2018, 143, 958–969. [Google Scholar] [CrossRef]
- Liu, G.; Wang, C.; Wang, H.; Zhu, L.; Zhang, H.; Wang, Y.; Pei, C.; Liu, L. Antiviral Efficiency of a Coumarin Derivative on Spring Viremia of Carp Virus In Vivo. Virus Res. 2019, 268, 11–17. [Google Scholar] [CrossRef]
- Liu, G.L.; Hu, Y.; Chen, X.H.; Wang, G.X.; Ling, F. Synthesis and Anthelmintic Activity of Coumarin–Imidazole Hybrid Derivatives against Dactylogyrus intermedius in Goldfish. Bioorg Med. Chem. Lett. 2016, 26, 5039–5043. [Google Scholar] [CrossRef]
- Krasnovskaya, O.; Naumov, A.; Guk, D.; Gorelkin, P.; Erofeev, A.; Beloglazkina, E.; Majouga, A. Copper Coordination Compounds as Biologically Active Agents. Int. J. Mol. Sci. 2020, 21, 3965. [Google Scholar] [CrossRef] [PubMed]
- Djoko, K.Y.; S Achard, M.E.; Phan, M.-D.; Lo, A.W.; Miraula, M.; Prombhul, S.; Hancock, S.J.; Peters, K.M.; Sidjabat, H.E.; Harris, P.N.; et al. Copper Ions and Coordination Complexes as Novel Carbapenem Adjuvants. Antimicrob. Agents Chemother. 2018, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, J.; Xu, L.; Wang, B.; Lin, W.; Luo, Y. Copper Regulation of Immune Response and Potential Implications for Treating Orthopedic Disorders. Front. Mol. Biosci. 2022, 9, 1065265. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O. Dietary Copper Requirements for Aquatic Animals: A Review. Biol. Trace Elem. Res. 2022, 200, 5273–5282. [Google Scholar] [CrossRef] [PubMed]
- Schrader, K.; Harries, M.D.; Darwish, A.M. In vitro comparisons of the inhibitory activity of florfenicol copper sulfate and potassium permanganate towards Aeromonas hydrophila and Flavobacterium columnare. Aquac. Res. 2013, 44, 212–219. [Google Scholar] [CrossRef]
- Cáceres-Vásquez, J.; Jara, D.H.; Costamagna, J.; Martínez-Gómez, F.; Silva, C.P.; Lemus, L.; Freire, E.; Baggio, R.; Vera, C.; Guerrero, J. Effect of Non-Covalent Self-Dimerization on the Spectroscopic and Electrochemical Properties of Mixed Cu(i) Complexes. RSC Adv. 2023, 13, 825–838. [Google Scholar] [CrossRef]
- Aldabaldetrecu, M.; Parra, M.; Soto, S.; Arce, P.; Tello, M.; Guerrero, J.; Modak, B. New Copper(I) Complex with a Coumarin as Ligand with Antibacterial Activity against Flavobacterium psychrophilum. Molecules 2020, 25, 3183. [Google Scholar] [CrossRef]
- Aldabaldetrecu, M.; Parra, M.; Soto-Aguilera, S.; Arce, P.; de la Vega Quiroz, A.P.; Segura, R.; Tello, M.; Guerrero, J.; Modak, B. Protective Effect of [Cu(NN1)2](ClO4) Complex in Rainbow Trout Challenged against Flavobacterium psychrophilum. Microorganisms 2022, 10, 2296. [Google Scholar] [CrossRef]
- Figueroa, J.; Castro, D.; Lagos, F.; Cartes, C.; Isla, A.; Yáñez, A.J.; Avendaño-Herrera, R.; Haussmann, D. Analysis of Single Nucleotide Polymorphisms (SNPs) Associated with Antibiotic Resistance Genes in Chilean Piscirickettsia salmonis Strains. J. Fish. Dis. 2019, 42, 1645–1655. [Google Scholar] [CrossRef]
- Tandberg, J.I.; Lagos, L.X.; Langlete, P.; Berger, E.; Rishovd, A.L.; Roos, N.; Varkey, D.; Paulsen, I.T.; Winther-Larsen, H.C. Comparative Analysis of Membrane Vesicles from Three Piscirickettsia salmonis Isolates Reveals Differences in Vesicle Characteristics. PLoS ONE 2016, 11, e0165099. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.J.; Romero, A.; Gonzalez-Stegmaier, R.; Dantagnan, P. The Effects of Supplemented Diets with a Phytopharmaceutical Preparation from Herbal and Macroalgal Origin on Disease Resistance in Rainbow Trout against Piscirickettsia salmonis. Aquaculture 2016, 454, 109–117. [Google Scholar] [CrossRef]
- Romero, A.; Pérez, T.; Santibáñez, N.; Vega, M.; Miranda, P. Phytogenic Feed Additive (PFA) Standardized in Labdane Diterpens Have a Protective Effect in Salmo salar against Piscirickettsia salmonis. Aquaculture 2021, 533, 736170. [Google Scholar] [CrossRef]
- Lichtenheld, M.G.; Olsen, K.J.; Lu, P.; Lowrey, D.M.; Hameed, A.; Hengartner, H.; Podack, E.R. Structure and Function of Human Perforin. Nature 1988, 335, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.H.; Jung, S.J. CpG ODN 1668 Induce Innate and Adaptive Immune Responses in Rock Bream (Oplegnathus fasciatus) against Rock Bream Iridovirus (RBIV) Infection. Fish Shellfish Immunol. 2017, 69, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Samsing, F.; Alexandre, P.; Rigby, M.; Taylor, R.S.; Chong, R.; Wynne, J.W. Transcriptome Response of Atlantic Salmon (Salmo salar) to a New Piscine Orthomyxovirus. Pathogens 2020, 9, 807. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.; Baranski, M.; Mahapatra, K.D.; Saha, J.N.; Das, S.; Mishra, J.; Das, P.; Kent, M.; Arnyasi, M.; Sahoo, P.K. A Linkage Map of Transcribed Single Nucleotide Polymorphisms in Rohu (Labeo rohita) and QTL Associated with Resistance to Aeromonas hydrophila. BMC Genom. 2014, 15, 541. [Google Scholar] [CrossRef]
- Li, T.; Wang, L.; Zhang, Y.; Guo, X.; Chen, X.; Zhang, F.; Yang, G.; Wen, W.; Li, H. Molecular Characterization of Three Novel Perforins in Common Carp (Cyprinus carpio L.) and Their Expression Patterns during Larvae Ontogeny and in Response to Immune Challenges. BMC Vet. Res. 2018, 14, 299. [Google Scholar] [CrossRef]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An Important Defence Molecule of Fish Innate Immune System. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Song, Q.; Xiao, Y.; Xiao, Z.; Liu, T.; Li, J.; Li, P.; Han, F. Lysozymes in Fish. J. Agric. Food Chem. 2021, 69, 15039–15051. [Google Scholar] [CrossRef]
- Buonocore, F.; Randelli, E.; Trisolino, P.; Facchiano, A.; de Pascale, D.; Scapigliati, G. Molecular Characterization, Gene Structure and Antibacterial Activity of a g-Type Lysozyme from the European Sea Bass (Dicentrarchus labrax L.). Mol. Immunol. 2014, 62, 10–18. [Google Scholar] [CrossRef]
- Wei, S.; Huang, Y.; Huang, X.; Cai, J.; Wei, J.; Li, P.; Ouyang, Z.; Qin, Q. Molecular Cloning and Characterization of a New G-Type Lysozyme Gene (Ec-LysG) in Orange-Spotted Grouper, Epinephelus coioides. Dev. Comp. Immunol. 2014, 46, 401–412. [Google Scholar] [CrossRef]
- Jahazi, M.A.; Hoseinifar, S.H.; Jafari, V.; Hajimoradloo, A.; Van Doan, H.; Paolucci, M. Dietary Supplementation of Polyphenols Positively Affects the Innate Immune Response, Oxidative Status, and Growth Performance of Common Carp, Cyprinus carpio L. Aquaculture 2020, 517, 734709. [Google Scholar] [CrossRef]
- Ahmadi, A.; Bagheri, D.; Hoseinifar, S.H.; Morshedi, V.; Paolucci, M. Beneficial Role of Polyphenols as Feed Additives on Growth Performances, Immune Response and Antioxidant Status of Lates calcarifer (Bloch, 1790) Juveniles. Aquaculture 2022, 552, 737955. [Google Scholar] [CrossRef]
- Li, M.; Zhu, X.; Tian, J.; Liu, M.; Wang, G. Dietary Flavonoids from Allium mongolicum Regel Promotes Growth, Improves Immune, Antioxidant Status, Immune-Related Signaling Molecules and Disease Resistance in Juvenile Northern Snakehead Fish (Channa argus). Aquaculture 2019, 501, 473–481. [Google Scholar] [CrossRef]
- Boehm, U.; Klamp, T.; Groot, M.; Howard, J.C. 40 Annual Reviews AR026-28 AR26-28. Annu. Rev. Immunol. 1997, 8, 749–795. [Google Scholar] [CrossRef] [PubMed]
- Green, D.S.; Young, H.A.; Valencia, J.C. Current Prospects of Type II Interferon γ Signaling & Autoimmunity. J. Biol. Chem. 2017, 292, 13925–13933. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage Plasticity and Polarization: In Vivo Veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Pereiro, P.; Figueras, A.; Novoa, B. Insights into Teleost Interferon-Gamma Biology: An Update. Fish. Shellfish. Immunol. 2019, 90, 150–164. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An Overview of Signals, Mechanisms and Functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Yilmaz, S. Effects of Dietary Caffeic Acid Supplement on Antioxidant, Immunological and Liver Gene Expression Responses, and Resistance of Nile Tilapia, Oreochromis niloticus to Aeromonas veronii. Fish. Shellfish. Immunol. 2019, 86, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Metwally, A.E.S.; El-Sharawy, M.E.; Ghozlan, A.M.; Abdel-Latif, H.M.R.; Van Doan, H.; Ali, M.A.M. The Influences of Ferulic Acid on the Growth Performance, Haemato-Immunological Responses, and Immune-Related Genes of Nile Tilapia (Oreochromis niloticus) Exposed to Heat Stress. Aquaculture 2020, 525, 735320. [Google Scholar] [CrossRef]
- Yılmaz, S.; Ergün, S. Trans-Cinnamic Acid Application for Rainbow Trout (Oncorhynchus mykiss): I. Effects on Haematological, Serum Biochemical, Non-Specific Immune and Head Kidney Gene Expression Responses. Fish. Shellfish. Immunol. 2018, 78, 140–157. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.F.; Liu, L.; Feng, C.Z.; Hu, Y.; Chen, C.; Wang, G.X.; Zhu, B. Synthesis and Antiviral Activity of a New Coumarin Derivative against Spring Viraemia of Carp Virus. Fish. Shellfish. Immunol. 2018, 81, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.S.; Shin, N.R.; Lee, J.B.; Kim, M.S.; Whon, T.W.; Hyun, D.W.; Yun, J.H.; Jung, M.J.; Kim, J.Y.; Bae, J.W. Host Habitat Is the Major Determinant of the Gut Microbiome of Fish. Microbiome 2021, 9, 166. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jaramillo-Torres, A.; Li, Y.; Kortner, T.M.; Gajardo, K.; Brevik, Ø.J.; Jakobsen, J.V.; Krogdahl, Å. Microbiota in Intestinal Digesta of Atlantic Salmon (Salmo salar), Observed from Late Freshwater Stage until One Year in Seawater, and Effects of Functional Ingredients: A Case Study from a Commercial Sized Research Site in the Arctic Region. Anim. Microbiome 2021, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y.; Hlordzi, V.; Sakyi, M.E.; Afriyie, G.; Wang, Z.; Li, Y.; Xie, C.X. Mechanisms and the Role of Probiotic Bacillus in Mitigating Fish Pathogens in Aquaculture. Fish. Physiol. Biochem. 2020, 46, 819–841. [Google Scholar] [CrossRef]
- Soltani, M.; Pakzad, K.; Taheri-Mirghaed, A.; Mirzargar, S.; Shekarabi, S.P.H.; Yosefi, P.; Soleymani, N. Dietary Application of the Probiotic Lactobacillus plantarum 426951 Enhances Immune Status and Growth of Rainbow Trout (Oncorhynchus mykiss) Vaccinated Against Yersinia ruckeri. Probiotics Antimicrob. Proteins 2019, 11, 207–219. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, Z.; Zhao, F.; Liu, H.; Yu, L.; Zha, J.; Wang, G. Probiotic Potential of Bacillus velezensis JW: Antimicrobial Activity against Fish Pathogenic Bacteria and Immune Enhancement Effects on Carassius auratus. Fish. Shellfish. Immunol. 2018, 78, 322–330. [Google Scholar] [CrossRef]
- Xia, Y.; Lu, M.; Chen, G.; Cao, J.; Gao, F.; Wang, M.; Liu, Z.; Zhang, D.; Zhu, H.; Yi, M. Effects of Dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis Subsp. Lactis JCM5805 on the Growth, Intestinal Microbiota, Morphology, Immune Response and Disease Resistance of Juvenile Nile Tilapia, Oreochromis niloticus. Fish. Shellfish. Immunol. 2018, 76, 368–379. [Google Scholar] [CrossRef]
- Baumgärtner, S.; James, J.; Ellison, A. The Supplementation of a Prebiotic Improves the Microbial Community in the Gut and the Skin of Atlantic Salmon (Salmo salar). Aquac. Rep. 2022, 25, 101204. [Google Scholar] [CrossRef]
- Ou, W.; Hu, H.; Yang, P.; Dai, J.; Ai, Q.; Zhang, W.; Zhang, Y.; Mai, K. Dietary Daidzein Improved Intestinal Health of Juvenile Turbot in Terms of Intestinal Mucosal Barrier Function and Intestinal Microbiota. Fish. Shellfish. Immunol. 2019, 94, 132–141. [Google Scholar] [CrossRef]
- Zheng, Y.; Wu, W.; Hu, G.; Qiu, L.; Meng, S.; Song, C.; Fan, L.; Zhao, Z.; Bing, X.; Chen, J. Gut Microbiota Analysis of Juvenile Genetically Improved Farmed Tilapia (Oreochromis niloticus) by Dietary Supplementation of Different Resveratrol Concentrations. Fish. Shellfish. Immunol. 2018, 77, 200–207. [Google Scholar] [CrossRef]
- Coca, Y.; Godoy, M.; Pontigo, J.P.; Caro, D.; Maracaja-Coutinho, V.; Arias-Carrasco, R.; Rodríguez-Córdova, L.; de Oca, M.M.; Sáez-Navarrete, C.; Burbulis, I. Bacterial Networks in Atlantic Salmon with Piscirickettsiosis. Sci. Rep. 2023, 13, 17321. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.P. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standars Institute: Wyne, NJ, USA, 2020; pp. 1–332. ISBN 9781684400324. [Google Scholar]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, 2001–2007. [Google Scholar] [CrossRef] [PubMed]
- Vargas, R.A.; Soto-Aguilera, S.; Parra, M.; Herrera, S.; Santibañez, A.; Kossack, C.; Saavedra, C.P.; Mora, O.; Pineda, M.; Gonzalez, O.; et al. Analysis of Microbiota-Host Communication Mediated by Butyrate in Atlantic Salmon. Comput. Struct. Biotechnol. J. 2023, 21, 2558–2578. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S RRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolated | MIC µg/mL | IC50 µg/mL |
|---|---|---|
| CGRO2 | 29.0 ± 2.5 | 14.6 ± 5.5 |
| 12201 | 33.7 ± 2.4 | 12.6 ± 1.3 |
| 8148 | 15.0 ± 7.1 | 5.3 ± 0.8 |
| 727 | 33.9 ± 14.5 | 11.0 ± 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra, M.; Aldabaldetrecu, M.; Arce, P.; Soto-Aguilera, S.; Vargas, R.; Guerrero, J.; Tello, M.; Modak, B. [Cu(NN1)2]ClO4, a Copper (I) Complex as an Antimicrobial Agent for the Treatment of Piscirickettsiosis in Atlantic Salmon. Int. J. Mol. Sci. 2024, 25, 3700. https://doi.org/10.3390/ijms25073700
Parra M, Aldabaldetrecu M, Arce P, Soto-Aguilera S, Vargas R, Guerrero J, Tello M, Modak B. [Cu(NN1)2]ClO4, a Copper (I) Complex as an Antimicrobial Agent for the Treatment of Piscirickettsiosis in Atlantic Salmon. International Journal of Molecular Sciences. 2024; 25(7):3700. https://doi.org/10.3390/ijms25073700
Chicago/Turabian StyleParra, Mick, Maialen Aldabaldetrecu, Pablo Arce, Sarita Soto-Aguilera, Rodrigo Vargas, Juan Guerrero, Mario Tello, and Brenda Modak. 2024. "[Cu(NN1)2]ClO4, a Copper (I) Complex as an Antimicrobial Agent for the Treatment of Piscirickettsiosis in Atlantic Salmon" International Journal of Molecular Sciences 25, no. 7: 3700. https://doi.org/10.3390/ijms25073700
APA StyleParra, M., Aldabaldetrecu, M., Arce, P., Soto-Aguilera, S., Vargas, R., Guerrero, J., Tello, M., & Modak, B. (2024). [Cu(NN1)2]ClO4, a Copper (I) Complex as an Antimicrobial Agent for the Treatment of Piscirickettsiosis in Atlantic Salmon. International Journal of Molecular Sciences, 25(7), 3700. https://doi.org/10.3390/ijms25073700