
Robust Fluorometric Aptamer Assay for Direct and Rapid Detection of Clinical Isolates of Candida spec.

 ,
,  ,
,
Abstract
1. Introduction
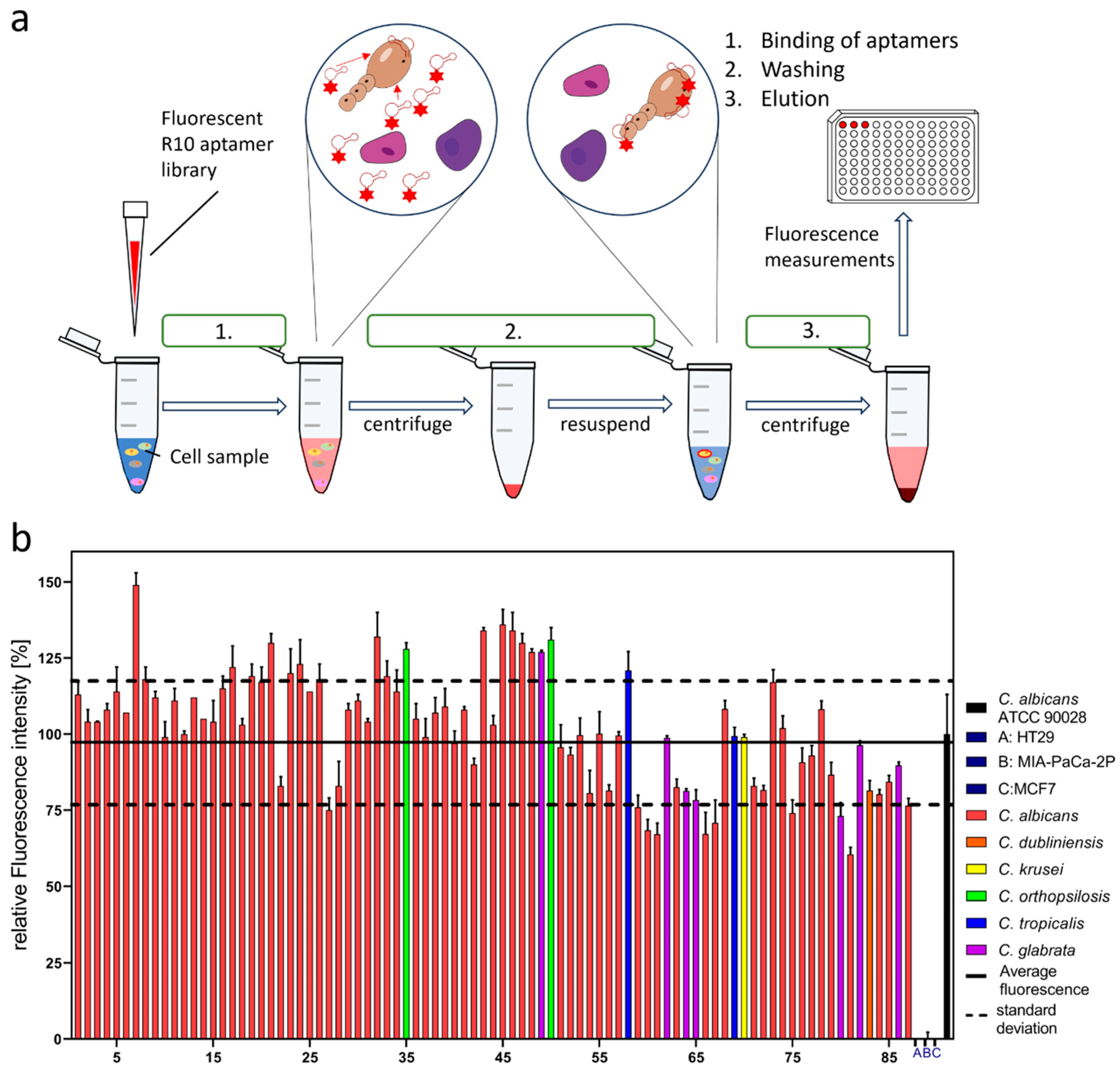
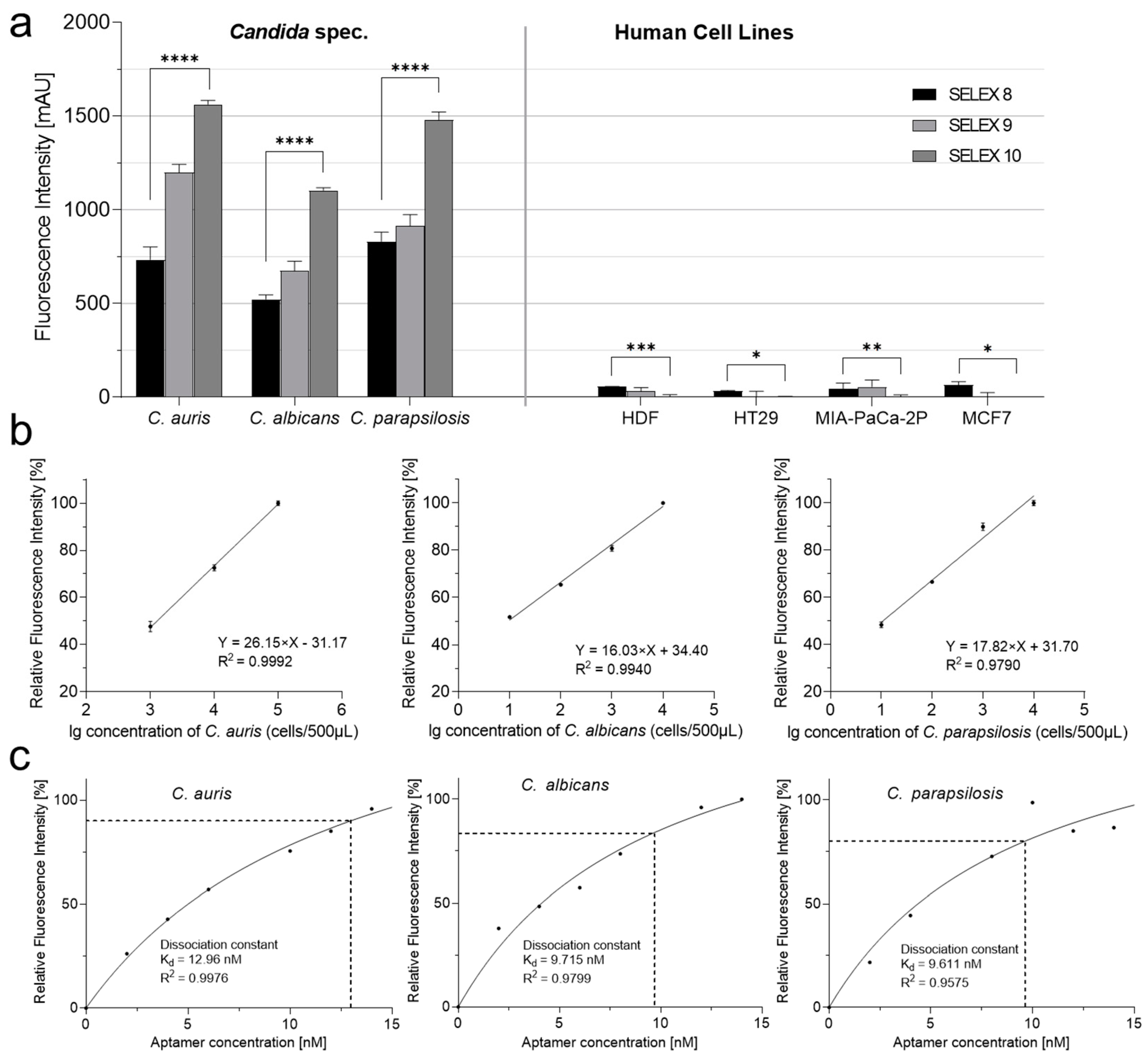
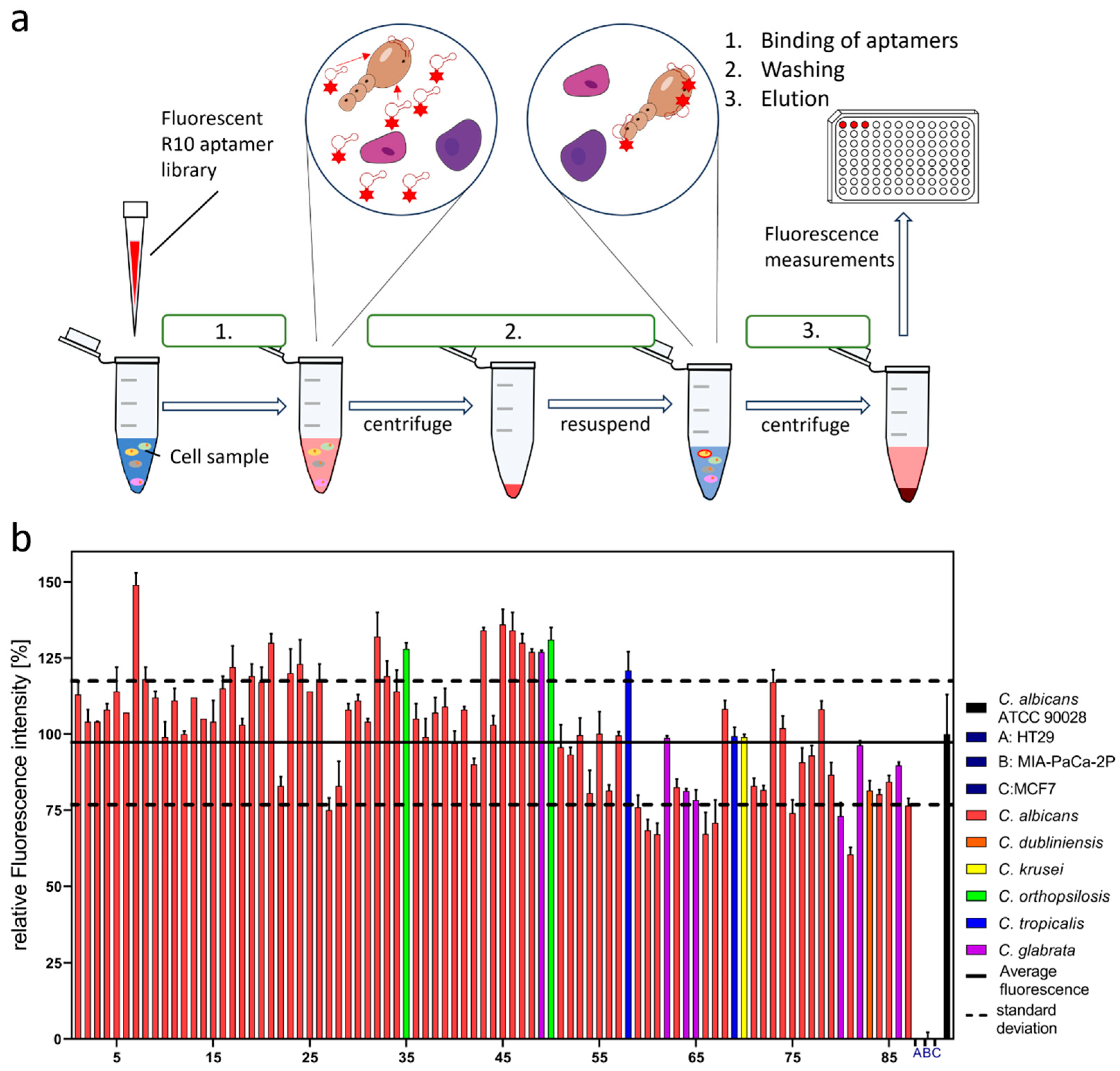
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Cell Pretreatment
4.3. Cell SELEX
4.3.1. Pretreatment before Counter SELEX
4.3.2. Pretreatment before Target SELEX
4.3.3. Aptamer Activation
4.3.4. Screening
4.3.5. Elution
4.3.6. Library Amplification
4.3.7. Binding Assay
Candida Binding Assay
Cell Binding Assay
4.4. Affinity Analysis
4.5. Sensitivity Test
4.6. Detection of Clinic Isolates
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nobile, C.J.; Johnson, A.D. Candida Albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and Emerged Pathogenic Candida Species: Beyond the Candida Albicans Paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef]
- Fakhim, H.; Vaezi, A.; Dannaoui, E.; Chowdhary, A.; Nasiry, D.; Faeli, L.; Meis, J.F.; Badali, H. Comparative Virulence of Candida Auris with Candida Haemulonii, Candida Glabrata and Candida Albicans in a Murine Model. Mycoses 2018, 61, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R. Current Epidemiology of Candida Infection. Clin. Microbiol. Newsl. 2014, 36, 131–136. [Google Scholar] [CrossRef]
- McCarty, T.P.; Pappas, P.G. Invasive Candidiasis. Infect. Dis. Clin. N. Am. 2016, 30, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Perlroth, J.; Choi, B.; Spellberg, B. Nosocomial Fungal Infections: Epidemiology, Diagnosis, and Treatment. Med. Mycol. 2007, 45, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections. Methods Mol. Biol. 2017, 1508, 17–65. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Gaitán, A.C.; Moret, A.; López Hontangas, J.L.; Molina, J.M.; Aleixandre López, A.I.; Cabezas, A.H.; Mollar Maseres, J.; Arcas, R.C.; Gómez Ruiz, M.D.; Chiveli, M.Á.; et al. Nosocomial Fungemia by Candida Auris: First Four Reported Cases in Continental Europe. Rev. Iberoam. Micol. 2017, 34, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Sobel, J.D.; Faro, S.; Force, R.W.; Foxman, B.; Ledger, W.J.; Nyirjesy, P.R.; Reed, B.D.; Summers, P.R. Vulvovaginal Candidiasis: Epidemiologic, Diagnostic, and Therapeutic Considerations. Am. J. Obstet. Gynecol. 1998, 178, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global Burden of Recurrent Vulvovaginal Candidiasis: A Systematic Review. Lancet Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive Candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of Invasive Candidiasis: A Persistent Public Health Problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed]
- Harrington, R.; Kindermann, S.L.; Hou, Q.; Taylor, R.J.; Azie, N.; Horn, D.L. Candidemia and Invasive Candidiasis among Hospitalized Neonates and Pediatric Patients. Curr. Med. Res. Opin. 2017, 33, 1803–1812. [Google Scholar] [CrossRef] [PubMed]
- Quindós, G.; Marcos-Arias, C.; San-Millán, R.; Mateo, E.; Eraso, E. The Continuous Changes in the Aetiology and Epidemiology of Invasive Candidiasis: From Familiar Candida Albicans to Multiresistant Candida Auris. Int. Microbiol. 2018, 21, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Xiao, M.; Liao, K.; Kudinha, T.; Wang, H.; Zhang, L.; Hou, X.; Kong, F.; Xu, Y.-C. Notable Increasing Trend in Azole Non-Susceptible Candida Tropicalis Causing Invasive Candidiasis in China (August 2009 to July 2014): Molecular Epidemiology and Clinical Azole Consumption. Front. Microbiol. 2017, 8, 464. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Preston, T.; Bale, M.; Koontz, F.P.; Body, B.A. Comparison of the Quantum II, API Yeast Ident, and AutoMicrobic Systems for Identification of Clinical Yeast Isolates. J. Clin. Microbiol. 1988, 26, 2054–2058. [Google Scholar] [CrossRef]
- Shankland, G.S.; Hopwood, V.; Forster, R.A.; Evans, E.G.; Richardson, M.D.; Warnock, D.W. Multicenter Evaluation of Microring YT, a New Method of Yeast Identification. J. Clin. Microbiol. 1990, 28, 2808–2810. [Google Scholar] [CrossRef]
- Dhiman, N.; Hall, L.; Wohlfiel, S.L.; Buckwalter, S.P.; Wengenack, N.L. Performance and Cost Analysis of Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry for Routine Identification of Yeast. J. Clin. Microbiol. 2011, 49, 1614–1616. [Google Scholar] [CrossRef]
- Pinto, A.; Halliday, C.; Zahra, M.; van Hal, S.; Olma, T.; Maszewska, K.; Iredell, J.R.; Meyer, W.; Chen, S.C.-A. Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry Identification of Yeasts Is Contingent on Robust Reference Spectra. PLoS ONE 2011, 6, e25712. [Google Scholar] [CrossRef]
- Jamal, W.; Saleem, R.; Rotimi, V.O. Rapid Identification of Pathogens Directly from Blood Culture Bottles by Bruker Matrix-Assisted Laser Desorption Laser Ionization-Time of Flight Mass Spectrometry versus Routine Methods. Diagn. Microbiol. Infect. Dis. 2013, 76, 404–408. [Google Scholar] [CrossRef]
- Jamal, W.Y.; Ahmad, S.; Khan, Z.U.; Rotimi, V.O. Comparative Evaluation of Two Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) Systems for the Identification of Clinically Significant Yeasts. Int. J. Infect. Dis. 2014, 26, 167–170. [Google Scholar] [CrossRef]
- Stevenson, L.G.; Drake, S.K.; Shea, Y.R.; Zelazny, A.M.; Murray, P.R. Evaluation of Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry for Identification of Clinically Important Yeast Species. J. Clin. Microbiol. 2010, 48, 3482–3486. [Google Scholar] [CrossRef]
- Felix, G.N.; de Freitas, V.L.T.; da Silva Junior, A.R.; Magri, M.M.C.; Rossi, F.; Sejas, O.N.E.; Abdala, E.; Malbouisson, L.M.S.; Guimarães, T.; Benard, G.; et al. Performance of a Real-Time PCR Assay for the Detection of Five Candida Species in Blood Samples from ICU Patients at Risk of Candidemia. J. Fungi 2023, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Busser, F.D.; Coelho, V.C.; de Abreu Fonseca, C.; Del Negro, G.M.B.; Shikanai-Yasuda, M.A.; Lopes, M.H.; Magri, M.M.C.; de Freitas, V.L.T. A Real Time PCR Strategy for the Detection and Quantification of Candida Albicans in Human Blood. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e9. [Google Scholar] [CrossRef] [PubMed]
- Camp, I.; Spettel, K.; Willinger, B. Molecular Methods for the Diagnosis of Invasive Candidiasis. J. Fungi 2020, 6, 101. [Google Scholar] [CrossRef] [PubMed]
- Ilgu, M.; Nilsen-Hamilton, M. Aptamers in Analytics. Analyst 2016, 141, 1551–1568. [Google Scholar] [CrossRef] [PubMed]
- Keefe, A.D.; Pai, S.; Ellington, A. Aptamers as Therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Chopra, A.; Shukla, R.; Sharma, T.K. Aptamers as an Emerging Player in Biology. Aptamers Synth. Antibodies 2014, 1, 1–11. [Google Scholar]
- Ali, M.H.; Elsherbiny, M.E.; Emara, M. Updates on Aptamer Research. Int. J. Mol. Sci. 2019, 20, 2511. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Yeung, W.; Hili, R. In Vitro Selection of Diversely Functionalized Aptamers. J. Am. Chem. Soc. 2017, 139, 13977–13980. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Huang, R.; Deng, Y.; He, N. Progress in Selection and Biomedical Applications of Aptamers. J. Biomed. Nanotechnol. 2014, 10, 3043–3062. [Google Scholar] [CrossRef] [PubMed]
- Sefah, K.; Shangguan, D.; Xiong, X.; O’Donoghue, M.B.; Tan, W. Development of DNA Aptamers Using Cell-SELEX. Nat. Protoc. 2010, 5, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Rossi, J. Aptamers as Targeted Therapeutics: Current Potential and Challenges. Nat. Rev. Drug Discov. 2017, 16, 440. [Google Scholar] [CrossRef]
- Ellington, A.D.; Szostak, J.W. In Vitro Selection of RNA Molecules That Bind Specific Ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic Evolution of Ligands by Exponential Enrichment: RNA Ligands to Bacteriophage T4 DNA Polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef]
- Shaban, S.M.; Kim, D.-H. Recent Advances in Aptamer Sensors. Sensors 2021, 21, 979. [Google Scholar] [CrossRef]
- Ku, T.-H.; Zhang, T.; Luo, H.; Yen, T.M.; Chen, P.-W.; Han, Y.; Lo, Y.-H. Nucleic Acid Aptamers: An Emerging Tool for Biotechnology and Biomedical Sensing. Sensors 2015, 15, 16281–16313. [Google Scholar] [CrossRef]
- Kusumawati, A.; Mustopa, A.Z.; Wibawan, I.W.T.; Setiyono, A.; Sudarwanto, M.B. A Sequential Toggle Cell-SELEX DNA Aptamer for Targeting Staphylococcus Aureus, Streptococcus Agalactiae, and Escherichia Coli Bacteria. J. Genet. Eng. Biotechnol. 2022, 20, 95. [Google Scholar] [CrossRef]
- Ansari, N.; Ghazvini, K.; Ramezani, M.; Shahdordizadeh, M.; Yazdian-Robati, R.; Abnous, K.; Taghdisi, S.M. Selection of DNA Aptamers against Mycobacterium Tuberculosis Ag85A, and Its Application in a Graphene Oxide-Based Fluorometric Assay. Microchim. Acta 2018, 185, 21. [Google Scholar] [CrossRef]
- Zhong, Z.; Gao, X.; Gao, R.; Jia, L. Selective Capture and Sensitive Fluorometric Determination of Pseudomonas Aeruginosa by Using Aptamer Modified Magnetic Nanoparticles. Microchim. Acta 2018, 185, 377. [Google Scholar] [CrossRef]
- Wu, Z.; He, D.; Cui, B. A Fluorometric Assay for Staphylococcal Enterotoxin B by Making Use of Platinum Coated Gold Nanorods and of Upconversion Nanoparticles. Microchim. Acta 2018, 185, 516. [Google Scholar] [CrossRef]
- Yuan, J.; Wu, S.; Duan, N.; Ma, X.; Xia, Y.; Chen, J.; Ding, Z.; Wang, Z. A Sensitive Gold Nanoparticle-Based Colorimetric Aptasensor for Staphylococcus Aureus. Talanta 2014, 127, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Meng, X.; Sun, Y.; Li, Q.; Zhao, R.; Zhang, Y.; Wang, J.; Yi, Z. Aptamer-Based Fluorometric Assay for Direct Identification of Methicillin-Resistant Staphylococcus Aureus from Clinical Samples. J. Microbiol. Methods 2018, 153, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.G.; Phillips, T.; Montez, T.; Garcia, A.; Sivils, J.C.; Mayo, M.W.; Greis, A. Development of a Fluorescent Enzyme-Linked DNA Aptamer-Magnetic Bead Sandwich Assay and Portable Fluorometer for Sensitive and Rapid Listeria Detection. J. Fluoresc. 2015, 25, 173–183. [Google Scholar] [CrossRef]
- Wu, W.; Li, J.; Pan, D.; Li, J.; Song, S.; Rong, M.; Li, Z.; Gao, J.; Lu, J. Gold Nanoparticle-Based Enzyme-Linked Antibody-Aptamer Sandwich Assay for Detection of Salmonella Typhimurium. ACS Appl. Mater. Interfaces 2014, 6, 16974–16981. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yue, F.; Kong, Q.; Liu, Z.; Guo, Y.; Sun, X. Aptamers against Pathogenic Bacteria: Selection Strategies and Apta-Assay/Aptasensor Application for Food Safety. J. Agric. Food Chem. 2022, 70, 5477–5498. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, L.; Lu, W.; Zhao, J.; Zhang, H.; Chen, W. Selection, Characterization and Interaction Studies of a DNA Aptamer for the Detection of Bifidobacterium Bifidum. Int. J. Mol. Sci. 2017, 18, 883. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Green, L.S.; Gold, L.; Janjic, N. Systematic Selection of Modified Aptamer Pairs for Diagnostic Sandwich Assays. Biotechniques 2014, 56, 125–133. [Google Scholar] [CrossRef]
- Kneißle, K.; Krämer, M.; Kissmann, A.-K.; Xing, H.; Müller, F.; Amann, V.; Noschka, R.; Gottschalk, K.-E.; Bozdogan, A.; Andersson, J.; et al. A Polyclonal SELEX Aptamer Library Allows Differentiation of Candida Albicans, C. Auris and C. Parapsilosis Cells from Human Dermal Fibroblasts. J. Fungi 2022, 8, 856. [Google Scholar] [CrossRef]
- Thevendran, R.; Navien, T.N.; Meng, X.; Wen, K.; Lin, Q.; Sarah, S.; Tang, T.-H.; Citartan, M. Mathematical Approaches in Estimating Aptamer-Target Binding Affinity. Anal. Biochem. 2020, 600, 113742. [Google Scholar] [CrossRef] [PubMed]
- Kubiczek, D.; Raber, H.; Bodenberger, N.; Oswald, T.; Sahan, M.; Mayer, D.; Wiese, S.; Stenger, S.; Weil, T.; Rosenau, F. The Diversity of a Polyclonal FluCell-SELEX Library Outperforms Individual Aptamers as Emerging Diagnostic Tools for the Identification of Carbapenem Resistant Pseudomonas Aeruginosa. Chem. A Eur. J. 2020, 26, 14536–14545. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Zhang, Y.; Krämer, M.; Kissmann, A.K.; Henkel, M.; Weil, T.; Knippschild, U.; Rosenau, F. A Polyclonal Selex Aptamer Library Directly Allows Specific Labelling of the Human Gut Bacterium Blautia Producta without Isolating Individual Aptamers. Molecules 2022, 27, 5693. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Kissmann, A.K.; Raber, H.F.; Krämer, M.; Amann, V.; Kohn, K.; Weil, T.; Rosenau, F. Polyclonal Aptamers for Specific Fluorescence Labeling and Quantification of the Health Relevant Human Gut Bacterium Parabacteroides Distasonis. Microorganisms 2021, 9, 2284. [Google Scholar] [CrossRef]
- Xing, H.; Zhang, Y.; Krämer, M.; Kissmann, A.K.; Amann, V.; Raber, H.F.; Weil, T.; Stieger, K.R.; Knippschild, U.; Henkel, M.; et al. A Polyclonal Aptamer Library for the Specific Binding of the Gut Bacterium Roseburia Intestinalis in Mixtures with Other Gut Microbiome Bacteria and Human Stool Samples. Int. J. Mol. Sci. 2022, 23, 7744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xing, H.; Bolotnikov, G.; Krämer, M.; Gotzmann, N.; Knippschild, U.; Kissmann, A.-K.; Rosenau, F. Enriched Aptamer Libraries in Fluorescence-Based Assays for Rikenella Microfusus-Specific Gut Microbiome Analyses. Microorganisms 2023, 11, 2266. [Google Scholar] [CrossRef] [PubMed]
- Raber, H.F.; Kubiczek, D.H.; Bodenberger, N.; Kissmann, A.K.; D’souza, D.; Hu, X.; Mayer, D.; Xu, P.; Knippschild, U.; Spellerberg, B.; et al. Flucell-selex Aptamers as Specific Binding Molecules for Diagnostics of the Health Relevant Gut Bacterium Akkermansia Muciniphila. Int. J. Mol. Sci. 2021, 22, 10425. [Google Scholar] [CrossRef]
- Kissmann, A.K.; Wolf, D.; Krämer, M.; Müller, F.; Amann, V.; Xing, H.; Gottschalk, K.E.; Weil, T.; Eichmann, R.; Schäfer, P.; et al. Polyclonal Aptamer Libraries from a FluRoot-SELEX for the Specific Labeling of the Apical and Elongation/Differentiation Zones of Arabidopsis Thaliana Roots. Int. J. Mol. Sci. 2022, 23, 12220. [Google Scholar] [CrossRef]
- Kissmann, A.-K.; Andersson, J.; Bozdogan, A.; Amann, V.; Kraemer, M.; Xing, H.; Raber, H.; Kubiczek, D.H.; Aspermair, P.; Knoll, W.; et al. Polyclonal Aptamer Libraries as Binding Entities on a Graphene FET Based Biosensor for the Discrimination of Apo- and Holo- Retinol Binding Protein 4. Nanoscale Horiz. 2022, 7, 770–778. [Google Scholar] [CrossRef]
- Wan, Q.; Liu, X.; Zu, Y. Oligonucleotide Aptamers for Pathogen Detection and Infectious Disease Control. Theranostics 2021, 11, 9133–9161. [Google Scholar] [CrossRef]
- Lee, S.-H.; Ahn, J.-Y.; Lee, K.-A.; Um, H.-J.; Sekhon, S.S.; Sun Park, T.; Min, J.; Kim, Y.-H. Analytical Bioconjugates, Aptamers, Enable Specific Quantitative Detection of Listeria Monocytogenes. Biosens. Bioelectron. 2015, 68, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Trunzo, N.E.; Hong, K.L. Recent Progress in the Identification of Aptamers Against Bacterial Origins and Their Diagnostic Applications. Int. J. Mol. Sci. 2020, 21, 5074. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, X.; Duan, N.; Wu, S.; Wang, Z.; Wei, X.; Wang, Y. Selection and Characterization of DNA Aptamers against Staphylococcus Aureus Enterotoxin C1. Food Chem. 2015, 166, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.-R.; Sekhon, S.S.; Kim, S.-G.; Rhee, S.J.; Yang, G.N.; Won, K.; Rhee, S.-K.; Ryu, H.; Kim, K.; Min, J.; et al. Aptamer-Based Pathogen Monitoring for Salmonella Enterica Ser. Typhimurium. J. Biomed. Nanotechnol. 2018, 14, 1992–2002. [Google Scholar] [CrossRef]
- Song, M.-S.; Sekhon, S.; Shin, W.-R.; Kim, H.; Min, J.; Ahn, J.-Y.; Kim, Y.-H. Detecting and Discriminating Shigella Sonnei Using an Aptamer-Based Fluorescent Biosensor Platform. Molecules 2017, 22, 825. [Google Scholar] [CrossRef]
- Chinnappan, R.; AlAmer, S.; Eissa, S.; Rahamn, A.A.; Abu Salah, K.M.; Zourob, M. Fluorometric Graphene Oxide-Based Detection of Salmonella Enteritis Using a Truncated DNA Aptamer. Microchim. Acta 2018, 185, 61. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| SELEX Rounds | Aptamer [pmol] | Counter SELEX with HDF | Target SELEX with Candida | Wash Times | BSA/tRNA [pmol] |
|---|---|---|---|---|---|
| 9 | 0.1 | 20,000 cells | 250 μL OD600 = 1 | 8 | 3000 |
| 10 | 0.1 | 100,000 cells | 250 μL OD600 = 1 | 8 | 3000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Xing, H.; Bolotnikov, G.; Krämer, M.; Bozdogan, A.; Kissmann, A.-K.; Weil, T.; Spellerberg, B.; Stenger, S.; Rosenau, F. Robust Fluorometric Aptamer Assay for Direct and Rapid Detection of Clinical Isolates of Candida spec. Int. J. Mol. Sci. 2024, 25, 3444. https://doi.org/10.3390/ijms25063444
Zhang Y, Xing H, Bolotnikov G, Krämer M, Bozdogan A, Kissmann A-K, Weil T, Spellerberg B, Stenger S, Rosenau F. Robust Fluorometric Aptamer Assay for Direct and Rapid Detection of Clinical Isolates of Candida spec. International Journal of Molecular Sciences. 2024; 25(6):3444. https://doi.org/10.3390/ijms25063444
Chicago/Turabian StyleZhang, Yiting, Hu Xing, Grigory Bolotnikov, Markus Krämer, Anil Bozdogan, Ann-Kathrin Kissmann, Tanja Weil, Barbara Spellerberg, Steffen Stenger, and Frank Rosenau. 2024. "Robust Fluorometric Aptamer Assay for Direct and Rapid Detection of Clinical Isolates of Candida spec." International Journal of Molecular Sciences 25, no. 6: 3444. https://doi.org/10.3390/ijms25063444
APA StyleZhang, Y., Xing, H., Bolotnikov, G., Krämer, M., Bozdogan, A., Kissmann, A.-K., Weil, T., Spellerberg, B., Stenger, S., & Rosenau, F. (2024). Robust Fluorometric Aptamer Assay for Direct and Rapid Detection of Clinical Isolates of Candida spec. International Journal of Molecular Sciences, 25(6), 3444. https://doi.org/10.3390/ijms25063444