
Cycloolivil Isolated from Nardostachys jatamansi Inhibits TNF-α/IFN-γ-Induced Chemokine Production by Blocking NF-κB and JAK/STAT Activation in HaCaT Keratinocytes

 ,
,
Abstract
1. Introduction
2. Results
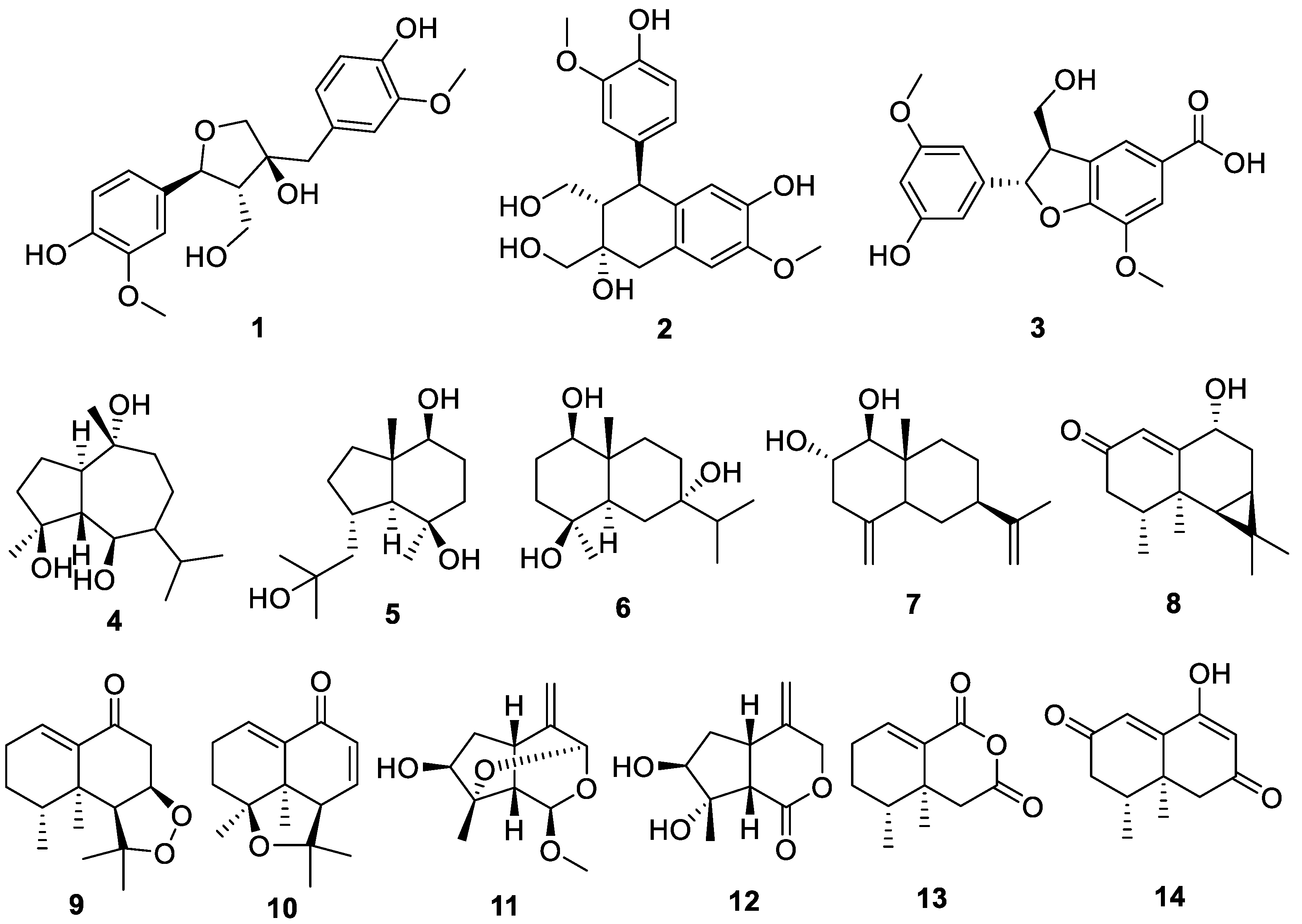
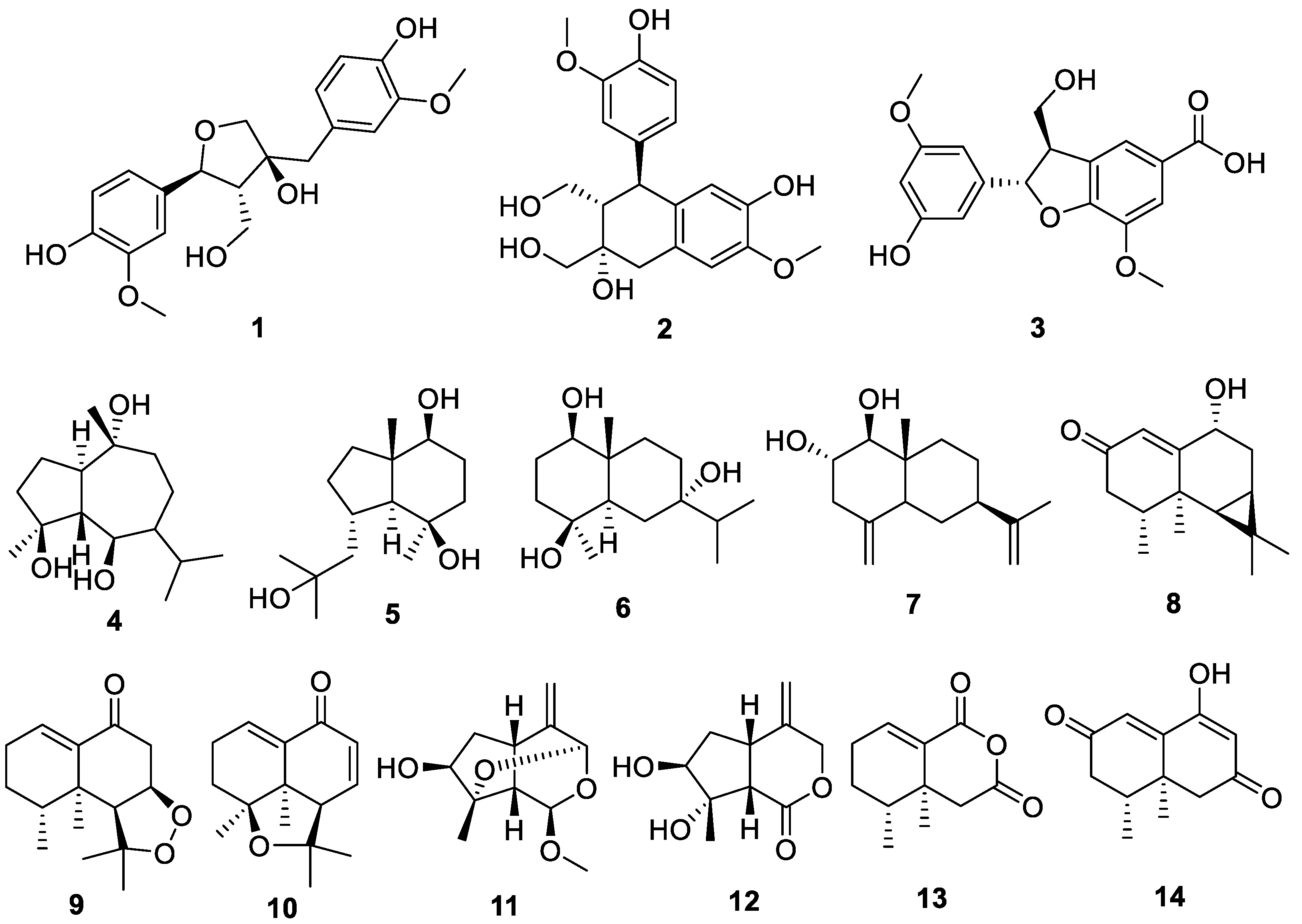
2.1. Chemical Structures of Compounds Isolated from N. jatamansi
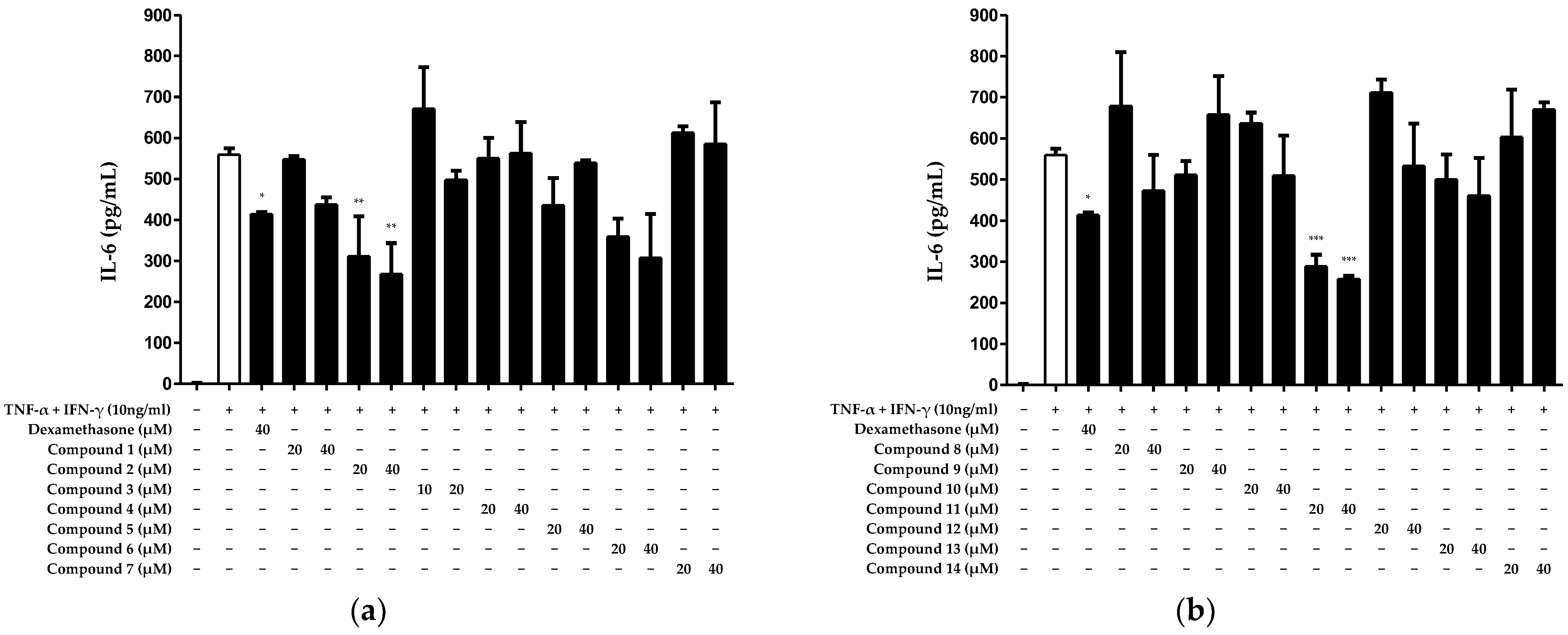
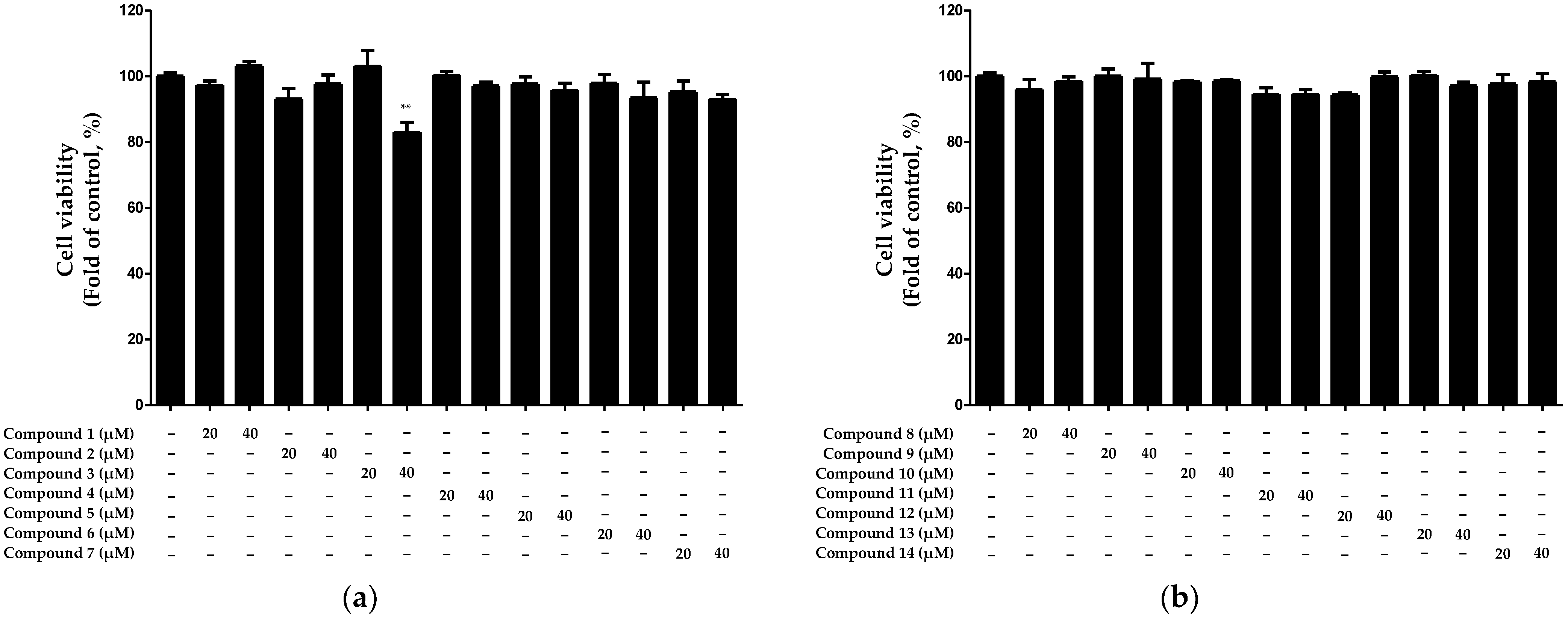
2.2. Inhibitory Effect of Compounds Isolated from N. jatamansi on Interleukin (IL)-6 Production in HaCaT Keratinocytes Induced by TNF-α/IFN-γ
2.3. Inhibitory Effect of Compounds Isolated from N. jatamansi on TNF-α/IFN-γ-Induced IL-8 and RANTES Production in HaCaT Keratinocytes
2.4. Inhibitory Effect of Cycloolivil on TNF-α/IFN-γ-Induced Cyclooxygenase (COX)-2 Expression in HaCaT Keratinocytes
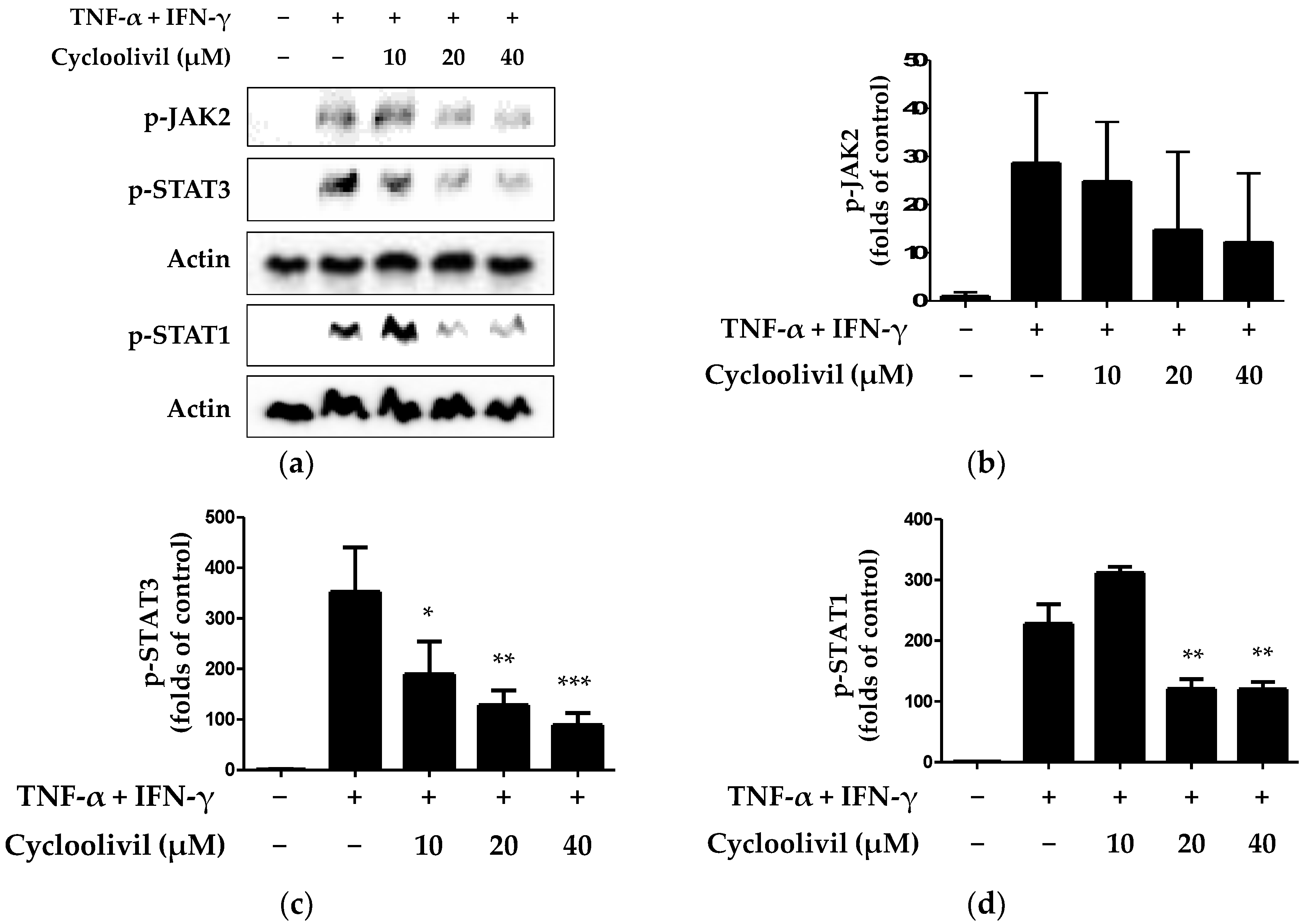
2.5. Effect of Cycloolivil on Regulation of the JAK/STAT Signaling Pathway in HaCaT Keratinocytes
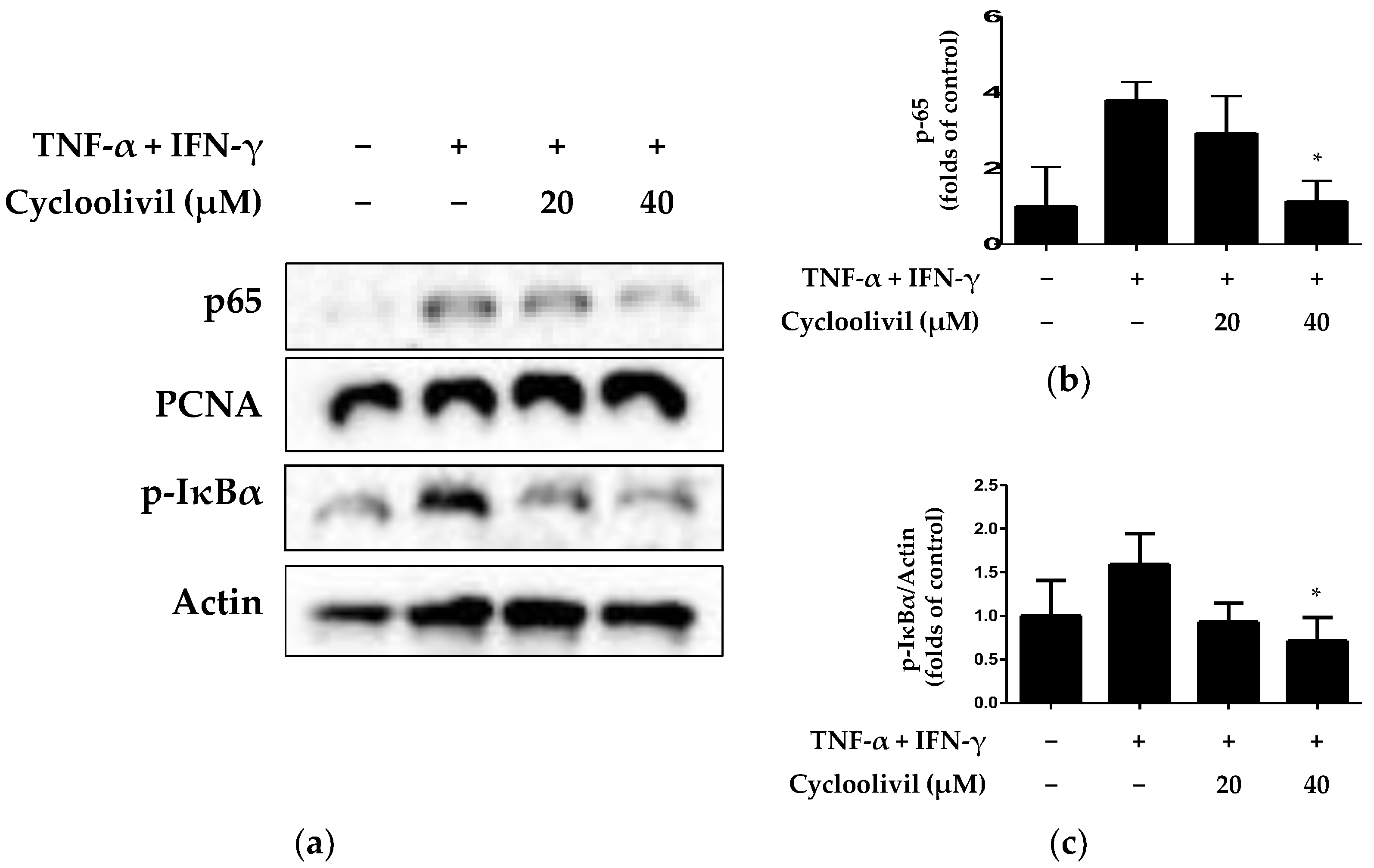
2.6. Effect of Cycloolivil on Regulation of the NF-κB Signaling Pathway in HaCaT Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Materials, Reagents, and Instruments
4.2. Extraction, Isolation, and Structural Identification
4.3. Cell Culture
4.4. MTT Assay
4.5. IL-6, IL-8, and RANTES Detection in the Cell Supernatant
4.6. Western Blot Analysis
4.7. Extraction of Total, Nuclear, and Cytosolic Proteins
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avena-Woods, C. Overview of atopic dermatitis. Am. J. Manag. Care 2017, 23, S115–S123. [Google Scholar]
- Kim, J.Y.; Yang, Y. Effects of Atopic Dermatitis on Suicidal Ideation, Plans and Attempts in Young Adult. J. Korean Acad. Community Health Nurs. 2017, 28, 504–512. [Google Scholar] [CrossRef]
- Arkwright, P.D.; Koplin, J.J. Impact of a Decade of Research into Atopic Dermatitis. J. Allergy Clin. Immunol. Pract. 2023, 11, 63–71. [Google Scholar] [CrossRef]
- Kim, M.; Kim, J. Phenotypes and endotypes of atopic dermatitis: Clinical implications. Allergy Asthma Respir. Dis. 2020, 8, 9–14. [Google Scholar] [CrossRef]
- Chang, Y.S.; Chiang, B.L. Mechanism of Sleep Disturbance in Children with Atopic Dermatitis and the Role of the Circadian Rhythm and Melatonin. Int. J. Mol. Sci. 2016, 17, 462. [Google Scholar] [CrossRef] [PubMed]
- Chieosilapatham, P.; Kiatsurayanon, C.; Umehara, Y.; Trujillo-Paez, J.V.; Peng, G.; Yue, H.; Nguyen, L.T.H.; Niyonsaba, F. Keratinocytes: Innate immune cells in atopic dermatitis. Clin. Exp. Immunol. 2021, 204, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Kim, E.Y.; Lim, S.E.; Kim, J.H.; Sohn, Y.; Jung, H.S. Dendrobium nobile Lindley Administration Attenuates Atopic Dermatitis-like Lesions by Modulating Immune Cells. Int. J. Mol. Sci. 2022, 23, 4470. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.Y.; Yu, H.S.; Ra, M.J.; Jung, S.M.; Yu, J.N.; Kim, J.C.; Kim, K.H. Phytochemical Investigation of Equisetum arvense and Evaluation of Their Anti-Inflammatory Potential in TNFα/INFγ-Stimulated Keratinocytes. Pharmaceuticals 2023, 16, 1478. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Liou, C.J.; Shen, S.C.; Hu, S.; Chao, J.C.; Huang, C.H.; Wu, S.J. Punicalagin from pomegranate ameliorates TNF-alpha/IFN-gamma-induced inflammatory responses in HaCaT cells via regulation of SIRT1/STAT3 axis and Nrf2/HO-1 signaling pathway. Int. Immunopharmacol. 2024, 130, 111665. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, P.; Zhang, J.; Hong, T. Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice. Molecules 2022, 28, 293. [Google Scholar] [CrossRef]
- Kundu, J.; Kim, D.H.; Chae, I.G.; Lee, J.K.; Lee, S.; Jeong, C.H.; Chun, K.S. Silicon dioxide nanoparticles induce COX-2 expression through activation of STAT3 signaling pathway in HaCaT cells. Toxicol. Vitr. 2018, 52, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.B.; Park, S.J. Anti-inflammatory Effects of 8α-hydroxy pinoresinol isolated from Nardostachys jatamansi on Lipopolysaccharide-induced Inflammatory Response in RAW 264.7 Cells. Korea J. Herbol. 2016, 31, 1–6. [Google Scholar]
- Choi, S.S.; Noh, H.S.; Cho, S.H.; Kong, K. Screening of inhibitors against tyrosinase activity from natural products. Yakhak Hoeji 2001, 45, 522–528. [Google Scholar]
- Pathak, S.; Godela, R. Nardostachys jatamansi: Phytochemistry, ethnomedicinal uses, and pharmacological activities: A comprehensive review. Fitoterapia 2023, 172, 105764. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, A.; Oshima, Y.; Hikino, H. Nardostachin, an iridoid of Nardostachys chinensis. Planta Med. 1988, 54, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Schulte, K.E.; Glauch, G.; Rücker, G. Nardosinone, a new constituent of Nardostachys chinensis Batalin. Tetrahedron. Lett. 1965, 35, 3083–3084. [Google Scholar] [CrossRef] [PubMed]
- Shide, L.; Mayer, R.; Rücker, G. Nardonoxide, a New Nardosinane-Type Sesquiterpene Ether from Nardostachys chinensis. Planta Med. 1987, 53, 332–334. [Google Scholar] [CrossRef] [PubMed]
- Itokawa, H.; Masuyama, K.; Morita, H.; Takeya, K. Cytotoxic sesquiterpenes from Nardostachys chinensis. Chem. Pharm. Bull. 1993, 41, 1183–1184. [Google Scholar] [CrossRef]
- Sharma, S.; Rana, A.K.; Rahmatkar, S.N.; Patial, V.; Singh, D. Protective effect of Nardostachys jatamansi extract against lithium-pilocarpine-induced spontaneous recurrent seizures and associated cardiac irregularities in a rat model. J. Ethnopharmacol. 2023, 308, 116280. [Google Scholar] [CrossRef]
- Rehman, T.; Ahmad, S. Nardostachys chinensis Batalin: A review of traditional uses, phytochemistry, and pharmacology. Phytother. Res. 2019, 33, 2622–2648. [Google Scholar] [CrossRef]
- Dong, L.; Kim, H.J.; Cao, T.Q.; Liu, Z.; Lee, H.; Ko, W.; Kim, Y.C.; Sohn, J.H.; Kim, T.K.; Yim, J.H.; et al. Anti-Inflammatory Effects of Metabolites from Antarctic Fungal Strain Pleosporales sp. SF-7343 in HaCaT Human Keratinocytes. Int. J. Mol. Sci. 2021, 22, 9674. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, H.K.; N’deh, K.P.U.; Choi, Y.J.; Fan, M.; Kim, E.K.; Chung, K.H.; An, A.J.H. Inhibitory Effect of Centella asiatica Extract on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Nutrients 2020, 12, 411. [Google Scholar] [CrossRef]
- Walter, M.J.; Look, D.C.; Tidwell, R.M.; Roswit, W.T.; Holtzman, M.J. Targeted inhibition of interferon-γ-dependent intercellular adhesion molecule-1 (ICAM-1) expression using dominant-negative Stat1. J. Biol. Chem. 1997, 272, 28582–28589. [Google Scholar] [CrossRef]
- James, E.; Darnell, J. STATs and gene regulation. Science 1997, 277, 1630–1635. [Google Scholar]
- Oh, J.E.; Kim, M.S.; Jeon, W.K. A nuclear factor kappa B-derived inhibitor tripeptide inhibits UVB-induced photoaging process. J. Dermatol. Sci. 2014, 76, 196–205. [Google Scholar] [CrossRef]
- Kim, K.W.; Yoon, C.S.; Kim, Y.C.; Oh, H. Desoxo-narchinol A and Narchinol B Isolated from Nardostachys jatamansi Exert Anti-neuroinflammatory Effects by Up-regulating of Nuclear Transcription Factor Erythroid-2-Related Factor 2/Heme Oxygenase-1 Signaling. Neurotox. Res. 2019, 35, 230–243. [Google Scholar] [CrossRef]
- Kim, K.W.; Yoon, C.S.; Park, S.J.; Bae, K.S.; Kim, D.G.; Kim, Y.C.; Oh, H. Chemical Analysis of the Ingredients of 20% Aqueous Ethanol Extract of Nardostachys jatamansi through Phytochemical Study and Evaluation of Anti-Neuroinflammatory Component. Evid. Based Complement. Alternat. Med. 2021, 2021, 5901653. [Google Scholar] [CrossRef] [PubMed]
- Juráňová, J.; Franková, J.; Ulrichová, J. The role of keratinocytes in inflammation. J. Appl. Biomed. 2017, 15, 169–179. [Google Scholar] [CrossRef]
- Zbidi, H.; Salido, S.; Altarejos, J.; Perez-Bonilla, M.; Bartegi, A.; Rosado, J.A.; Salido, G.M. Olive tree wood phenolic compounds with human platelet antiaggregant properties. Blood Cells Mol. Dis. 2009, 42, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Rashid, U.; Atta, A.; Khan, I.; Shah, Z.A.; Mobeen, B.; Javed, A.; Alomar, T.S.; Almasoud, N.; Naz, S.; et al. Antiproliferative Activity of Lignans from Olea ferruginea: In Vitro Evidence Supported by Docking Studies. Front. Biosci. Landmark Ed. 2023, 28, 216. [Google Scholar] [CrossRef]
- Yi, M.; Meng, F.C.; Qu, S.Y.; Mao, J.X.; Wang, G.; Liu, M.; Liao, Z.H.; Chen, M. Dolominol a and B, two new neolignans from Dolomiaea souliei (Franch.) C.Shih. Nat. Prod. Res. 2022, 36, 3909–3916. [Google Scholar] [CrossRef]
- Zafar, S.; Ur-Rehman, F.; Shah, Z.A.; Rauf, A.; Khan, A.; Humayun Khan, M.; Ur Rahman, K.; Khan, S.; Ullah, A.; Shaheen, F. Potent leishmanicidal and antibacterial metabolites from Olea ferruginea. J. Asian Nat. Prod. Res. 2019, 21, 679–687. [Google Scholar] [CrossRef]
- Pérez-Bonilla, M.; Salido, S.; van Beek, T.A.; Linares-Palomino, P.J.; Altarejos, J.; Nogueras, M.; Sánchez, A. Isolation and identification of radical scavengers in olive tree (Olea europaea) wood. J. Chromatogr. A 2006, 1112, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.S.; Kim, K.W.; Lee, S.C.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory effects of sesquiterpenoids isolated from Nardostachys jatamansi. Bioorg. Med. Chem. Lett. 2018, 28, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.J.; Chen, H.M.; Yang, F.; Deng, Y.; Ao, H.; Xie, X.F.; Li, H.X.; Zhang, H.; Cao, Z.X.; Zhu, L.X.; et al. Iridoids from the roots of Valeriana jatamansi Jones. Phytochemistry 2017, 141, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.S.; Kim, D.C.; Park, J.S.; Kim, K.W.; Kim, Y.C.; Oh, H. Isolation of Novel Sesquiterpeniods and Anti-neuroinflammatory Metabolites from Nardostachys jatamansi. Molecules 2018, 23, 2367. [Google Scholar] [CrossRef] [PubMed]
- Beken, B.; Serttas, R.; Yazicioglu, M.; Turkekul, K.; Erdogan, S. Quercetin Improves Inflammation, Oxidative Stress, and Impaired Wound Healing in Atopic Dermatitis Model of Human Keratinocytes. Pediatr. Allergy Immunol. Pulmonol. 2020, 33, 69–79. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (IL-6) |
|---|---|
| Cycloolivil (2) | 31.05 ± 0.93 μM |
| 4β-hydroxy-8β-methoxy-10-methylene-2,9-dioxatricyclo[4.3.1.03,7]decane (11) | 28.25 ± 0.21 μM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, C.-S.; Lee, H.; Liu, Z.; Dong, L.; Lee, G.; Kim, N.; Oh, H.; Lee, D.-S. Cycloolivil Isolated from Nardostachys jatamansi Inhibits TNF-α/IFN-γ-Induced Chemokine Production by Blocking NF-κB and JAK/STAT Activation in HaCaT Keratinocytes. Int. J. Mol. Sci. 2024, 25, 3342. https://doi.org/10.3390/ijms25063342
Yoon C-S, Lee H, Liu Z, Dong L, Lee G, Kim N, Oh H, Lee D-S. Cycloolivil Isolated from Nardostachys jatamansi Inhibits TNF-α/IFN-γ-Induced Chemokine Production by Blocking NF-κB and JAK/STAT Activation in HaCaT Keratinocytes. International Journal of Molecular Sciences. 2024; 25(6):3342. https://doi.org/10.3390/ijms25063342
Chicago/Turabian StyleYoon, Chi-Su, Hwan Lee, Zhiming Liu, Linsha Dong, Gyoyoung Lee, Nayeon Kim, Hyuncheol Oh, and Dong-Sung Lee. 2024. "Cycloolivil Isolated from Nardostachys jatamansi Inhibits TNF-α/IFN-γ-Induced Chemokine Production by Blocking NF-κB and JAK/STAT Activation in HaCaT Keratinocytes" International Journal of Molecular Sciences 25, no. 6: 3342. https://doi.org/10.3390/ijms25063342
APA StyleYoon, C.-S., Lee, H., Liu, Z., Dong, L., Lee, G., Kim, N., Oh, H., & Lee, D.-S. (2024). Cycloolivil Isolated from Nardostachys jatamansi Inhibits TNF-α/IFN-γ-Induced Chemokine Production by Blocking NF-κB and JAK/STAT Activation in HaCaT Keratinocytes. International Journal of Molecular Sciences, 25(6), 3342. https://doi.org/10.3390/ijms25063342