
Hypothermia Inhibits Dexmedetomidine-Induced Contractions in Isolated Rat Aortae

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
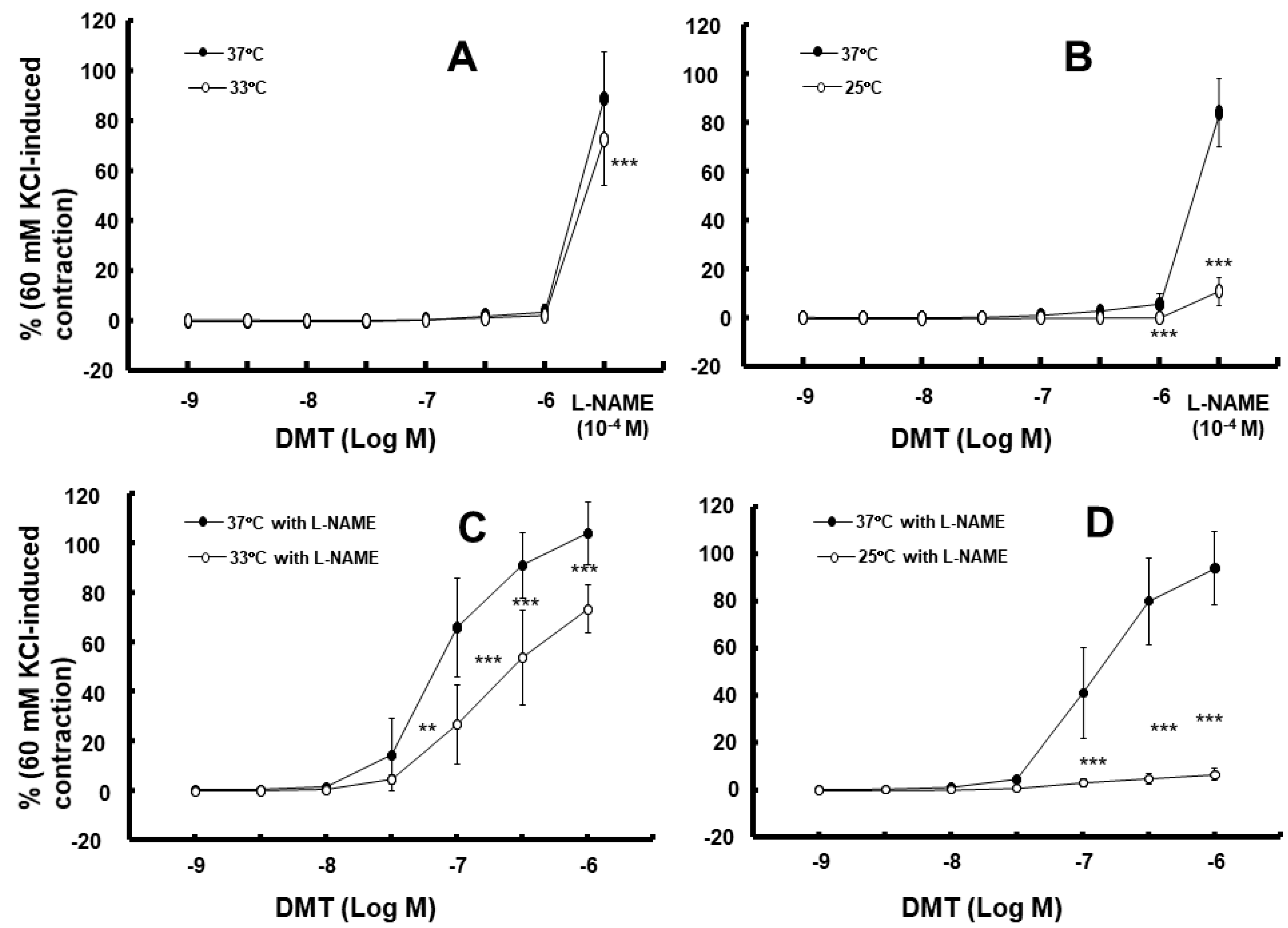
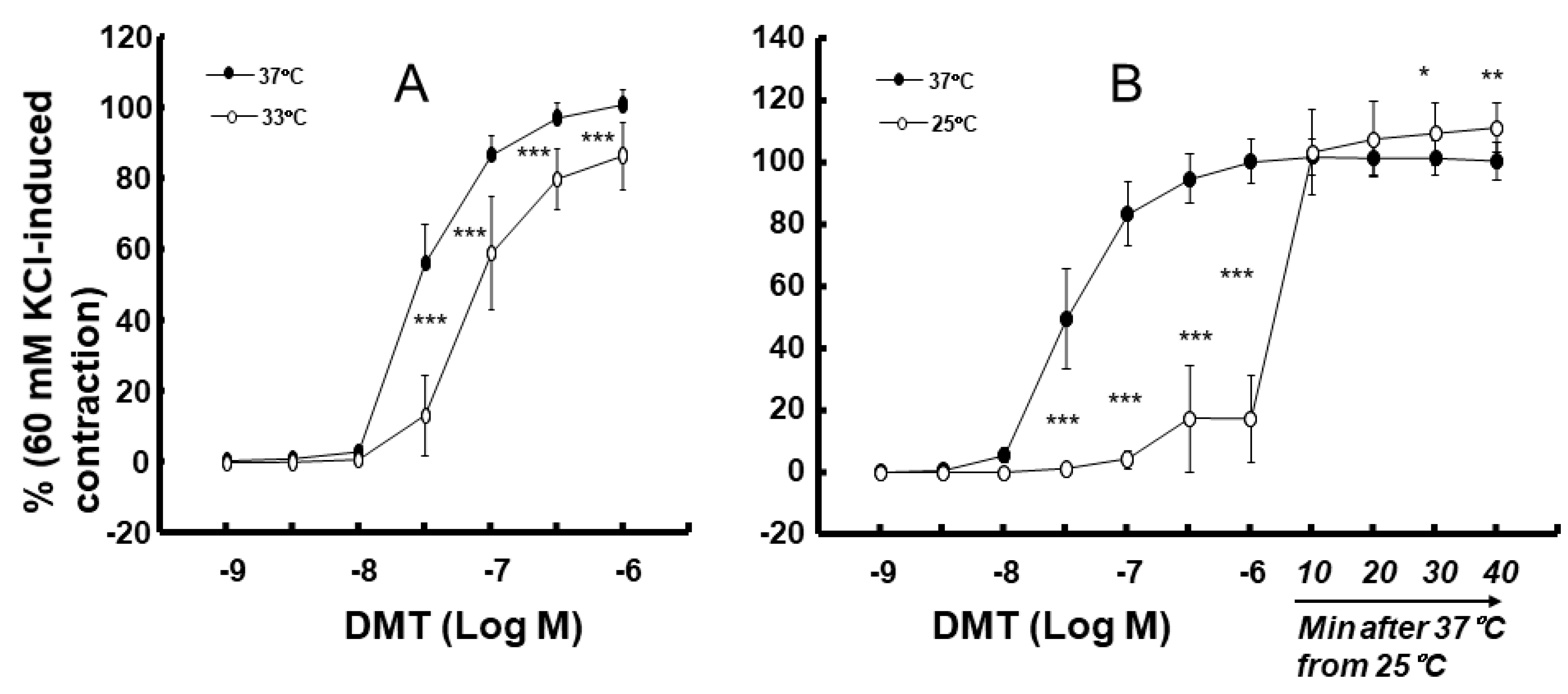
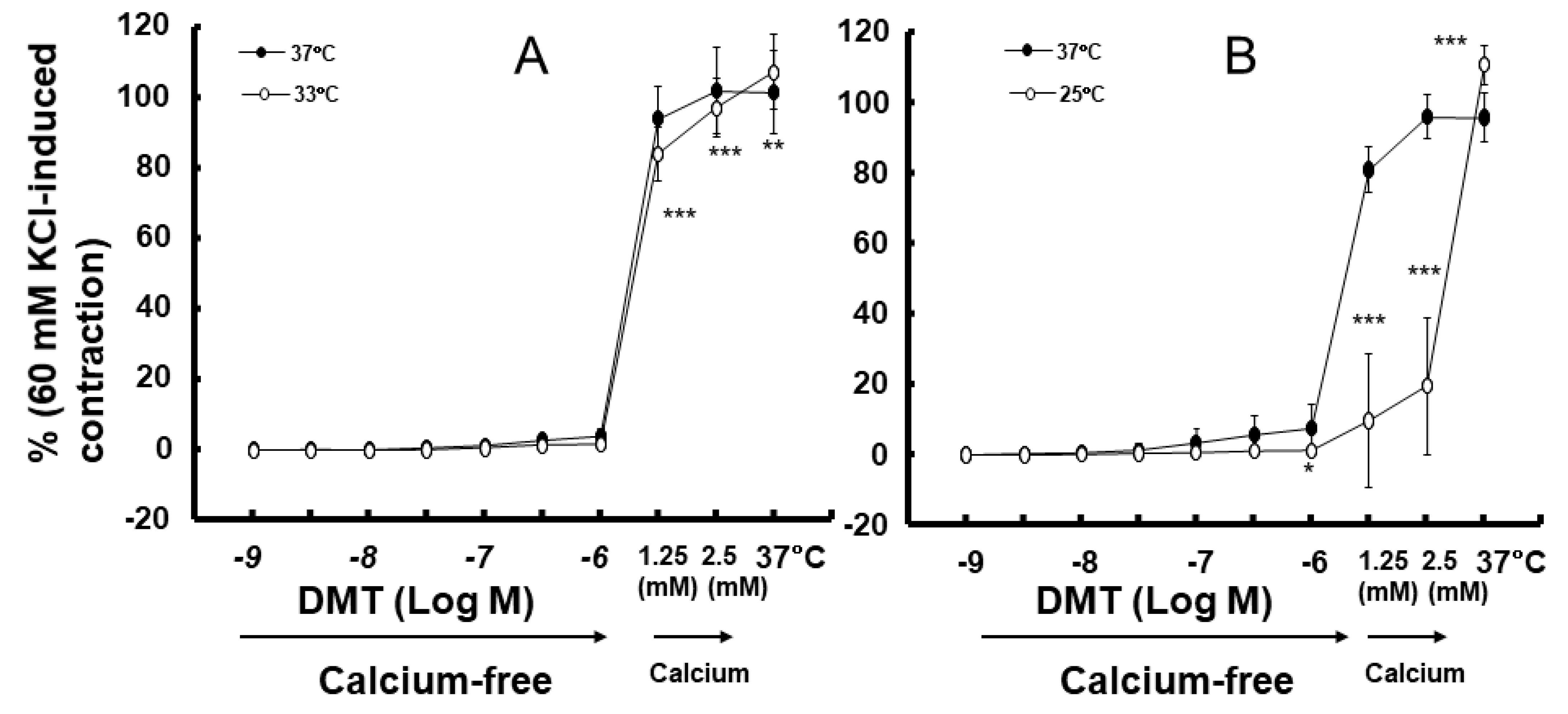
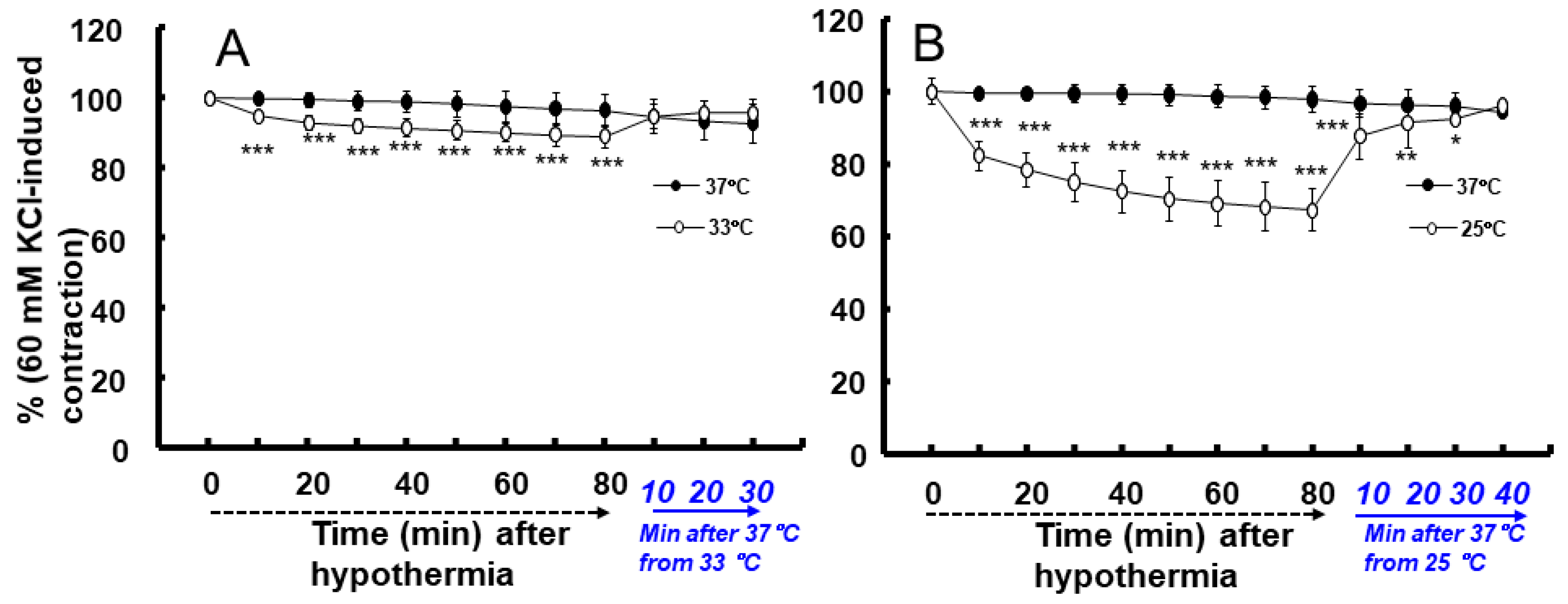
2. Results
3. Discussion
4. Materials and Methods
4.1. Experiment Protocols
4.2. Cell Culture
4.3. Western Blot
4.4. Materials
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Virtanen, R.; Savola, J.M.; Saano, V.; Nyman, L. Characterization of the selectivity, specificity and potency of medetomidine as an alpha 2-adrenoceptor agonist. Eur. J. Pharmacol. 1988, 150, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Gertler, R.; Brown, H.C.; Mitchell, D.H.; Silvius, E.N. Dexmedetomidine: A novel sedative-analgesic agent. Bayl. Univ. Med. Cent. Proc. 2001, 1, 13–21. [Google Scholar] [CrossRef]
- Sandestig, A.; Romner, B.; Grände, P.O. Therapeutic hypothermia in children and adults with severe traumatic brain injury. Ther. Hypothermia Temp. Manag. 2014, 4, 10–20. [Google Scholar] [CrossRef]
- Peberdy, M.A.; Callaway, C.W.; Neumar, R.W.; Geocadin, R.G.; Zimmerman, J.L.; Donnino, M.; Gabrielli, A.; Silvers, S.M.; Zaritsky, A.L.; Merchant, R.; et al. American Heart Association. Part 9: Post-cardiac arrest care: 2010 American Heart Association Guidelines for Cardiopulmonary Resuscitation and Emergency Cardiovascular Care. Circulation 2010, 122, S768–S786. [Google Scholar] [CrossRef] [PubMed]
- Karcioglu, O.; Topacoglu, H.; Dikme, O.; Dikme, O. A systematic review of safety and adverse effects in the practice of therapeutic hypothermia. Am. J. Emerg. Med. 2018, 36, 1886–1894. [Google Scholar] [CrossRef] [PubMed]
- Evora, P.R.; Cable, D.G.; Chua, Y.L.; Rodrigues, A.J.; Pearson, P.J.; Schaff, H.V. Nitric oxide and prostacyclin-dependent pathways involvement on in vitro induced hypothermia. Cryobiology 2007, 54, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Schmeling, W.T.; Kampine, J.P.; Roerig, D.L.; Warltier, D.C. The effects of the stereoisomers of the alpha 2-adrenergic agonist medetomidine on systemic and coronary hemodynamics in conscious dogs. Anesthesiology 1991, 75, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Mason, K.P.; Zurakowski, D.; Zgleszewski, S.; Prescilla, R.; Fontaine, P.J.; Dinardo, J.A. Incidence and predictors of hypertension during high-dose dexmedetomidine sedation for pediatric MRI. Pediatr. Anesth. 2010, 20, 516–523. [Google Scholar] [CrossRef]
- Erkonen, G.; Lamb, F.; Tobias, J.D. High-dose dexmedetomidine-induced hypertension in a child with traumatic brain injury. Neurocrit. Care 2008, 9, 366–369. [Google Scholar] [CrossRef]
- Shah, S.; Sangari, T.; Qasim, M.; Martin, T. Severe hypertension and bradycardia after dexmedetomidine for radiology sedation in a patient with acute transverse myelitis. Pediatr. Anesth. 2008, 18, 681–682. [Google Scholar] [CrossRef]
- Guimarães, S.; Moura, D. Vascular adrenoceptors: An update. Pharmacol. Rev. 2001, 53, 319–356. [Google Scholar] [PubMed]
- Akata, T. General anesthetics and vascular smooth muscle: Direct actions of general anesthetics on cellular mechanisms regulating vascular tone. Anesthesiology 2007, 106, 365–391. [Google Scholar] [CrossRef] [PubMed]
- Ok, S.H.; Bae, S.I.; Shim, H.S.; Sohn, J.T. Dexmedetomidine-induced contraction of isolated rat aorta is dependent on extracellular calcium concentration. Korean J. Anesthesiol. 2012, 63, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Sung, H.J.; Ok, S.H.; Kwon, S.C.; Cheon, K.S.; Kim, H.J.; Chang, K.C.; Shin, I.W.; Lee, H.K.; Chung, Y.K.; et al. Calcium sensitization involved in dexmedetomidine-induced contraction of isolated rat aorta. Can. J. Physiol. Pharmacol. 2011, 89, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Sohn, J.T.; Jeong, Y.S.; Cho, M.S.; Kim, H.J.; Chang, K.C.; Shin, M.K.; Park, C.S.; Chung, Y.K. Direct effect of dexmedetomidine on rat isolated aorta involves endothelial nitric oxide synthesis and activation of the lipoxygenase pathway. Clin. Exp. Pharmacol. Physiol. 2009, 36, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.H.; Oh, K.W.; Kim, S.T.; Park, E.S.; Je, H.D.; Yoon, H.J.; Sohn, U.D.; Jeong, J.H.; La, H.O. Hypothermia inhibits endothelium-independent vascular contractility via Rho-kinase inhibition. Biomol. Ther. 2018, 26, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Thoresen, M.; Whitelaw, A. Cardiovascular changes during mild therapeutic hypothermia and rewarming in infants with hypoxic-ischemic encephalopathy. Pediatrics 2000, 106, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Ok, S.H.; Kwon, S.C.; Baik, J.; Hong, J.M.; Oh, J.; Han, J.Y.; Sohn, J.T. Dexmedetomidine induced contraction involves CPI-17 phosphorylation in isolated rat aortas. Int. J. Mol. Sci. 2016, 17, 1663. [Google Scholar] [CrossRef]
- Jeon, S.B.; Jin, F.; Kim, J.I.; Kim, S.H.; Suk, K.; Chae, S.C.; Jun, J.E.; Park, W.H.; Kim, I.K. A role for Rho kinase in vascular contraction evoked by sodium fluoride. Biochem. Biophys. Res. Commun. 2006, 343, 27–33. [Google Scholar] [CrossRef]
- Ok, S.H.; Byon, H.J.; Kwon, S.C.; Park, J.; Lee, Y.; Hwang, Y.; Baik, J.; Choi, M.J.; Sohn, J.T. Lipid emulsion inhibits vasodilation induced by a toxic dose of bupivacaine via attenuated dephosphorylation of myosin phosphatase target subunit 1 in isolated rat aorta. Int. J. Med. Sci. 2015, 12, 958–967. [Google Scholar] [CrossRef]
- Clifford, P.S. Local control of blood flow. Adv. Physiol. Educ. 2011, 35, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Deussen, A. Hyperthermia and hypothermia. Effects on the cardiovascular system. Anaesthesist 2007, 56, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Park, K.E.; Lee, S.H.; Bae, S.I.; Hwang, Y.; Ok, S.H.; Ahn, S.H.; Sim, G.; Chung, S.; Sohn, J.T. Lipid emulsion inhibits amlodipine-induced nitric oxide-mediated vasodilation in isolated rat aorta. Int. J. Mol. Sci. 2023, 24, 8741. [Google Scholar] [CrossRef] [PubMed]
- Klöss, S.; Bouloumié, A.; Mülsch, A. Aging and chronic hypertension decrease expression of rat aortic soluble guanylyl cyclase. Hypertension 2000, 35, 43–47. [Google Scholar] [CrossRef]
- Lee, S.H.; Kwon, S.C.; Ok, S.H.; Hong, J.M.; Kim, J.Y.; Ahn, S.H.; Il Bae, S.; Shin, Y.; Sohn, J.T. Levobupivacaine-induced vasoconstriction involves caldesmon phosphorylation mediated by tyrosine kinase-induced ERK phosphorylation. Eur. J. Pharmacol. 2019, 842, 167–176. [Google Scholar] [CrossRef]
- Lavergne, C.; Martinez, M.J.; Trottier, C. Empirical model selection in generalized linear mixed effects models. Comput. Stat. 2008, 23, 99–109. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Hwang, Y.; Park, K.-E.; Bae, S.; Ok, S.-H.; Ahn, S.-H.; Sim, G.; Bae, M.; Sohn, J.-T. Hypothermia Inhibits Dexmedetomidine-Induced Contractions in Isolated Rat Aortae. Int. J. Mol. Sci. 2024, 25, 3017. https://doi.org/10.3390/ijms25053017
Lee S, Hwang Y, Park K-E, Bae S, Ok S-H, Ahn S-H, Sim G, Bae M, Sohn J-T. Hypothermia Inhibits Dexmedetomidine-Induced Contractions in Isolated Rat Aortae. International Journal of Molecular Sciences. 2024; 25(5):3017. https://doi.org/10.3390/ijms25053017
Chicago/Turabian StyleLee, Soohee, Yeran Hwang, Kyeong-Eon Park, Sungil Bae, Seong-Ho Ok, Seung-Hyun Ahn, Gyujin Sim, Moonju Bae, and Ju-Tae Sohn. 2024. "Hypothermia Inhibits Dexmedetomidine-Induced Contractions in Isolated Rat Aortae" International Journal of Molecular Sciences 25, no. 5: 3017. https://doi.org/10.3390/ijms25053017
APA StyleLee, S., Hwang, Y., Park, K.-E., Bae, S., Ok, S.-H., Ahn, S.-H., Sim, G., Bae, M., & Sohn, J.-T. (2024). Hypothermia Inhibits Dexmedetomidine-Induced Contractions in Isolated Rat Aortae. International Journal of Molecular Sciences, 25(5), 3017. https://doi.org/10.3390/ijms25053017