
Effects of Nutraceutical Compositions Containing Rhizoma Gastrodiae or Lipoic Acid in an In Vitro Induced Neuropathic Pain Model



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
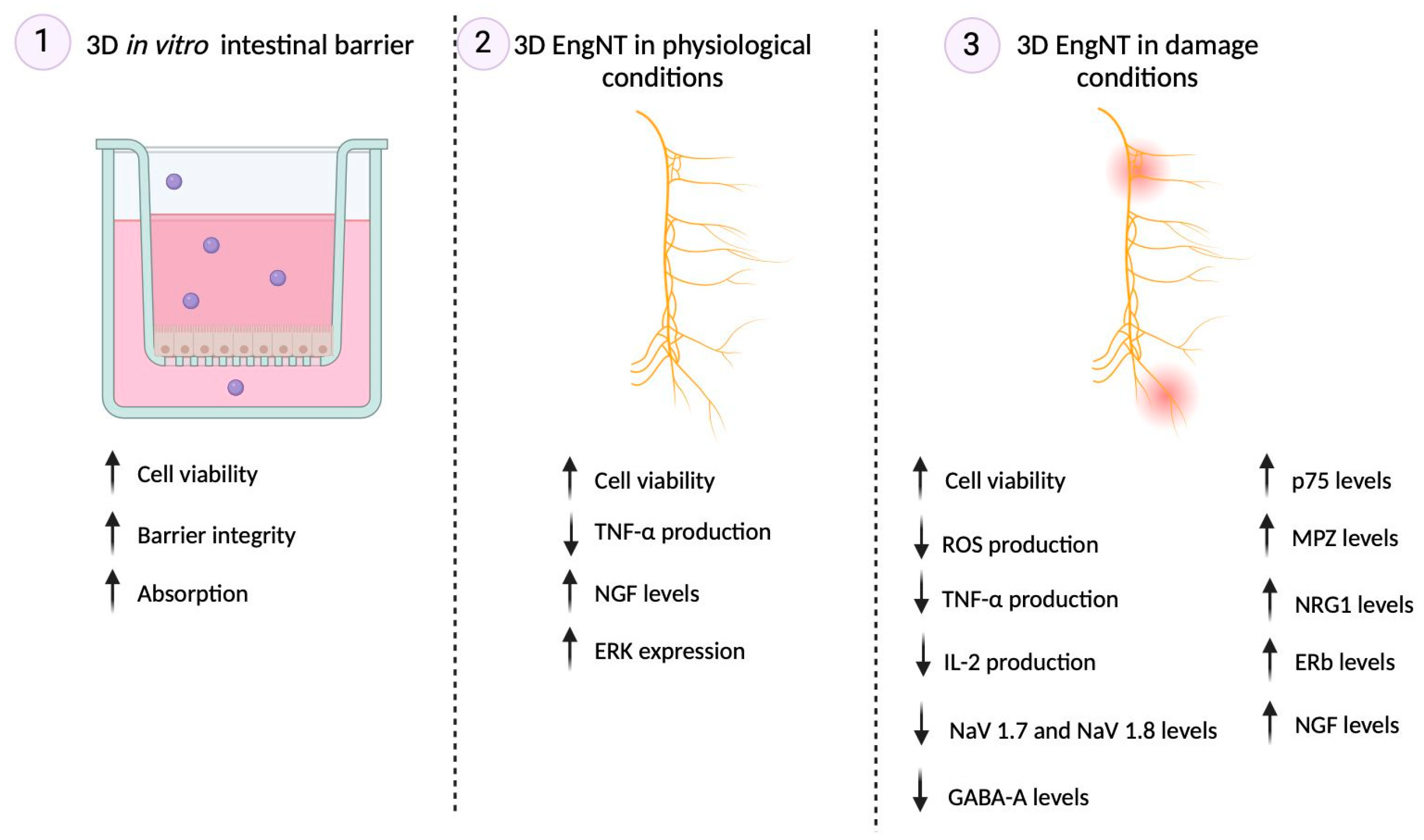
2. Results
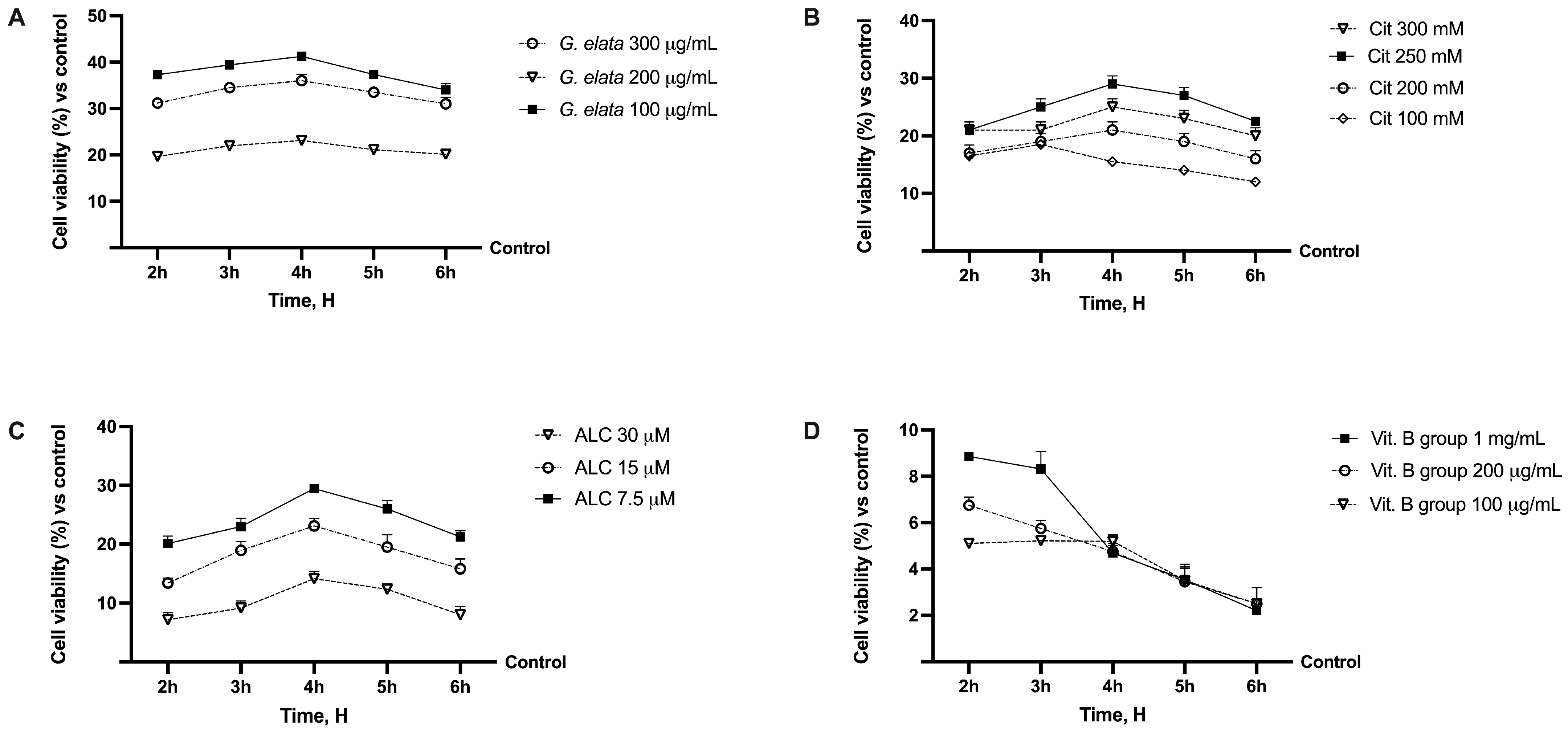
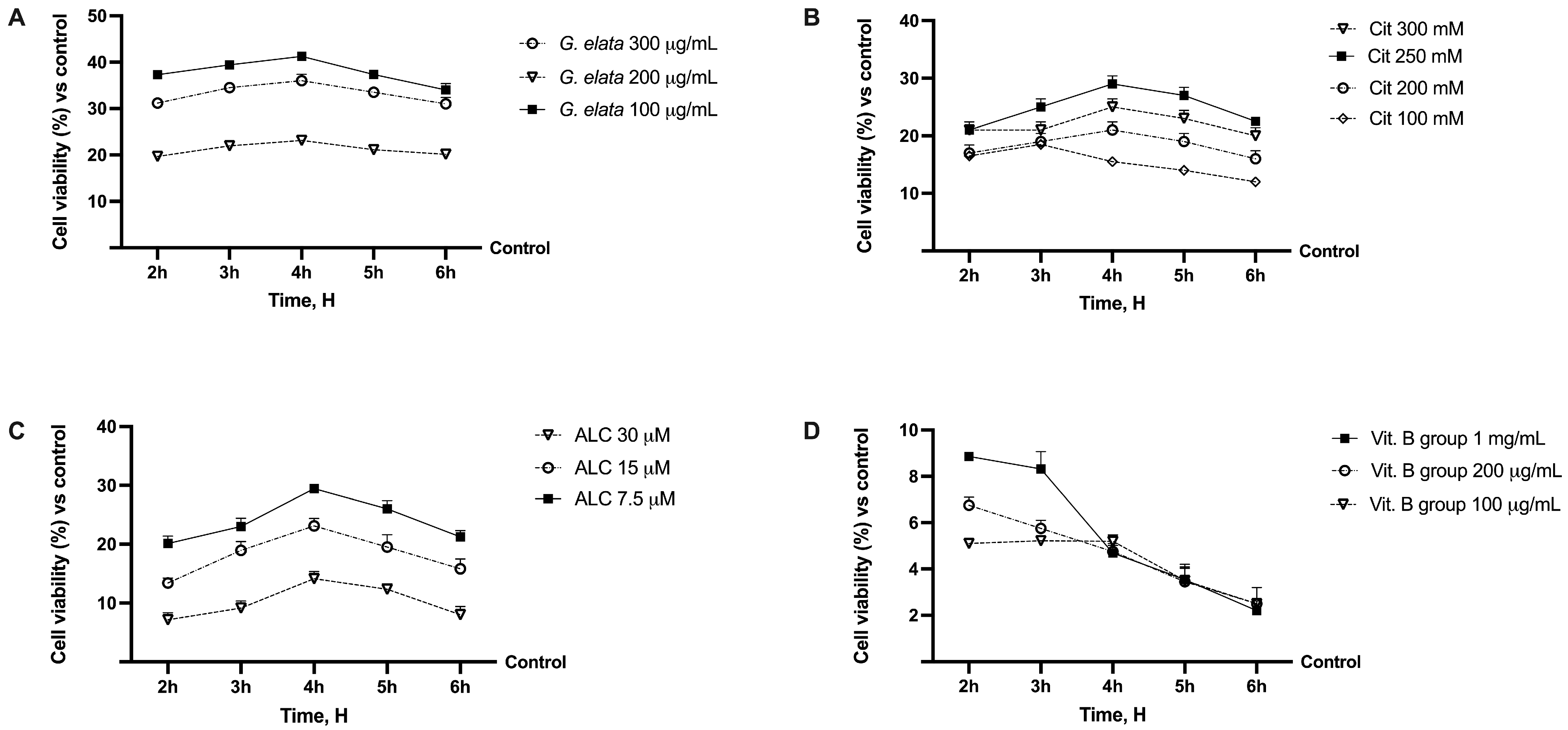
2.1. Dose–Response Study of the Single Components on Caco-2
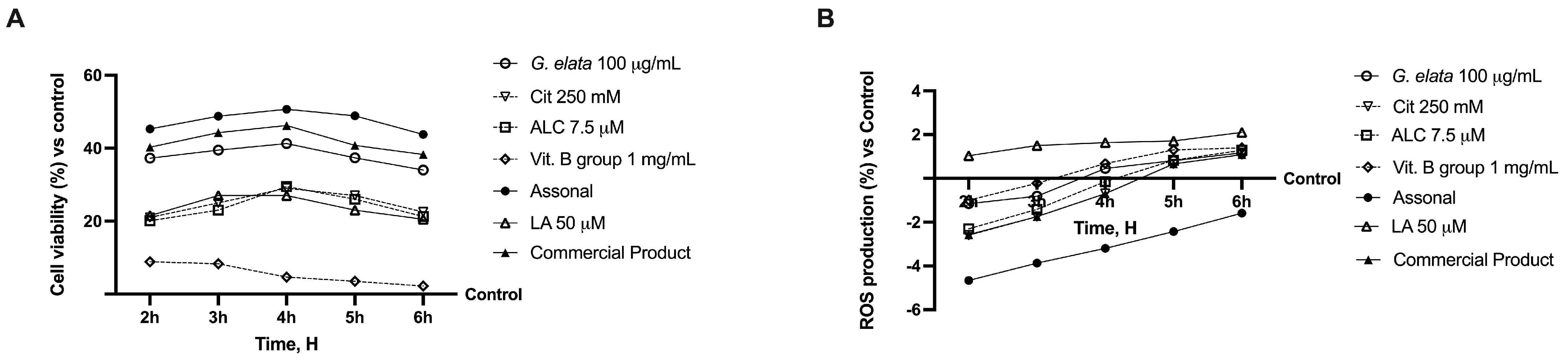
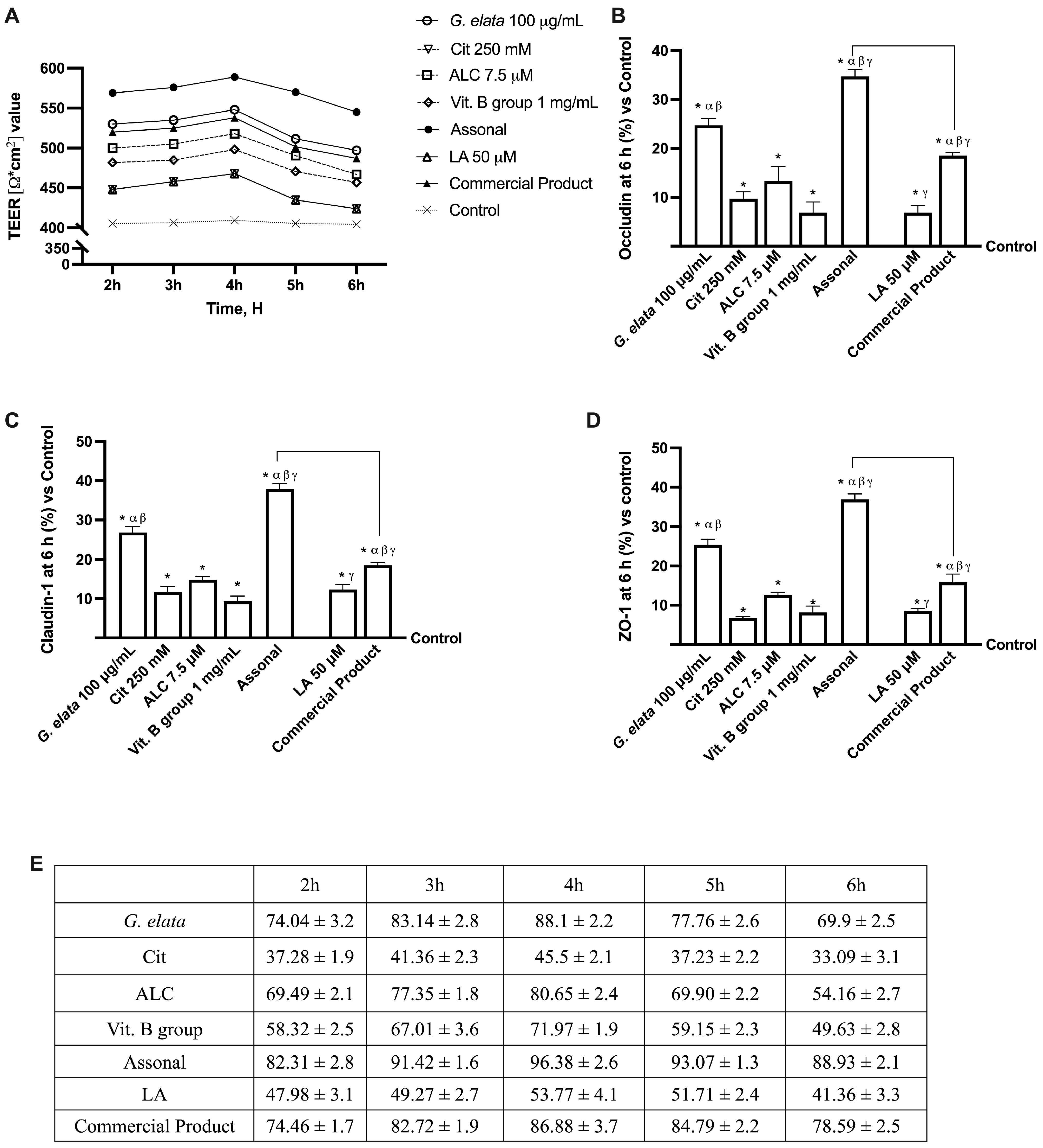
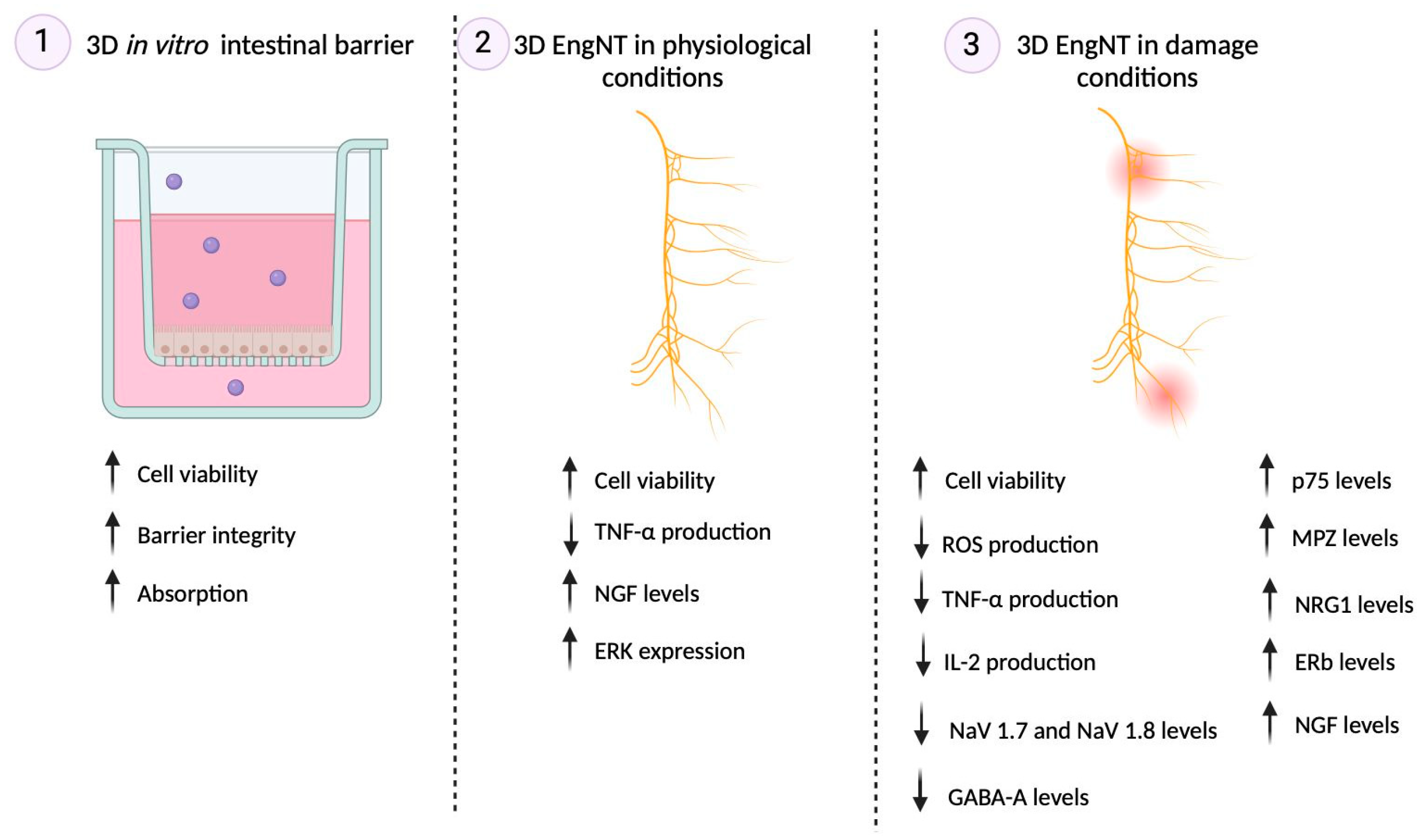
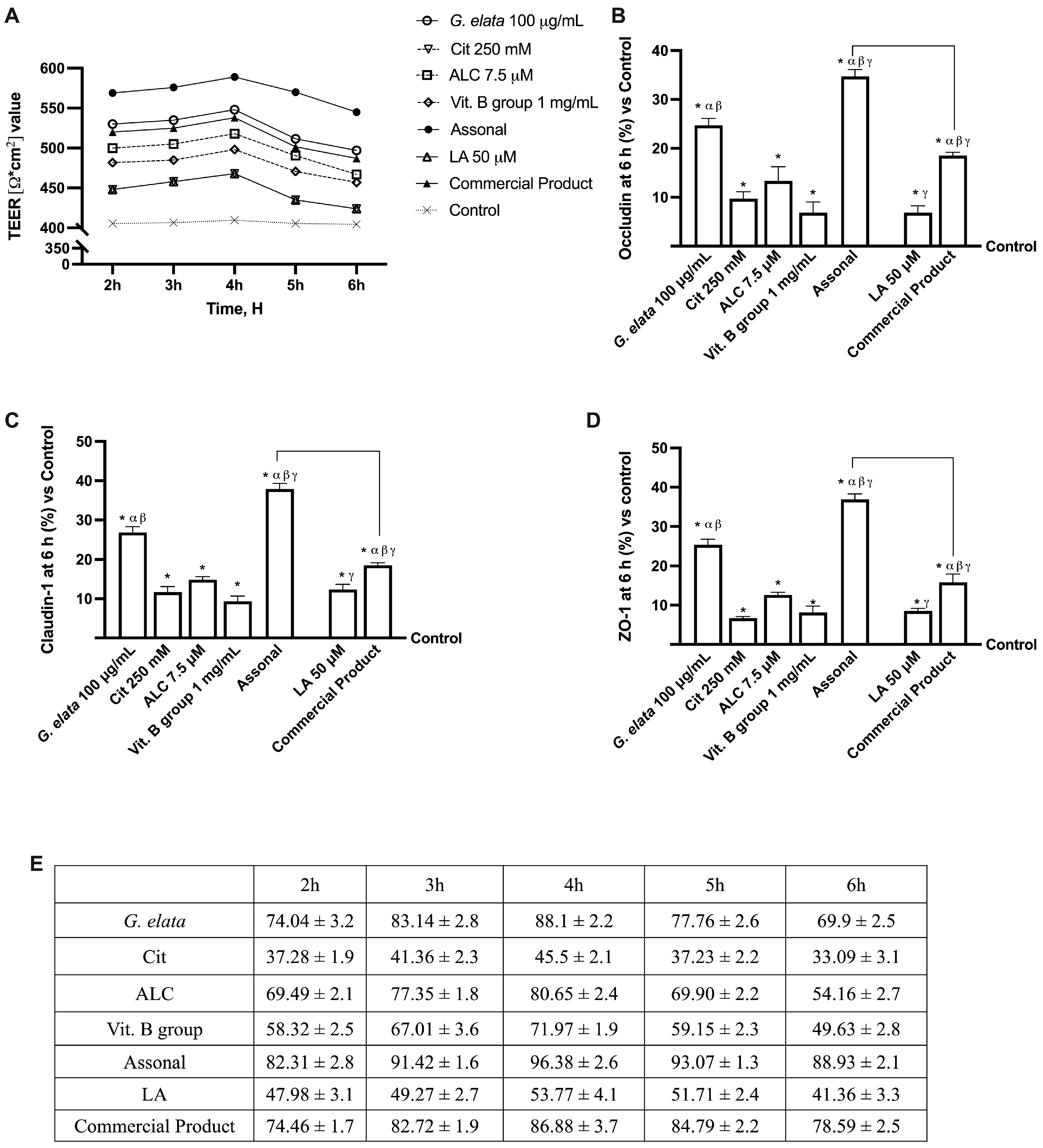
2.2. Permeability and Absorption Mechanisms Analyzed in an In Vitro Intestinal Barrier Model
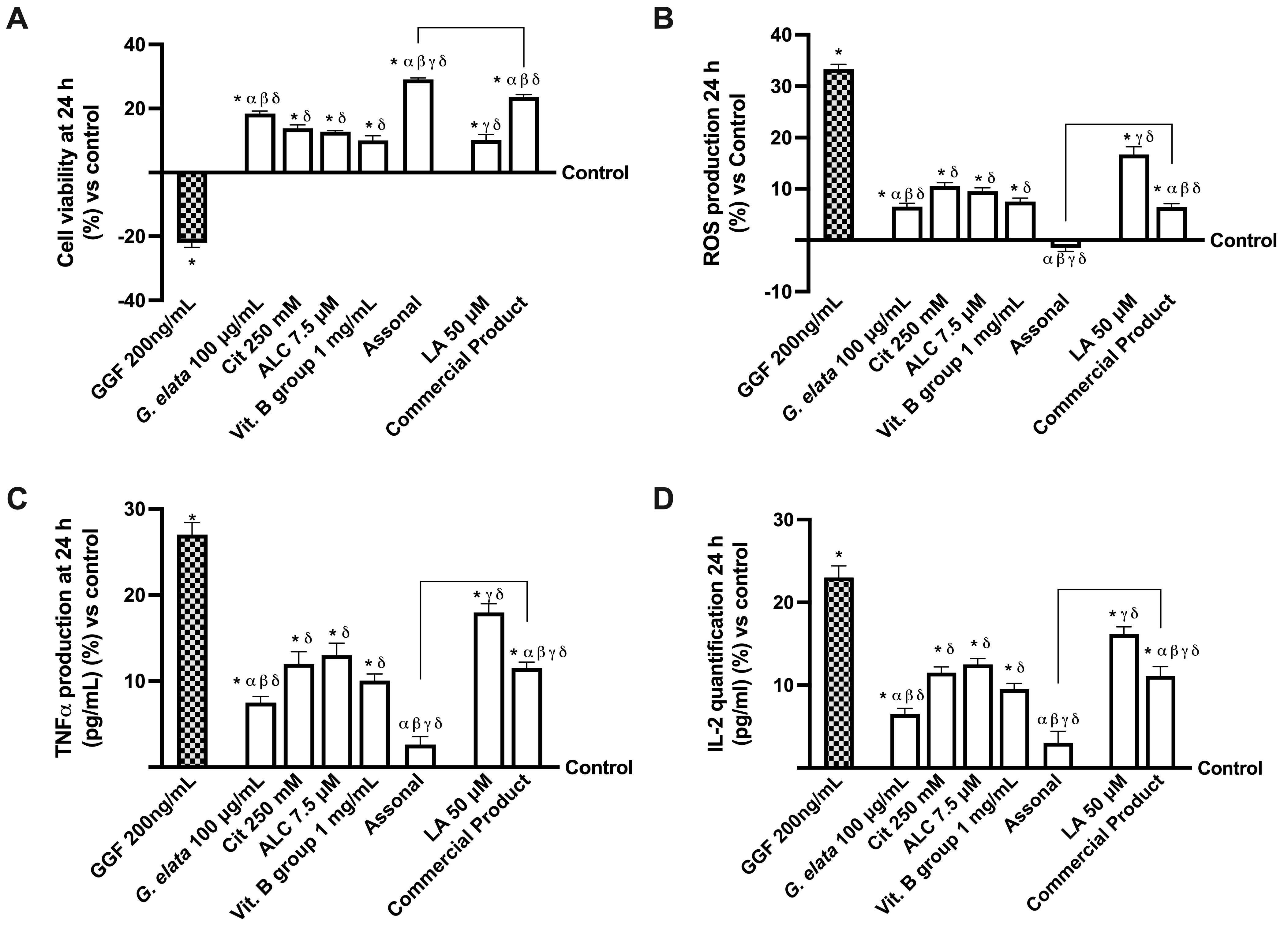
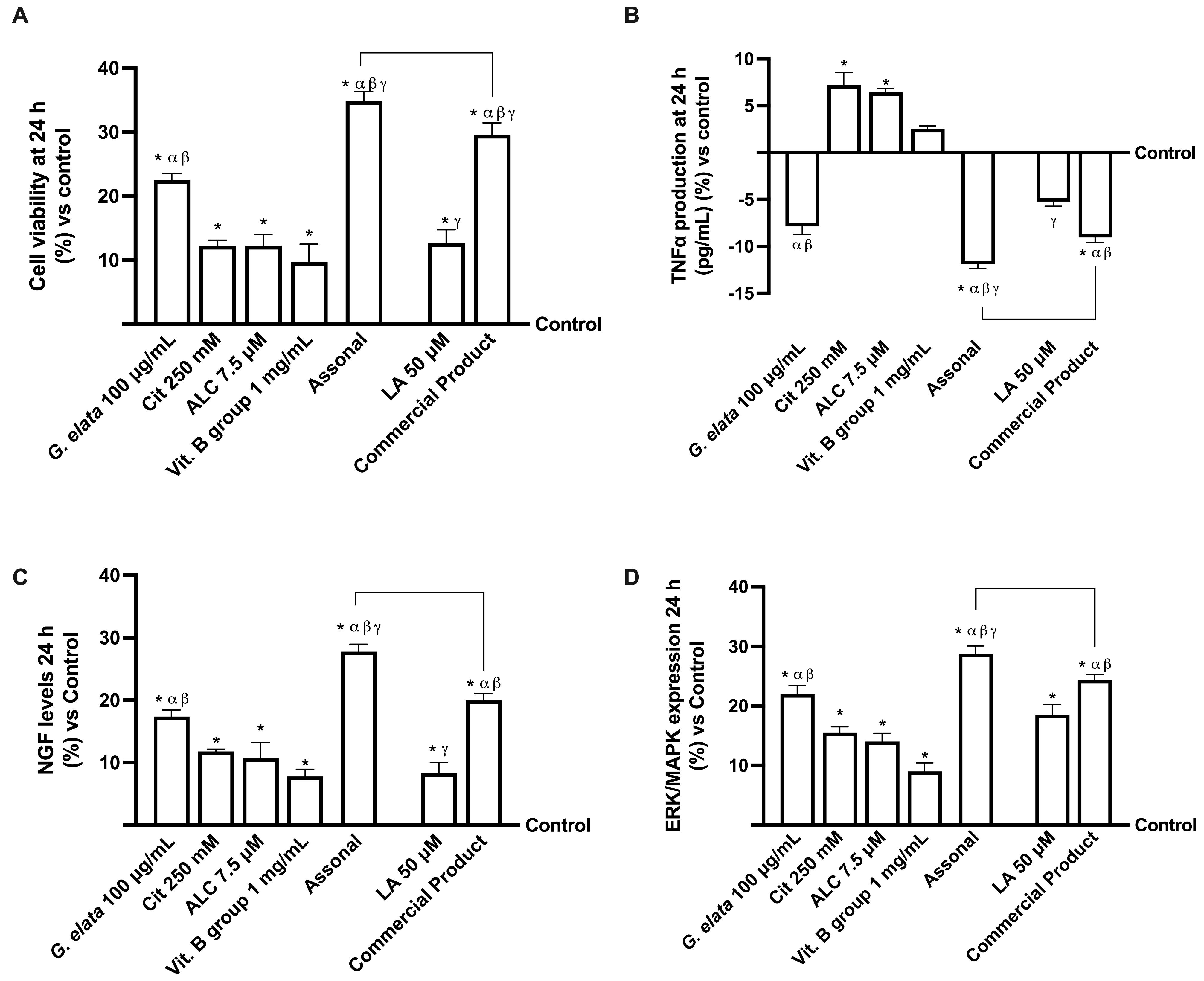
2.3. Effects of Single Components and Combination on 3D EngNT Co-Cultures
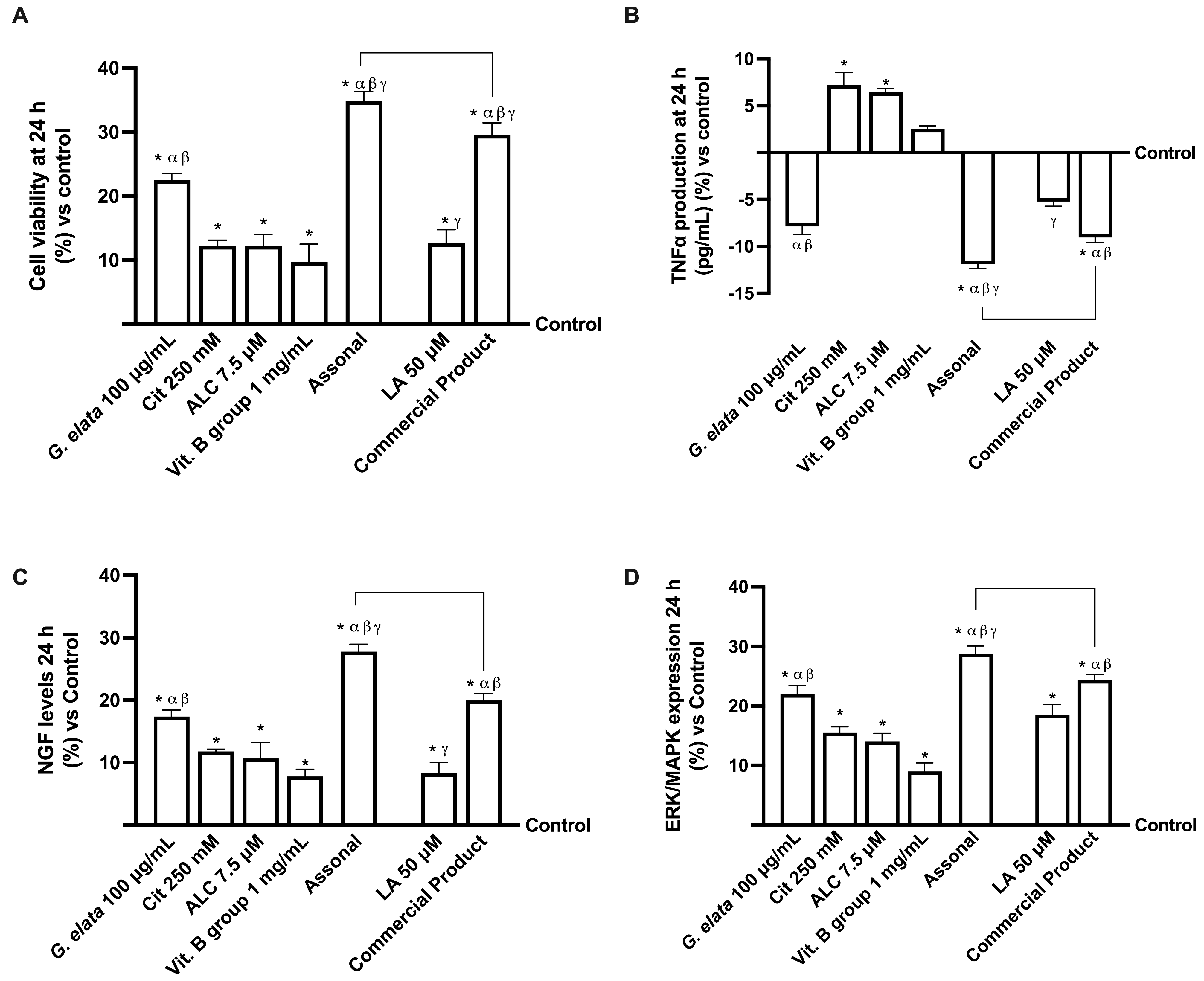
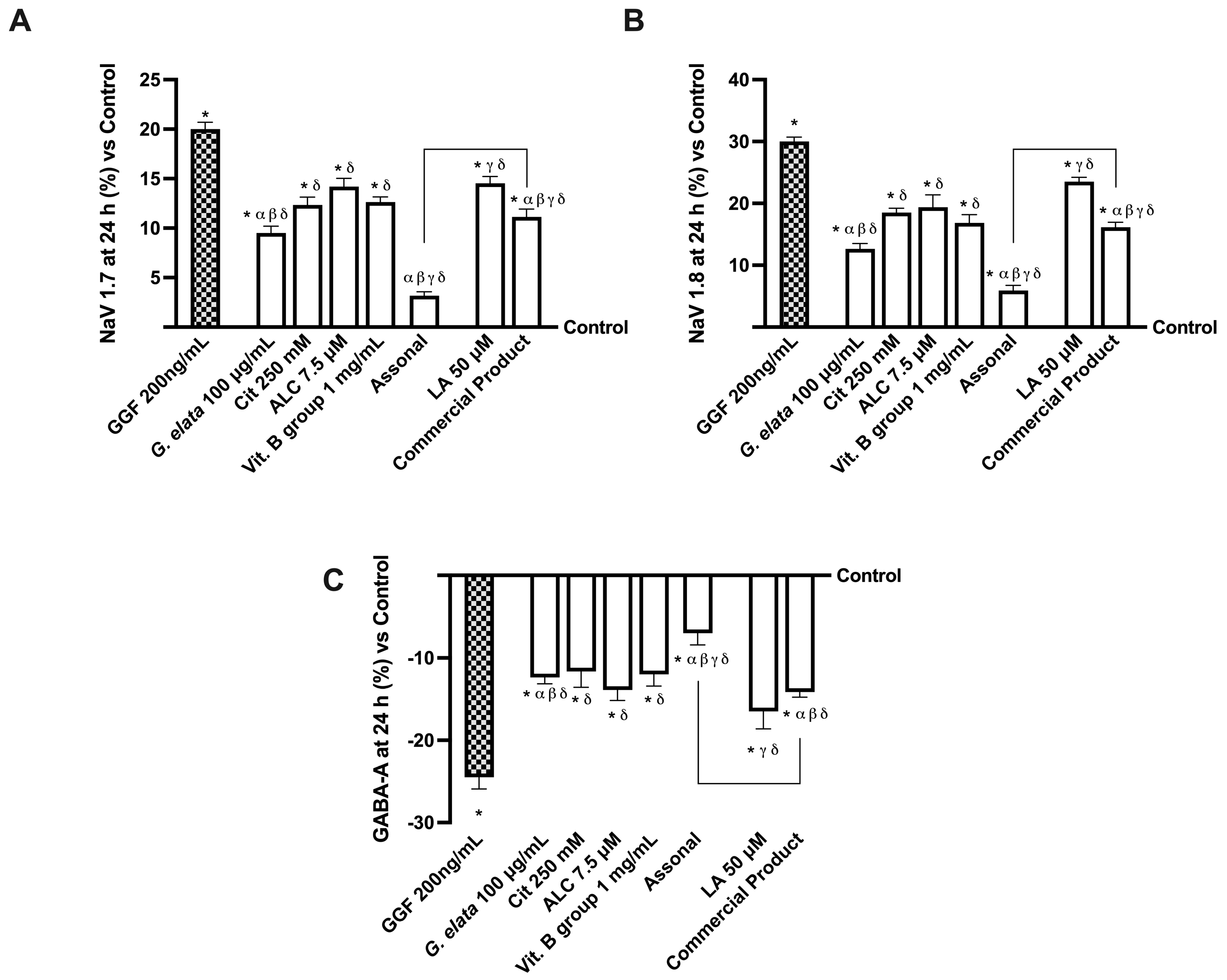
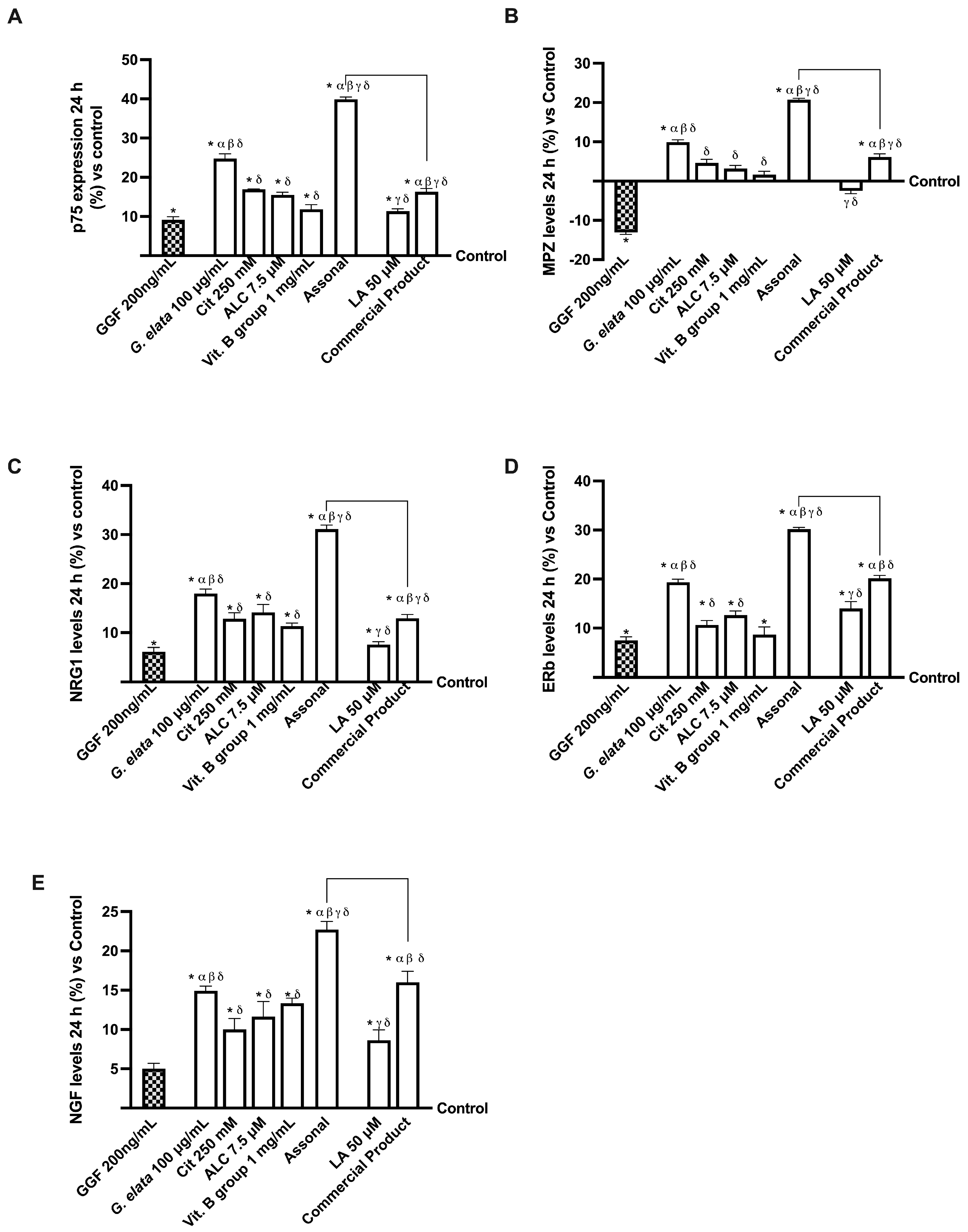
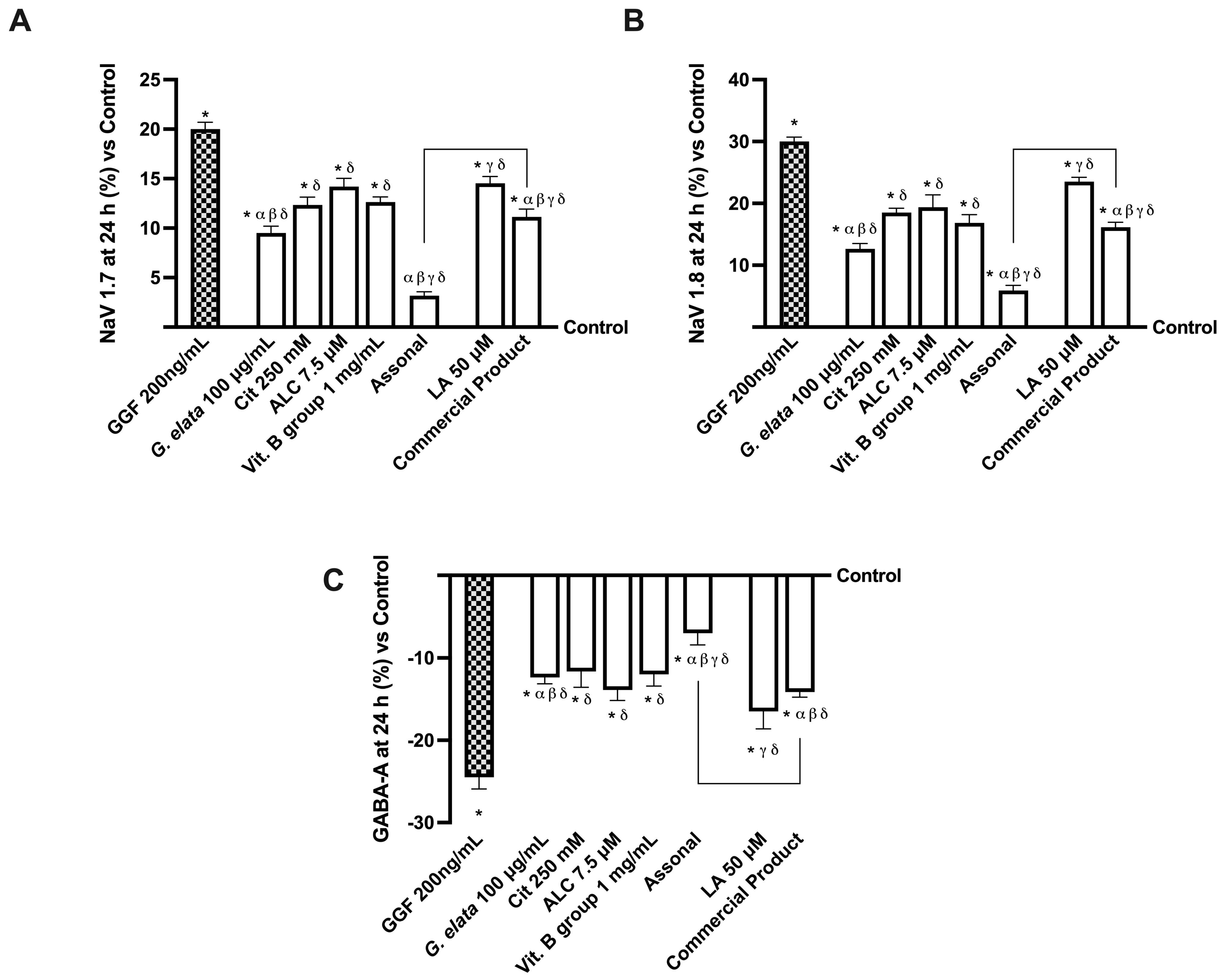
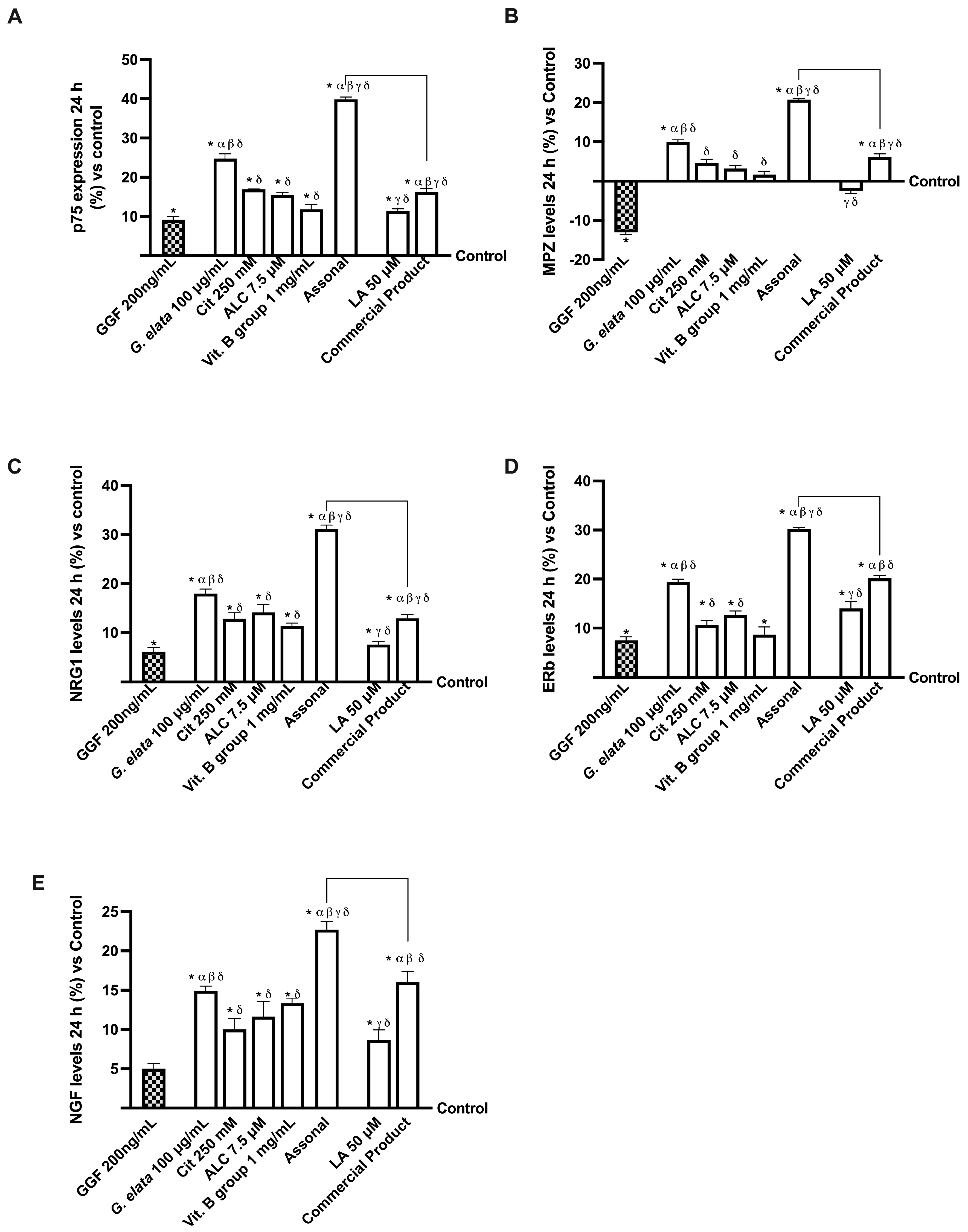
2.4. Effects of Single Components and Combination on an In Vitro Model of Peripheral Nervous Injury (PNI)
3. Discussion
3.1. Analysis of Assonal Effects on 3D In Vitro Intestinal Barrier Model
3.2. Analysis of Assonal Effects on a Physiological 3D EngNT Model in Physiological Conditions
3.3. Analysis of Assonal Effects on a 3D EngNT Model under PNI Conditions
4. Materials and Methods
4.1. Agents Preparation
4.2. Cell Culture
4.2.1. Epithelial Intestinal Cells
4.2.2. Schwann Cell
4.2.3. Neuronal Cell
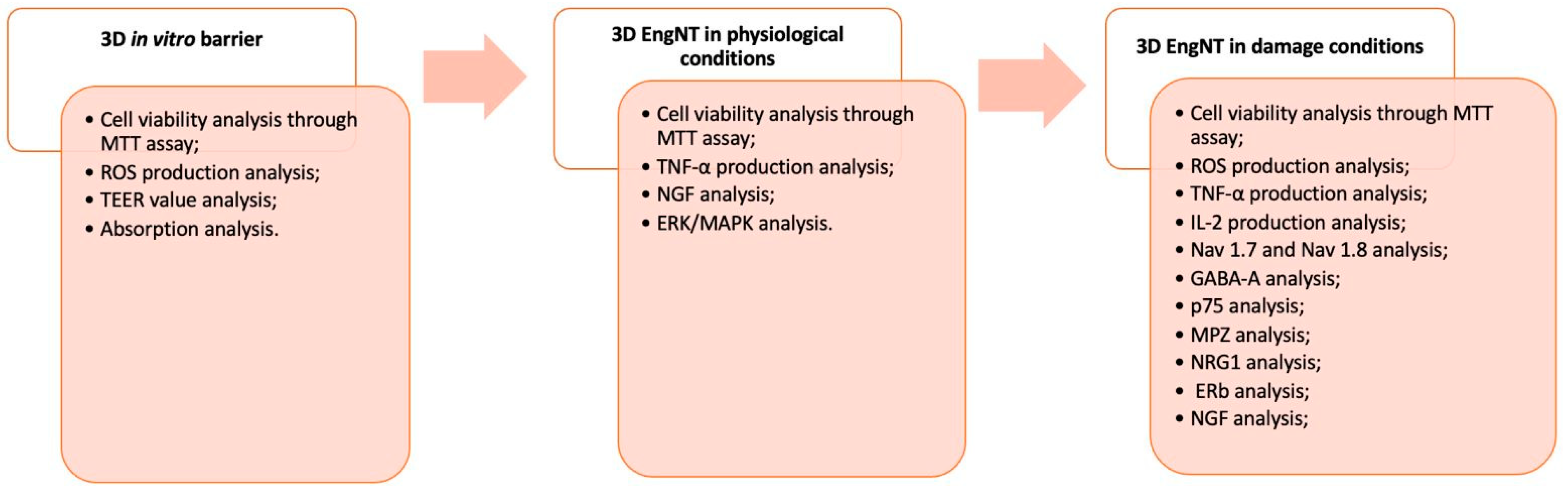
4.3. Experimental Protocol
4.4. Cell Viability
4.5. ROS Production
4.6. In Vitro Intestinal Barrier Model
- -
- Jmax: the maximum permeation rate;
- -
- [C]: the initial concentration of fluorescein;
- -
- Kt: the Michaelis–Menten constant.
4.7. TJs Analysis
4.8. 3D EngNT Co-Culture Setup
4.9. TNFα ELISA Assay
4.10. Human Beta-NGF Assay
4.11. ERK/MAPK Expression
4.12. Interleukin-2 ELISA Assay
4.13. NAV1.7 ELISA Assay
4.14. NAV1.8 ELISA Assay
4.15. γ-Aminobutyric Acid (GABA) ELISA Assay
4.16. p75 by NGFR ELISA Assay
4.17. Myelin Protein Zero (MPZ) ELISA Assay
4.18. (Neuregulin 1)NRG1 ELISA Assay
4.19. Estrogen Receptor β ELISA Assay
4.20. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D EngNT | 3D-engineered neural tissue |
| ADE | adverse drug event |
| ADR | adverse drug response |
| ALC | acetyl-L-carnitin |
| ANOVA | one-way analysis of variance |
| ATCC | American Type Culture Collection |
| BDNF | brain-derived neurotrophic factor |
| Cit | citicoline |
| CNS | central nervous system |
| DMEM-Adv | Dulbecco’s Modified Eagle’s Medium Advance |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| EMA | European Medicines Agency |
| FBS | fetal bovine serum |
| FDA | Food and Drug Administration |
| G. elata | Gastrodia elata Blume extract |
| GABA | γ-aminobutyric acid |
| GGF | glial growth factor 2 |
| GLTs | glucose transporters |
| HBA | p-hydroxy benzyl alcohol |
| LA | lipoic acid |
| MEM | Minimum Essential Medium |
| MPZ | myelin protein zero |
| NGF | nerve growth factor |
| NRG1 | neuregulin 1 |
| OS | oxidative stress |
| PN | peripheral neuropathy |
| PNI | Peripheral Nervous Injury |
| PNS | peripheral nervous system |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| RPMI-Adv | Roswell Park Memorial Institute-1640 Advanced |
| TEER | transepithelial electrical resistance |
| TJ | tight junction |
| Vit. B group | Vitamin of group B |
| ZO-1 | Zona Occludens 1 |
Appendix A
References
- Turk, D.C.; Melzack, R. The measurement of pain and the assessment of people experiencing pain. In Handbook of Pain Assessment, 3rd ed.; Guilford Press: New York, NY, USA, 2011; pp. 3–16. [Google Scholar]
- Murray, C.B.; de la Vega, R.; Murphy, L.K.; Kashikar-Zuck, S.; Palermo, T.M. The prevalence of chronic pain in young adults: A systematic review and meta-analysis. Pain 2022, 163, e972–e984. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M. Role of the immune system in neuropathic pain. Scand. J. Pain 2019, 20, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Marchettini, P.; Lacerenza, M.; Mauri, E.; Marangoni, C. Painful peripheral neuropathies. Curr. Neuropharmacol. 2006, 4, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Hulsebosch, C.E.; Hains, B.C.; Crown, E.D.; Carlton, S.M. Mechanisms of chronic central neuropathic pain after spinal cord injury. Brain Res. Rev. 2009, 60, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Ilari, S.; Giancotti, L.A.; Lauro, F.; Dagostino, C.; Gliozzi, M.; Malafoglia, V.; Sansone, L.; Palma, E.; Tafani, M.; Russo, M.A.; et al. Antioxidant modulation of sirtuin 3 during acute inflammatory pain: The ROS control. Pharmacol. Res. 2020, 157, 104851. [Google Scholar] [CrossRef] [PubMed]
- Tracey, D.J.; Walker, J.S. Pain due to nerve damage: Are inflammatory mediators involved? Inflamm. Res. 1995, 44, 407–411. [Google Scholar] [CrossRef]
- Mishra, G.; Singh, P.; Molla, M.; Yimer, Y.S.; Ewunetie, A.; Tadesse, T.Y.; Ayele, T.M.; Kefale, B. Nutraceuticals: A source of benefaction for neuropathic pain and fibromyalgia. J. Funct. Foods 2022, 97, 105260. [Google Scholar] [CrossRef]
- Papanas, N.; Ziegler, D. Efficacy of α-lipoic acid in diabetic neuropathy. Expert Opin. Pharmacother. 2014, 15, 2721–2731. [Google Scholar] [CrossRef]
- Gatti, M.; Ippoliti, I.; Poluzzi, E.; Antonazzo, I.C.; Moro, P.A.; Moretti, U.; Menniti-Ippolito, F.; Mazzanti, G.; De Ponti, F.; Raschi, E. Assessment of adverse reactions to α-lipoic acid containing dietary supplements through spontaneous reporting systems. Clin. Nutr. 2021, 40, 1176–1185. [Google Scholar] [CrossRef]
- Cassanego, G.; Rodrigues, P.; De Freitas Bauermann, L.; Trevisan, G. Evaluation of the analgesic effect of ɑ-lipoic acid in treating pain disorders: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2022, 177, 106075. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Liu, M.F.; Ji, X.Y.; Jiang, C.R.; Li, Z.L.; OuYang, B. Gastrodin combined with rhynchophylline inhibits cerebral ischaemia-induced inflammasome activation via upregulating miR-21-5p and miR-331-5p. Life Sci. 2019, 239, 116935. [Google Scholar] [CrossRef]
- Sun, X.; Jia, B.; Sun, J.; Lin, J.; Lu, B.; Duan, J.; Li, C.; Wang, Q.; Zhang, X.; Tan, M.; et al. Gastrodia elata Blume: A review of its mechanisms and functions on cardiovascular systems. Fitoterapia 2023, 167, 105511. [Google Scholar] [CrossRef]
- Zhan, H.D.; Zhou, H.Y.; Sui, Y.P.; Du, X.L.; Wang, W.H.; Dai, L.; Sui, F.; Huo, H.R.; Jiang, T.L. The rhizome of Gastrodia elata Blume—An ethnopharmacological review. J. Ethnopharmacol. 2016, 189, 361–385. [Google Scholar] [CrossRef]
- Wu, J.; Wu, B.; Tang, C.; Zhao, J. Analytical Techniques and Pharmacokinetics of Gastrodia elata Blume and Its Constituents. Molecules 2017, 22, 1137. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, J.; Peng, M.; Meng, H.; Ma, H.; Cai, P.; Xu, Y.; Zhao, Q.; Si, G. A Review on Central Nervous System Effects of Gastrodin. Front. Pharmacol. 2018, 9, 24. [Google Scholar] [CrossRef]
- Wang, X.D.; Zeng, S. The transport of gastrodin in Caco-2 cells and uptake in Bcap37 and Bcap37/MDR1 cells. Yao Xue Xue Bao = Acta Pharm. Sin. 2010, 45, 1497–1502. [Google Scholar]
- Cai, Z.; Huang, J.; Luo, H.; Lei, X.; Yang, Z.; Mai, Y.; Liu, Z. Role of glucose transporters in the intestinal absorption of gastrodin, a highly water-soluble drug with good oral bioavailability. J. Drug Target. 2013, 21, 574–580. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, G.; Zeng, S. Pharmacokinetics of Gastrodin in rat plasma and CSF after i.n. and i.v. Int. J. Pharm. 2007, 341, 20–25. [Google Scholar] [CrossRef]
- Huang, P.; Wan, H.; Shao, C.; Li, C.; Zhang, L.; He, Y. Recent Advances in Chinese Herbal Medicine for Cerebral Ischemic Reperfusion Injury. Front. Pharmacol. 2022, 12, 688596. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Chen, Y.; Wu, W.; Feng, X.; Du, K. Gastrodia elata Blume Polysaccharides Attenuate Vincristine-Evoked Neuropathic Pain through the Inhibition of Neuroinflammation. Mediat. Inflamm. 2021, 2021, 9965081. [Google Scholar] [CrossRef]
- Roohi-Azizi, M.; Torkaman-Boutorabi, A.; Akhondzadeh, S.; Nejatisafa, A.A.; Sadat-Shirazi, M.S.; Zarrindast, M.R. Influence of citicoline on citalopram-induced antidepressant activity in depressive-like symptoms in male mice. Physiol. Behav. 2018, 195, 151–157. [Google Scholar] [CrossRef]
- Hurtado, O.; Moro, M.A.; Cárdenas, A.; Sánchez, V.; Fernández-Tomé, P.; Leza, J.C.; Lorenzo, P.; Secades, J.J.; Lozano, R.; Dávalos, A.; et al. Neuroprotection afforded by prior citicoline administration in experimental brain ischemia: Effects on glutamate transport. Neurobiol. Dis. 2005, 18, 336–345. [Google Scholar] [CrossRef]
- Sarzi-Puttini, P.; Giorgi, V.; Di Lascio, S.; Fornasari, D. Acetyl-L-carnitine in chronic pain: A narrative review. Pharmacol. Res. 2021, 173, 105874. [Google Scholar] [CrossRef]
- Niafar, M.; Hai, F.; Porhomayon, J.; Nader, N.D. The role of metformin on vitamin B12 deficiency: A meta-analysis review. Intern. Emerg. Med. 2015, 10, 93–102. [Google Scholar] [CrossRef]
- Sil, A.; Kumar, H.; Mondal, R.D.; Anand, S.S.; Ghosal, A.; Datta, A.; Sawant, S.V.; Kapatkar, V.; Kadhe, G.; Rao, S. A randomized, open labeled study comparing the serum levels of cobalamin after three doses of 500 mcg vs. a single dose methylcobalamin of 1500 mcg in patients with peripheral neuropathy. Korean J. Pain 2018, 31, 183–190. [Google Scholar] [CrossRef]
- Menezes, R.R.; Godin, A.M.; Rodrigues, F.F.; Coura, G.M.E.; Melo, I.S.F.; Brito, A.M.S.; Bertollo, C.M.; Paulino, T.P.; Rachid, M.A.; Machado, R.R.; et al. Thiamine and riboflavin inhibit production of cytokines and increase the anti-inflammatory activity of a corticosteroid in a chronic model of inflammation induced by complete Freund’s adjuvant. Pharmacol. Rep. 2017, 69, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Onk, D.; Mammadov, R.; Suleyman, B.; Cimen, F.K.; Cankaya, M.; Gul, V.; Altuner, D.; Senol, O.; Kadioglu, Y.; Malkoc, I.; et al. The effect of thiamine and its metabolites on peripheral neuropathic pain induced by cisplatin in rats. Exp. Anim. 2018, 67, 259–269. [Google Scholar] [CrossRef]
- Alemanno, F.; Ghisi, D.; Westermann, B.; Bettoni, A.; Fanelli, A.; La Colla, L.; Danelli, G.; Cesana, B.M. The use of vitamin B1 as a perineural adjuvant to middle interscalene block for postoperative analgesia after shoulder surgery. Acta Biomed. 2016, 87, 22–27. [Google Scholar] [PubMed]
- Hedaya, R. Five herbs plus thiamine reduce pain and improve functional mobility in patients with pain: A pilot study. Altern. Ther. Health Med. 2017, 23, 14–19. [Google Scholar] [PubMed]
- Baltrusch, S. The Role of Neurotropic B Vitamins in Nerve Regeneration. BioMed Res. Int. 2021, 2021, 9968228. [Google Scholar] [CrossRef]
- Ma, W.; Quirion, R. The ERK/MAPK pathway, as a target for the treatment of neuropathic pain. Expert Opin. Ther. Targets 2005, 9, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Levinson, S.R.; Luo, S.; Henry, M.A. The role of sodium channels in chronic pain. Muscle Nerve 2012, 46, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, K.M.; Hackshaw, K.V. Nutritional Supplements for the Treatment of Neuropathic Pain. Biomedicines 2021, 9, 674. [Google Scholar] [CrossRef] [PubMed]
- Casale, R.; Symeonidou, Z.; Ferfeli, S.; Micheli, F.; Scarsella, P.; Paladini, A. Food for special medical purposes and nutraceuticals for pain: A narrative review. Pain Ther. 2021, 10, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Smith, M.T. Neurotrophins and neuropathic pain: Role in pathobiology. Molecules 2015, 20, 10657–10688. [Google Scholar] [CrossRef]
- Pan, P.; Dobrowsky, R.T. Differential expression of neuregulin-1 isoforms and downregulation of erbin are associated with Erb B2 receptor activation in diabetic peripheral neuropathy. Acta Neuropathol. Commun. 2013, 1, 39. [Google Scholar] [CrossRef]
- Wang, C.; Hao, H.; He, K.; An, Y.; Pu, Z.; Gamper, N.; Zhang, H.; Du, X. Neuropathic Injury–Induced Plasticity of GABAergic System in Peripheral Sensory Ganglia. Front. Pharmacol. 2021, 12, 702218. [Google Scholar] [CrossRef]
- Barker, P.A.; Mantyh, P.; Arendt-Nielsen, L.; Viktrup, L.; Tive, L. Nerve Growth Factor Signaling and Its Contribution to Pain. J. Pain Res. 2020, 13, 1223–1241. [Google Scholar] [CrossRef]
- Packer, L.; Cadenas, E. Lipoic acid: Energy metabolism and redox regulation of transcription and cell signaling. J. Clin. Biochem. Nutr. 2011, 48, 26–32. [Google Scholar] [CrossRef]
- Lu, C.; Qu, S.; Zhong, Z.; Luo, H.; Lei, S.S.; Zhong, H.J.; Su, H.; Wang, Y.; Chong, C.M. The effects of bioactive components from the rhizome of. Front. Pharmacol. 2022, 13, 963327. [Google Scholar] [CrossRef]
- Molinari, C.; Ruga, S.; Farghali, M.; Galla, R.; Fernandez-Godino, R.; Clemente, N.; Uberti, F. Effects of a New Combination of Natural Extracts on Glaucoma-Related Retinal Degeneration. Foods 2021, 10, 1885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, M.; Liu, G.; Li, D.; Zhou, B. Oral absorption mechanism of the polysaccharides from Gastrodia elata Blume base on fluorescence labeling. Food Res. Int. 2021, 144, 110342. [Google Scholar] [CrossRef]
- Demirbas, S.; Stavchansky, S. Effects of A and dimethylsulfoxide on transepithelial transport of passively diffused drugs in the Caco-2 cell culture model. Int. J. Pharm. 2003, 251, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Albogami, S. The Potential Inhibitory Role of Acetyl-L-Acetyl-L-Carnitine on Proliferation, Migration, and Gene Expression in HepG2 and HT29 Human Adenocarcinoma Cell Lines. Curr. Issues Mol. Biol. 2023, 45, 2393–2408. [Google Scholar] [CrossRef]
- Soh, Y.; Lee, D.H.; Won, C.W. Association between Vitamin B12 levels and cognitive function in the elderly Korean population. Medicine 2020, 99, e21371. [Google Scholar] [CrossRef] [PubMed]
- Malouf, R.; Grimley Evans, J. The effect of vitamin B6 on cognition. Cochrane Database Syst. Rev. 2003, 4, CD004393. [Google Scholar] [CrossRef]
- Molinari, C.; Morsanuto, V.; Ghirlanda, S.; Ruga, S.; Notte, F.; Gaetano, L.; Uberti, F. Role of Combined Lipoic Acid and Vitamin D3 on Astrocytes as a Way to Prevent Brain Ageing by Induced Oxidative Stress and Iron Accumulation. Oxidative Med. Cell. Longev. 2019, 2019, 2843121. [Google Scholar] [CrossRef]
- Lea, T. Caco-2 cell line. In The Impact of Food Bioactives on Health; Springer: Cham, Switzerland, 2015; pp. 103–111. [Google Scholar]
- DiMarco, R.L.; Hunt, D.R.; Dewi, R.E.; Heilshorn, S.C. Improvement of paracellular transport in the Caco-2 drug screening model using protein-engineered substrates. Biomaterials 2017, 129, 152–162. [Google Scholar] [CrossRef]
- Galla, R.; Ruga, S.; Aprile, S.; Ferrari, S.; Brovero, A.; Grosa, G.; Molinari, C.; Uberti, F. New Hyaluronic Acid from Plant Origin to Improve Joint Protection-An In Vitro Study. Int. J. Mol. Sci. 2022, 23, 8114. [Google Scholar] [CrossRef]
- Ceriotti, L.; Meloni, M. La valutazione dell’assorbimento intestinale in vitro. L’integratore Nutr. 2014, 17, 62–65. [Google Scholar]
- Rayner, M.L.D.; Laranjeira, S.; Evans, R.E.; Shipley, R.J.; Healy, J.; Phillips, J.B. Developing an In Vitro Model to Screen Drugs for Nerve Regeneration. Anat. Rec. 2018, 301, 1628–1637. [Google Scholar] [CrossRef]
- Chua, P.; Lim, W.K. Optimisation of a PC12 cell-based in vitro stroke model for screening neuroprotective agents. Sci. Rep. 2021, 11, 8096. [Google Scholar] [CrossRef] [PubMed]
- Lahiani, A.; Brand-Yavin, A.; Yavin, E.; Lazarovici, P. Neuroprotective Effects of Bioactive Compounds and MAPK Pathway Modulation in “Ischemia”—Stressed PC12 Pheochromocytoma Cells. Brain Sci. 2018, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Christides, T.; Wray, D.; McBride, R.; Fairweather, R.; Sharp, P. Iron bioavailability from commercially available iron supplements. Eur. J. Nutr. 2015, 54, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Fda.Gov. Available online: https://www.fda.gov/media/117974/download (accessed on 4 September 2023).
- Ema.Eu. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/ich-m9-biopharmaceutics-classification-system-based-biowaivers-step-2b-first-version_en.pdf (accessed on 4 September 2023).
- Guha, S.; Alvarez, S.; Majumder, K. Transport of Dietary Anti-Inflammatory Peptide, γ-Glutamyl Valine (γ-EV), across the Intestinal Caco-2 Monolayer. Nutrients 2021, 13, 1448. [Google Scholar] [CrossRef] [PubMed]
- Hubatsch, I.; Ragnarsson, E.G.; Artursson, P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat. Protoc. 2007, 2, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Morsanuto, V.; Galla, R.; Molinari, C.; Uberti, F. A New Palmitoylethanolamide Form Combined with Antioxidant Molecules to Improve Its Effectivess on Neuronal Aging. Brain Sci. 2020, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Schuh, C.M.A.P.; Day, A.G.E.; Redl, H.; Phillips, J. An Optimized Collagen-Fibrin Blend Engineered Neural Tissue Promotes Peripheral Nerve Repair. Tissue Eng. Part A 2018, 24, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Muangsanit, P.; Day, A.; Dimiou, S.; Ataç, A.F.; Kayal, C.; Park, H.; Nazhat, S.N.; Phillips, J.B. Rapidly formed stable and aligned dense collagen gels seeded with Schwann cells support peripheral nerve regeneration. J. Neural Eng. 2020, 17, 046036. [Google Scholar] [CrossRef]
- Uberti, F.; Ruga, S.; Farghali, M.; Galla, R.; Molinari, C. A Combination of α-Lipoic Acid (ALA) and Palmitoylethanolamide (PEA) Blocks Endotoxin-Induced Oxidative Stress and Cytokine Storm: A Possible Intervention for COVID-19. J. Diet. Suppl. 2023, 20, 133–155. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Kadoya, K.; Kawamura, D.; Iwasaki, N. Evidence for cell-contact factor involvement in neurite outgrowth of dorsal root ganglion neurons stimulated by Schwann cells. Exp. Physiol. 2019, 104, 1447–1454. [Google Scholar] [CrossRef]
- Du, Y.; Luan, J.; Jiang, R.P.; Liu, J.; Ma, Y. Myrcene exerts anti-asthmatic activity in neonatal rats via modulating the matrix remodeling. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420954948. [Google Scholar] [CrossRef]
- Han, C.; Estacion, M.; Huang, J.; Vasylyev, D.; Zhao, P.; Dib-Hajj, S.D.; Waxman, S.G. Human Na(v)1.8: Enhanced persistent and ramp currents contribute to distinct firing properties of human DRG neurons. J. Neurophysiol. 2015, 113, 3172–3185. [Google Scholar] [CrossRef] [PubMed]
- Al Suhaibani, A.; Ben Bacha, A.; Alonazi, M.; Bhat, R.S.; El-Ansary, A. Testing the combined effects of probiotics and prebiotics against neurotoxic effects of propionic acid orally administered to rat pups. Food Sci. Nutr. 2021, 9, 4440–4451. [Google Scholar] [CrossRef] [PubMed]
- Ruga, S.; Galla, R.; Ferrari, S.; Invernizzi, M.; Uberti, F. Novel Approach to the Treatment of Neuropathic Pain Using a Combination with Palmitoylethanolamide and Equisetum arvense L. in an In Vitro Study. Int. J. Mol. Sci. 2023, 24, 5503. [Google Scholar] [CrossRef] [PubMed]
- Rzemieniec, J.; Litwa, E.; Wnuk, A.; Lason, W.; Gołas, A.; Krzeptowski, W.; Kajta, M. Neuroprotective action of raloxifene against hypoxia-induced damage in mouse hippocampal cells depends on ERα but not ERβ or GPR30 signalling. J. Steroid Biochem. Mol. Biol. 2015, 146, 26–37. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrari, S.; Mulè, S.; Galla, R.; Brovero, A.; Genovese, G.; Molinari, C.; Uberti, F. Effects of Nutraceutical Compositions Containing Rhizoma Gastrodiae or Lipoic Acid in an In Vitro Induced Neuropathic Pain Model. Int. J. Mol. Sci. 2024, 25, 2376. https://doi.org/10.3390/ijms25042376
Ferrari S, Mulè S, Galla R, Brovero A, Genovese G, Molinari C, Uberti F. Effects of Nutraceutical Compositions Containing Rhizoma Gastrodiae or Lipoic Acid in an In Vitro Induced Neuropathic Pain Model. International Journal of Molecular Sciences. 2024; 25(4):2376. https://doi.org/10.3390/ijms25042376
Chicago/Turabian StyleFerrari, Sara, Simone Mulè, Rebecca Galla, Arianna Brovero, Giulia Genovese, Claudio Molinari, and Francesca Uberti. 2024. "Effects of Nutraceutical Compositions Containing Rhizoma Gastrodiae or Lipoic Acid in an In Vitro Induced Neuropathic Pain Model" International Journal of Molecular Sciences 25, no. 4: 2376. https://doi.org/10.3390/ijms25042376
APA StyleFerrari, S., Mulè, S., Galla, R., Brovero, A., Genovese, G., Molinari, C., & Uberti, F. (2024). Effects of Nutraceutical Compositions Containing Rhizoma Gastrodiae or Lipoic Acid in an In Vitro Induced Neuropathic Pain Model. International Journal of Molecular Sciences, 25(4), 2376. https://doi.org/10.3390/ijms25042376