
In Silico Analysis of Genes Associated with the Pathogenesis of Odontogenic Keratocyst
 ,
,  , and
, and
Abstract
1. Introduction
2. Results
2.1. GO and KEGG Enrichment
2.2. IPP Network and Modular Analysis
2.3. Post-Functional Enrichment of Candidate Genes
3. Discussion
4. Materials and Methods
- GO and KEGG Enrichment
- Protein–protein interaction (PPI)
- Post-functional enrichment of candidate genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Speight, P.; Devilliers, P.; Li, T.J.; Odell, E.W.; Wright, J.M. Odontogenic keratocyst. In WHO Classification of Head and Neck Tumours, 4th ed.; El-Naggar, A.K., Chan, J.K.C., Grandis, J.R., Takata, T., Slootweg, P.J., Eds.; IARC: Lyon, France, 2017; pp. 235–236. [Google Scholar]
- Ram, H.; Mohammad, S.; Husain, N.; Gupta, S.; Kumar, A. Bilateral odontogenic keratocyst of the mandible. J. Maxillofac. Oral Surg. 2014, 13, 341–345. [Google Scholar] [CrossRef]
- Priya, K.; Karthikeyan, P.; Nirmal Coumare, V. Odontogenic Keratocyst: A Case Series of five Patients. Indian J. Otolaryngol. Head Neck Surg. 2014, 66, 17–21. [Google Scholar] [CrossRef][Green Version]
- Portes, J.; Cunha, K.S.G.; da Silva, L.E.; da Silva, A.K.F.; Conde, D.C.; Silva Junior, A. Computerized Evaluation of the Immunoexpression of Ki-67 Protein in Odontogenic Keratocyst and Dentigerous Cyst. Head Neck Pathol. 2020, 14, 598–605. [Google Scholar] [CrossRef]
- Kitisubkanchana, J.; Reduwan, N.H.; Poomsawat, S.; Pornprasertsuk-Damrongsri, S.; Wongchuensoontorn, C. Odontogenic keratocyst and ameloblastoma: Radiographic evaluation. Oral Radiol. 2021, 37, 55–65. [Google Scholar] [CrossRef]
- Gutiérrez-Patiño, P.A.; Rivadeneyra-Rodriguez, A. Queratoquiste odontogenico recurrente en paciente con sindrome de Gorlin-Goltz. Rev. Estomatol. Herediana 2020, 30, 53–62. [Google Scholar] [CrossRef]
- Valdivia, A.D.C.M.; Ramos-Ibarra, M.L.; Franco-Barrera, M.J.; Arias-Ruiz, L.F.; García-Cruz, J.M.; Torres-Bugarín, O. What is Currently Known about Odontogenic Keratocysts? Oral Health Prev. Dent. 2022, 20, 321–330. [Google Scholar] [PubMed]
- Moctezuma-Bravo, G.S.; Magallanes-González, E. Study of 103 cases of odontogenic cysts. Rev. Med. Inst. Mex. Seguro Soc. 2009, 47, 493–496. [Google Scholar] [PubMed]
- Gaitán-Cepeda, L.A.; Quezada-Rivera, D.; Tenorio-Rocha, F.; Leyva-Huerta, E.R. Reclassification of odontogenic keratocyst as tumour. Impact on the odontogenic tumours prevalence. Oral Dis. 2010, 16, 185–187. [Google Scholar] [CrossRef]
- Kammer, P.V.; Mello, F.W.; Rivero, E.R.C. Comparative analysis between developmental and inflammatory odontogenic cysts: Retrospective study and literature review. Oral Maxillofac. Surg. 2020, 24, 73–84. [Google Scholar] [CrossRef]
- Borghesi, A.; Nardi, C.; Giannitto, C.; Tironi, A.; Maroldi, R.; Di Bartolomeo, F.; Preda, L. Odontogenic keratocyst: Imaging features of a benign lesion with an aggressive behaviour. Insights Imaging 2018, 9, 883–897. [Google Scholar] [CrossRef] [PubMed]
- de França, G.M.; da Silva, L.B.A.; Mafra, R.P.; da Silva, W.R.; de Lima, K.C.; Galvão, H.C. Recurrence-free survival and prognostic factors of odontogenic keratocyst: A single-center retrospective cohort. Eur. Arch. Otorhinolaryngol. 2021, 278, 1223–1231. [Google Scholar] [CrossRef]
- Onodera, S.; Nakamura, Y.; Azuma, T. Gorlin Syndrome: Recent Advances in Genetic Testing and Molecular and Cellular Biological Research. Int. J. Mol. Sci. 2020, 21, 7559. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Zhang, J.Y.; Li, X.F.; Luo, H.Y.; Chen, F.; Li, T.J. PTCH1 gene mutations in Keratocystic odontogenic tumors: A study of 43 Chinese patients and a systematic review. PLoS ONE 2013, 8, e77305. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.R.; Guimarães, M.M.; Guimarães, A.L.; Diniz, M.G.; Gomes, C.C.; Brito, J.A.; Gomez, R.S. Methylation of P16, P21, P27, RB1 and P53 genes in odontogenic keratocysts. J. Oral Pathol. Med. 2009, 38, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Kadashetti, V.; Patil, N.; Datkhile, K.; Kanetakar, S.; Shivakumar, K.M. Analysis of expression of p53, p63 and proliferating cell nuclear antigen proteins in odontogenic keratocyst: An immunohistochemical study. J. Oral Maxillofac. Pathol. 2020, 24, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Mascitti, M.; Togni, L.; Balercia, A.; Balercia, P.; Rubini, C.; Santarelli, A. p53-Family Proteins in Odontogenic Cysts: An Immunohistochemical Study. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.M.; Nagrash, O.M.; Safadi, R.A. Maspin, Syndecan-1, and Ki-67 in the Odontogenic Keratocyst: An Immunohistochemical Analysis. Int. J. Dent. 2020, 2020, 7041520. [Google Scholar] [CrossRef] [PubMed]
- Li, T.J.; Browne, R.M.; Matthews, J.B. Epithelial cell proliferation in odontogenic keratocysts: A comparative immunocytochemical study of Ki67 in simple, recurrent and basal cell naevus syndrome (BCNS)-associated lesions. J. Oral Pathol. Med. 1995, 24, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Li, T.J.; Browne, R.M.; Matthews, J.B. Quantification of PCNA+ cells within odontogenic jaw cyst epithelium. J. Oral Pathol. Med. 1994, 23, 184–189. [Google Scholar] [CrossRef]
- Kadlub, N.; Coudert, A.; Gatibelza, M.E.; El Houmami, N.; Soufir, N.; Ruhin-Poncet, B.; L’hermine, A.C.; Berdal, A.; Vazquez, M.P.; Descroix, V.; et al. PTCH1 mutation and local aggressiveness of odontogenic keratocystic tumors in children: Is there a relationship? Hum. Pathol. 2013, 44, 1071–1078. [Google Scholar] [CrossRef]
- Pavelić, B.; Levanat, S.; Crnić, I.; Kobler, P.; Anić, I.; Manojlović, S.; Sutalo, J. PTCH gene altered in dentigerous cysts. J. Oral Pathol. Med. 2001, 30, 569–576. [Google Scholar] [CrossRef]
- Kuźniarz, K.; Luchowska-Kocot, D.; Tomaszewski, T.; Kurzepa, J. Role of matrix metalloproteinases and their tissue inhibitors in the pathological mechanisms underlying maxillofacial cystic lesions. Biomed. Rep. 2021, 15, 65. [Google Scholar] [CrossRef]
- Wahlgren, J.; Väänänen, A.; Teronen, O.; Sorsa, T.; Pirilä, E.; Hietanen, J.; Maisi, P.; Tjäderhane, L.; Salo, T. Laminin-5 gamma 2 chain is colocalized with gelatinase-A (MMP-2) and collagenase-3 (MMP-13) in odontogenic keratocysts. J. Oral Pathol. Med. 2003, 32, 100–107. [Google Scholar] [CrossRef]
- Poomsawat, S.; Punyasingh, J.; Vejchapipat, P. Expression of basement membrane components in odontogenic tumors. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2007, 104, 666–675. [Google Scholar] [CrossRef]
- Poomsawat, S.; Punyasingh, J.; Weerapradist, W. Expression of basement membrane components in odontogenic cysts. Oral Dis. 2006, 12, 290–296. [Google Scholar] [CrossRef]
- de Oliveira, M.D.; de Miranda, J.L.; de Amorim, R.F.; de Souza, L.B.; de Almeida Freitas, R. Tenascin and fibronectin expression in odontogenic cysts. J. Oral Pathol. Med. 2004, 33, 354–359. [Google Scholar] [CrossRef]
- Cota, J.E.; Spadigam, A.; Dhupar, A. Detection of Type VII collagen in odontogenic keratocyst: An immunohistochemical study. J. Clin. Exp. Dent. 2019, 11, e310–e314. [Google Scholar] [CrossRef]
- Loreto, C.; Polizzi, A.; Filetti, V.; Pannone, G.; Dos Santos, J.N.; Venezia, P.; Leonardi, R.; Isola, G. Expression of Matrix Metalloproteinases 7 and 9, Desmin, Alpha-Smooth Muscle Actin and Caldesmon, in Odontogenic Keratocyst Associated with NBCCS, Recurrent and Sporadic Keratocysts. Biomolecules 2022, 12, 775. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Zhao, D.; Yin, Z.; Liu, P. BST-2 binding with cellular MT1-MMP blocks cell growth and migration via decreasing MMP2 activity. J. Cell Biochem. 2012, 113, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- de França, G.M.; Felipe Junior, J.; de Freitas, C.T.S.; de Lucena, H.F.; de Andrade, A.L.D.L.; Galvão, H.C. Langerhans Cells Correlate With Macrophages for Defense Mechanisms in the Atrophic Epithelium of Radicular Cysts. Appl. Immunohistochem. Mol. Morphol. 2021, 29, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Hadziabdic, N.; Kurtovic-Kozaric, A.; Frkatovic, A.; Smajlovic, S.; Letra, A. Quantitative Analysis of CCL5 and ep300 in Periapical Inflammatory Lesions. Acta Med. Acad. 2019, 48, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Zhong, N.N.; Li, S.R.; Man, Q.W.; Liu, B. Identification of Immune Infiltration in Odontogenic Keratocyst by Integrated Bioinformatics Analysis. BMC Oral Health 2023, 23, 454. [Google Scholar] [CrossRef] [PubMed]
- Man, Q.W.; Li, R.F.; Liu, J.F.; Zhao, Y.; Liu, B. Single-cell analysis reveals immune cellular components in odontogenic keratocysts. Oral Dis. 2023, 29, 3420–3432. [Google Scholar] [CrossRef] [PubMed]
- Alsaegh, M.A.; Mahmoud, O.; Varma, S.R.; Zhu, S. The Prevalence of EBV and KSHV in Odontogenic Lesions. Int. Dent. J. 2023, 73, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A.; Campadelli-Fiume, G.; Mocarski, E.; Moore, P.S.; Roizman, B.; Whitley, R.; Yamanishi, K. Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Iftode, N.; Rădulescu, M.A.; Aramă, Ș.S.; Aramă, V. Update on Kaposi sarcoma-associated herpesvirus (KSHV or HHV8)—Review. Rom. J. Intern. Med. 2020, 58, 199–208. [Google Scholar] [CrossRef]
- Hakeem, A.; Fitzpatrick, S.G.; Gonsalves, C.A.; Isom, J.; Islam, M.N.; Bhattacharyya, I.; Cohen, D.M.; Drew, P.A. p16INK4a as a proliferation marker unrelated to HPV expression in odontogenic cysts and tumors. J. Oral Pathol. Med. 2020, 49, 72–81. [Google Scholar] [CrossRef]
- Wang, S.; Hong, Y.; Qu, J.; Zhang, J.; Zhang, Y.; Zhai, J.; Li, T. PTCH/SMO gene mutations in odontogenic keratocysts and drug interventions. J. Oral Pathol. Med. 2023, 52, 867–876. [Google Scholar] [CrossRef]
- Noguchi, K.; Wakai, K.; Kiyono, T.; Kawabe, M.; Yoshikawa, K.; Hashimoto-Tamaoki, T.; Kishimoto, H.; Nakano, Y. Molecular analysis of keratocystic odontogenic tumor cell lines derived from sporadic and basal cell nevus syndrome patients. Int. J. Oncol. 2017, 51, 1731–1738. [Google Scholar] [CrossRef]
- DAVID Bioinformatics Resources 6.8. Available online: https://david.ncifcrf.gov/ (accessed on 17 September 2023).
- STRING. Available online: https://string-db.org/ (accessed on 17 September 2023).


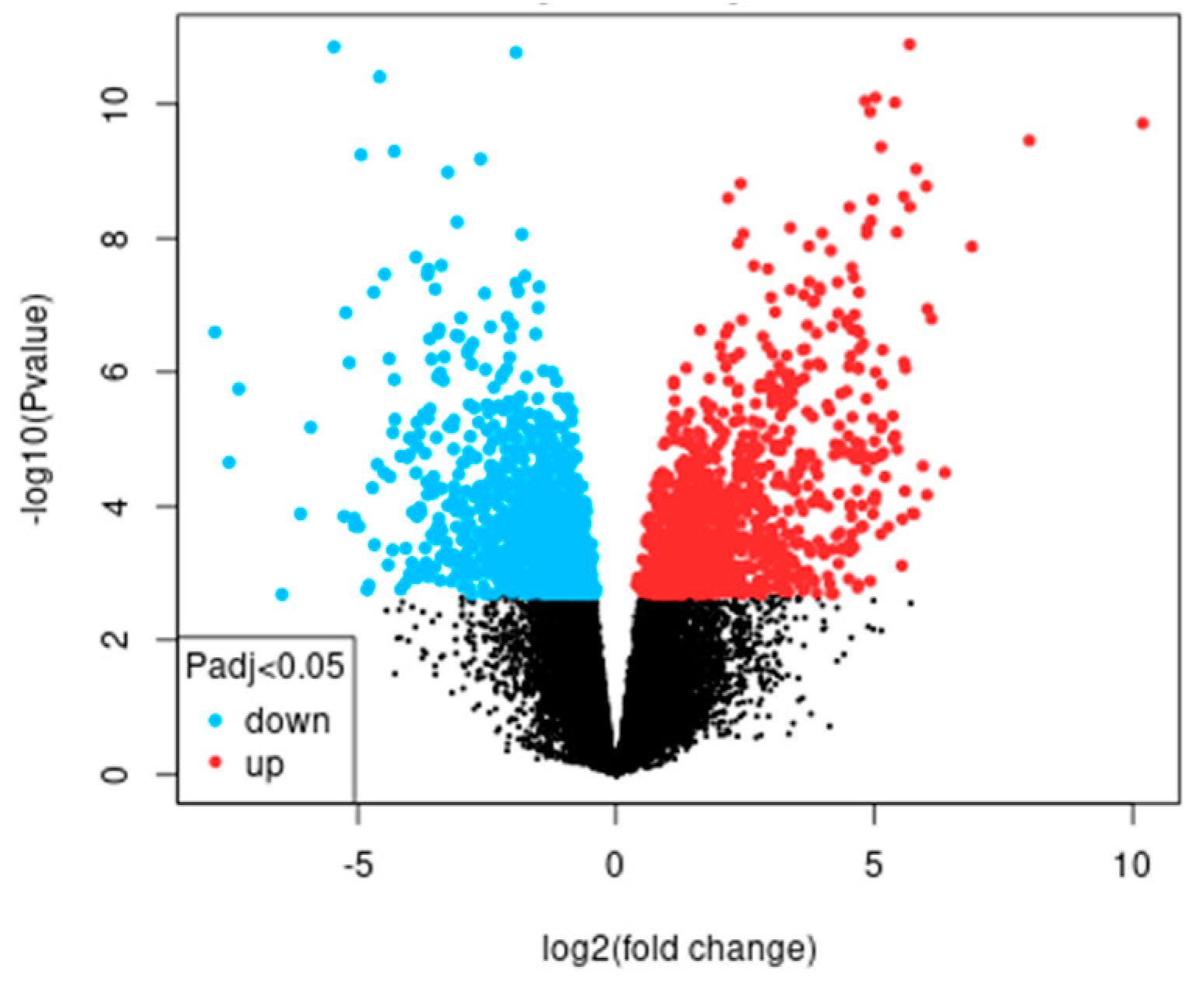{kind=link}
| Category | Terms | Genes Number | p-Value | FDR |
|---|---|---|---|---|
| GO | GO:0007155~cell adhesión | 69 | 1.16 × 10−27 | 3.07 × 10−24 |
| GO | GO:0030199~collagen fibril organization | 27 | 5.47 × 10−23 | 7.23 × 10−20 |
| GO | GO:0030198~extracellular matrix organization | 35 | 6.05 × 10−20 | 5.34 × 10−17 |
| GO | GO:0001501~skeletal system development | 21 | 1.46 × 10−10 | 9.67 × 10−8 |
| GO | GO:0001649~osteoblast differentiation | 19 | 2.87 × 10−9 | 1.52 × 10−6 |
| GO | GO:0008284~positive regulation of cell proliferation | 39 | 1.14 × 10−8 | 4.83 × 10−6 |
| GO | GO:0007411~axon guidance | 22 | 1.35 × 10−8 | 4.83 × 10−6 |
| GO | GO:0001503~ossification | 15 | 1.46 × 10−8 | 4.83 × 10−6 |
| GO | GO:0071711~basement membrane organization | 9 | 3.42 × 10−8 | 1.01 × 10−5 |
| GO | GO:0071230~cellular response to amino acid stimulus | 12 | 2.04 × 10−7 | 4.97 × 10−5 |
| KEGG | hsa04974:Protein digestion and absorption | 23 | 8.90 × 10−14 | 2.18 × 10−11 |
| KEGG | hsa04512:ECM–receptor interaction | 17 | 3.54 × 10−9 | 4.34 × 10−7 |
| KEGG | hsa04151:PI3K-Akt signaling pathway | 29 | 9.53 × 10−7 | 7.78 × 10−5 |
| KEGG | hsa04510:Focal adhesion | 21 | 1.35 × 10−6 | 8.27 × 10−5 |
| KEGG | hsa05205:Proteoglycans in cancer | 18 | 8.00 × 10−5 | 0.00391793 |
| KEGG | hsa04933:AGE-RAGE signaling pathway in diabetic complications | 12 | 1.29 × 10−4 | 0.00527679 |
| KEGG | hsa05200:Pathways in cancer | 31 | 2.66 × 10−4 | 0.00932276 |
| KEGG | hsa04310:Wnt signaling pathway | 15 | 3.91 × 10−4 | 0.01196999 |
| KEGG | hsa05165:Human papillomavirus infection | 22 | 5.28 × 10−4 | 0.01437369 |
| KEGG | hsa04360:Axon guidance | 15 | 7.29 × 10−4 | 0.01787028 |
| Category | Terms | Genes Number | p-Value | FDR |
| GO | GO:0045087~innate immune response | 37 | 7.88 × 10−11 | 1.64 × 10−7 |
| GO | GO:0051607~defense response to virus | 22 | 3.48 × 10−10 | 3.62 × 10−7 |
| GO | GO:0009615~response to virus | 15 | 3.65 × 10−9 | 2.53 × 10−6 |
| GO | GO:0006954~inflammatory response | 26 | 5.11 × 10−8 | 2.66 × 10−5 |
| GO | GO:0071346~cellular response to interferon-gamma | 12 | 7.13 × 10−7 | 2.62 × 10−4 |
| GO | GO:0045071~negative regulation of viral genome replication | 9 | 7.55 × 10−7 | 2.62 × 10−4 |
| GO | GO:0008544~epidermis development | 9 | 7.99 × 10−5 | 0.02205982 |
| GO | GO:0006955~immune response | 22 | 8.48 × 10−5 | 0.02205982 |
| GO | GO:0002503~peptide antigen assembly with MHC class II protein complex | 5 | 1.46 × 10−4 | 0.03076427 |
| GO | GO:0070106~interleukin−27-mediated signaling pathway | 4 | 1.48 × 10−4 | 0.03076427 |
| KEGG | hsa05164:Influenza A | 17 | 7.84 × 10−7 | 1.85 × 10−4 |
| KEGG | hsa05150:Staphylococcus aureus infection | 12 | 6.37 × 10−6 | 7.52 × 10−4 |
| KEGG | hsa05330:Allograft rejection | 8 | 1.41 × 10−5 | 0.00110822 |
| KEGG | hsa05145:Toxoplasmosis | 11 | 1.46 × 10−4 | 0.00864153 |
| KEGG | hsa05332:Graft-versus-host disease | 7 | 2.58 × 10−4 | 0.00994006 |
| KEGG | hsa04612:Antigen processing and presentation | 9 | 2.64 × 10−4 | 0.00994006 |
| KEGG | hsa04940:Type I diabetes mellitus | 7 | 2.95 × 10−4 | 0.00994006 |
| KEGG | hsa05169:Epstein–Barr virus infection | 14 | 4.22 × 10−4 | 0.01244118 |
| KEGG | hsa05310:Asthma | 6 | 4.81 × 10−4 | 0.01261624 |
| KEGG | hsa05160:Hepatitis C | 12 | 5.77 × 10−4 | 0.01361529 |
| Category | Genes |
|---|---|
| Genes derived from >2FC | COL6A3, FBLN1, DCN, THBS2, COL1A1, COL1A2, COL3A1, FN1, POSTN, COL5A1, LUM, COL4A1, COL6A1, COL6A2, NID1, COL5A2, COL14A1, COL15A1, COL8A1, FMOD, COL12A1, NID2, COL11A1, COL4A2, SERPINH1, FBN1, FBLN2, MMP2, CCN2, COL10A1, LAMA4, THBS1, COL16A1, COL18A1, LOXL1, ELN, ADAMTS2, COL21A1, TIMP3, CDH11, THY1, CXCL12, SPP1, PXDN, MMP13, COMP |
| Genes derived from <2FC | IRF1, CXCL10, OAS2, STAT1, MX1, OAS1, RSAD2, GBP2, GBP1, IFIH1, IFIT3, CCL5, PARP9, DDX60, CXCL9, GBP4, NLRC5, BST2, IFI27, TNF, GBP3, MX2, AIM2, SAMD9 |
| The MCODE algorithm was applied to enrich clusters of the IPP network and identify proteins that are densely connected (MCODE score > 4). | |
| ID | Terms | p-Value with Bonferroni Correction | Genes Number | Associated Genes |
|---|---|---|---|---|
| GO:0030198 | extracellular matrix organization | 5.623 × 10−42 | 31 | ADAMTS2, CCN2, COL10A1, COL11A1, COL12A1, COL14A1, COL15A1, COL16A1, COL18A1, COL1A1, COL1A2, COL3A1, COL4A1, COL4A2, COL5A1, COL5A2, COL8A1, COMP, ELN, FBLN1, FBLN2, FMOD, LOXL1, LUM, MMP13, MMP2, NID1, NID2, POSTN, PXDN, SERPINH1 |
| GO:0043062 | extracellular structure organization | 6.005 × 10−42 | 31 | ADAMTS2, CCN2, COL10A1, COL11A1, COL12A1, COL14A1, COL15A1, COL16A1, COL18A1, COL1A1, COL1A2, COL3A1, COL4A1, COL4A2, COL5A1, COL5A2, COL8A1, COMP, ELN, FBLN1, FBLN2, FMOD, LOXL1, LUM, MMP13, MMP2, NID1, NID2, POSTN, PXDN, SERPINH1 |
| GO:0045229 | external encapsulating structure organization | 7.033 × 10−42 | 31 | ADAMTS2, CCN2, COL10A1, COL11A1, COL12A1, COL14A1, COL15A1, COL16A1, COL18A1, COL1A1, COL1A2, COL3A1, COL4A1, COL4A2, COL5A1, COL5A2, COL8A1, COMP, ELN, FBLN1, FBLN2, FMOD, LOXL1, LUM, MMP13, MMP2, NID1, NID2, POSTN, PXDN, SERPINH1 |
| GO:0030199 | collagen fibril organization | 5.279 × 10−25 | 15 | ADAMTS2, COL11A1, COL12A1, COL14A1, COL1A1, COL1A2, COL3A1, COL5A1, COL5A2, COMP, FMOD, LOXL1, LUM, PXDN, SERPINH1 |
| GO:0035987 | endodermal cell differentiation | 1.849 × 10−15 | 10 | COL11A1, COL12A1, COL4A2, COL5A1, COL5A2, COL6A1, COL8A1, FN1, LAMA4, MMP2 |
| GO:0001706 | endoderm formation | 1.154 × 10−14 | 10 | COL11A1, COL12A1, COL4A2, COL5A1, COL5A2, COL6A1, COL8A1, FN1, LAMA4, MMP2 |
| GO:0007492 | endoderm development | 5.997 × 10−13 | 10 | COL11A1, COL12A1, COL4A2, COL5A1, COL5A2, COL6A1, COL8A1, FN1, LAMA4, MMP2 |
| GO:0001704 | formation of primary germ layer | 2.526 × 10−11 | 10 | COL11A1, COL12A1, COL4A2, COL5A1, COL5A2, COL6A1, COL8A1, FN1, LAMA4, MMP2 |
| GO:0071230 | cellular response to amino acid stimulus | 3.293 × 10−10 | 8 | COL16A1, COL1A1, COL1A2, COL3A1, COL4A1, COL5A2, COL6A1, MMP2 |
| GO:0043200 | response to amino acid | 4.674 × 10−10 | 9 | CCN2, COL16A1, COL1A1, COL1A2, COL3A1, COL4A1, COL5A2, COL6A1, MMP2 |
| ID | Terms | p-Value with Bonferroni Correction | Genes Number | Associated Genes |
|---|---|---|---|---|
| GO:0009615 | response to virus | 1.985 × 10−30 | 21 | AIM2, BST2, CCL5, CXCL10, CXCL9, DDX60, GBP1, GBP3, IFI27, IFIH1, IFIT3, IRF1, MX1, MX2, NLRC5, OAS1, OAS2, PARP9, RSAD2, STAT1, TNF |
| GO:0051607 | defense response to virus | 2.420 × 10−28 | 19 | AIM2, BST2, CXCL10, CXCL9, DDX60, GBP1, GBP3, IFI27, IFIH1, IFIT3, IRF1, MX1, MX2, NLRC5, OAS1, OAS2, PARP9, RSAD2, STAT1 |
| GO:0140546 | defense response to symbiont | 2.483 × 10−28 | 19 | AIM2, BST2, CXCL10, CXCL9, DDX60, GBP1, GBP3, IFI27, IFIH1, IFIT3, IRF1, MX1, MX2, NLRC5, OAS1, OAS2, PARP9, RSAD2, STAT1 |
| GO:0034341 | response to interferon-gamma | 4.479 × 10−16 | 11 | BST2, CCL5, GBP1, GBP2, GBP3, GBP4, IRF1, NLRC5, PARP9, STAT1, TNF |
| GO:0071346 | cellular response to interferon-gamma | 9.175 × 10−15 | 10 | CCL5, GBP1, GBP2, GBP3, GBP4, IRF1, NLRC5, PARP9, STAT1, TNF |
| GO:0071346 | cellular response to interferon-gamma | 9.175 × 10−15 | 10 | CCL5, GBP1, GBP2, GBP3, GBP4, IRF1, NLRC5, PARP9, STAT1, TNF |
| GO:0048525 | negative regulation of viral process | 4.501 × 10−14 | 9 | BST2, CCL5, IFIH1, MX1, OAS1, OAS2, RSAD2, STAT1, TNF |
| GO:0045071 | negative regulation of viral genome replication | 1.100 × 10−13 | 8 | BST2, CCL5, IFIH1, MX1, OAS1, OAS2, RSAD2, TNF |
| GO:0019079 | viral genome replication | 9.466 × 10−13 | 9 | BST2, CCL5, IFI27, IFIH1, MX1, OAS1, OAS2, RSAD2, TNF |
| GO:0045069 | regulation of viral genome replication | 3.289 × 10−12 | 8 | BST2, CCL5, IFIH1, MX1, OAS1, OAS2, RSAD2, TNF |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Martínez, C.M.; Legorreta-Villegas, I.; Mejía-Velázquez, C.P.; Portilla-Robertson, J.; Gaitán-Cepeda, L.A.; Paramo-Sánchez, J.T.; Chanes-Cuevas, O.A.; Alonso-Moctezuma, A.; Jacinto-Alemán, L.F. In Silico Analysis of Genes Associated with the Pathogenesis of Odontogenic Keratocyst. Int. J. Mol. Sci. 2024, 25, 2379. https://doi.org/10.3390/ijms25042379
Ramírez-Martínez CM, Legorreta-Villegas I, Mejía-Velázquez CP, Portilla-Robertson J, Gaitán-Cepeda LA, Paramo-Sánchez JT, Chanes-Cuevas OA, Alonso-Moctezuma A, Jacinto-Alemán LF. In Silico Analysis of Genes Associated with the Pathogenesis of Odontogenic Keratocyst. International Journal of Molecular Sciences. 2024; 25(4):2379. https://doi.org/10.3390/ijms25042379
Chicago/Turabian StyleRamírez-Martínez, Carla Monserrat, Itzel Legorreta-Villegas, Claudia Patricia Mejía-Velázquez, Javier Portilla-Robertson, Luis Alberto Gaitán-Cepeda, Jessica Tamara Paramo-Sánchez, Osmar Alejandro Chanes-Cuevas, Alejandro Alonso-Moctezuma, and Luis Fernando Jacinto-Alemán. 2024. "In Silico Analysis of Genes Associated with the Pathogenesis of Odontogenic Keratocyst" International Journal of Molecular Sciences 25, no. 4: 2379. https://doi.org/10.3390/ijms25042379
APA StyleRamírez-Martínez, C. M., Legorreta-Villegas, I., Mejía-Velázquez, C. P., Portilla-Robertson, J., Gaitán-Cepeda, L. A., Paramo-Sánchez, J. T., Chanes-Cuevas, O. A., Alonso-Moctezuma, A., & Jacinto-Alemán, L. F. (2024). In Silico Analysis of Genes Associated with the Pathogenesis of Odontogenic Keratocyst. International Journal of Molecular Sciences, 25(4), 2379. https://doi.org/10.3390/ijms25042379