1. Introduction
Embryo implantation is a tightly regulated process by which a competent blastocyst reaches the maternal endometrium at its most receptive phase (window of implantation). This process begins when the outer layer of the blastocyst, the trophectoderm cells, come into contact and attach to the endometrial epithelium (apposition-attachment) [
1,
2,
3]. Shortly thereafter, the trophectoderm undergoes differentiation, giving rise to two different regions: (i) syncytiotrophoblast, a multinucleated outer cell layer that penetrates the endometrial epithelial membrane, allowing the embryo’s implantation into the endometrium; and (ii) mononuclear cytotrophoblast cells that proliferate to form cell columns that further penetrate the endometrial stroma [
1,
3,
4]. Emerging from the tips of these anchoring villi structures are extravillous cytotrophoblast cells (EVT), known for their highly migratory, proliferative, and invasive characteristics [
5,
6,
7].
Trophoblast invasion of the uterus involves attachment of EVT to the extracellular matrix and degradation of the matrix as well as the uterine vasculature (endovascular invasion) to ultimately establish the uteroplacental circulation [
3,
5]. When this physiological equilibrium is disrupted, implantation failure and subsequent spontaneous abortion may occur [
3]. Moreover, shallow trophoblast invasion and impaired uterine vasculature remodeling during the early stages of gestation can also perpetuate during pregnancy and lead to pregnancy complications such as preeclampsia, intrauterine growth restriction (IUGR), and premature birth [
7,
8,
9]. Despite significant progress in reproductive research, efforts are still needed towards a better understanding of the physiological processes initiated during implantation, such as trophoblast invasion and migration, as well as the proteins that might orchestrate them. These two processes are closely related since both require changes in the cell microenvironment and concomitant activation of extracellular proteases [
10]. The proteolytic activity of extracellular matrix-degrading metalloproteinases (MMPs) has been largely implicated in the efficiency of trophoblast invasion, specifically in the disruption of the extracellular matrix at the fetal-maternal interface during embryo implantation [
11].
We have previously demonstrated, in a model of extravillous trophoblast of choriocarcinoma cell lines, that the forkhead box M1 (FOXM1) transcription factor is an important mediator of angiogenesis and migration, which are key processes involved in early placentation [
12]. Additionally, we have also showed in a rat model that placental FOXM1 protein expression decreased as gestational age progresses, further indicating a role for FOXM1 in the early stages of pregnancy [
12]. Although our previous results are compelling, the precise role of FOXM1 in implantation and whether FOXM1 is expressed in the pre-implantation blastocyst stage remain unknown. FOXM1 plays a crucial role in a plethora of biological processes, by directly regulating the transcription of downstream target genes involved in cell proliferation, cell cycle progression, angiogenesis, migration, invasion, cell differentiation, and DNA damage repair [
13,
14]. FOXM1 activity is also required in embryonic and fetal development [
15,
16]. Indeed, during mice embryogenesis, FOXM1 expression is found in the mesenchymal and epithelial cells of the liver, lung, intestine, renal cortex, and urinary tract; yet, its adult expression pattern is restricted to actively proliferating epithelial cells of the intestine, the spermatocytes and spermatids of the testis, the thymus, and colon [
17]. Moreover, Foxm1 knockout mice are characterized by an embryonic lethal phenotype due to severe abnormalities in the development of the heart and liver [
18]. In this study, we tested the hypothesis that FOXM1 is expressed in the trophectoderm of the blastocyst and participates in embryo implantation by contributing to trophoblast migration and early trophoblast invasion, by inducing transcription activation of genes involved in these processes.
3. Discussion
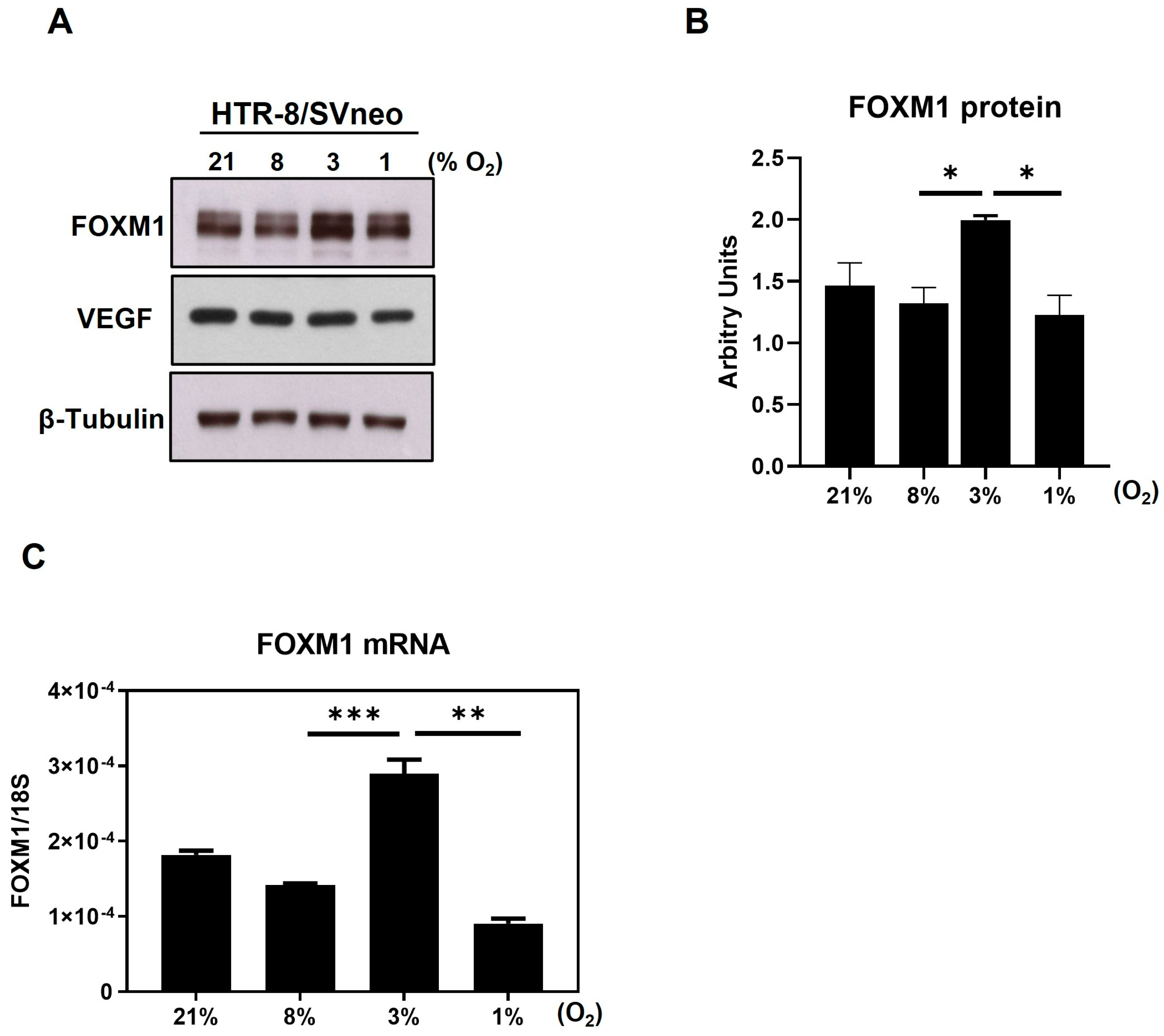
Implantation and early development of the placenta are highly regulated processes that are pivotal for a successful pregnancy. Indeed, there is large body of evidence showing that the impairment of these processes contributes to the etiopathology of pregnancy complications such as preeclampsia, intrauterine growth restriction, and preterm labor. An improved understanding of the molecular mechanisms involved in normal implantation and early trophoblast development will improve the ability of clinicians to understand and treat these pregnancy disorders. Using a model of choriocarcinoma cell lines that exhibits an extravillous trophoblast phenotype, we showed that FOXM1 is an important mediator of key processes involved in early placentation including angiogenesis and migration [
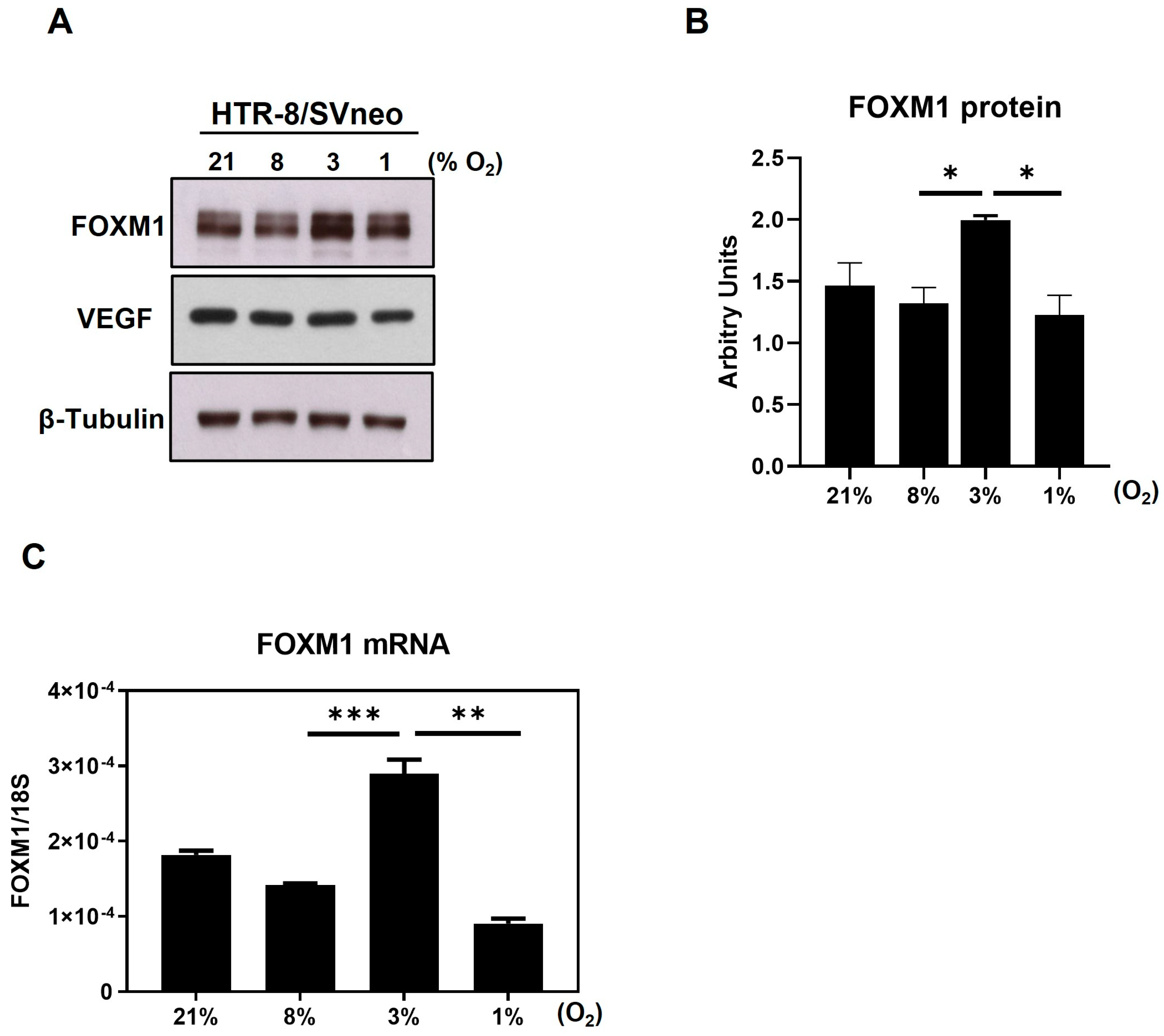
12]. Here, we further demonstrate that, in a model of first trimester trophoblast cells, FOXM1 expression significantly increases in response to 3% O
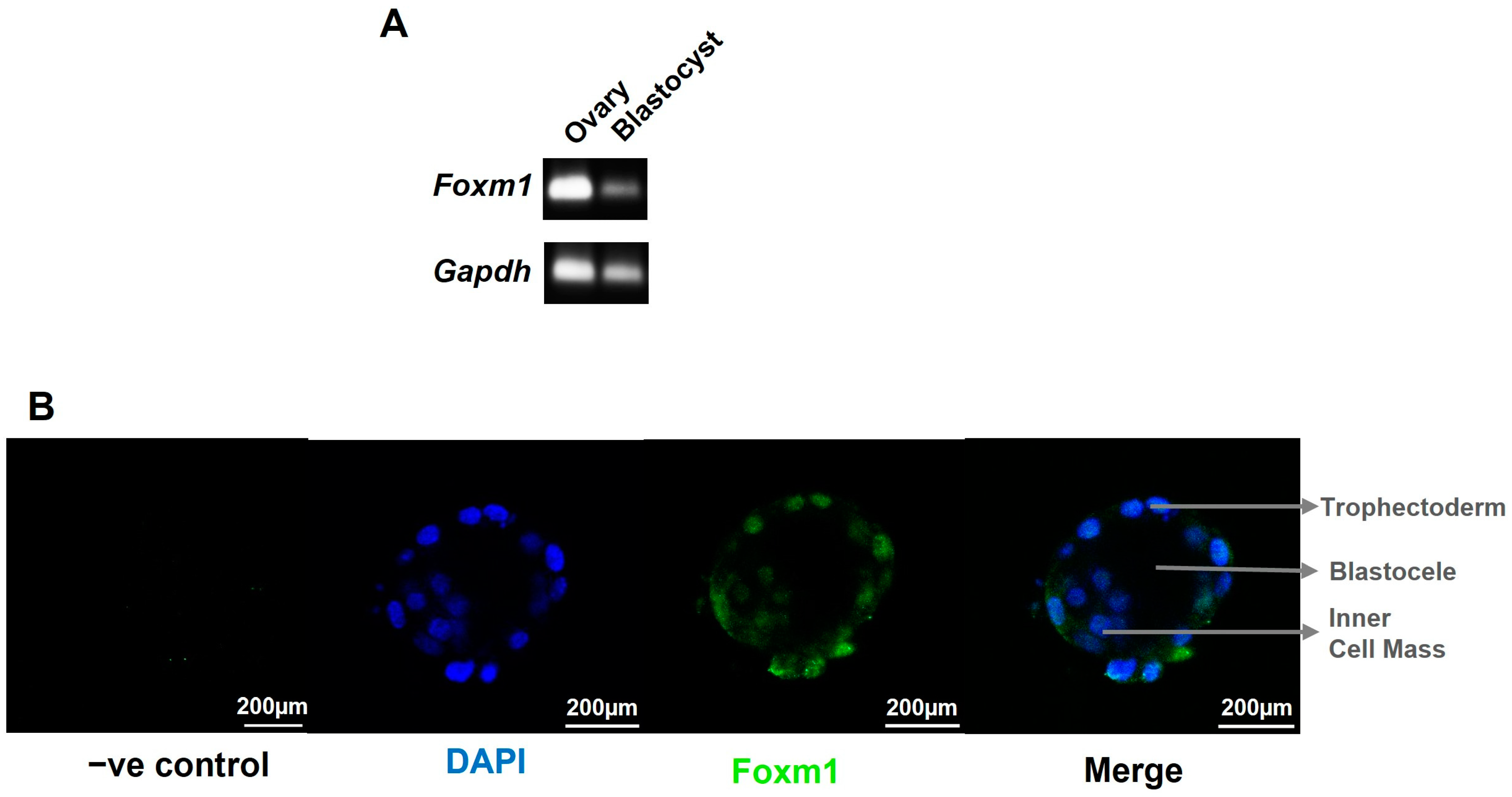
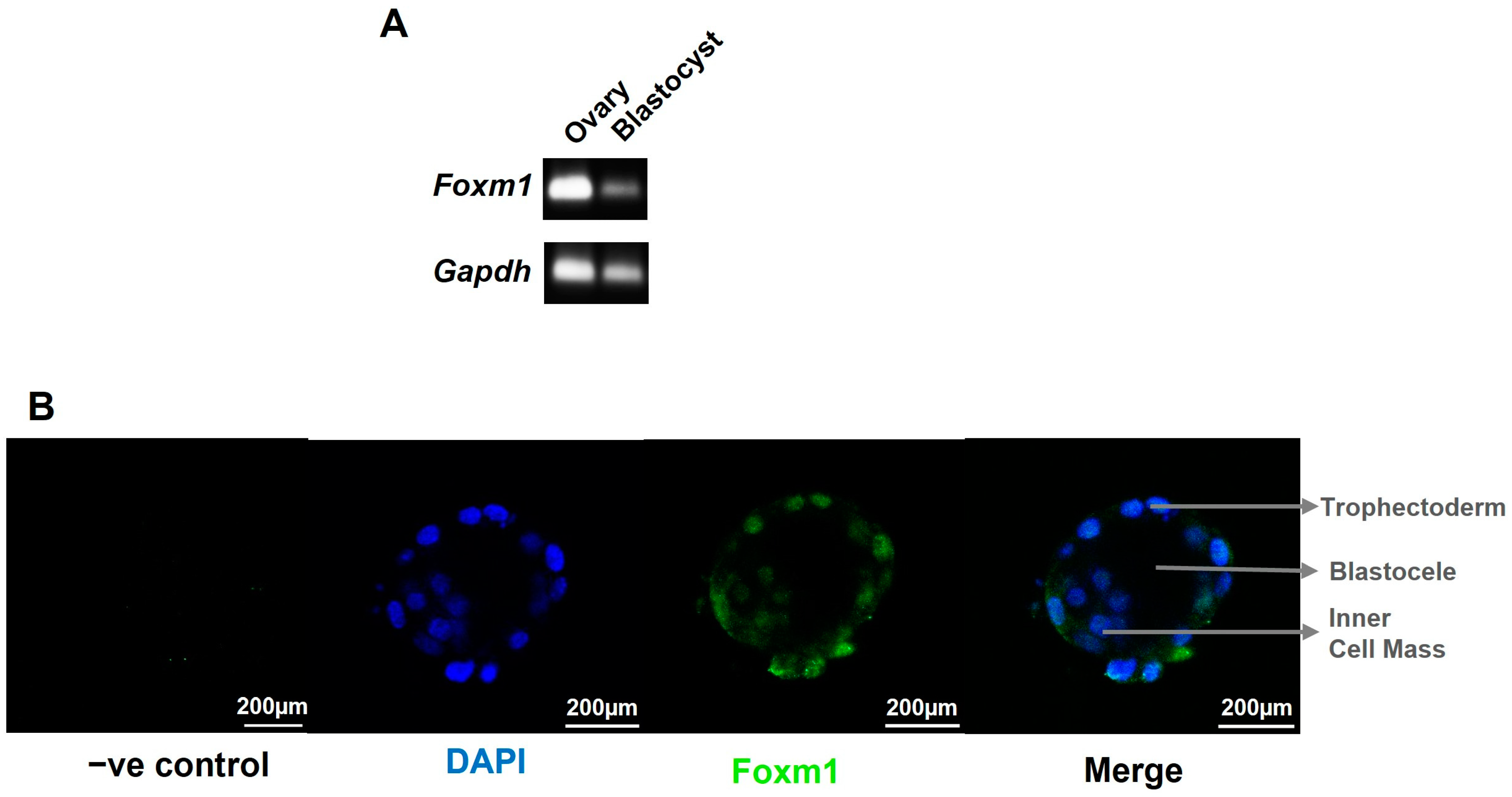
2, which coincides with the oxygen concentration within the uterine surface at implantation, and 1% oxygen, resembling a hypoxic environment, diminished FOXM1 expression [
19]. Moreover, we have observed, for the first time, that FOXM1 is expressed as early as the blastocyst stage in mice embryos and that it is not only is expressed in the inner cell mass but also in the trophectoderm, highlighting the potential role of this protein in the early phases of trophoblast differentiation and blastocyst implantation. This process begins when the trophectoderm cells of the blastocyst attach to the endometrial epithelium, start to migrate out of the blastocyst, and invade the endometrium [
1,
2,
3]. The detection of FOXM1 in the inner cell mass was not surprising since FOXM1 has been shown to be required for embryonic and fetal development [
15,
16]. Indeed, during embryogenesis, FOXM1 is expressed to ensure correct development of both epithelial and mesenchymal tissues [
17]. Additionally, FOXM1 has been demonstrated to be essential for embryonic development of mice pulmonary vasculature [
18].
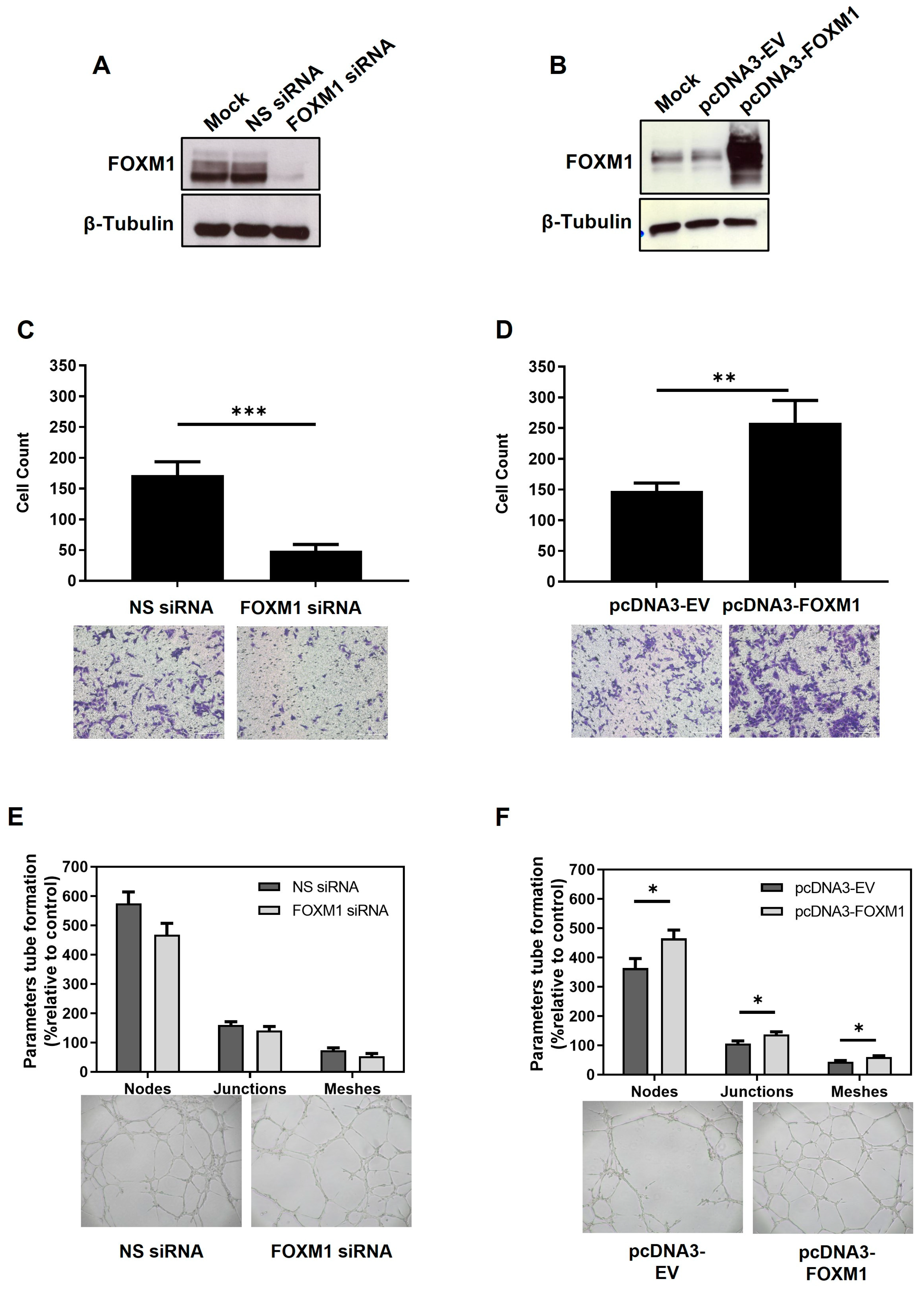
In the cancer scenario, it has been demonstrated that FOXM1 induces tumor proliferation, migration, invasion, and angiogenesis [
14], which are processes shared by trophoblast cells during the peri-implantation phase. Accordingly, we have also demonstrated that FOXM1 is necessary for the migration and angiogenesis of the human HTR8/Svneo first trimester trophoblast cell line while not influencing its proliferative ability (
Supplementary Figure S2). Moreover, neither silencing nor overexpression of FOXM1 led to statistically significant effects on the ability of HTR8/Svneo cells to invade MATRIGEL, as assessed by transwell invasion assays (
Supplementary Figure S1).
Successful implantation requires effective maternal-embryonic communication [
2,
3,
4,
27]. Using an in vitro 3D trophoblast invasion model, we have confirmed that trophoblast cells were only able to migrate from the trophosphere and invade through the MATRIGEL in the presence of MenSCs of endometrial origin, which was pre-treated with the endometrial mimic, suggesting that MenSCs secrete important factors that trigger the motility and invasion of the trophoblast cells, as described previously [
10,
23,
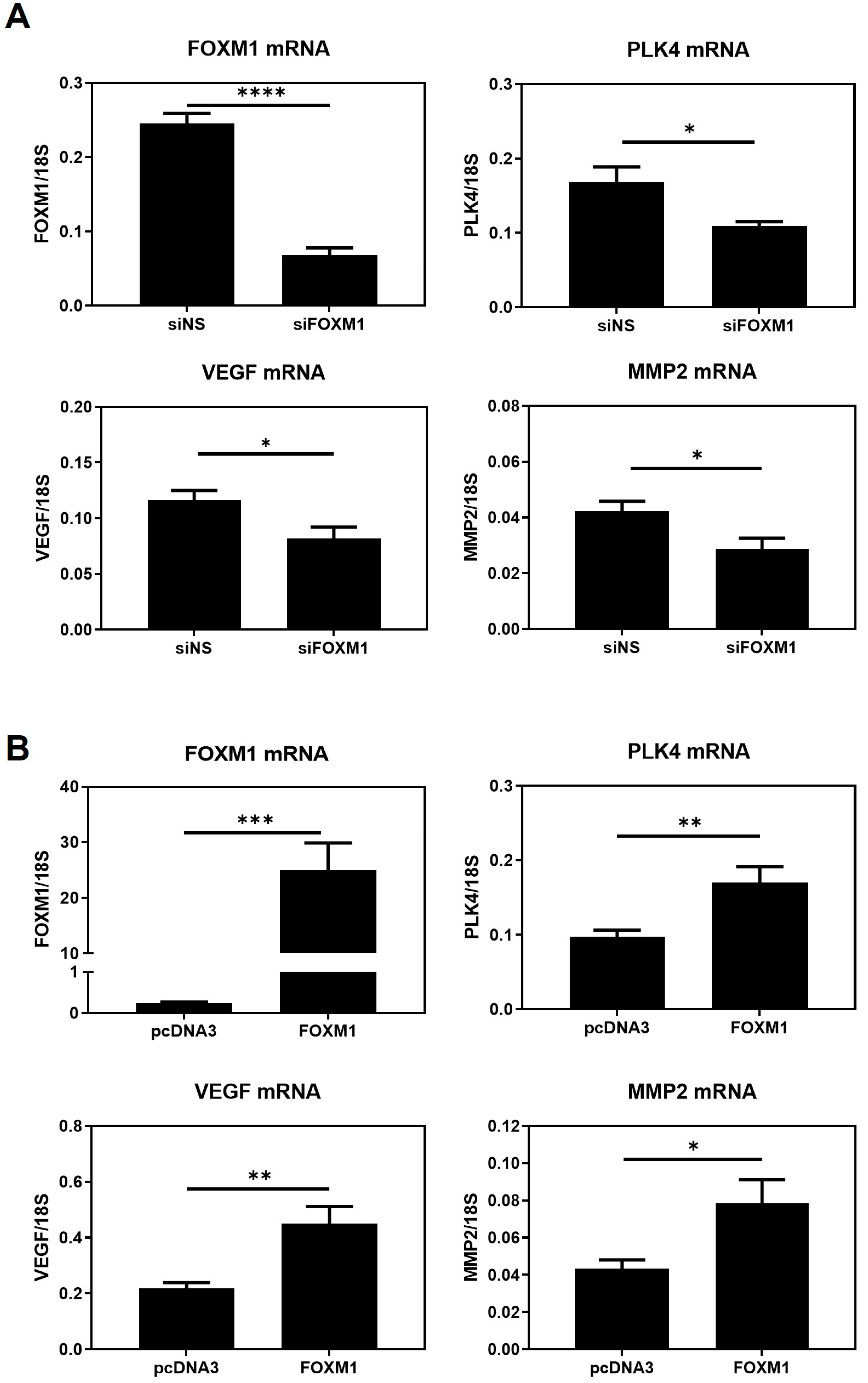
24]. More importantly, FOXM1 mRNA expression was significantly higher in those trophoblast cells that invaded greater into the MATRIGEL, i.e., the trophospheres that were co-cultured with MATRIGEL and MenSCs compared to pre-invasion trophospheres. Together, these results suggest that trophoblast invasion strongly relies on maternal-fetal communication and that maternal decidual cells may induce trophoblast cells to express higher levels of FOXM1 that subsequently activate genes involved in trophoblast migration and invasion. Mitochondria or extracellular vesicle transfer could be studied as potential communication mechanisms. To further validate that FOXM1 is indeed required for trophoblast invasion, we demonstrated that FOXM1-overexpressed trophospheres exhibited a significant increase in the area invaded by the trophosphere, compared to those cells transfected with the pcDNA3-empty vector control. It is worth mentioning that spheroid formation from HTR-8/Svneo morphologically resembles a blastocyst and expresses higher stemness markers, NANOG and SOX2, and lower CDH1 epithelial marker, in comparison with its adherent counterpart [
23].
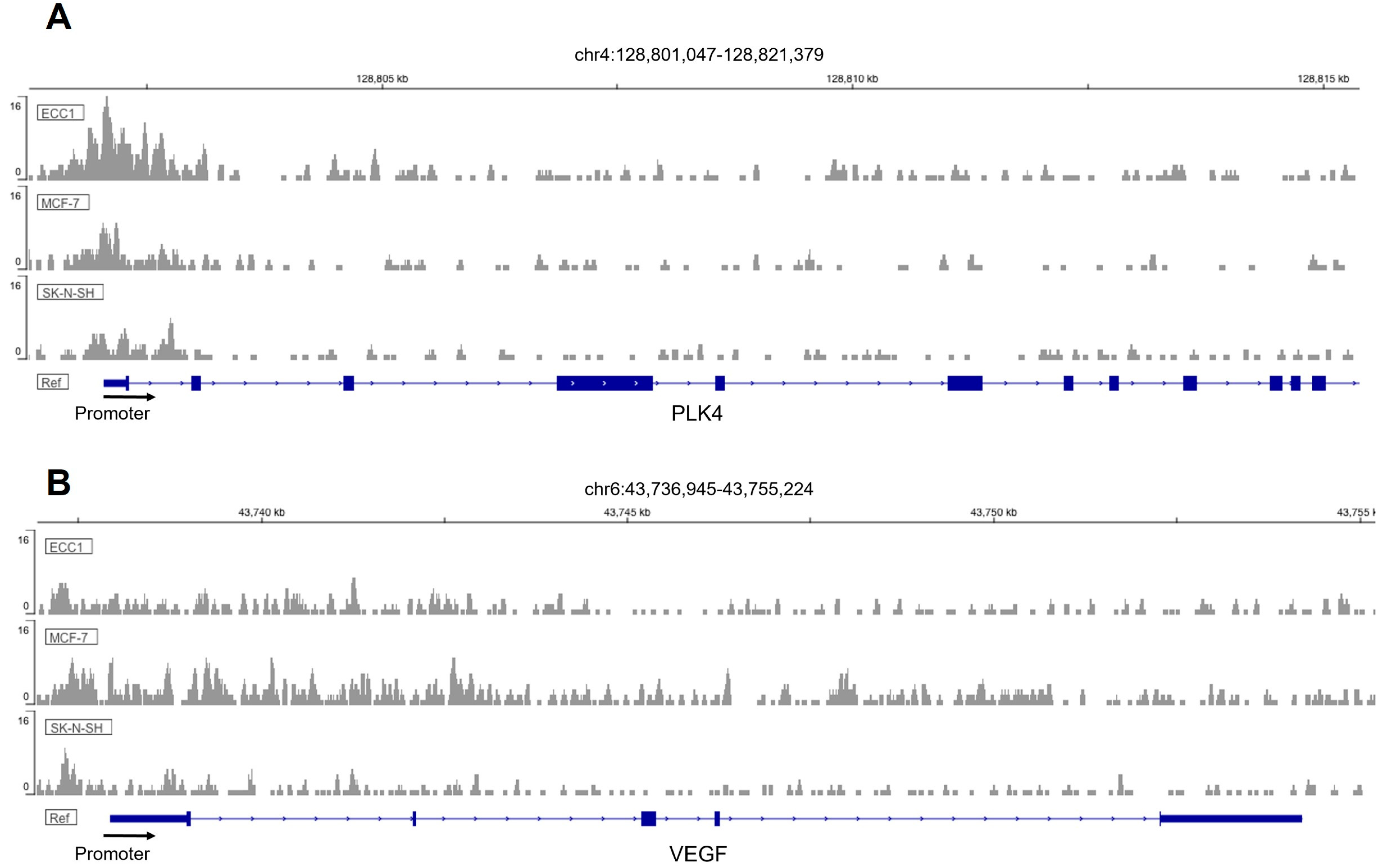
As a transcription factor, it is plausible that FOXM1 exerts its effects by directly binding the promoters and inducing the transcription of genes involved in trophoblast differentiation, migration, invasion, and angiogenesis. Thus, we propose that
PLK4,
VEGF, and
MMP2 are FOXM1 downstream targets involved in these processes. Invasion and migration are closely related mechanisms that involve the ability of the cells to move in response to a stimulus and to advance through the extracellular matrix within a tissue by activating extracellular proteases [
29]. The proteolytic activity of extracellular matrix-degrading metalloproteinases (MMPs) has been largely implicated in the efficiency of trophoblast invasion, specifically in disrupting the extracellular matrix at the fetal-maternal interface during embryo implantation [
11]. Specifically, MMP2 has been identified to be secreted by the cytotrophoblast cells that are responsible in digesting the major constituents of the endometrial matrix [
27]. Indeed, MMP2 can be directly regulated by FOXM1 in human retinoblastoma Y-79 cells [
28]. Consistently, a large-scale microarray study of the transcriptome of the rat placenta throughout mid-late gestation also reveals the concomitant expression of FOXM1 and angiogenic genes, including VEGFA, MMP14, Caveolin-1, and Angiopoietin-4 [
30]. Moreover, VEGF has also been demonstrated to be a direct target of FOXM1 in breast cancer cells [
22]. Polo-like kinase four (PLK4) is a member of the serine/threonine kinase family that plays an important role in cell cycle regulation, by participating in centriole duplication and maintaining mitotic accuracy in normal cells. Its deregulation, henceforth, has been associated with a prominent role in cancer and metastasis [
31,
32]. PLK4 has also been demonstrated to be involved in the differentiation of trophoblast stem cells into trophoblast giant cells during placental development [
31,
33]. This evidence, together with the ChIP-Seq data, indicates that
VEFG,
PLK4, and
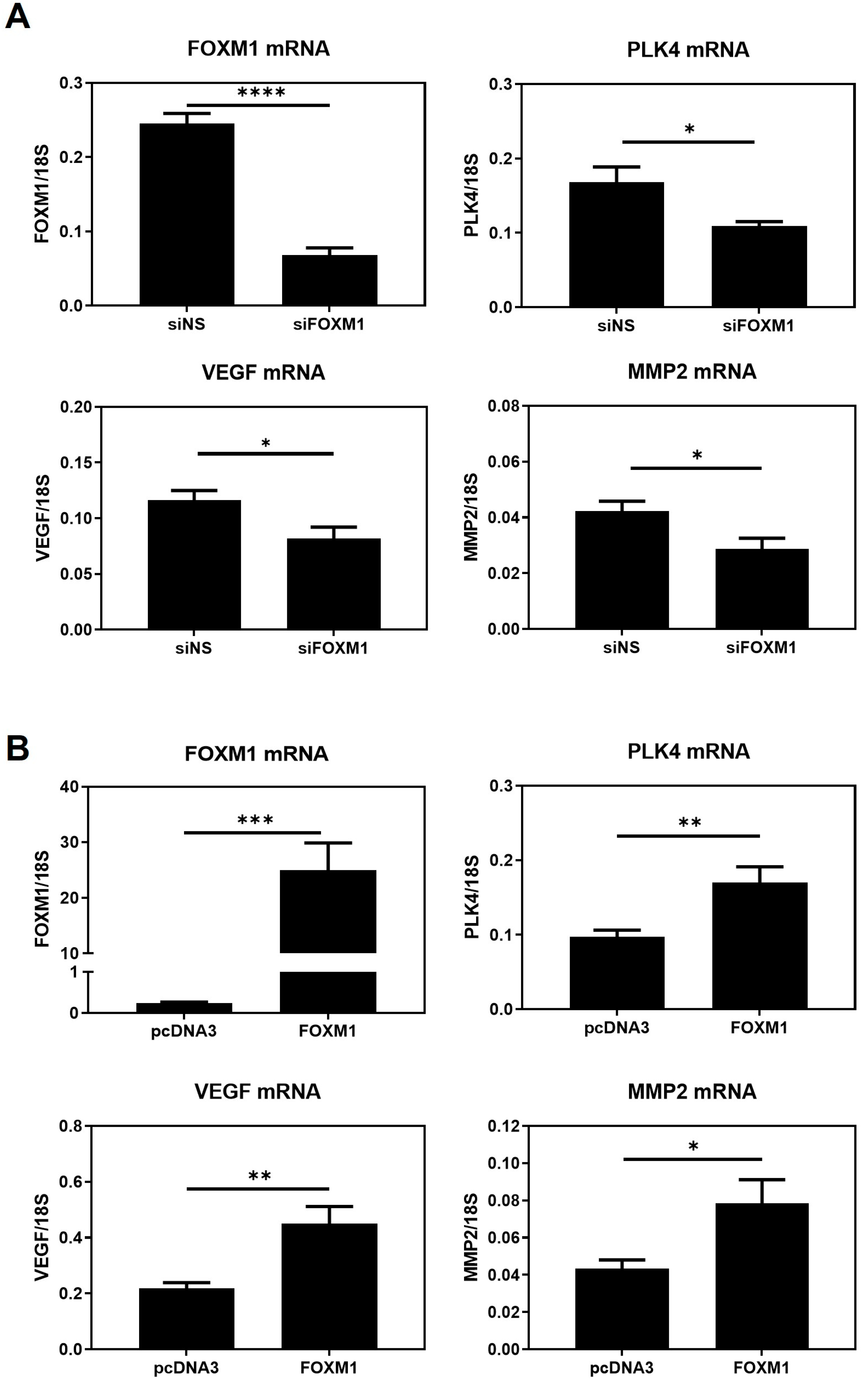
MMP2 are potential downstream targets of FOXM1, through which it promotes trophoblast differentiation, migration, invasion, and angiogenesis during early placentation. To support this conjecture, we have also shown that the mRNA expression of these genes is significantly downregulated following silencing of FOXM1 and significantly upregulated following FOXM1 overexpression in HTR-8/Svneo cells. Further studies using ChIP are required to validate that
VEFG,
PLK4, and
MMP2 are downstream targets of FOXM1 in HTR-8/Svneo cells.
Our results demonstrate, for the first time, that FOXM1 is present in the trophectoderm of mice blastocysts. By using a 3D in vitro model of implantation, we have confirmed that maternal-fetal communication is crucial for trophoblast invasion and that maternal stromal cells may induce trophoblast cells to express higher levels of FOXM1, which subsequently activates the transcription of its downstream targets genes that are important for successful implantation: PLK4, VEGF, and MMP2.
4. Materials and Methods
4.1. Cell Culture
The human HTR8/Svneo first trimester trophoblast cell line was purchased from the American Type Culture Collection (CRL-3271; Lot #70016636, ATCC, Manassas, VA, USA) and cultured in RPMI-1640 medium (GE Healthcare, Piscataway, NJ, USA), supplemented with 10% heat-inactivated fetal bovine serum (FBS), 1 mmol/L sodium pyruvate, and 1% penicillin/streptomycin (P/S) (all Gibco, ThermoFisher, Waltham, MA, USA). Cells were maintained at 37 °C in a humidified incubator with 5% CO2. For hypoxia experiments, cells were seeded in the appropriate cell culture plastic wells and maintained at 21%, 8%, 3%, or 1% O2 for 24 h to 72 h prior to experiments. Hypoxic chamber C-474 equipped with Pro-Ox 110 oxygen controlling regulator (BioSpherix, Parish, NY, USA) was used.
4.2. Embryo Collection
C57BL/6 mice were purchased from Universidad de Chile. Animals were maintained in the animal facility of Universidad de los Andes at 25 °C and 12 h light:dark cycling and fed standard chow and water ad libitum. Protocols were conducted in agreement with the National Research Council (NRC) publication Guide for the Care and Use of Laboratory Animals (eighth edition, The National Academies Press, Washington, DC, USA). The animal study was reviewed and approved by the Ethical Scientific Committees of Universidad de los Andes, Santiago, Chile. For blastocyst collection, female mice were superovulated by intraperitoneal injections with 5 IU of pregnant mares’ serum gonadotropin (PMSG) (Novormon, Buenos Aires, Argentina) and 5 IU of human chorionic gonadotropin (HCG) (Sigma-Aldrich, St Louis, MO, USA) 48 h later. Immediately after HCG injection, female mice were mated 1:2 to male mice of the same strain. Mating was evaluated by inspection of the vaginal plug on the following day. The presence of sperm plug was considered as embryonic day 0.5 (E0.5) of pregnancy. Pregnant females were euthanized at E3.5 and subsequently, blastocysts were obtained. The procedure involved initial anesthesia with ketamine:xylazine, followed by euthanasia by cervical dislocation. Embryos were collected by flushing the uterus with M2 medium (Sigma-Aldrich, St Louis, MO, USA). Then, freshly harvested embryos were washed once in M2, followed by three washes with M2 medium containing polyvinyl pyrrolidone (PVP; 6 mg/mL, H6PVP Sigma-Aldrich, USA) and three washes with 1X phosphate-buffered saline (PBS) (Cytiva, Logan, UT, USA). Embryos and oocytes with signs of disintegration were classified as degenerated embryos or oocytes. At least 14 embryos at the blastocyst stage were used for qRT-PCR. The blastocysts were transferred to a tube with a minimal volume (1-2 µL) of PBS, snap-frozen in liquid nitrogen, and stored at −80 °C. For blastocyst differential labelling of cell lineages by immunofluorescence, embryos were washed and used immediately.
4.3. Immunofluorescent Staining
Immunofluorescence staining of the blastocyst was carried out as described previously [
34]. After embryo collection, blastocysts (E3.5) were washed three times in M2 medium, and the
zona pellucida was removed using acidified Tyrodes solution (Sigma-Aldrich, USA), at pH 2.3 at 37 °C for 30–60 s, followed by extensive washing in M2. Blastocysts were fixed with 4% PFA (ThermoFisher, Rockford, IL, USA) in PBS for 15 min at room temperature. The fixative was removed, and the embryos were washed three times with 1X PBS and then permeabilized with 0.2% Triton X-100 (Sigma-Aldrich, USA) in 1X PBS for 10 min at room temperature. Embryos were washed again three times with PBS containing 0.1% (
v/
v) Tween 20 (Winkler, Santiago, Chile) (PBST), followed by blocking with 1% BSA (Rockland, Philadelphia, PA, USA) and 5% goat serum G9023 (Sigma-Aldrich, St. Louis, MO, USA) in PBST for 1 h at room temperature. FOXM1 G-5 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) primary antibody in PBST, 1% BSA, 1% goat serum, and glycine at 0.05 M (Winkler, Chile) were added overnight at 4 °C. The next day, the embryos were washed three times with PBST with 1% BSA, followed by secondary antibody Alexa Fluor 488 anti-mouse (ThermoFisher, Eugene, OR, USA) incubation for 1 h at room temperature in a dark environment. Final PBST with BSA washes were performed. A drop of Fluoromount-G (Invitrogen, Carlsbad CA, USA) mounting medium with 4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA, USA) was added to each drop of blastocysts before addition of a coverslip and sealing with clear nail varnish. After drying, slides were stored at 4 °C. Images were collected with a Leica TCS SP8 confocal microscope (Leica Microsystems, Mannhein, Germany) with 20× objective (N.A. 0.5) using LAS-X software v3.0 (Leica Microsystems, Mannhein, Germany).
4.4. Western Blot Analysis
Cells were harvested by trypsinization and whole cell lysates prepared as described previously [
35]. Primary antibodies, FOXM1 D12D5 (Cell Signaling, Danvers, MA, USA), β-Tubulin H-235 (Santa Cruz Biotechnology, USA), and VEFG-A ab46154 (Abcam, Cambridge, MA, USA), were detected using horseradish peroxidase-linked anti-rabbit or anti-mouse conjugates as appropriate (KPL, Gaithersburg, MD, USA), and proteins were visualized using enhanced chemiluminescence (ECL) Pierce ECL Western Blotting Substrate detection system (ThermoFisher, USA) with X-ray films. The obtained images were analyzed using ImageJ software v1.54h (National Institutes of Health, Bethesda, MD, USA). All samples were normalized for protein loading using β-tubulin.
4.5. RNA Isolation and Quantitative Real-Time PCR
Total RNA was extracted from HTR8/SVneo cells with the TRIzol Reagent (Ambion, Carlsbad CA, USA) according to the manufacturer’s protocols. RNA from the pooled blastocyst (n = 14) was isolated using PureLink RNA Micro Kit (Invitrogen, Carlsbad CA, USA) according to the manufacturer’s protocols. RNA concentration and quality were evaluated using the NanoDrop One spectrophotometer (ThermoFisher, Madison WI, USA). Before cDNA synthesis, total RNA was treated with DNase I (Invitrogen, Carlsbad CA, USA). Complementary DNA generated by SuperScript II Reverse Transcription System (Invitrogen, Carlsbad CA, USA), according to the manufacturer’s instructions, was analyzed using quantitative real-time PCR (qRT-PCR) in the Stratagene Mx3000P system (Agilent Technologies, Santa Clara, CA, USA), using Brilliant III SYBR Green qPCR Master Mix (Agilent Technologies, USA). The qRT-PCR was set to 95 °C for 10 min for enzyme activation, followed by 35 cycles of denaturation and primer annealing/extension consisting of 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 15 s, respectively. After the PCR runs, a dissociation curve was generated to confirm the absence of nonspecific amplification. Transcription levels were quantified using the 2−ΔΔCT method. 18S ribosomal RNA or GAPDH housekeeping genes were used to normalize input complementary DNA. The following gene-specific primers were used for mouse: Foxm1-sense: 5′-GGACATCTACACTTGGATTGAGG-3′ and Foxm1-antisense: 5′-TGTCATGGAGAGAAAGGTTGTG-3′; Gapdh-sense: 5′-AGTGGCAAAGTGGAGATT-3′ and Gapdh-antisense 5′-GTGGAGTCATACTGGAACA-3′; and for human HTR8/SVneo cells: FOXM1-sense: 5′-TGCAGCTAGGGATGTGAATCTTC-3′ and FOXM1-antisense: 5′-GGAGCCCAGTCCATCAGAACT-3′; VEGF-sense: 5′-TATGCGGATCAAACCTCACC-3′ and VEGF-antisense: 5′-CTTGTCTTGCTCTATCTTTCTTTGG-3′; MMP2-sense: 5′-TGTGACGCCACGTGACAAG-3′ and MMP2-antisense: 5′-CCAGTATTCATTCCCTGCAAAGA-3′; PLK-4-sense: 5′-GATAGACCACCCTCACCTACT-3′ and PLK-4-antisense: 5′-CTGTACAAACCTGGAAGCATATTG-3′; 18S-sense: 5′-GCCGCTAGAGGTGAAATTCTTGGA-3′ and 18S-antisense: 5′-ATCGCCAGTCGGCATCGTTTAT-3′; GAPDH-sense: GTCAGGGTCTCTCTCTTCCT and GAPDH-antisense: GCTCTCCTCTGACTTGAACA.
4.6. Transient Gene Silencing and Overexpression
For gene silencing, HTR8/SVneo cells were transiently transfected with ON-TARGETplus SMARTpool siRNA (Dharmacon ThermoFisher, Lafayette, CO, USA), using Oligofectamine (Invitrogen, Carlsbad, CA, USA) and Opti-MEM (Gibco, ThermoFisher, Waltham, MA, USA) according to the manufacturer’s instructions. siRNA FOXM1 (L-009762-00-0005) and the non-specific (NS) control siRNA (D-001810-10-05) SMARTpools were used. RNA interference experiments were carried out at 3% O
2. For gene overexpression, HTR8/SVneo cells were transfected with either the pcDNA3-empty vector or pcDNA3-FOXM1 [
36] using FuGENE 6 Transfection Reagent (Promega, Madison, WI, USA) in a 3:1 ratio (μL of FuGENE: μg of DNA) according to the manufacturer’s recommendations. The overexpression experiments were performed at 1% O
2.
4.7. Transmigration Assay
HTR8/SVneo cells transiently transfected with either FOXM1 siRNA or control NS siRNA, or pcDNA3-FOXM1 or empty vector EV-pcDNA3, under different oxygen tensions were used to examine cell migration in vitro. Following 24 h of transfection, 50,000 HTR-8/SVneo cells were seeded in a millicell insert (pore 8 µm, 12 mm, Millipore, Billerica, MA, USA) with 400 µL RPMI with reduced serum (0.5% FBS). Inserts were placed on 500 µL of RPMI supplemented with 10% FBS in 48-well plates. Transmigration capacity was evaluated following 12 h incubation either at 3 or 1% O2. Briefly, the insert was washed with 1X PBS, fixed with cold methanol for 2 min, and stained with 0.5% crystal violet (Winkler, Santiago, Chile). Cells inside the inserts were scraped with cotton swabs moistened with 1X PBS to ensure that only migrated cells were analyzed. Five fields were captured for each insert at 4× objective magnification (N.A. = 0.10, W.D. = 10.5 mm) before and after scraping under an inverted microscope (Primo Vert, Zeiss, Jena, Germany), using the AxioCam ERc5s camera (Zeiss, Jena, Germany). Images were analyzed with AxioVision Rel analysis software v4.8 (Zeiss, Jena, Germany). The percentage of migrated cells was calculated as follows: number of cells after/before scraping × 100 (average of the 5 fields). The experiments were performed in duplicate.
4.8. MATRIGEL-Based Tube Forming Assay
HTR8/SVneo cells transiently transfected with either FOXM1 siRNA or control NS siRNA, or pcDNA3-FOXM1 or empty vector EV-pcDNA3, under different oxygen tensions were resuspended in the RPMI medium with 2% FBS or in the endothelial growth media-2 (EGM-2, Bullet Kit, Lonza, Verviers, Belgium) (positive control) and seeded in triplicate in 96-well plates pre-coated with 70 μL growth factor reduced phenol-red free MATRIGEL matrix (Corning Life Sciences, Union City, CA, USA). Cells were incubated at 37 °C and under 3 or 1% O2 (accordingly) for 6 h and tube-like structures were examined with a phase-contrast microscope Primo Vert (Zeiss, Jena, Germany). One image per well was captured using an AxioCam ERc5s camera (Zeiss, Jena, Germany). Number of nodes, junctions, and meshes were analyzed using ImageJ-Angiogenesis Analyzer-HUVEC Phase Contrast software v1.0.c.
4.9. Trophosphere Formation
Trophoblast spheres (Trophospheres) are blastocyst like-structures formed in vitro using adherent HTR8/SVneo cells, as described previously [
23]. In brief, 20,000 HTR8/SVneo cells were suspended in 200 μL of supplemented RPMI media and placed into each well of an ultra-low attachment 96-well plate (Costar, Kennebunk, ME, USA). Following centrifugation at 300×
g for 5 min, cells were incubated for 72 h at 37 °C in a humidified atmosphere hypoxia chamber with 5% O
2 and 5% CO
2. The trophospheres were washed with 1X PBS and used for 3D invasion assays. The differential cellular characteristics of trophoblast cells as a 3D model or monolayer were previously described in detail [
23].
4.10. 3D Trophoblast Invasion Assay
To mimic the structure of the endometrium, mesenchymal stem cells isolated from menstrual fluid of healthy donors, who had not used hormonal contraceptives for at least three months, were isolated and cultured as described in [
23,
24]. All experiments were performed using MenSCs at early passages (P) P3 to P7 (
n = 4). Menstrual fluid was self-collected by consenting donors following their written informed consent to participate in this study according to a protocol reviewed and approved by the Ethical Scientific Committee of the Universidad de los Andes, Santiago, Chile. To achieve the 3D invasion assay [
23], growth factor reduced phenol-red free MATRIGEL (Corning Life Sciences, Union City, CA, USA) was mixed with Dulbecco’s Modified Eagle Medium (DMEM) without phenol red (Mediatech Inc., Manassas, VA, USA) containing 10% charcoal-stripped FBS and 1% P/S in a 1:1 ratio and added onto 3000 MenSCs that had been previously seeded in a 96-well plate and subjected to 213 pg/mL 17β-estradiol (E2) for 24 h, followed by 146 pg/mL E2 and 11 ng/mL progesterone (P4) for an additional 24 h at 5% O
2 (endometrial mimic), in order to resemble the physiological hormonal and O
2 conditions within the endometrium, throughout the menstrual cycle [
23]. The plate was incubated for 30 min at 37 °C to allow the MATRIGEL to solidify. A single trophosphere was placed on each well containing MenSCs and MATRIGEL, and 150 μL of endometrial mimic media were added to embed the sphere. Plates were incubated at 37 °C in a humidified atmosphere hypoxia chamber with 5% O
2 and 5% CO
2. Trophosphere invasion was evaluated after 72 h. Phase contrast images were captured by the contrast microscope Olympus CKX41 and Axiocam 208 camara (10× objective lens, N.A. = 0.25, W.D. = 10.5 mm) and AxioVision Rel software v4.8 (Zeiss, Jena, Germany). The invasion level (area) was quantified by using ImageJ software v1.51j8, and the trophospheres were collected for qRT-PCR assay. For mRNA analysis of the trophospheres following 3D invasion assay, the trophospheres were recovered from MATRIGEL and incubated with 300 μL of Cell Recovery Solution (Corning Life Sciences, Bedford, MA, USA) for 15 min on ice, followed by centrifugation at 400×
g for 5 min at 4 °C. The supernatant was removed and the trophospheres were washed with PBS, followed with centrifugation at 350×
g for 2 min at 4 °C. PBS was removed and the trophospheres were lysed with TRIzol for RNA extraction.
4.11. Statistical Analyses
Statistical comparisons between two means were performed using Student’s t-tests. Results were deemed statistically significant when p ≤ 0.05. Data normality was tested using the D’Agostino-Pearson test. For analysis of 2 groups, unpaired t-test was used. Graphing and statistical analyses were performed using GraphPad Prism 9.0.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}