
Altered Levels of Natural Autoantibodies against Heat Shock Proteins in Pregnant Women with Hashimoto’s Thyroiditis
 , , , ,
, , , ,  , and
, and 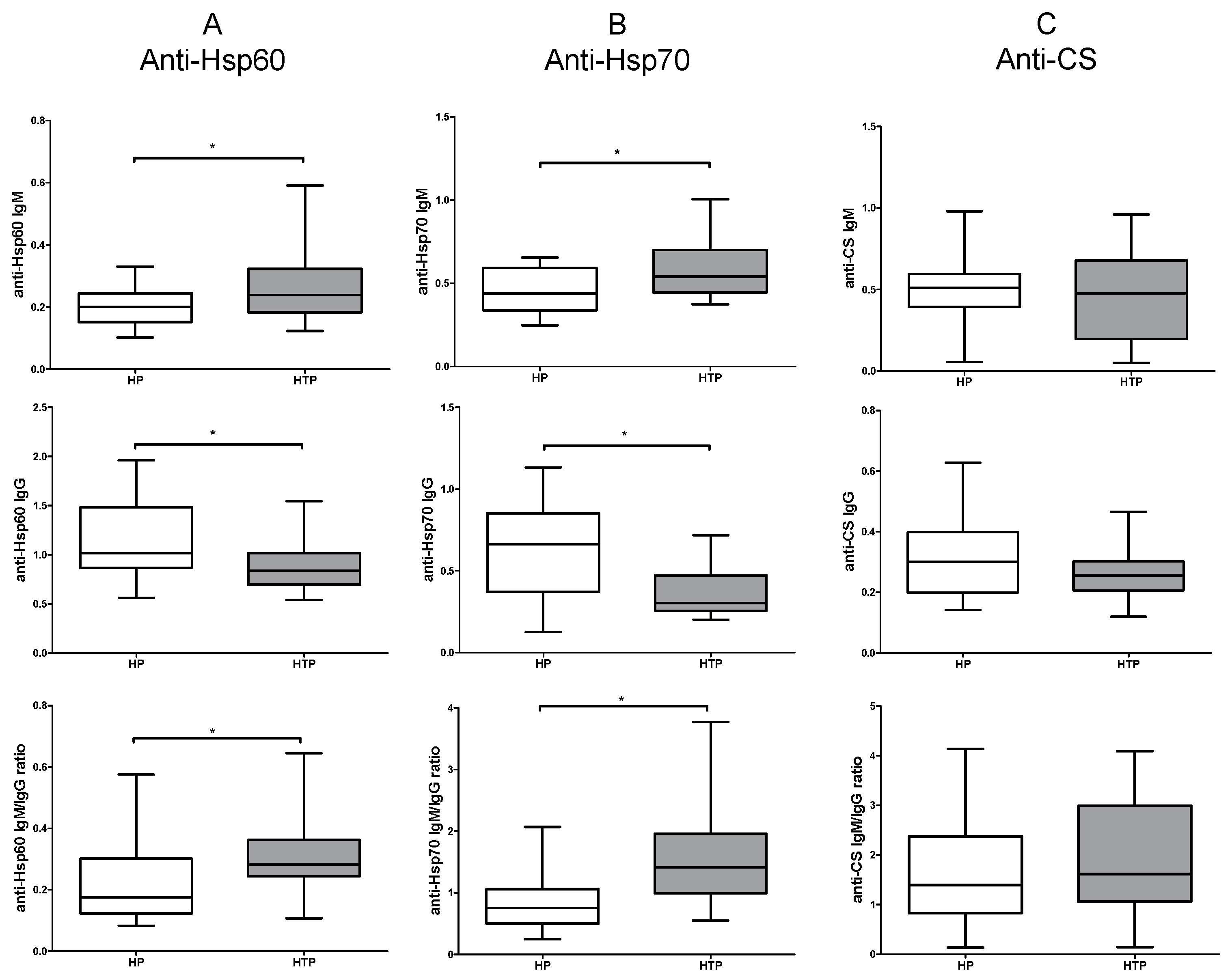{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
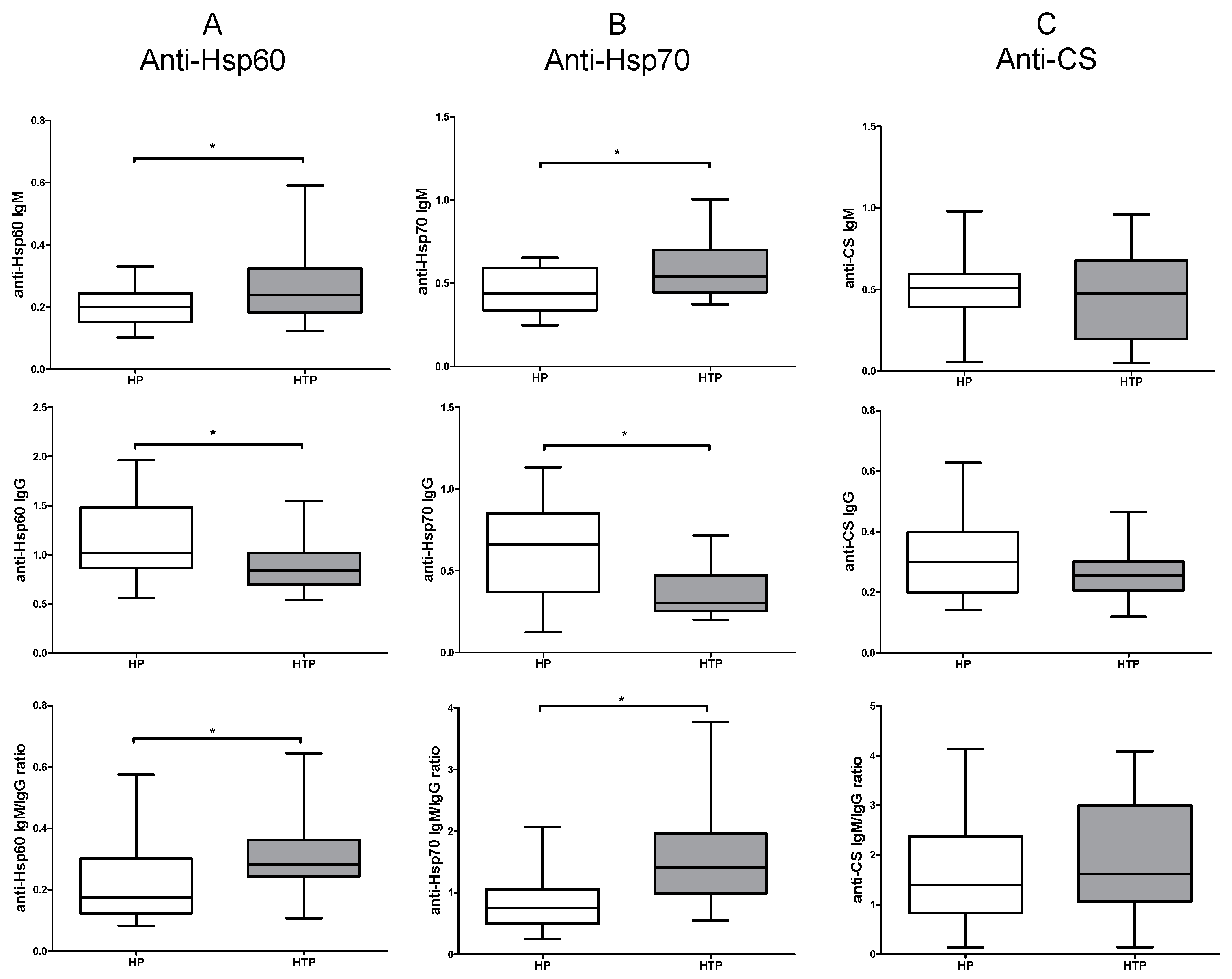
2.1. Increased IgM and Decreased IgG Levels of Anti-Hsp60 and Hsp70 Autoantibodies in HTP
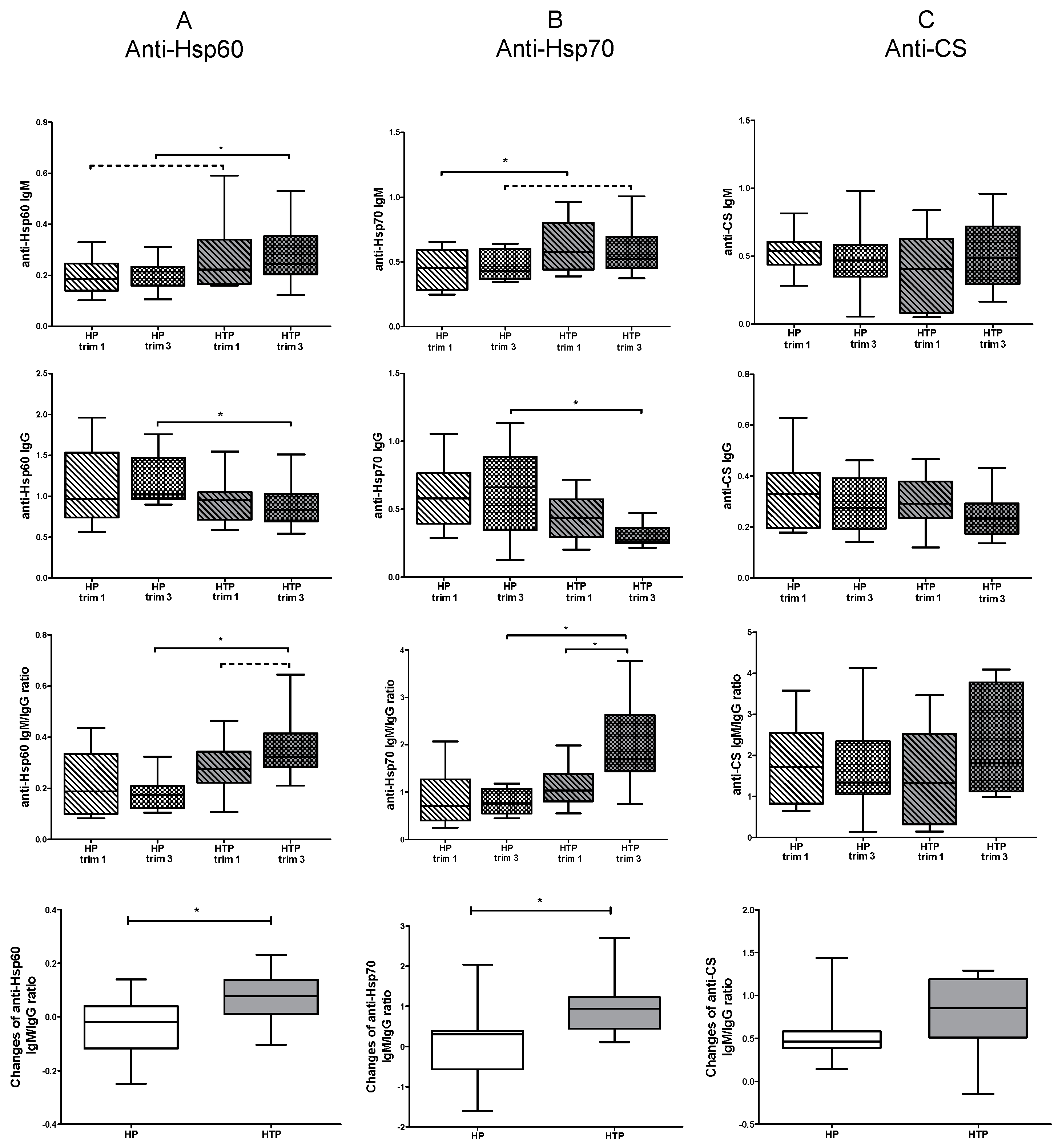
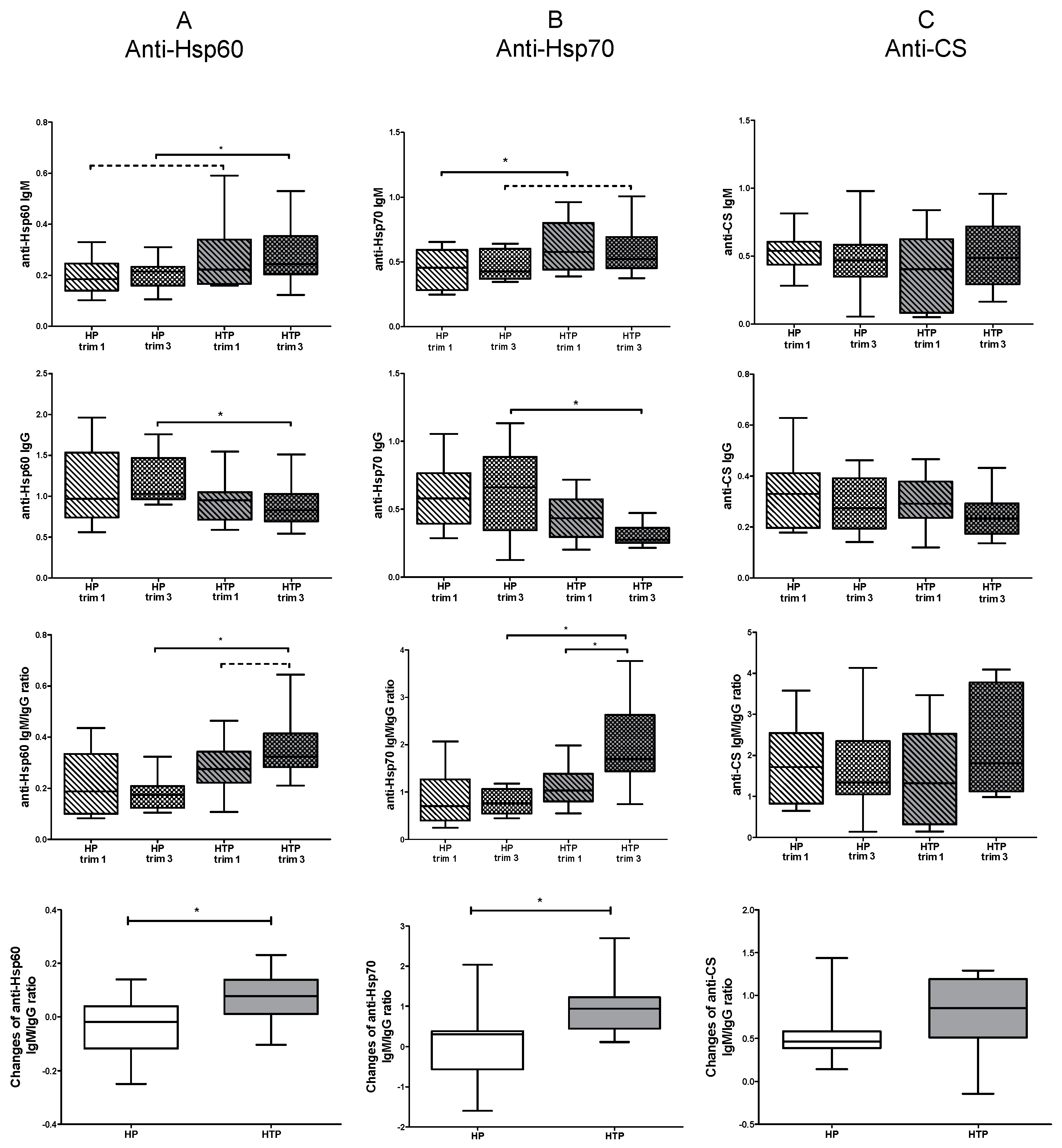
2.2. Altered Levels of Both IgM and IgG nAAbs against Hsp60 and Hsp70 in the Third Trimester of HTP Women Resulting in Their Elevated Ratios
3. Discussion
4. Materials and Methods
4.1. Selected Individuals
4.2. Measurement of nAAbs against CS
4.3. Measurement of nAAbs against Hsp60 and Hsp70
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biggs, A.T.; Littlejohn, L.F. Vaccination and Natural Immunity: Advantages and Risks as a Matter of Public Health Policy. Lancet Reg. Health Am. 2022, 8, 100242. [Google Scholar] [CrossRef] [PubMed]
- Franchi, M.; Pellegrini, G.; Cereda, D.; Bortolan, F.; Leoni, O.; Pavesi, G.; Galli, M.; Valenti, G.; Corrao, G. Natural and Vaccine-Induced Immunity Are Equivalent for the Protection against SARS-CoV-2 Infection. J. Infect. Public Health 2023, 16, 1137. [Google Scholar] [CrossRef] [PubMed]
- Maddur, M.S.; Lacroix-Desmazes, S.; Dimitrov, J.D.; Kazatchkine, M.D.; Bayry, J.; Kaveri, S.V. Natural Antibodies: From First-Line Defense Against Pathogens to Perpetual Immune Homeostasis. Clin. Rev. Allergy Immunol. 2020, 58, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.; Tokarz-Deptuła, B.; Deptuła, J.; Deptuła, W. Natural Antibodies—Facts Known and Unknown. Cent. Eur. J. Immunol. 2018, 43, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Amital, H.; Shoenfeld, Y. Natural autoantibodies, heralding, protecting and inducing autoimmunity. In Autoantibodies, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 7–12. [Google Scholar] [CrossRef]
- Baumgarth, N.; Tung, J.W.; Herzenberg, L.A. Inherent Specificities in Natural Antibodies: A Key to Immune Defense against Pathogen Invasion. Springer Semin. Immunopathol. 2005, 26, 347–362. [Google Scholar] [CrossRef]
- Lutz, H.U.; Binder, C.J.; Kaveri, S. Naturally Occurring Auto-Antibodies in Homeostasis and Disease. Trends Immunol. 2009, 30, 43–51. [Google Scholar] [CrossRef]
- Cohen, I.R. Autoantibody Repertoires, Natural Biomarkers, and System Controllers. Trends Immunol. 2013, 34, 620–625. [Google Scholar] [CrossRef]
- Avrameas, S.; Alexopoulos, H.; Moutsopoulos, H.M. Natural Autoantibodies: An Undersugn Hero of the Immune System and Autoimmune Disorders-A Point of View. Front. Immunol. 2018, 9, 1320. [Google Scholar] [CrossRef]
- Palm, A.K.E.; Kleinau, S. Marginal Zone B Cells: From Housekeeping Function to Autoimmunity? J. Autoimmun. 2021, 119, 102627. [Google Scholar] [CrossRef]
- Panda, S.; Zhang, J.; Tan, N.S.; Ho, B.; Ding, J.L. Natural IgG Antibodies Provide Innate Protection against Ficolin-Opsonized Bacteria. EMBO J. 2013, 32, 2905–2919. [Google Scholar] [CrossRef]
- Muramatsu, M.; Sankaranand, V.S.; Anant, S.; Sugai, M.; Kinoshita, K.; Davidson, N.O.; Honjo, T. Specific Expression of Activation-Induced Cytidine Deaminase (AID), a Novel Member of the RNA-Editing Deaminase Family in Germinal Center B Cells. J. Biol. Chem. 1999, 274, 18470–18476. [Google Scholar] [CrossRef] [PubMed]
- Irrgang, P.; Gerling, J.; Kocher, K.; Lapuente, D.; Steininger, P.; Habenicht, K.; Wytopil, M.; Beileke, S.; Schäfer, S.; Zhong, J.; et al. Class Switch toward Noninflammatory, Spike-Specific IgG4 Antibodies after Repeated SARS-CoV-2 mRNA Vaccination. Sci. Immunol. 2023, 8, eade2798. [Google Scholar] [CrossRef] [PubMed]
- Avrameas, S.; Ternynck, T. The Natural Autoantibodies System: Between Hypotheses and Facts. Mol. Immunol. 1993, 30, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Uchida, K.; Nakagaki, K.; Trapnell, B.C.; Nakata, K. High Avidity Cytokine Autoantibodies in Health and Disease: Pathogenesis and Mechanisms. Cytokine Growth Factor Rev. 2010, 21, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Mannoor, K.; Xu, Y.; Chen, C. Natural Autoantibodies and Associated B Cells in Immunity and Autoimmunity. Autoimmunity 2013, 46, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Grönwall, C.; Silverman, G.J. Natural IgM: Beneficial Autoantibodies for the Control of Inflammatory and Autoimmune Disease. J. Clin. Immunol. 2014, 34, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Nagele, E.P.; Han, M.; Acharya, N.K.; DeMarshall, C.; Kosciuk, M.C.; Nagele, R.G. Natural IgG Autoantibodies Are Abundant and Ubiquitous in Human Sera, and Their Number Is Influenced By Age, Gender, and Disease. PLoS ONE 2013, 8, e60726. [Google Scholar] [CrossRef]
- Czömpöly, T.; Olasz, K.; Simon, D.; Nyárády, Z.; Pálinkás, L.; Czirják, L.; Berki, T.; Németh, P. A Possible New Bridge between Innate and Adaptive Immunity: Are the Anti-Mitochondrial Citrate Synthase Autoantibodies Components of the Natural Antibody Network? Mol. Immunol. 2006, 43, 1761–1768. [Google Scholar] [CrossRef]
- Simon, D.; Balogh, P.; Erdő-Bonyár, S.; Böröcz, K.; Minier, T.; Czirják, L.; Berki, T. Increased Frequency of Activated Switched Memory B Cells and Its Association With the Presence of Pulmonary Fibrosis in Diffuse Cutaneous Systemic Sclerosis Patients. Front. Immunol. 2021, 12, 686483. [Google Scholar] [CrossRef]
- Böröcz, K.; Kinyó, Á.; Simon, D.; Erdő-Bonyár, S.; Németh, P.; Berki, T. Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. Int. J. Mol. Sci. 2023, 24, 6439. [Google Scholar] [CrossRef]
- Böröcz, K.; Simon, D.; Erdő-Bonyár, S.; Kovács, K.T.; Tuba, É.; Czirják, L.; Németh, P.; Berki, T. Relationship between Natural and Infection-Induced Antibodies in Systemic Autoimmune Diseases (SAD): SLE, SSc and RA. Clin. Exp. Immunol. 2021, 203, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Szinger, D.; Berki, T.; Németh, P.; Erdo-Bonyar, S.; Simon, D.; Drenjančević, I.; Samardzic, S.; Zelić, M.; Sikora, M.; Požgain, A.; et al. Following Natural Autoantibodies: Further Immunoserological Evidence Regarding Their Silent Plasticity and Engagement in Immune Activation. Int. J. Mol. Sci. 2023, 24, 14961. [Google Scholar] [CrossRef]
- Molvarec, A.; Derzsy, Z.; Kocsis, J.; Bäze, T.; Nagy, B.; Balogh, K.; Makó, V.; Cervenak, L.; Mézes, M.; Karádi, I.; et al. Circulating Anti-Heat-Shock-Protein Antibodies in Normal Pregnancy and Preeclampsia. Cell Stress Chaperones 2009, 14, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, K.B.; Muzzio, D.O.; Matzner, F.; Bommer, I.; Malinowsky, K.; Ehrhardt, J.; Ventimiglia, M.S.; Zygmunt, M.; Jensen, F. Human Pregnancy Is Accompanied by Modifications in B Cell Development and Immunoglobulin Profile. J. Reprod. Immunol. 2018, 129, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Thangaratinam, S.; Tan, A.; Knox, E.; Kilby, M.D.; Franklyn, J.; Coomarasamy, A. Association between Thyroid Autoantibodies and Miscarriage and Preterm Birth: Meta-Analysis of Evidence. BMJ 2011, 342, d2616. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.R. Biomarkers, Self-Antigens and the Immunological Homunculus. J. Autoimmun. 2007, 29, 246–249. [Google Scholar] [CrossRef]
- Pockley, A.G. Heat Shock Proteins as Regulators of the Immune Response. Lancet 2003, 362, 469–476. [Google Scholar] [CrossRef]
- Prohászka, Z.; Singh, M.; Nagy, K.; Kiss, E.; Lakos, G.; Duba, J.; Füst, G. Heat Shock Protein 70 Is a Potent Activator of the Human Complement System. Cell Stress Chaperones 2002, 7, 17. [Google Scholar] [CrossRef]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The Chaperone Function of hsp70 Is Required for Protection against Stress-Induced Apoptosis. Mol. Cell. Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef]
- Jee, B.; Dhar, R.; Singh, S.; Karmakar, S. Heat Shock Proteins and Their Role in Pregnancy: Redefining the Function of “Old Rum in a New Bottle”. Front. Cell Dev. Biol. 2021, 9, 648463. [Google Scholar] [CrossRef]
- Li, D.G.; Gordon, C.B.; Stagg, C.A.; Udelsman, R. Heat Shock Protein Expression in Human Placenta and Umbilical Cord. Shock 1996, 5, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Kawahara, H.; Isurugi, C.; Syoji, T.; Oyama, R.; Sugiyama, T.; Horiuchi, S. Changes in Serum Levels of Heat Shock Protein 70 in Preterm Delivery and Pre-Eclampsia. J. Obstet. Gynaecol. Res. 2005, 31, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Saghafi, N.; Poural, L.; Ghanbarabadi, V.G.; Mirzamarjani, F.; Mirteimouri, M. Serum Heat Shock Protein 70 in Preeclampsia and Normal Pregnancy: A Systematic Review and Meta-Analysis. Int. J. Reprod. Biomed. 2018, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Molvarec, A.; Tamási, L.; Losonczy, G.; Madách, K.; Prohászka, Z.; Rigó, J. Circulating Heat Shock Protein 70 (HSPA1A) in Normal and Pathological Pregnancies. Cell Stress Chaperones 2010, 15, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Madách, K.; Molvarec, A.; Rigó, J.; Nagy, B.; Pénzes, I.; Karádi, I.; Prohászka, Z. Elevated Serum 70 kDa Heat Shock Protein Level Reflects Tissue Damage and Disease Severity in the Syndrome of Hemolysis, Elevated Liver Enzymes, and Low Platelet Count. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 139, 133–138. [Google Scholar] [CrossRef]
- Garamvölgyi, Z.; Prohászka, Z.; Rigó, J.; Kecskeméti, A.; Molvarec, A. Increased Circulating Heat Shock Protein 70 (HSPA1A) Levels in Gestational Diabetes Mellitus: A Pilot Study. Cell Stress Chaperones 2015, 20, 575–581. [Google Scholar] [CrossRef]
- O’Keefe, D.J. Brief Report: Post Hoc Power, Observed Power, A Priori Power, Retrospective Power, Prospective Power, Achieved Power: Sorting Out Appropriate Uses of Statistical Power Analyses. Commun. Methods Meas. 2007, 1, 291–299. [Google Scholar] [CrossRef]
- Hoenig, J.M.; Heisey, D.M. The Abuse of Power: The Pervasive Fallacy of Power Calculations for Data Analysis. Am. Stat. 2001, 55, 19–24. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simon, D.; Erdő-Bonyár, S.; Böröcz, K.; Balázs, N.; Badawy, A.; Bajnok, A.; Nörenberg, J.; Serény-Litvai, T.; Várnagy, Á.; Kovács, K.; et al. Altered Levels of Natural Autoantibodies against Heat Shock Proteins in Pregnant Women with Hashimoto’s Thyroiditis. Int. J. Mol. Sci. 2024, 25, 1423. https://doi.org/10.3390/ijms25031423
Simon D, Erdő-Bonyár S, Böröcz K, Balázs N, Badawy A, Bajnok A, Nörenberg J, Serény-Litvai T, Várnagy Á, Kovács K, et al. Altered Levels of Natural Autoantibodies against Heat Shock Proteins in Pregnant Women with Hashimoto’s Thyroiditis. International Journal of Molecular Sciences. 2024; 25(3):1423. https://doi.org/10.3390/ijms25031423
Chicago/Turabian StyleSimon, Diána, Szabina Erdő-Bonyár, Katalin Böröcz, Noémi Balázs, Ahmed Badawy, Anna Bajnok, Jasper Nörenberg, Tímea Serény-Litvai, Ákos Várnagy, Kálmán Kovács, and et al. 2024. "Altered Levels of Natural Autoantibodies against Heat Shock Proteins in Pregnant Women with Hashimoto’s Thyroiditis" International Journal of Molecular Sciences 25, no. 3: 1423. https://doi.org/10.3390/ijms25031423
APA StyleSimon, D., Erdő-Bonyár, S., Böröcz, K., Balázs, N., Badawy, A., Bajnok, A., Nörenberg, J., Serény-Litvai, T., Várnagy, Á., Kovács, K., Hantosi, E., Mezősi, E., Németh, P., & Berki, T. (2024). Altered Levels of Natural Autoantibodies against Heat Shock Proteins in Pregnant Women with Hashimoto’s Thyroiditis. International Journal of Molecular Sciences, 25(3), 1423. https://doi.org/10.3390/ijms25031423