

Genome-Wide Identification of the Trihelix Transcription Factor Family and Functional Analysis of ZmTHX15 in Maize
 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Identification and Physicochemical Properties of ZmTHXs
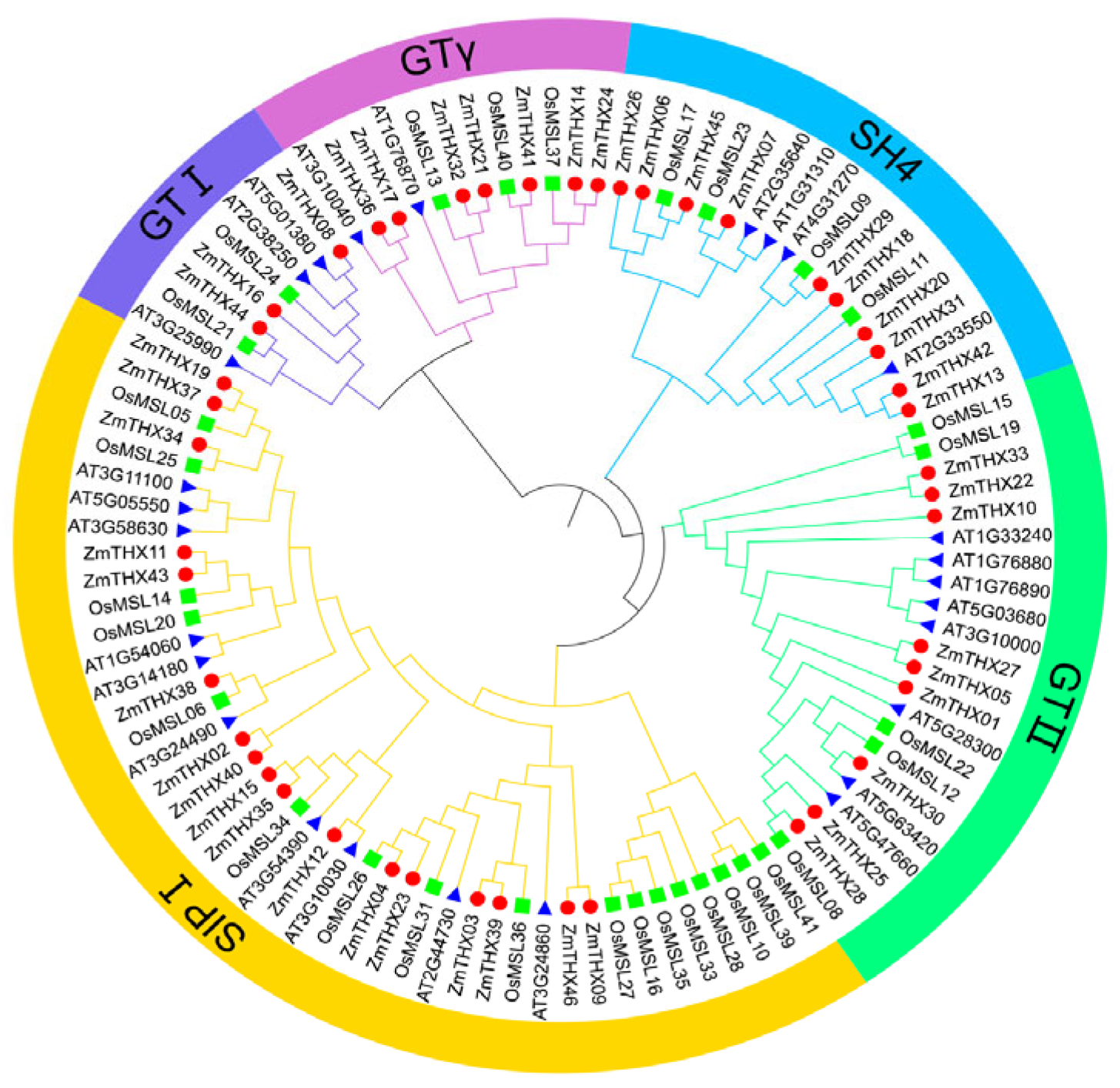
2.2. Phylogenetic Analysis and Subfamily Classification of ZmTHXs
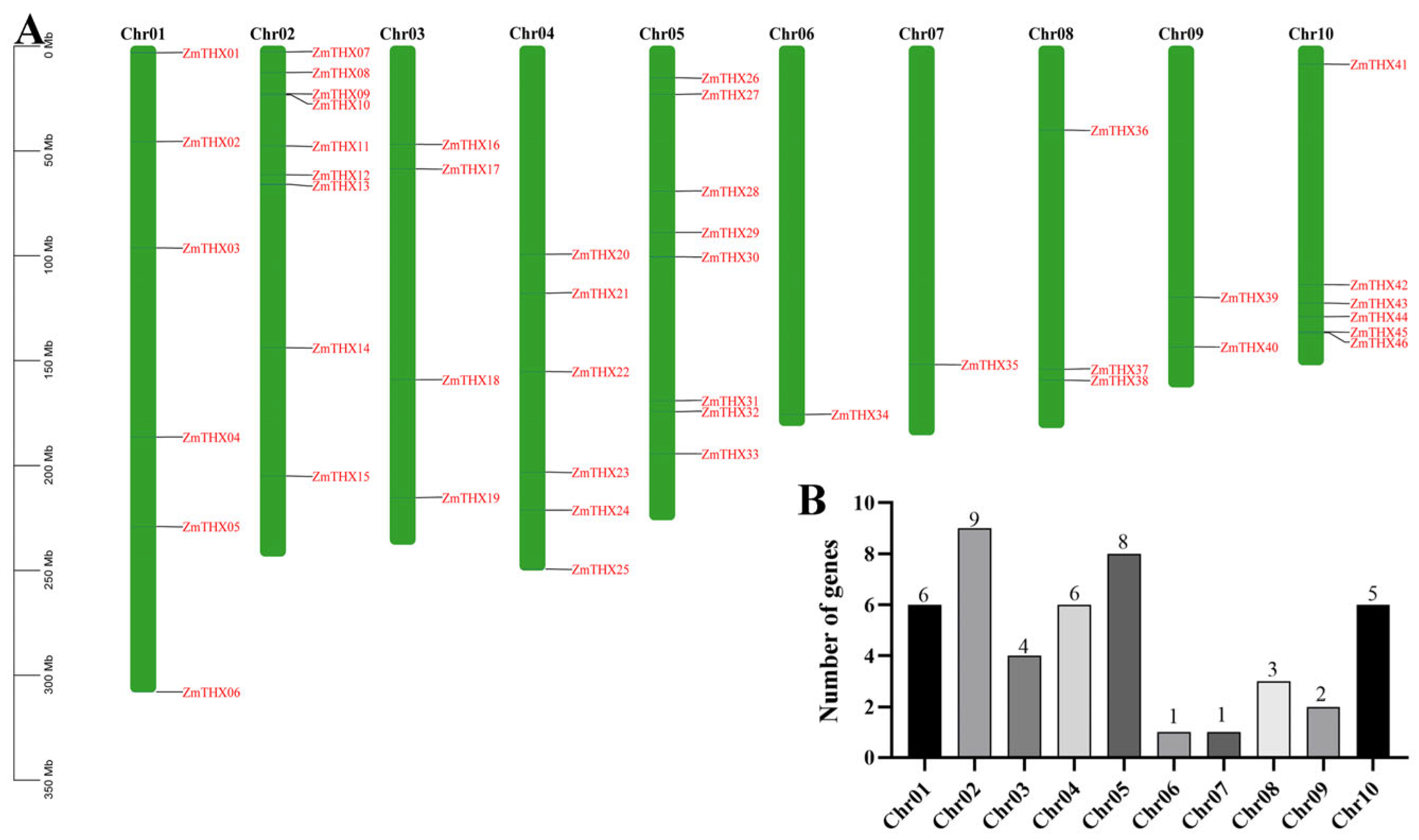
2.3. Chromosomal Localization and Synteny Analysis of ZmTHXs
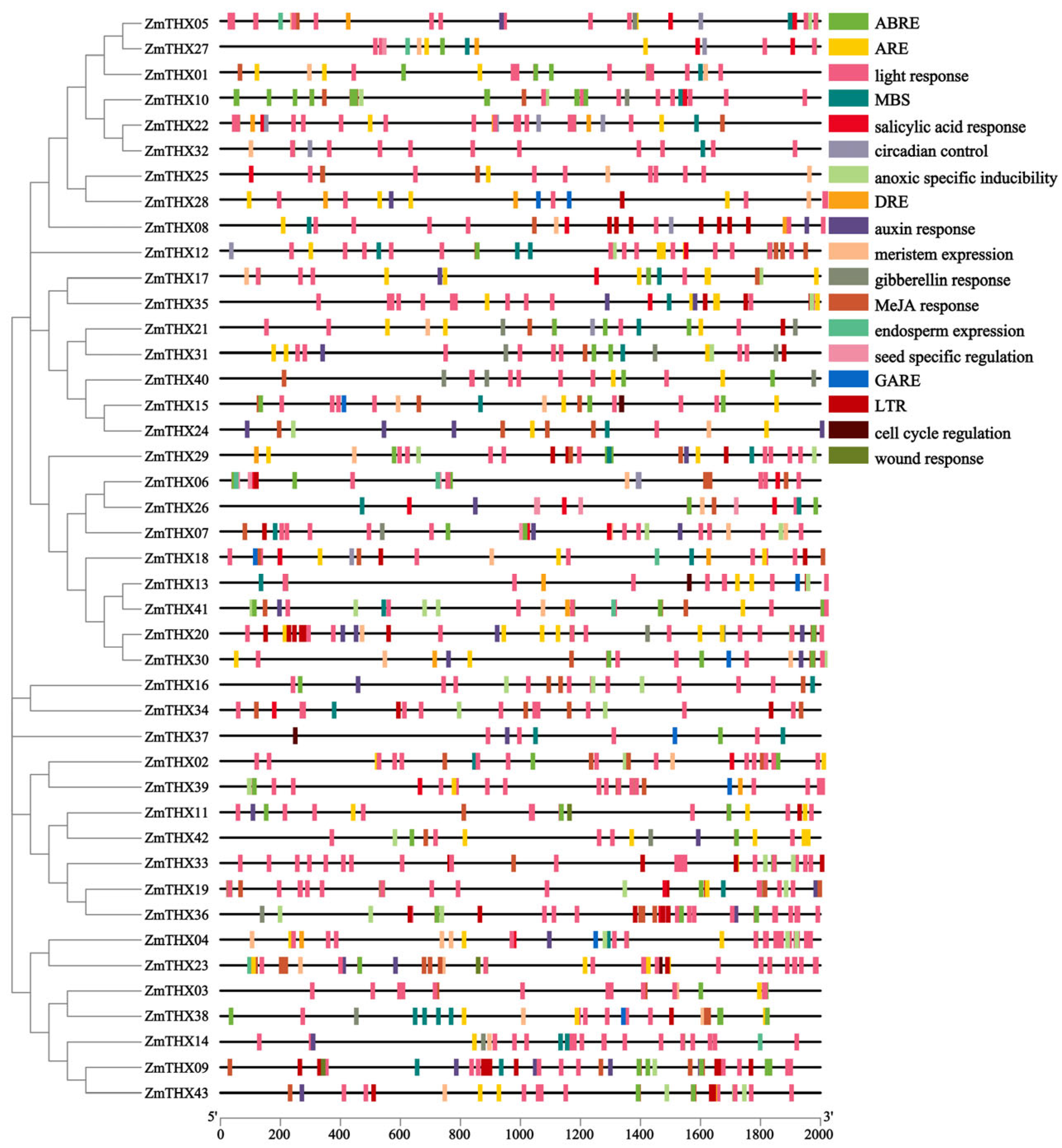
2.4. Analysis of Cis-Acting Elements in ZmTHXs
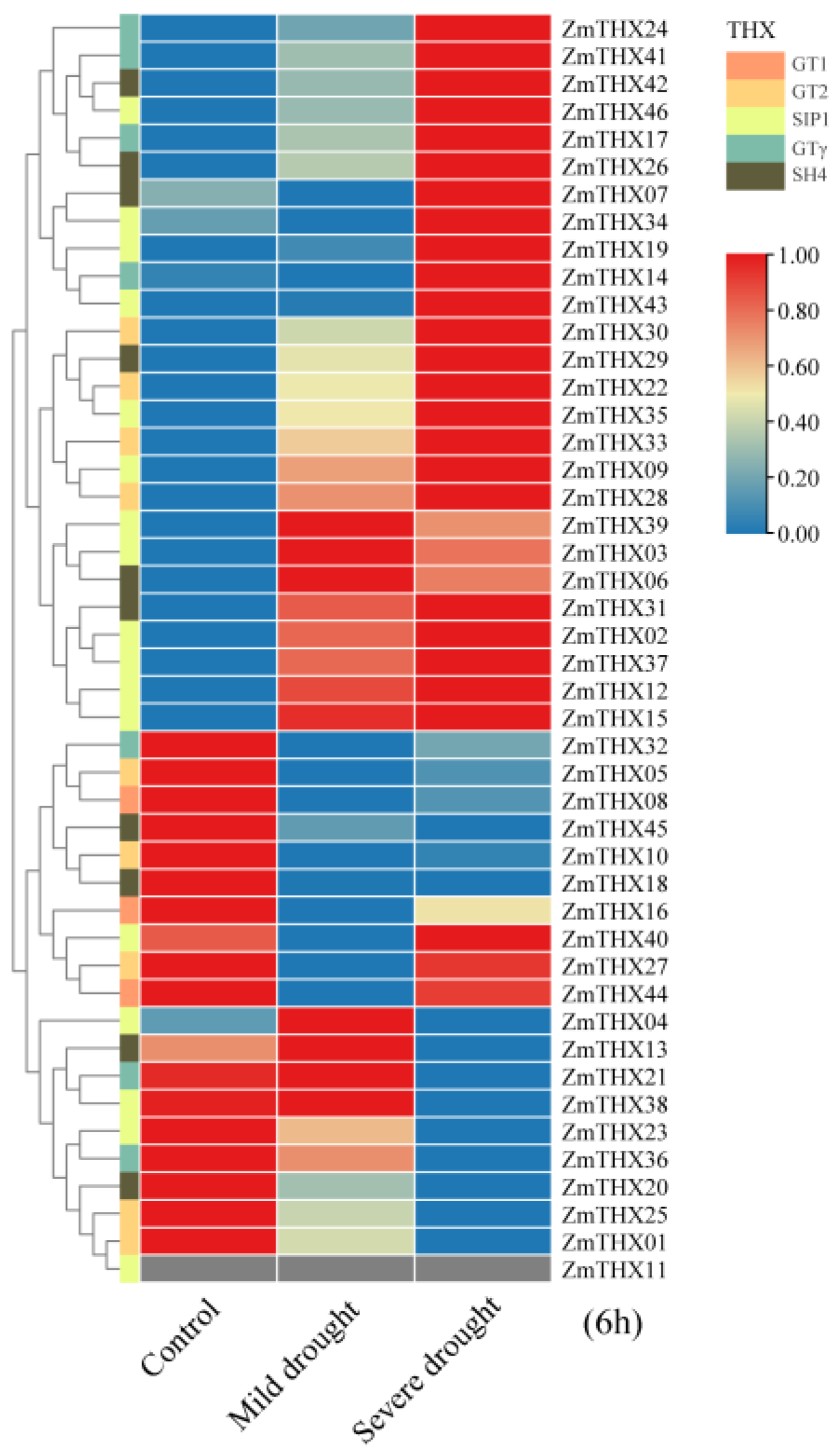
2.5. Gene Expression Profile Analysis
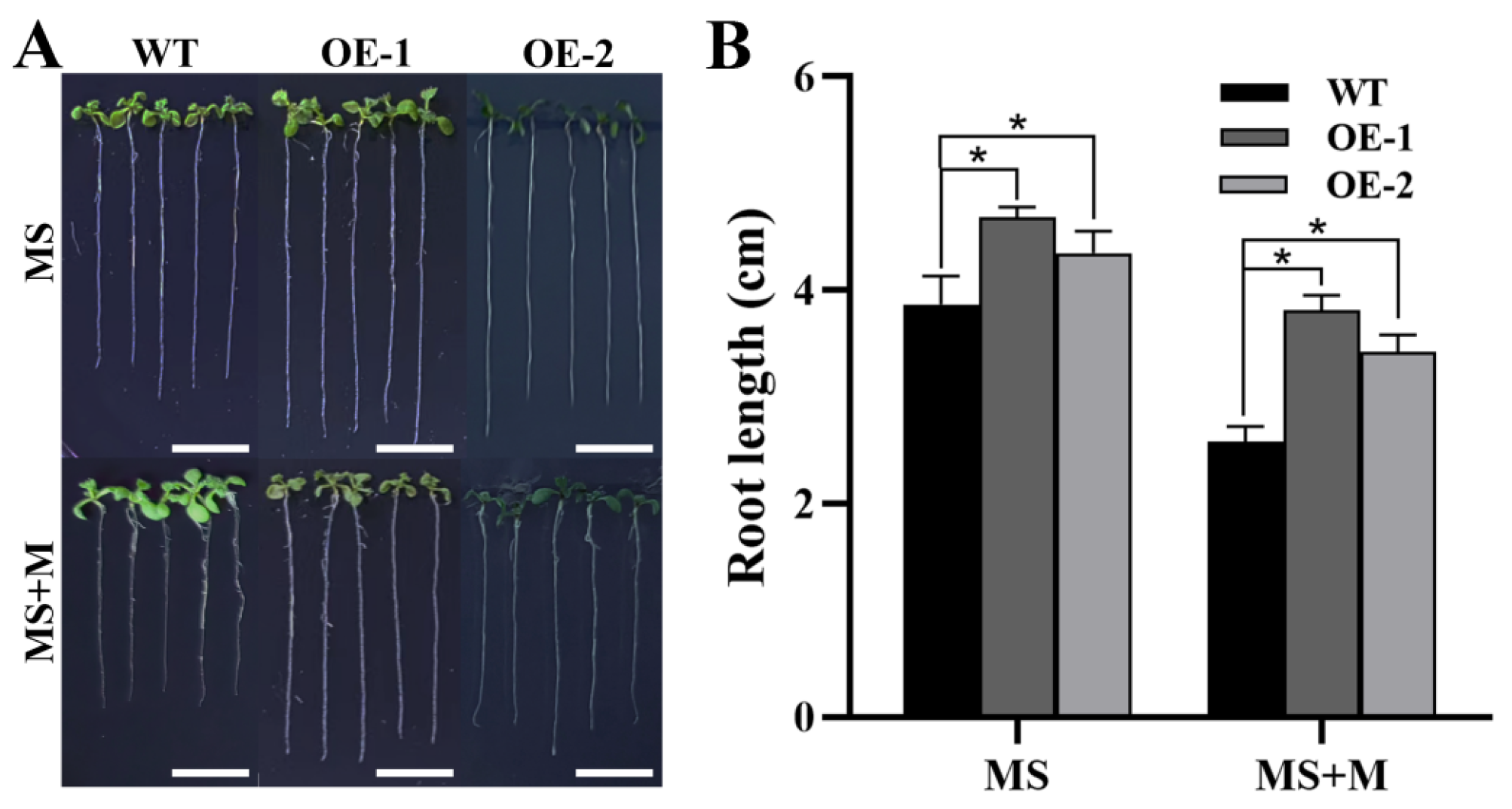
2.6. ZmTHX15 Involved in Osmotic Stress Regulation
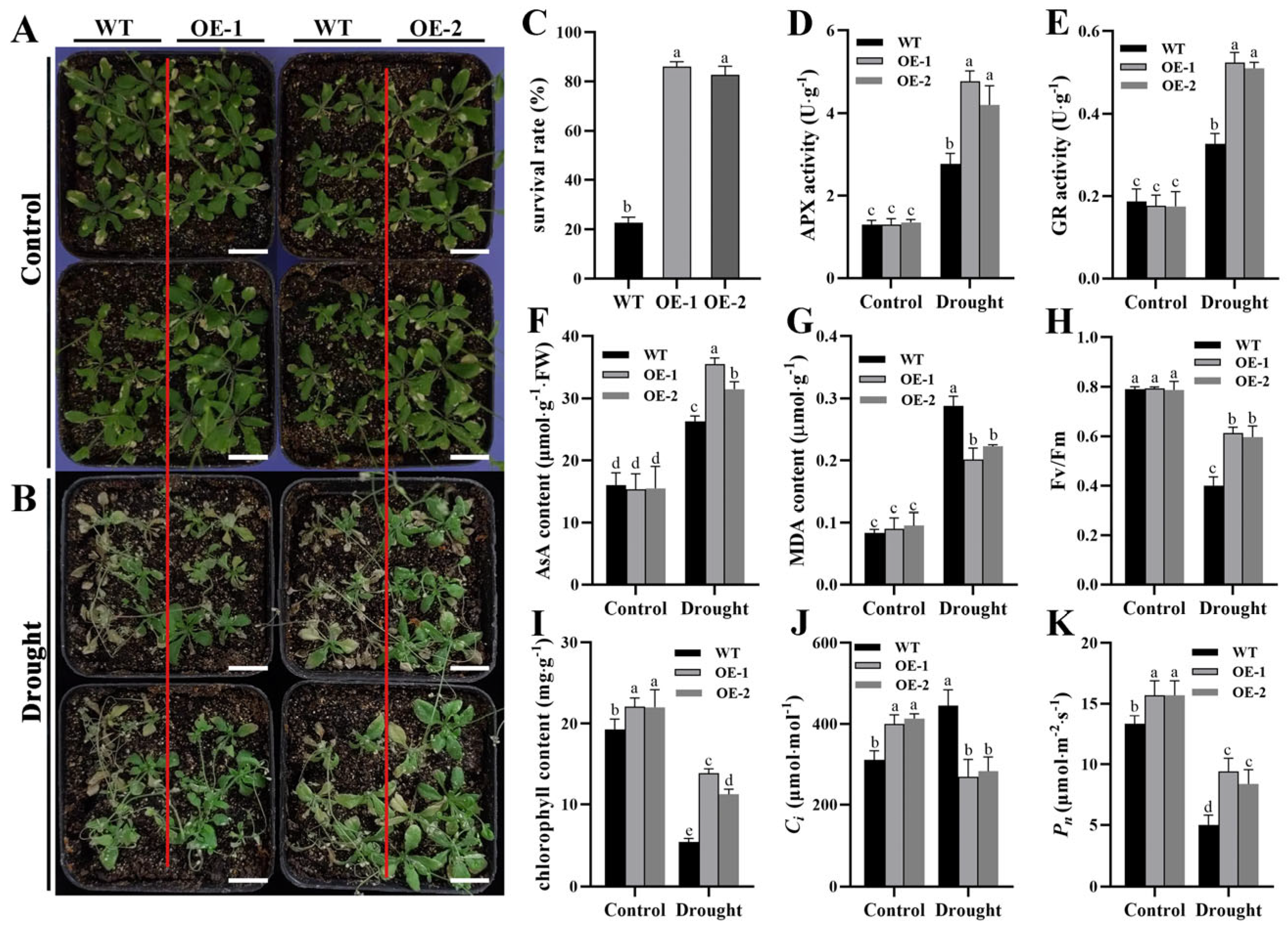
2.7. ZmTHX15-Overexpressing Arabidopsis Is More Drought Tolerant than WT
3. Discussion
4. Materials and Methods
4.1. Identification and Physicochemical Properties of ZmTHXs
4.2. Phylogenetic Analysis of the ZmTHXs Gene Family
4.3. Cis-Acting Element Analysis of ZmTHXs
4.4. Chromosomal Localization and Synteny Analysis of ZmTHXs
4.5. Differential Expression Analysis of ZmTHXs Under Drought Stress
4.6. Gene Cloning
4.7. Generation of Transgenic Plants
4.8. Mannitol Treatment of Transgenic Arabidopsis
4.9. Measurement of Chlorophyll Content
4.10. Measurement of Photosynthetic Rate (Pn) and Intercellular CO2 Concentration
4.11. Chlorophyll Fluorescence Measurement
4.12. APX and GR Enzyme Activity
4.13. Determination of AsA and MDA Content
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yanagisawa, S. Dof domain proteins: Plant-specific transcription factors associated with diverse phenomena unique to plants. Plant Cell Physiol. 2004, 45, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Manna, M.; Thakur, T.; Chirom, O.; Mandlik, R.; Deshmukh, R.; Salvi, P. Transcription factors as key molecular target to strengthen the drought stress tolerance in plants. Physiol. Plant. 2021, 172, 847–868. [Google Scholar] [CrossRef] [PubMed]
- Green, P.J.; Yong, M.H.; Cuozzo, M.; Kano-Murakami, Y.; Silverstein, P.; Chua, N.H. Binding site requirements for pea nuclear protein factor GT-1 correlate with sequences required for light-dependent transcriptional activation of the rbcS-3A gene. EMBO J. 1988, 7, 4035–4044. [Google Scholar] [CrossRef] [PubMed]
- Green, P.J.; Kay, S.A.; Chua, N.H. Sequence-specific interactions of a pea nuclear factor with light-responsive elements upstream of the rbcS-3A gene. EMBO J. 1987, 6, 2543–2549. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xie, K.; Hou, X.; Hu, H.; Xiong, L. Systematic analysis of GT factor family of rice reveals a novel subfamily involved in stress responses. Mol. Genet. Genom. 2010, 283, 157–169. [Google Scholar] [CrossRef]
- Nagano, Y. Several features of the GT-factor trihelix domain resemble those of the Myb DNA-binding domain. Plant Physiol. 2000, 124, 491–494. [Google Scholar] [CrossRef]
- Qin, Y.; Ma, X.; Yu, G.; Wang, Q.; Wang, L.; Kong, L.; Kim, W.; Wang, H.W. Evolutionary history of trihelix family and their functional diversification. DNA Res. 2014, 21, 499–510. [Google Scholar] [CrossRef]
- Ayadi, M.; Delaporte, V.; Li, Y.F.; Zhou, D.X. Analysis of GT-3a identifies a distinct subgroup of trihelix DNA-binding transcription factors in Arabidopsis. FEBS Lett. 2004, 562, 147–154. [Google Scholar] [CrossRef]
- Barr, M.S.; Willmann, M.R.; Jenik, P.D. Is there a role for trihelix transcription factors in embryo maturation? Plant Signal. Behav. 2012, 7, 205–209. [Google Scholar] [CrossRef]
- Breuer, C.; Kawamura, A.; Ichikawa, T.; Tominaga-Wada, R.; Wada, T.; Kondou, Y.; Muto, S.; Matsui, M.; Sugimoto, K. The trihelix transcription factor GTL1 regulates ploidy-dependent cell growth in the Arabidopsis trichome. Plant Cell 2009, 21, 2307–2322. [Google Scholar] [CrossRef]
- Fu, M.; Li, F.; Zhou, S.; Guo, P.; Chen, Y.; Xie, Q.; Chen, G.; Hu, Z. Trihelix transcription factor SlGT31 regulates fruit ripening mediated by ethylene in tomato. J. Exp. Bot. 2023, 74, 5709–5721. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.J.; Lydiate, D.J.; Li, X.; Lui, H.; Gjetvaj, B.; Hegedus, D.D.; Rozwadowski, K. Repression of seed maturation genes by a trihelix transcriptional repressor in Arabidopsis seedlings. Plant Cell 2009, 21, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Levy, R.N.; Brewer, P.B.; Quon, T.; Smyth, D.R. The trihelix family of transcription factors—Light, stress and development. Trends Plant Sci. 2012, 17, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Breuer, C.; Kawamura, A.; Clark, N.M.; Rymen, B.; Braidwood, L.; Morohashi, K.; Busch, W.; Benfey, P.N.; Sozzani, R.; et al. GTL1 and DF1 regulate root hair growth through transcriptional repression of root hair defective 6-LIKE 4 in Arabidopsis. Development 2018, 145, dev159707. [Google Scholar] [CrossRef]
- Brewer, P.B.; Howles, P.A.; Dorian, K.; Griffith, M.E.; Ishida, T.; Kaplan-Levy, R.N.; Kilinc, A.; Smyth, D.R. PETAL LOSS, a trihelix transcription factor gene, regulates perianth architecture in the Arabidopsis flower. Development 2004, 131, 4035–4045. [Google Scholar] [CrossRef]
- Chen, H.; Zou, W.; Zhao, J. Ribonuclease J is required for chloroplast and embryo development in Arabidopsis. J. Exp. Bot. 2015, 66, 2079–2091. [Google Scholar] [CrossRef]
- Lin, Z.; Griffith, M.E.; Li, X.; Zhu, Z.; Tan, L.; Fu, Y.; Zhang, W.; Wang, X.; Xie, D.; Sun, C. Origin of seed shattering in rice (Oryza sativa L.). Planta 2007, 226, 11–20. [Google Scholar] [CrossRef]
- Hofmann, N.R. Downstream of a kinase cascade: A trihelix transcription factor represses immune genes. Plant Cell 2015, 27, 481. [Google Scholar] [CrossRef]
- Li, B.; Jiang, S.; Yu, X.; Cheng, C.; Chen, S.; Cheng, Y.; Yuan, J.S.; Jiang, D.; He, P.; Shan, L. Phosphorylation of trihelix transcriptional repressor ASR3 by MAP KINASE4 negatively regulates Arabidopsis immunity. Plant Cell 2015, 27, 839–856. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, M.; Zhang, Y.; Huang, M.; Wei, L.; Lin, Y.; Xie, J.; Liu, J.; Fu, Y.; Jiang, D.; et al. Coordinated regulation of plant defense and autoimmunity by paired trihelix transcription factors ASR3/AITF1 in Arabidopsis. New Phytol. 2023, 237, 914–929. [Google Scholar] [CrossRef]
- Mao, H.; Zhang, W.; Lv, J.; Yang, J.; Yang, S.; Jia, B.; Song, J.; Wu, M.; Pei, W.; Ma, J.; et al. Overexpression of cotton Trihelix transcription factor GhGT-3b_A04 enhances resistance to Verticillium dahliae and affects plant growth in Arabidopsis thaliana. J. Plant Physiol. 2023, 283, 153947. [Google Scholar] [CrossRef] [PubMed]
- Volz, R.; Kim, S.K.; Mi, J.; Mariappan, K.G.; Guo, X.; Bigeard, J.; Alejandro, S.; Pflieger, D.; Rayapuram, N.; Al-Babili, S.; et al. The Trihelix transcription factor GT2-like 1 (GTL1) promotes salicylic acid metabolism, and regulates bacterial-triggered immunity. PLoS Genet. 2018, 14, e1007708. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, G.; Zhang, J.; Xuan, J. E3 Ubiquitin Ligase PUB23 in Kiwifruit Interacts with Trihelix Transcription Factor GT1 and Negatively Regulates Immune Responses Against Pseudomonas syringae pv. actinidiae. Int. J. Mol. Sci. 2024, 25, 1930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhong, T.; Linzhu, E.; Xu, M.; Dai, W.; Sun, S.; Ye, J. GT Factor ZmGT-3b Is Associated with Regulation of Photosynthesis and Defense Response to Fusarium graminearum Infection in Maize Seedling. Front. Plant Sci. 2021, 12, 724133. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Li, Q.T.; Chen, H.W.; Zhang, W.K.; Ma, B.; Chen, S.Y.; Zhang, J.S. Trihelix transcription factor GT-4 mediates salt tolerance via interaction with TEM2 in Arabidopsis. BMC Plant Biol. 2014, 14, 339. [Google Scholar] [CrossRef]
- Liu, X.; Wu, D.; Shan, T.; Xu, S.; Qin, R.; Li, H.; Negm, M.; Wu, D.; Li, J. The trihelix transcription factor OsGTgamma-2 is involved adaption to salt stress in rice. Plant Mol. Biol. 2020, 103, 545–560. [Google Scholar] [CrossRef]
- Xi, J.; Qiu, Y.; Du, L.; Poovaiah, B.W. Plant-specific trihelix transcription factor AtGT2L interacts with calcium/calmodulin and responds to cold and salt stresses. Plant Sci. 2012, 185–186, 274–280. [Google Scholar] [CrossRef]
- Li, Y.; Hu, Z.; Dong, Y.; Xie, Z. Trihelix Transcriptional Factor GhGT26 of Cotton Enhances Salinity Tolerance in Arabidopsis. Plants 2022, 11, 2694. [Google Scholar] [CrossRef]
- Xu, H.; Shi, X.; He, L.; Guo, Y.; Zang, D.; Li, H.; Zhang, W.; Wang, Y. Arabidopsis thaliana Trihelix Transcription Factor AST1 Mediates Salt and Osmotic Stress Tolerance by Binding to a Novel AGAG-Box and Some GT Motifs. Plant Cell Physiol. 2018, 59, 946–965. [Google Scholar] [CrossRef]
- Yoo, C.Y.; Pence, H.E.; Jin, J.B.; Miura, K.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. The Arabidopsis GTL1 transcription factor regulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1. Plant Cell 2010, 22, 4128–4141. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, H.; Ji, H.; Wang, Y.; Dong, B.; Qiao, Y.; Liu, M.; Li, X. The Wheat GT Factor TaGT2L1D Negatively Regulates Drought Tolerance and Plant Development. Sci. Rep. 2016, 6, 27042. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.; Yoo, C.Y.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. Poplar GTL1 is a Ca2+/calmodulin-binding transcription factor that functions in plant water use efficiency and drought tolerance. PLoS ONE 2012, 7, e32925. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hu, Z.; Dong, Y.; Xie, Z. Overexpression of the cotton trihelix transcription factor GhGT23 in Arabidopsis mediates salt and drought stress tolerance by binding to GT and MYB promoter elements in stress-related genes. Front. Plant Sci. 2023, 14, 1144650. [Google Scholar] [CrossRef] [PubMed]
- Magwanga, R.O.; Kirungu, J.N.; Lu, P.; Yang, X.; Dong, Q.; Cai, X.; Xu, Y.; Wang, X.; Zhou, Z.; Hou, Y.; et al. Genome wide identification of the trihelix transcription factors and overexpression of Gh_A05G2067 (GT-2), a novel gene contributing to increased drought and salt stresses tolerance in cotton. Physiol. Plant. 2019, 167, 447–464. [Google Scholar] [CrossRef]
- Yu, C.; Song, L.; Song, J.; Ouyang, B.; Guo, L.; Shang, L.; Wang, T.; Li, H.; Zhang, J.; Ye, Z. ShCIGT, a Trihelix family gene, mediates cold and drought tolerance by interacting with SnRK1 in tomato. Plant Sci. 2018, 270, 140–149. [Google Scholar] [CrossRef]
- Li, F.; Chen, G.; Xie, Q.; Zhou, S.; Hu, Z. Down-regulation of SlGT-26 gene confers dwarf plants and enhances drought and salt stress resistance in tomato. Plant Physiol. Biochem. 2023, 203, 108053. [Google Scholar] [CrossRef]
- Lv, H.; Wang, X.; Dong, X.; Gao, M.; Dong, D.; Li, C.; Jing, S.; Guo, Y.D.; Zhang, N. CRISPR/Cas9 edited SlGT30 improved both drought resistance and fruit yield through endoreduplication. Plant. Cell Environ. 2024. early view. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Ng, D.W.; Abeysinghe, J.K.; Kamali, M. Regulating the Regulators: The Control of Transcription Factors in Plant Defense Signaling. Int. J. Mol. Sci. 2018, 19, 3737. [Google Scholar] [CrossRef]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2008, 133, 481–489. [Google Scholar] [CrossRef]
- Umezawa, T.; Nakashima, K.; Miyakawa, T.; Kuromori, T.; Tanokura, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular basis of the core regulatory network in ABA responses: Sensing, signaling and transport. Plant Cell Physiol. 2010, 51, 1821–1839. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zeng, B.; Zhao, H.; Zhang, M.; Xie, S.; Lai, J. Genome-wide transcription factor gene prediction and their expressional tissue-specificities in maize. J. Integr. Plant Biol. 2012, 54, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Reboledo, G.; Agorio, A.; Ponce De Leon, I. Moss transcription factors regulating development and defense responses to stress. J. Exp. Bot. 2022, 73, 4546–4561. [Google Scholar] [CrossRef]
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27, 922–935. [Google Scholar] [CrossRef]
- Zhao, Y.; Liang, J.; Wang, Z.; Yan, T.; Yan, X.; Wei, W.; Le, M.; Sun, J. Genome-wide identification and expression analysis of the trihelix transcription factor family in sesame (Sesamum indicum L.) under abiotic stress. Mol. Biol. Rep. 2023, 50, 8281–8295. [Google Scholar] [CrossRef]
- Li, K.; Fan, Y.; Zhou, G.; Liu, X.; Chen, S.; Chang, X.; Wu, W.; Duan, L.; Yao, M.; Wang, R.; et al. Genome-wide identification, phylogenetic analysis, and expression profiles of trihelix transcription factor family genes in quinoa (Chenopodium quinoa Willd.) under abiotic stress conditions. BMC Genom. 2022, 23, 499. [Google Scholar] [CrossRef]
- Li, K.; Duan, L.; Zhang, Y.; Shi, M.; Chen, S.; Yang, M.; Ding, Y.; Peng, Y.; Dong, Y.; Yang, H.; et al. Genome-wide identification and expression profile analysis of trihelix transcription factor family genes in response to abiotic stress in sorghum [Sorghum bicolor (L.) Moench]. BMC Genom. 2021, 22, 738. [Google Scholar] [CrossRef]
- Ma, Z.; Liu, M.; Sun, W.; Huang, L.; Wu, Q.; Bu, T.; Li, C.; Chen, H. Genome-wide identification and expression analysis of the trihelix transcription factor family in tartary buckwheat (Fagopyrum tataricum). BMC Plant Biol. 2019, 19, 344. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Ma, L.; Wang, Z.; Wang, K. Genome-Wide Identification and Expression Profiling Analysis of the Trihelix Gene Family Under Abiotic Stresses in Medicago truncatula. Genes 2020, 11, 1389. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Hu, R.; Gu, T.; Han, J.; Qiu, D.; Su, P.; Feng, J.; Chang, J.; Yang, G.; He, G. Genome-wide identification and expression profiling of trihelix gene family under abiotic stresses in wheat. BMC Genom. 2019, 20, 287. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Wang, J.; Liu, H.; Zheng, H.; Zhen, Z.; Zhao, H.; et al. Genome-Wide Characterization and Identification of Trihelix Transcription Factor and Expression Profiling in Response to Abiotic Stresses in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 251. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Cai, X.; Ye, Z.; Li, H. Genome-wide identification and expression profiling analysis of trihelix gene family in tomato. Biochem. Biophys. Res. Commun. 2015, 468, 653–659. [Google Scholar] [CrossRef]
- Zhao, D.; Gao, F.; Guan, P.; Gao, J.; Guo, Z.; Guo, J.; Cui, H.; Li, Y.; Zhang, G.; Li, Z.; et al. Identification and analysis of differentially expressed trihelix genes in maize (Zea mays) under abiotic stresses. PeerJ 2023, 11, e15312. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef]
- Hung, J.H.; Weng, Z. Sequence Alignment and Homology Search with BLAST and ClustalW. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot093088. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis Across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Rombauts, S.; Dehais, P.; Van Montagu, M.; Rouze, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Zhang, J.; Pai, Q.; Yue, L.; Wu, X.; Liu, H.; Wang, W. Cytokinin regulates female gametophyte development by cell cycle modulation in Arabidopsis thaliana. Plant Sci. 2022, 324, 111419. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, T.; Duan, X.; Wei, X.; Shi, T.; Li, J.; Russell, S.D.; Gou, X. Cis-Regulatory Elements Determine Germline Specificity and Expression Level of an Isopentenyltransferase Gene in Sperm Cells of Arabidopsis. Plant Physiol. 2016, 170, 1524–1534. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Gene ID | Amino Acids | MW (KDa) | pI | Instability Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| ZmTHX01 | Zm00001eb000880 | 809 | 84,901.28 | 6.55 | 69.6 | −0.736 | Nucleus |
| ZmTHX02 | Zm00001eb013590 | 377 | 40,843.3 | 10.12 | 57.94 | −0.649 | Nucleus |
| ZmTHX03 | Zm00001eb023870 | 321 | 34,106.44 | 9.41 | 74.38 | −0.567 | Nucleus |
| ZmTHX04 | Zm00001eb033570 | 319 | 35,873.99 | 5.6 | 75.37 | −0.966 | Nucleus |
| ZmTHX05 | Zm00001eb043580 | 669 | 70,672.04 | 9.22 | 63.48 | −0.734 | Nucleus |
| ZmTHX06 | Zm00001eb065500 | 204 | 22,859.11 | 9.38 | 41.64 | −0.412 | Nucleus |
| ZmTHX07 | Zm00001eb066770 | 381 | 40,870.93 | 9.38 | 80.33 | −0.64 | Nucleus |
| ZmTHX08 | Zm00001eb071780 | 273 | 32,311.31 | 8.37 | 62.18 | −1.259 | Nucleus |
| ZmTHX09 | Zm00001eb075080 | 208 | 22,949.16 | 11.09 | 96.5 | −0.682 | Nucleus |
| ZmTHX10 | Zm00001eb075250 | 774 | 83,274.2 | 5.9 | 73.29 | −0.882 | Nucleus |
| ZmTHX11 | Zm00001eb081720 | 385 | 40,978.96 | 6.68 | 64.43 | −0.783 | Nucleus |
| ZmTHX12 | Zm00001eb084300 | 354 | 37,623.59 | 8.08 | 41.35 | −0.716 | Nucleus |
| ZmTHX13 | Zm00001eb084860 | 348 | 37,680.89 | 8.69 | 52.69 | −0.773 | Nucleus |
| ZmTHX14 | Zm00001eb092040 | 436 | 49,833.69 | 6.32 | 53.26 | −1.023 | Nucleus |
| ZmTHX15 | Zm00001eb104640 | 357 | 38,923.08 | 8.23 | 32.16 | −0.642 | Nucleus |
| ZmTHX16 | Zm00001eb129190 | 78 | 88,55.39 | 10.01 | 53.05 | −0.553 | Nucleus |
| ZmTHX17 | Zm00001eb130740 | 537 | 57,507.76 | 7.1 | 63.71 | −0.731 | Nucleus |
| ZmTHX18 | Zm00001eb142600 | 295 | 32,148.04 | 8.6 | 43.47 | −0.617 | Nucleus |
| ZmTHX19 | Zm00001eb157080 | 318 | 34,724.07 | 9.59 | 70.81 | −0.75 | Nucleus |
| ZmTHX20 | Zm00001eb180800 | 341 | 36,829.89 | 5.27 | 61.81 | −0.743 | Nucleus |
| ZmTHX21 | Zm00001eb181760 | 398 | 45,440.35 | 5.97 | 49.73 | −0.814 | Nucleus |
| ZmTHX22 | Zm00001eb186200 | 668 | 71,709.6 | 5.78 | 55.45 | −0.758 | Nucleus |
| ZmTHX23 | Zm00001eb199110 | 262 | 28,997.61 | 9.49 | 64.97 | −0.834 | Nucleus |
| ZmTHX24 | Zm00001eb202410 | 426 | 48,757.53 | 6.64 | 55.07 | −1.055 | Nucleus |
| ZmTHX25 | Zm00001eb209880 | 714 | 76,269.47 | 5.93 | 56.8 | −0.862 | Nucleus |
| ZmTHX26 | Zm00001eb217780 | 360 | 39,267.53 | 6.2 | 64.99 | −0.882 | Nucleus |
| ZmTHX27 | Zm00001eb220250 | 777 | 82,583.32 | 6.11 | 57.98 | −0.827 | Nucleus |
| ZmTHX28 | Zm00001eb229400 | 476 | 51,138.97 | 4.98 | 59.64 | −0.848 | Nucleus |
| ZmTHX29 | Zm00001eb233350 | 387 | 40,507.55 | 4.55 | 66.65 | −0.522 | Nucleus |
| ZmTHX30 | Zm00001eb234620 | 874 | 96,306.71 | 8.96 | 40.31 | −0.296 | Chloroplast |
| ZmTHX31 | Zm00001eb242030 | 349 | 37,637.71 | 5.05 | 64.55 | −0.662 | Nucleus |
| ZmTHX32 | Zm00001eb243260 | 405 | 46,187.92 | 5.91 | 47.34 | −0.896 | Nucleus |
| ZmTHX33 | Zm00001eb248900 | 672 | 72,812.61 | 5.87 | 51.7 | −0.865 | Nucleus |
| ZmTHX34 | Zm00001eb295700 | 343 | 36,918.88 | 9.91 | 62.4 | −0.57 | Nucleus |
| ZmTHX35 | Zm00001eb320470 | 337 | 36,717.5 | 6.94 | 37.51 | −0.631 | Nucleus |
| ZmTHX36 | Zm00001eb340550 | 533 | 57,191.2 | 6.09 | 57.65 | −0.727 | Nucleus |
| ZmTHX37 | Zm00001eb360120 | 315 | 34,672.99 | 9.87 | 70.51 | −0.824 | Nucleus |
| ZmTHX38 | Zm00001eb361590 | 350 | 38,799.78 | 9.7 | 74.3 | −0.847 | Nucleus |
| ZmTHX39 | Zm00001eb390910 | 318 | 33,599.95 | 9.17 | 70.04 | −0.458 | Nucleus |
| ZmTHX40 | Zm00001eb396910 | 251 | 27,105.29 | 6.27 | 61.82 | −0.65 | Nucleus |
| ZmTHX41 | Zm00001eb407690 | 446 | 50,509.25 | 6.11 | 46.7 | −1.011 | Nucleus |
| ZmTHX42 | Zm00001eb421380 | 334 | 36,440.24 | 5.89 | 51.63 | −0.794 | Nucleus |
| ZmTHX43 | Zm00001eb423330 | 392 | 42,106.22 | 6.61 | 66.72 | −0.846 | Nucleus |
| ZmTHX44 | Zm00001eb425230 | 379 | 41,935.71 | 6.28 | 48.46 | −0.727 | Nucleus |
| ZmTHX45 | Zm00001eb427630 | 616 | 66,455.62 | 9.34 | 71.11 | −0.931 | Nucleus |
| ZmTHX46 | Zm00001eb427790 | 206 | 22,628.8 | 11.19 | 96.72 | −0.657 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Cheng, Z.; Sun, X.; Zhu, M.; Yue, L.; Liu, H.; Wu, X.; Zhang, J.; Duan, C. Genome-Wide Identification of the Trihelix Transcription Factor Family and Functional Analysis of ZmTHX15 in Maize. Int. J. Mol. Sci. 2024, 25, 13257. https://doi.org/10.3390/ijms252413257
Cao Y, Cheng Z, Sun X, Zhu M, Yue L, Liu H, Wu X, Zhang J, Duan C. Genome-Wide Identification of the Trihelix Transcription Factor Family and Functional Analysis of ZmTHX15 in Maize. International Journal of Molecular Sciences. 2024; 25(24):13257. https://doi.org/10.3390/ijms252413257
Chicago/Turabian StyleCao, Yanyong, Zeqiang Cheng, Xinyan Sun, Meichen Zhu, Ling Yue, Hui Liu, Xiaolin Wu, Jinghua Zhang, and Canxing Duan. 2024. "Genome-Wide Identification of the Trihelix Transcription Factor Family and Functional Analysis of ZmTHX15 in Maize" International Journal of Molecular Sciences 25, no. 24: 13257. https://doi.org/10.3390/ijms252413257
APA StyleCao, Y., Cheng, Z., Sun, X., Zhu, M., Yue, L., Liu, H., Wu, X., Zhang, J., & Duan, C. (2024). Genome-Wide Identification of the Trihelix Transcription Factor Family and Functional Analysis of ZmTHX15 in Maize. International Journal of Molecular Sciences, 25(24), 13257. https://doi.org/10.3390/ijms252413257