
Recent Advances in Gene Mining and Hormonal Mechanism for Brown Planthopper Resistance in Rice
 ,
,
Abstract
1. Introduction
2. Evaluation of Rice Resistance to BPH
3. Screening of BPH-Resistant Rice Germplasms
4. Mapping of BPH Resistance Genes/QTLs in Rice
5. Cloning and Mechanistic Analysis of BPH Resistance Genes in Rice
6. Roles of Phytohormones in BPH Resistance
7. Discussion and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, L. Development of Hybrid Rice to Ensure Food Security. Rice Sci. 2014, 21, 1–2. [Google Scholar] [CrossRef]
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia-Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Du, B.; Chen, R.; Guo, J.; He, G. Current understanding of the genomic, genetic, and molecular control of insect resistance in rice. Mol. Breed. 2020, 40, 24. [Google Scholar] [CrossRef]
- Sōgawa, K. The Rice Brown Planthopper: Feeding Physiology and Host Plant Interactions. Annu. Rev. Entomol. 1982, 27, 49–73. [Google Scholar] [CrossRef]
- Min, S.; Lee, S.W.; Choi, B.R.; Lee, S.H.; Kwon, D.H. Insecticide resistance monitoring and correlation analysis to select appropriate insecticides against Nilaparvata lugens (Stål), a migratory pest in Korea. J. Asia-Pac. Entomol. 2014, 17, 711–716. [Google Scholar] [CrossRef]
- Wu, S.; Zeng, B.; Zheng, C.; Mu, X.; Zhang, Y.; Hu, J.; Zhang, S.; Gao, C.; Shen, J. The evolution of insecticide resistance in the brown planthopper (Nilaparvata lugens Stål) of China in the period 2012–2016. Sci. Rep. 2018, 8, 4586. [Google Scholar] [CrossRef]
- Ling, S.; Zhang, H.; Zhang, R. Effect of fenvalerate on the reproduction and fitness costs of the brown planthopper, Nilaparvata lugens and its resistance mechanism. Pestic. Biochem. Phys. 2011, 101, 148–153. [Google Scholar] [CrossRef]
- Liu, F.; Li, X.; Zhao, M.; Guo, M.; Han, K.; Dong, X.; Zhao, J.; Cai, W.; Zhang, Q.; Hua, H. Ultrabithorax is a key regulator for the dimorphism of wings, a main cause for the outbreak of planthoppers in rice. Nat. Sci. Rev. 2020, 7, 1181–1189. [Google Scholar] [CrossRef]
- Khush; Gurdev, S. Green revolution: The way forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef]
- Li, C.; Wu, D.; Huang, S.; Meng, M.; Shih, H.; Lai, M.; Chen, L.; Jena, K.K.; Hechanova, S.L.; Ke, T.; et al. The Bph45 gene confers resistance against brown planthopper in rice by reducing the production of limonene. Int. J. Mol. Sci. 2023, 24, 1798. [Google Scholar] [CrossRef]
- Du, B.; Zhang, W.; Liu, B.; Hu, J.; Wei, Z.; Shi, Z.; He, R.; Zhu, L.; Chen, R.; Han, B. Identification and characterization of Bph14, a gene conferring resistance to brown planthopper in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 22163–22168. [Google Scholar] [CrossRef] [PubMed]
- Fujita, D.; Kohli, A.; Horgan, F.G. Rice Resistance to Planthoppers and Leafhoppers. Crit. Rev. Plant Sci. 2013, 32, 162–191. [Google Scholar] [CrossRef]
- Hao, P.; Liu, C.; Wang, Y.; Chen, R.; Tang, M.; Du, B.; Zhu, L.; He, G. Herbivore-induced callose deposition on the sieve plates of rice: An important mechanism for host resistance. Plant Physiol. 2008, 146, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Liu, Y.; Liu, Y.; Jiang, L.; Wu, H.; Kang, H.; Liu, S.; Chen, L.; Liu, X.; Cheng, X.; et al. High-resolution mapping of brown planthopper (BPH) resistance gene Bph27(t) in rice (Oryza sativa L.). Mol. Breed. 2012, 31, 549–557. [Google Scholar] [CrossRef]
- Painter, R.H. Resistance of Plants to Insects. Annu. Rev. Entomol. 1958, 3, 267–290. [Google Scholar] [CrossRef]
- Alam, S.N.; Cohen, M.B. Detection and analysis of QTLs for resistance to the brown planthopper, Nilaparvata lugens, in a doubled-haploid rice population. Theor. Appl. Genet. 1998, 97, 1370–1379. [Google Scholar] [CrossRef]
- Pathak, M.D.; Cheng, C.H.; Fortuno, M.E. Resistance to Nephotettix impicticeps and Nilaparvata lugens in Varieties of Rice. Nature 1969, 223, 502–504. [Google Scholar] [CrossRef]
- Jackson, M.T.; Jackson, M.T. Conservation of rice genetic resources: The role of the International Rice Genebank at IRRI. Plant Mol. Biol. 1997, 35, 61–67. [Google Scholar] [CrossRef]
- Ling, Y.; Weilin, Z. Genetic and biochemical mechanisms of rice resistance to planthopper. Plant Cell Rep. 2016, 35, 1559–1572. [Google Scholar] [CrossRef]
- Ikeda, R.; Vaughan, D. The distribution of resistance genes to the brown planthopper in rice germplasm. RGN 1991, 8, 1–3. [Google Scholar]
- Wang, Y.; Cao, L.; Zhang, Y.; Cao, C.; Liu, F.; Huang, F.; Qiu, Y.; Li, R.; Lou, X. Map-based cloning and characterization of BPH29, a B3 domain-containing recessive gene conferring brown planthopper resistance in rice. J. Exp. Bot. 2015, 66, 6035–6045. [Google Scholar] [CrossRef] [PubMed]
- Athwal, D. Genetics of resistance to brown planthoppers and green leafhoppers in Oryza sativa L. Crop Sci. 1971, 11, 747–750. [Google Scholar] [CrossRef]
- Kim, S.M.; Sohn, J.K. Identification of a rice gene (Bph1) conferring resistance to brown planthopper (Nilaparvata lugens Stål) using STS markers. Mol. Cells 2005, 20, 30–34. [Google Scholar] [CrossRef]
- Sun, L.; Liu, Y.; Jiang, L.; Su, C.; Wang, C.; Zhai, H.; Wan, J. Identification of quantitative trait loci associated with resistance to brown planthopper in the indica rice cultivar Col.5 Thailand. Hereditas 2007, 144, 48–52. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Su, C.; Liu, Y.; Zhai, H.; Wan, J. Mapping and marker-assisted selection of a brown planthopper resistance gene bph2 in rice (Oryza sativa L.). Acta Genet. Sin. 2006, 33, 717–723. [Google Scholar] [CrossRef]
- Cha, Y.; Ji, H.; Yun, D.; Ahn, B.; Lee, M.C.; Suh, S.; Lee, C.S.; Ahn, E.K.; Jeon, Y.; Jin, I. Fine mapping of the rice Bph1 gene, which confers resistance to the brown planthopper (Nilaparvata lugens Stål), and development of STS markers for marker-assisted selection. Mol. Cells 2008, 26, 146–151. [Google Scholar] [CrossRef]
- Bhanu, K.V.; Lakshmi, V.J.; Katti, G.; Reddy, A.V. Antibiosis and tolerance mechanisms of resistance in rice varieties carrying brown planthopper resistance genes. Asian J. Biol. Life Sci. 2014, 3, 108–113. [Google Scholar]
- Liu, Y.; Wu, H.; Chen, H.; Liu, Y.; He, J.; Kang, H.; Sun, Z.; Pan, G.; Wang, Q.; Hu, J.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2014, 33, 301–305. [Google Scholar] [CrossRef]
- Jairin, J.; Sansen, K.; Wongboon, W.; Kothcharerk, J. Detection of a brown planthopper resistance gene bph4 at the same chromosomal position of Bph3 using two different genetic backgrounds of rice. Breed. Sci. 2010, 60, 71–75. [Google Scholar] [CrossRef]
- Sidhu, G.S.; Khush, G.S. Genetic analysis of brown planthopper resistance in twenty varieties of rice, Oryza saliva L. Theor. Appl. Genet. 1978, 53, 199–203. [Google Scholar] [CrossRef]
- Khush, G.S.; Karim, A.N.M.R.; Angeles, E.R. Genetics of resistance of rice cultivar ARC10550 to Bangladesh brown pianthopper teletype. J. Genet. 1985, 64, 121–125. [Google Scholar] [CrossRef]
- Guo, J.; Xu, C.; Wu, D.; Zhao, Y.; Qiu, Y.; Wang, X.; Ouyang, Y.; Cai, B.; Liu, X.; Jing, S.; et al. Bph6 encodes an exocyst-localized protein and confers broad resistance to planthoppers in rice. Nat. Genet. 2018, 50, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. High-resolution mapping of the brown planthopper resistance gene Bph6 in rice and characterizing its resistance in the 9311 and Nipponbare near isogenic backgrounds. Theor. Appl. Genet. 2010, 121, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. Fine mapping of the rice brown planthopper resistance gene BPH7 and characterization of its resistance in the 93-11 background. Euphytica 2014, 198, 369–379. [Google Scholar] [CrossRef]
- Nemoto, H.; Ikeda, R.; Kaneda, C. New genes for resistance to brown planthopper, Nilaparvata lugens Stål, in Rice. Jpn. J. Breed. 1989, 39, 23–28. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, J.; Wang, Z.; Jing, S.; Wang, Y.; Ouyang, Y.; Cai, B.; Xin, X.-F.; Liu, X.; Zhang, C.; et al. Allelic diversity in an NLR gene BPH9 enables rice to combat planthopper variation. Proc. Natl. Acad. Sci. USA 2016, 113, 12850–12855. [Google Scholar] [CrossRef]
- Su, C.; Zhai, H.; Wang, C.; Sun, L.; Wan, J. SSR mapping of brown planthopper resistance gene Bph9 in kaharamana, an indica rice (Oryza sativa L.). Acta Genet. Sin. 2006, 33, 262–268. [Google Scholar] [CrossRef]
- Ishii, T.; Brar, D.S.; Multani, D.S.; Khush, G.S. Molecular tagging of genes for brown planthopper resistance and earliness introgressed from Oryza australiensis into cultivated rice, O. sativa. Genome 1994, 37, 217–221. [Google Scholar] [CrossRef]
- Hirabayashi, H. Identification of brown planthopper resistance gene derived from O. officinalis using molecular markers in rice. Breed. Sci. 1998, 48, 82. [Google Scholar]
- Qiu, Y.; Guo, J.; Jing, S.; Zhu, L.; He, G. Development and characterization of japonica rice lines carrying the brown planthopper-resistance genes BPH12 and BPH6. Theor. Appl. Genet. 2011, 124, 485–494. [Google Scholar] [CrossRef]
- Yang, H.; Ren, X.; Weng, Q.; Zhu, L.; He, G. Molecular mapping and genetic analysis of a rice brown planthopper (Nilaparvata lugens Stål) resistance gene. Hereditas 2002, 136, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yan, H.; Fu, Q.; Qian, Q.; Zhang, Z.; Zhai, W.; Zhu, L. Mapping of a new gene for brown planthopper resistance in cultivated rice introgressed from Oryza eichingeri. Chin. Sci. Bull. 2001, 46, 1459–1462. [Google Scholar] [CrossRef]
- Renganayaki, K.; Fritz, A.K.; Sadasivam, S.; Pammi, S.; Harrington, S.E.; McCouch, S.R.; Kumar, S.M.; Reddy, A.S. Mapping and progress toward map-based cloning of brown planthopper biotype-4 resistance gene introgressed from Oryza officinalis into cultivated rice, O. sativa. Crop Sci. 2002, 42, 2112–2117. [Google Scholar] [CrossRef]
- Yang, H.; You, A.; Yang, Z.; Zhang, F.; He, R.; Zhu, L.; He, G. High-resolution genetic mapping at the Bph15 locus for brown planthopper resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2004, 110, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wu, Y.; Guo, J.; Du, B.; Chen, R.; Zhu, L.; He, G. A rice lectin receptor-like kinase that is involved in innate immune responses also contributes to seed germination. Plant J. 2013, 76, 687–698. [Google Scholar] [CrossRef]
- Sun, L.; Su, C.; Wang, C.; Zhai, H.; Wan, J. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed. Sci. 2005, 55, 391–396. [Google Scholar] [CrossRef]
- Ji, H.; Kim, S.; Kim, Y.-H.; Suh, J.-P.; Park, H.-M.; Sreenivasulu, N.; Misra, G.; Kim, S.-M.; Hechanova, S.L.; Kim, H.; et al. Map-based cloning and characterization of the BPH18 Gene from wild rice conferring resistance to brown planthopper (BPH) insect pest. Sci. Rep. 2016, 6, 34376. [Google Scholar] [CrossRef]
- Li, R.; Li, L.; Wei, S.; Wei, Y.; Chen, Y.; Bai, D.; Yang, L.; Huang, F.; Lu, W.; Zhang, X.; et al. The evaluation and utilization of new genes for brown planthopper resistance in common wild rice (Oryza rufipogon Griff.). Mol. Entomol. 2010, 1, 365–371. [Google Scholar]
- Chen, J.W.; Wang, L.; Pang, X.F.; Pan, Q.H. Genetic analysis and fine mapping of a rice brown planthopper (Nilaparvata lugens Stål) resistance gene bph19(t). Mol. Genet. Genom. 2006, 275, 321–329. [Google Scholar] [CrossRef]
- Rahman, M.L.; Jiang, W.; Chu, S.H.; Qiao, Y.; Ham, T.; Woo, M.; Lee, J.; Khanam, M.S.; Chin, J.; Jeung, J.; et al. High-resolution mapping of two rice brown planthopper resistance genes, Bph20(t) and Bph21(t), originating from Oryza minuta. Theor. Appl. Genet. 2009, 119, 1237–1246. [Google Scholar] [CrossRef]
- Yang, L.; Li, R.B.; Li, Y.R.; Huang, F.K.; Chen, Y.Z.; Huang, S.S.; Huang, L.F.; Liu, C.; Ma, Z.F.; Huang, D.H.; et al. Genetic mapping of bph20(t) and bph21(t) loci conferring brown planthopper resistance to Nilaparvata lugens Stål in rice (Oryza sativa L.). Euphytica 2011, 183, 161–171. [Google Scholar] [CrossRef]
- Ram, T.; Deen, R.; Gautam, S.; Ramesh, K.; Rao, Y.; Brar, D. Identification of new genes for brown planthopper resistance in rice introgressed from O. glaberrima and O. minuta. Rice Genet. Newsl. 2010, 25, 67–69. [Google Scholar]
- Hou, L.; Yu, P.; Xu, Q.; Yuan, X.; Yu, H.; Wang, Y.; Wang, C.; Wan, G.; Tang, S.; Peng, S.; et al. Genetic analysis and preliminary mapping of two recessive resistance genes to brown planthopper, Nilaparvata lugens Stål in rice. Rice Sci. 2011, 18, 238–242. [Google Scholar] [CrossRef]
- Deen, R.; Ramesh, K.; Gautam, S.; Rao, Y.; Lakshmi, V.; Viraktamath, B.; Brar, D.; Ram, T. Identification of new gene for BPH resistance introgressed from O. rufipogon. Rice Genet. Newsl. 2010, 25, 70–71. [Google Scholar]
- Myint, K.K.M.; Fujita, D.; Matsumura, M.; Sonoda, T.; Yoshimura, A.; Yasui, H. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens Stål) in the rice cultivar ADR52. Theor. Appl. Genet. 2011, 124, 495–504. [Google Scholar] [CrossRef]
- Tamura, Y.; Hattori, M.; Yoshioka, H.; Yoshioka, M.; Takahashi, A.; Wu, J.; Sentoku, N.; Yasui, H. Map-based cloning and characterization of a brown planthopper resistance gene BPH26 from Oryza sativa L. ssp. indica Cultivar ADR52. Sci. Rep. 2014, 4, 5872. [Google Scholar]
- Huang, D.; Qiu, Y.; Zhang, Y.; Huang, F.; Meng, J.; Wei, S.; Li, R.; Chen, B. Fine mapping and characterization of BPH27, a brown planthopper resistance gene from wild rice (Oryza rufipogon Griff.). Theor. Appl. Genet. 2013, 126, 219–229. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Y.; He, J.; Liu, Y.; Jiang, L.; Liu, L.; Wang, C.; Cheng, X.; Wan, J. Fine mapping of brown planthopper (Nilaparvata lugens Stål) resistance gene Bph28(t) in rice (Oryza sativa L.). Mol. Breed. 2014, 33, 909–918. [Google Scholar] [CrossRef]
- Shi, S.; Wang, H.; Nie, L.; Tan, D.; Zhou, C.; Zhang, Q.; Li, Y.; Du, B.; Guo, J.; Huang, J.; et al. Bph30 confers resistance to brown planthopper by fortifying sclerenchyma in rice leaf sheaths. Mol. Plant 2021, 14, 1714–1732. [Google Scholar] [CrossRef]
- Prahalada, G.D.; Shivakumar, N.; Lohithaswa, H.C.; Sidde Gowda, D.K.; Ramkumar, G.; Kim, S.-R.; Ramachandra, C.; Hittalmani, S.; Mohapatra, T.; Jena, K.K. Identification and fine mapping of a new gene, BPH31 conferring resistance to brown planthopper biotype 4 of India to improve rice, Oryza sativa L. Rice 2017, 10, 41. [Google Scholar] [CrossRef]
- Ren, J.; Gao, F.; Wu, X.; Lu, X.; Zeng, L.; Lv, J.; Su, X.; Luo, H.; Ren, G. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 2016, 6, 37645. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chang, X.; Zou, L.; Tang, W.; Wu, W. Identification and fine mapping of Bph33, a new brown planthopper resistance gene in rice (Oryza sativa L.). Rice 2018, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.B.; Divya, D.; Sahu, N.; Sundaram, R.M.; Sarao, P.S.; Singh, K.; Lakshmi, V.J.; Bentur, J.S. A new gene Bph33(t) conferring resistance to brown planthopper (BPH), Nilaparvata lugens (Stål) in rice line RP2068-18-3-5. Euphytica 2018, 214, 53. [Google Scholar] [CrossRef]
- Kumar, K.; Sarao, P.S.; Bhatia, D.; Neelam, K.; Kaur, A.; Mangat, G.S.; Brar, D.S.; Singh, K. High-resolution genetic mapping of a novel brown planthopper resistance locus, Bph34 in Oryza sativa L. X Oryza nivara (Sharma & Shastry) derived interspecific F2 population. Theor. Appl. Genet. 2018, 131, 1163–1171. [Google Scholar]
- Zhang, Y.; Qin, G.; Ma, Q.; Wei, M.; Tang, X.; Ma, Z.; Liang, H.; Liu, C.; Li, Z.; Liu, F.; et al. Identification of major locus Bph35 resistance to brown planthopper in rice. Rice Sci. 2020, 27, 237–245. [Google Scholar]
- Li, Z.; Xue, Y.; Zhou, H.; Li, Y.; Usman, B.; Jiao, X.; Wang, X.; Liu, F.; Qin, B.; Li, R.; et al. High-resolution mapping and breeding application of a novel brown planthopper resistance gene derived from wild rice (Oryza. rufipogon Griff). Rice 2019, 12, 1–13. [Google Scholar] [CrossRef]
- Yang, M.; Cheng, L.; Yan, L.; Shu, W.; Wang, X.; Qiu, Y. Mapping and characterization of a quantitative trait locus resistance to the brown planthopper in the rice variety IR64. Hereditas 2019, 156, 22. [Google Scholar] [CrossRef]
- Zhou, C.; Zhang, Q.; Chen, Y.; Huang, J.; Guo, Q.; Li, Y.; Wang, W.; Qiu, Y.; Guan, W.; Zhang, J.; et al. Balancing selection and wild gene pool contribute to resistance in global rice germplasm against planthopper. J. Integr. Plant Biol. 2021, 63, 1695–1711. [Google Scholar] [CrossRef]
- Yang, M.; Lin, J.; Cheng, L.; Zhou, H.; Chen, S.; Liu, F.; Li, R.; Qiu, Y. Identification of a novel planthopper resistance gene from wild rice (Oryza rufipogon Griff.). Crop J. 2020, 8, 1057–1070. [Google Scholar] [CrossRef]
- Balachiranjeevi, C.H.; Prahalada, G.D.; Mahender, A.; Jamaloddin, M.; Sevilla, M.A.L.; Marfori-Nazarea, C.M.; Vinarao, R.; Sushanto, U.; Baehaki, S.E.; Li, Z.K.; et al. Identification of a novel locus, BPH38(t), conferring resistance to brown planthopper (Nilaparvata lugens Stål.) using early backcross population in rice (Oryza sativa L.). Euphytica 2019, 215, 185. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, Y.; Zou, L.; Wu, X.; Zhang, F.; Chen, C.; Xiong, S.; Liang, B.; Zhu, Z.; Wu, W.; et al. Identification and candidate analysis of a new brown planthopper resistance locus in an Indian landrace of rice, paedai kalibungga. Mol. Breed. 2024, 44, 45. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Pusuluri, M.; Balakrishnan, D.; Vattikuti, J.L.; Neelamraju, S.; Sundaram, R.M.; Mangrauthia, S.K.; Ram, T. Identification and functional characterization of two major loci associated with resistance against brown planthoppers (Nilaparvata lugens Stål) derived from Oryza nivara. Genes 2023, 14, 2066. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Han, Y.; Zhang, Y.; Deng, B.; Wu, B.; Guo, X.; Qin, Y.; Fang, Y.; Liu, F.; Qin, B.; et al. QTL mapping integrated with BSA-Seq analysis identifies a novel gene conferring resistance to brown planthopper from common wild rice (Oryza rufipogon Griff.). Euphytica 2022, 218, 34–55. [Google Scholar] [CrossRef]
- Tan, H.Q.; Palyam, S.; Gouda, J.; Kumar, P.P.; Chellian, S.K. Identification of two QTLs, BPH41 and BPH42, and their respective gene candidates for brown planthopper resistance in rice. Sci. Rep. 2022, 12, 18538–18550. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Neelam, K.; Sarao, P.S.; Babbar, A.; Kumar, K.; Vikal, Y.; Khanna, R.; Kaur, R.; Mangat, G.S.; Singh, K. Molecular mapping and transfer of a novel brown planthopper resistance gene bph42 from Oryza rufipogon (Griff.) to cultivated rice (Oryza sativa L.). Mol. Biol. Rep. 2022, 49, 8597–8606. [Google Scholar] [CrossRef]
- Kim, J.; An, X.; Yang, K.; Miao, S.; Qin, Y.; Hu, Y.; Du, B.; Zhu, L.; He, G.; Chen, R. Molecular mapping of a new brown planthopper resistance gene Bph43 in rice (Oryza sativa L.). Agronomy 2022, 12, 808. [Google Scholar] [CrossRef]
- Kiswanto, I.; Soetopo, L.; Adiredjo, A.L. Identification of novel candidate of brown planthopper resistance gene Bph44 in rice (Oryza sativa L.). Genome 2022, 65, 505–511. [Google Scholar] [CrossRef]
- Li, F.; Yan, L.; Shen, J.; Liao, S.; Ren, X.; Cheng, L.; Li, Y.; Qiu, Y. Fine mapping and breeding application of two brown planthopper resistance genes derived from landrace rice. PLoS ONE 2024, 19, e0297945. [Google Scholar] [CrossRef]
- Hu, J.; Xiao, C.; Cheng, M.; Gao, G.; Zhang, Q.; He, Y. Fine mapping and pyramiding of brown planthopper resistance genes QBph3 and QBph4 in an introgression line from wild rice O. officinalis. Mol. Breed. 2015, 35, 3–12. [Google Scholar] [CrossRef]
- Van Mai, T.; Fujita, D.; Matsumura, M.; Yoshimura, A.; Yasui, H. Genetic basis of multiple resistance to the brown planthopper (Nilaparvata lugens Stål) and the green rice leafhopper (Nephotettix cincticeps Uhler) in the rice cultivar ‘ASD7’(Oryza sativa L. ssp. indica). Breed. Sci. 2015, 65, 420–429. [Google Scholar] [CrossRef]
- Qiu, Y.; Cheng, L.; Liu, F.; Li, R.B. Identification of a new locus conferring antixenosis to the brown planthopper in rice cultivar Swarnalata (Oryza sativa L.). Genet. Mol. Res. 2013, 12, 3201–3211. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Wan, J.; Zhai, H.; Wang, C.; Sun, L.; Yasui, H.; Yoshimura, A. A new locus for resistance to brown planthopper identified in the indica rice variety DV85. Plant Breed. 2005, 124, 93–95. [Google Scholar] [CrossRef]
- Kamolsukyeunyong, W.; Ruengphayak, S.; Chumwong, P.; Kusumawati, L.; Chaichoompu, E.; Jamboonsri, W.; Saensuk, C.; Phoonsiri, K.; Toojinda, T.; Vanavichit, A. Identification of spontaneous mutation for broad-spectrum brown planthopper resistance in a large, long-term fast neutron mutagenized rice population. Rice 2019, 12, 16–41. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Xiao, C.; Cheng, M.; Gao, G.; Zhang, Q.; He, Y. A new finely mapped Oryza australiensis-derived QTL in rice confers resistance to brown planthopper. Gene 2015, 561, 132–137. [Google Scholar] [CrossRef]
- Mohanty, S.K.; Panda, R.S.; Mohapatra, S.L.; Nanda, A.; Behera, L.; Jena, M.; Sahu, R.K.; Sahu, S.C.; Mohapatra, T. Identification of novel quantitative trait loci associated with brown planthopper resistance in the rice landrace Salkathi. Euphytica 2017, 213, 38–52. [Google Scholar] [CrossRef]
- Hu, L.; Wu, Y.; Wu, D.; Rao, W.; Guo, J.; Ma, Y.; Wang, Z.; Shangguan, X.; Wang, H.; Xu, C.; et al. The coiled-coil and nucleotide binding domains of BROWN PLANTHOPPER RESISTANCE14 function in signaling and resistance against planthopper in rice. Plant Cell 2017, 29, 3157–3185. [Google Scholar] [CrossRef]
- Guo, J.; Wang, H.; Guan, W.; Guo, Q.; Wang, J.; Yang, J.; Peng, Y.; Shan, J.; Gao, M.; Shi, S.; et al. A tripartite rheostat controls self-regulated host plant resistance to insects. Nature 2023, 618, 799–807. [Google Scholar] [CrossRef]
- Hatchett, J.H.; Gallun, R.L. Genetics of the ability of the hessian fly, mayetiola destructor, to survive on wheats having different genes for resistance. Ann. Entomol. Soc. Am. 1970, 63, 1400–1407. [Google Scholar] [CrossRef]
- Laksminarayana, A.; Khush, G.S. New genes for resistance to the brown planthopper in rice. Crop Sci. 1977, 17, 96–100. [Google Scholar] [CrossRef]
- Cheng, X.; Zhu, L.; He, G. Towards understanding of molecular interactions between rice and the brown planthopper. Mol. Plant 2013, 6, 621–634. [Google Scholar] [CrossRef]
- Jairin, J.; Phengrat, K.; Teangdeerith, S.; Vanavichit, A.; Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breed. 2007, 19, 35–44. [Google Scholar] [CrossRef]
- Wu, D.; Guo, J.; Zhang, Q.; Shi, S.; Guan, W.; Zhou, C.; Chen, R.; Du, B.; Zhu, L.; He, G. Necessity of rice resistance to planthoppers for OsEXO70H3 regulating SAMSL excretion and lignin deposition in cell walls. New Phytol. 2022, 234, 1031–1046. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, X.; Wang, Q.; Huangfu, J.; Schuman, M.C.; Lou, Y. A group D MAPK protects plants from autotoxicity by suppressing herbivore-induced defense signaling. Plant Physiol. 2019, 179, 1386–1401. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Yu, H.; Jian, Y.; Zeng, J.; Ji, R.; Chen, H.; Lou, Y. A salivary EF-hand calcium-binding protein of the brown planthopper Nilaparvata lugens functions as an effector for defense responses in rice. Sci. Rep. 2017, 7, 40498–40509. [Google Scholar] [CrossRef]
- Dai, Z.; Tan, J.; Zhou, C.; Yang, X.; Yang, F.; Zhang, S.; Sun, S.; Miao, X.; Shi, Z. The OsmiR396-OsGRF8-OsF3H-flavonoid pathway mediates resistance to the brown planthopper in rice (Oryza sativa). Plant Biotechnol. J. 2019, 17, 1657–1669. [Google Scholar] [CrossRef]
- Liu, M.; Hong, G.; Li, H.; Bing, X.; Chen, Y.; Jing, X.; Gershenzon, J.; Lou, Y.; Baldwin, I.T.; Li, R. Sakuranetin protects rice from brown planthopper attack by depleting its beneficial endosymbionts. Proc. Natl. Acad. Sci. USA 2023, 120, e2305007120. [Google Scholar] [CrossRef]
- Sun, B.; Shen, Y.; Zhu, L.; Yang, X.; Liu, X.; Li, D.; Zhu, M.; Miao, X.; Shi, Z. OsmiR319-OsPCF5 modulate resistance to brown planthopper in rice through association with MYB proteins. BMC Biol. 2024, 22, 68. [Google Scholar] [CrossRef]
- He, J.; Liu, Y.; Yuan, D.; Duan, M.; Liu, Y.; Shen, Z.; Yang, C.; Qiu, Z.; Liu, D.; Wen, P.; et al. An R2R3 MYB transcription factor confers brown planthopper resistance by regulating the phenylalanine ammonia-lyase pathway in rice. Proc. Natl. Acad. Sci. USA 2019, 117, 271–277. [Google Scholar] [CrossRef]
- Li, S.; Tan, X.; He, Z.; Jiang, L.; Li, Y.; Yang, L.; Hoffmann, A.A.; Zhao, C.; Fang, J.; Ji, R. Transcriptome-wide N6-methyladenosine profiling reveals growth-defense trade-offs in the response of rice to brown planthopper (Nilaparvata lugens) infestation. Pest. Manag. Sci. 2024, 80, 5364–5376. [Google Scholar] [CrossRef]
- Tao, Z.; Zhu, L.; Li, H.; Sun, B.; Liu, X.; Li, D.; Hu, W.; Wang, S.; Miao, X.; Shi, Z. ACL1-ROC4/5 complex reveals a common mechanism in rice response to brown planthopper infestation and drought. Nat. Commun. 2024, 15, 8107–8123. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, X.; Li, S.; Xie, Y.; Luo, X.; Yang, Y.; Pu, Z.; Zhang, L.; Lu, J.; Huang, H.; et al. Rapid intracellular acidification is a plant defense response countered by the brown planthopper. Curr. Biol. 2024, 34, 5017–5027.e4. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.L.; Li, W.; Wang, L.N.; Wang, S.C.; Falert, S.; Wang, C.; Yu, S.Y.; Abdelkhalek, S.T.; Lu, J.; Lin, Y.J.; et al. Limonene enhances rice plant resistance to a piercing-sucking herbivore and rice pathogens. Plant Biotechnol. J. 2024, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Liu, Y.; Ji, L.; Zhang, X.; He, J.; Huang, J.; Qiu, Z.; Liu, D.; Sun, Z.; Xu, T.; et al. Brassinosteroids mediate susceptibility to brown planthopper by integrating with the salicylic acid and jasmonic acid pathways in rice. J. Exp. Bot. 2018, 69, 4433–4442. [Google Scholar] [CrossRef] [PubMed]
- Huangfu, J.; Li, J.; Li, R.; Ye, M.; Kuai, P.; Zhang, T.; Lou, Y. The transcription factor OsWRKY45 negatively modulates the resistance of rice to the brown planthopper Nilaparvata lugens. Int. J. Mol. Sci. 2016, 17, 697. [Google Scholar] [CrossRef]
- Bürger, M.; Chory, J. Stressed out about hormones: How plants orchestrate immunity. Cell Host Microbe 2019, 26, 163–172. [Google Scholar] [CrossRef]
- Mishra, A.; Barik, S.R.; Pandit, E.; Yadav, S.S.; Das, S.R.; Pradhan, S.K. Genetics, mechanisms and deployment of brown planthopper resistance genes in rice. Crit. Rev. Plant Sci. 2022, 41, 91–127. [Google Scholar] [CrossRef]
- Zhou, G.; Qi, J.; Ren, N.; Cheng, J.; Erb, M.; Mao, B.; Lou, Y. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J. 2009, 60, 638–648. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Zu, H.; Zeng, X.; Baldwin, I.T.; Lou, Y.; Li, R. Molecular dissection of rice phytohormone signaling involved in resistance to a piercing-sucking herbivore. New Phytol. 2021, 230, 1639–1652. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Liu, S.; Fu, W.; Zhuang, Y.; Xu, J.; Lou, Y.; Baldwin, I.T.; Li, R. Functional dissection of rice jasmonate receptors involved in development and defense. New Phytol. 2023, 238, 2144–2158. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, D.; Gao, D.; Zhao, W.; Du, H.; Qiu, Z.; Huang, J.; Wen, P.; Wang, Y.; Li, Q.; et al. Cytokinin confers brown planthopper resistance by elevating jasmonic acid pathway in rice. Int. J. Mol. Sci. 2022, 23, 5946. [Google Scholar] [CrossRef]
- Li, M.; Yu, G.; Cao, C.; Liu, P. Metabolism, signaling, and transport of jasmonates. Plant Commun. 2021, 2, 100231. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Jin, G.; Jiménez-Alemán, G.H.; Wang, X.; Song, J.; Li, S.; Lou, Y.; Li, R. The jasmonic acid-amino acid conjugates JA-Val and JA-Leu are involved in rice resistance to herbivores. Plant Cell Environ. 2022, 45, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.S.; Liu, D.; Guo, W.; Liu, Z.X.; Zhang, X.; Shi, L.L.; Zhou, D.M.; Wang, L.N.; Kang, K.; Wang, F.Z.; et al. Poaceae-specific β-1,3;1,4-d-glucans link jasmonate signalling to OsLecRK1-mediated defence response during rice-brown planthopper interactions. Plant Biotechnol. J. 2023, 21, 1286–1300. [Google Scholar] [CrossRef]
- Lu, J.; Li, J.; Ju, H.; Liu, X.; Erb, M.; Wang, X.; Lou, Y. Contrasting effects of ethylene biosynthesis on induced plant resistance against a chewing and a piercing-sucking herbivore in rice. Mol. Plant 2014, 7, 1670–1682. [Google Scholar] [CrossRef]
- Ma, F.; Yang, X.; Shi, Z.; Miao, X. Novel crosstalk between ethylene- and jasmonic acid-pathway, 45, responses to a piercing-sucking insect in rice. New Phytol. 2019, 225, 474–487. [Google Scholar] [CrossRef]
- Huang, J.; Qiu, Z.; He, J.; Xu, H.; Wang, K.; Du, H.; Gao, D.; Zhao, W.; Sun, Q.; Wang, Y.; et al. Phytochrome B mediates dim-light-reduced insect resistance by promoting the ethylene pathway in rice. Plant Physiol. 2023, 191, 1272–1287. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, L.; Wang, S.; Xie, P.; Liu, J. A key ABA hydrolase gene, OsABA8ox3 is involved in rice resistance to Nilaparvata lugens by affecting callose deposition. J. Asia-Pac. Entomol. 2019, 22, 625–631. [Google Scholar] [CrossRef]
- Li, J.; Liu, H.; Lv, X.; Wang, W.; Liang, X.; Chen, L.; Wang, Y.; Liu, J. A key ABA biosynthetic gene OsNCED3 is a positive regulator in resistance to Nilaparvata lugens in Oryza sativa. Front. Plant Sci. 2024, 15, 1359315–1359327. [Google Scholar] [CrossRef]
- Liu, J.; Du, H.; Ding, X.; Zhou, Y.; Xie, P.; Wu, J. Mechanisms of callose deposition in rice regulated by exogenous abscisic acid and its involvement in rice resistance to Nilaparvata lugens Stål (Hemiptera: Delphacidae). Pest. Manag. Sci. 2017, 73, 2559–2568. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Li, J.; Zhou, G.; Wang, Q.; Bian, W.; Erb, M.; Lou, Y. Prioritizing plant defence over growth through WRKY regulation facilitates infestation by non-target herbivores. eLife 2015, 4, e04805. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, T.; Wang, W.; Cao, T.; Li, R.; Lou, Y. Silencing OsSLR1 enhances the resistance of rice to the brown planthopper Nilaparvata lugens. Plant Cell Environ. 2017, 40, 2147–2159. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cao, T.; Zhang, J.; Lou, Y. Overexpression of OsGID1 enhances the resistance of rice to the brown planthopper Nilaparvata lugens. Int. J. Mol. Sci. 2018, 19, 2744. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Jin, N.; Mo, X.; Wu, J.; Lu, J.; Lou, Y. Exogenous gibberellin GA3 enhances defense responses in rice to the brown planthopper Nilaparvata lugens (Stål). J. Plant Biol. 2020, 64, 379–387. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, T.; Gao, M.; Ye, M.; Lin, M.; Wu, D.; Guo, J.; Guan, W.; Wang, J.; Yang, K.; et al. Transcriptome and metabolome profiling reveal the resistance mechanisms of rice against brown planthopper. Int. J. Mol. Sci. 2022, 23, 4083. [Google Scholar] [CrossRef]
- Shi, S.; Zha, W.; Yu, X.; Wu, Y.; Li, S.; Xu, H.; Li, P.; Li, C.; Liu, K.; Chen, J.; et al. Integrated transcriptomics and metabolomics analysis provide insight into the resistance response of rice against brown planthopper. Front. Plant Sci. 2023, 14, 1213257. [Google Scholar] [CrossRef]
- Huang, S.; Jia, A.; Ma, S.; Sun, Y.; Chang, X.; Han, Z.; Chai, J. NLR signaling in plants: From resistosomes to second messengers. Trends Biochem. Sci. 2023, 48, 776–787. [Google Scholar] [CrossRef]
- Kuai, P.; Lin, N.; Ye, M.; Ye, M.; Chen, L.; Chen, S.; Zu, H.; Hu, L.; Gatehouse, A.M.R.; Lou, Y. Identification and knockout of a herbivore susceptibility gene enhances planthopper resistance and increases rice yield. Nat. Food 2024, 5, 846–859. [Google Scholar] [CrossRef]
- McDowell, J.M.; Argueso, C.T.; Ferreira, F.J.; Epple, P.; To, J.P.C.; Hutchison, C.E.; Schaller, G.E.; Dangl, J.L.; Kieber, J.J. Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 2012, 8, e1002448. [Google Scholar]
- Cao, M.; Chen, R.; Li, P.; Yu, Y.; Zheng, R.; Ge, D.; Zheng, W.; Wang, X.; Gu, Y.; Gelová, Z.; et al. TMK1-mediated auxin signalling regulates differential growth of the apical hook. Nature 2019, 568, 240–243. [Google Scholar] [CrossRef]
- Gao, J.; Chen, H.; Yang, H.; He, Y.; Tian, Z.; Li, J. A brassinosteroid responsive miRNA-target module regulates gibberellin biosynthesis and plant development. New Phytol. 2018, 220, 488–501. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, G.; Miao, X.; Shi, Z. OsmiR159 modulate bph resistance through regulating g-protein γ subunit GS3 gene in rice. Rice 2023, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Gheysen, G.; Yang, M.; Xiao, X.; Xu, L.; Guo, X.; Yang, L.; Liu, W.; He, Y.; Peng, D.; et al. Brown planthopper infestation on rice reduces plant susceptibility to Meloidogyne graminicola by reducing root sugar allocation. New Phytol. 2024, 242, 262–277. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Durán, R.; Zipfel, C. Trade-off between growth and immunity: Role of brassinosteroids. Trends Plant Sci. 2015, 20, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Qi, J.; Zu, H.; Liu, S.; Gershenzon, J.; Lou, Y.; Baldwin, I.T.; Li, R. Jasmonate-mediated gibberellin catabolism constrains growth during herbivore attack in rice. Plant Cell 2023, 35, 3828–3844. [Google Scholar] [CrossRef]
- Ning, Y.; Liu, W.; Wang, G.-L. Balancing immunity and yield in crop plants. Trends Plant Sci. 2017, 22, 1069–1079. [Google Scholar] [CrossRef]
- Nomoto, M.; Skelly, M.J.; Itaya, T.; Mori, T.; Suzuki, T.; Matsushita, T.; Tokizawa, M.; Kuwata, K.; Mori, H.; Yamamoto, Y.Y.; et al. Suppression of MYC transcription activators by the immune cofactor NPR1 fine-tunes plant immune responses. Cell Rep. 2021, 37, 110125. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, L.; Liu, Y.; Dai, H.; He, J.; Kang, H.; Pan, G.; Huang, J.; Qiu, Z.; Wang, Q.; et al. Marker assisted pyramiding of two brown planthopper resistance genes, Bph3 and Bph27(t), into elite rice Cultivars. Rice 2016, 9, 1–7. [Google Scholar] [CrossRef]
- Liu, M.; Fan, F.; He, S.; Guo, Y.; Chen, G.; Li, N.; Li, N.; Yuan, H.; Si, F.; Yang, F.; et al. Creation of elite rice with high-yield, superior-quality and high resistance to brown planthopper based on molecular design. Rice 2022, 15, 17. [Google Scholar] [CrossRef]
- van Schie, C.C.; Takken, F.L. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Gao, M.; He, Y.; Yin, X.; Zhong, X.; Yan, B.; Wu, Y.; Chen, J.; Li, X.; Zhai, K.; Huang, Y.; et al. Ca2+ sensor-mediated ROS scavenging suppresses rice immunity and is exploited by a fungal effector. Cell 2021, 184, 5391–5404. [Google Scholar] [CrossRef]
- Li, S.; Lin, D.; Zhang, Y.; Deng, M.; Chen, Y.; Lv, B.; Li, B.; Lei, Y.; Wang, Y.; Zhao, L.; et al. Genome-edited powdery mildew resistance in wheat without growth penalties. Nature 2022, 602, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, Y.; Zeng, X.; Lou, Y.; Baldwin, I.T.; Li, R. Brown planthoppers manipulate rice sugar transporters to benefit their own feeding. Curr. Biol. 2024, 34, 2990–2996. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.C.; Felton, G.W.; Tumlinson, J.H. The dual function of elicitors and effectors from insects: Reviewing the ‘arms race’ against plant defenses. Plant Mol. Biol. 2022, 109, 427–445. [Google Scholar] [CrossRef] [PubMed]
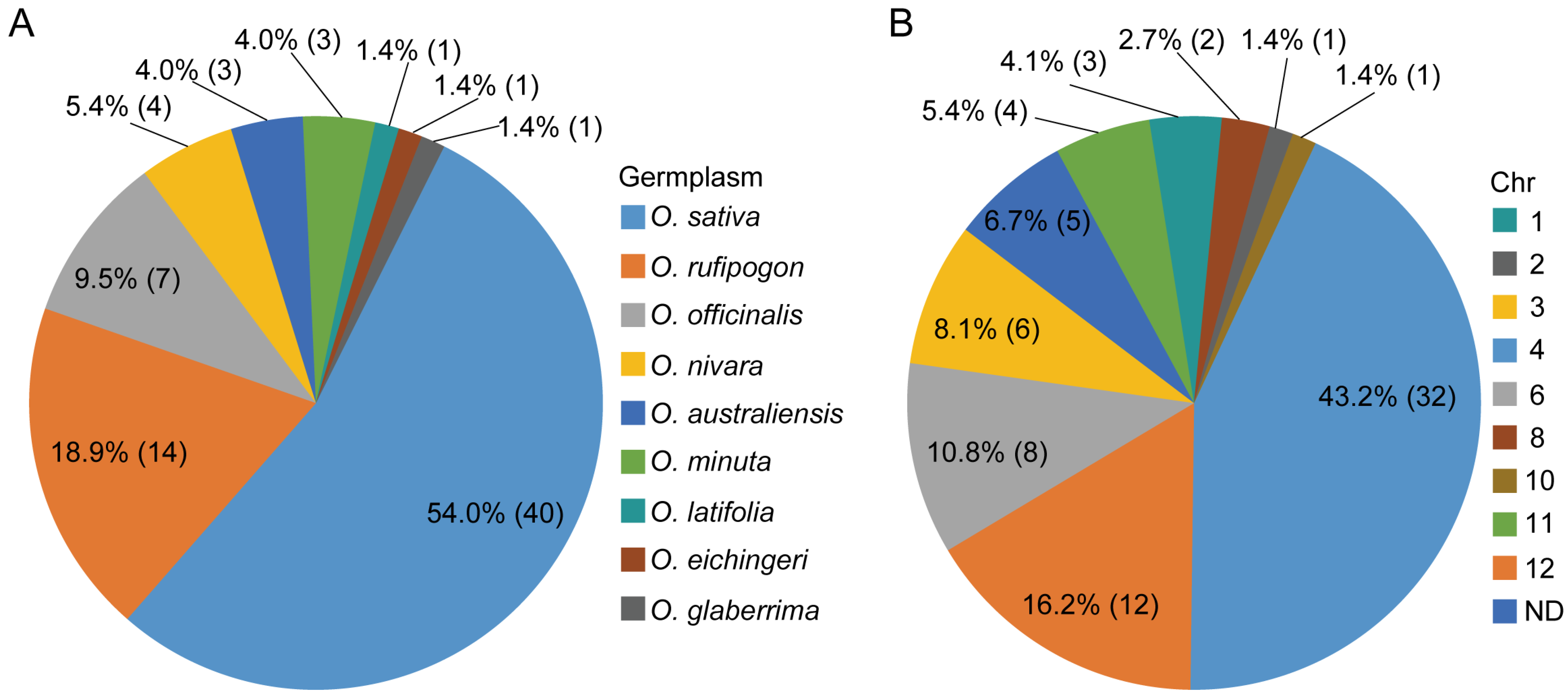
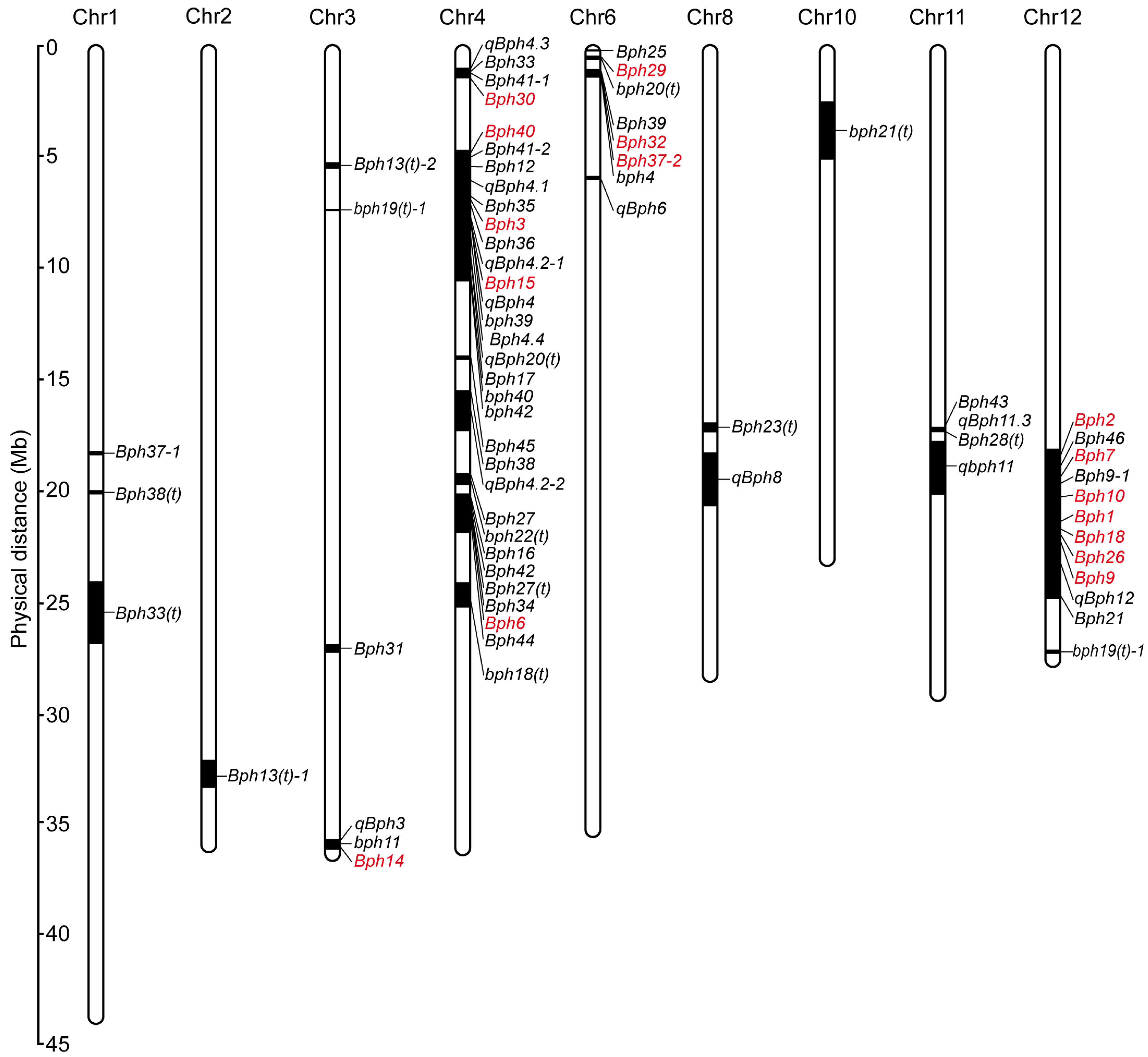


{kind=link}
{kind=link}
{kind=link}
| Genes/QTLs | Chr. | Germplasm | Linked Markers | Position (Mb) | Resistance Mechanism | Resistance to Biotype | Reference |
|---|---|---|---|---|---|---|---|
| Bph1* | 12L | Mudgo | pBPH4, pBPH14 | 22.86 | Antibiosis, antixenosis | 1, 3 | [22,26] |
| Bph2* | 12L | ASD7 | RM463, RM7102 | 13.21–22.13 | Antibiosis, tolerance | 1, 2 | [25,27] |
| Bph3* | 4S | Rathu Heenati | RHD9, RHC10 | 6.20–6.97 | Antibiosis, antixenosis | 1, 2, 3, 4 | [28] |
| bph4 | 6S | Babawee | RM589, RM586 | 1.38–1.47 | Antibiosis, antixenosis | 1, 2, 3, 4 | [29,30] |
| bph5 | ND | ARC10550 | ND | ND | ND | 4 | [31] |
| Bph6* | 4L | Swarnalata | Y19, Y9 | 21.36–21.39 | Antibiosis, antixenosis | 1, 2, 3, 4 | [32,33] |
| Bph7* | 12L | T12 | RM3448, RM313 | 19.95–20.87 | Antibiosis, tolerance | 4 | [27,34] |
| bph8 | ND | Chin Saba | ND | ND | ND | 1, 2, 3 | [35] |
| Bph9* | 12L | Pokkali | InD2, RsaI | 22.85–22.97 | Antibiosis, antixenosis | 1, 2, 3 | [36] |
| Bph9-1 | 12L | Kaharamana | RM463, RM5341 | 19.16–22.13 | ND | 1, 2, 3 | [37] |
| Bph10* | 12L | IR65482-4-136-2-2 (O. australiensis) | RG457 | 19.55–26.98 | ND | 1 | [36,38] |
| bph11 | 3L | O. officinalis | G1318 | 35.60–35.80 | ND | 1, 2 | [39] |
| Bph12 | 4S | B14 (O. latifolia) | RM16459, RM1305 | 5.21–5.56 | Antibiosis, antixenosis | 1, 2 | [40,41] |
| Bph13(t)-1 | 2L | O. eichingeri | RM240, RM250 | 31.50–32.78 | ND | ND | [42] |
| Bph13(t)-2 | 3S | O.officinalis | RG100, RG191 | 5.18–5.70 | ND | 4 | [43] |
| Bph14* | 3L | B5 (O. officinalis) | SM1, G1318 | 35.68~35.70 | Antibiosis | 1, 2, 3 | [11] |
| Bph15* | 4S | B5 (O. officinalis) | RG1, RG2 | 6.68~6.90 | Antixenosis | 1, 2, 3 | [44,45] |
| Bph16 | 4L | O. officinalis | G271, R93 | 20.17~21.14 | ND | 1, 2 | [39] |
| Bph17 | 4S | Rathu Heenati | RM8213, RM5953 | 4.44~9.38 | ND | ND | [46] |
| Bph18* | 12L | IR65482-7-216-1-2 (O. australiensis) | BIM3, BN162 | 22.88 | Antibiosis, antixenosis | 1 | [36,47] |
| bph18(t) | 4L | O. rufipogon | RM273, RM6506 | 24.05–25.05 | ND | 1, 2 | [48] |
| bph19(t)-1 | 3S | AS20-1 | RM6308, RM3134 | 7.18–7.24 | ND | 2 | [49] |
| bph19(t)-2 | 12L | O. rufipogon | RM17 | 26.98 | ND | 1, 2 | [48] |
| Bph20(t) | 4S | IR71033-121-15 (O. minuta) | B42, B44 | 8.76 | ND | 1 | [50] |
| bph20(t) | 6S | O. rufipogon | BYL7, BYL8 | 0.47–0.53 | ND | 2 | [51] |
| Bph21* | 12L | IR71033-121-15 (O. minuta) | S12094A, B122 | 24.20–24.36 | ND | 1 | [50] |
| bph21(t) | 10S | O. rufipogon | RM222, RM244 | 2.62–5.00 | ND | 2 | [51] |
| Bph22(t) | ND | O. glaberrima | ND | ND | ND | 4 | [52] |
| Bph23(t) | ND | O. minuta | ND | ND | ND | 4 | [52] |
| bph22(t) | 4L | O. rufipogon | RM8212, RM261 | 19.11–19.57 | ND | 1, 2 | [53] |
| bph23(t) | 8L | O. rufipogon | RM2655, RM3572 | 16.63–17.07 | ND | 1, 2 | [53] |
| bph24(t) | ND | IR73678-6-9-B (O. rufipogon) | ND | ND | ND | 4 | [54] |
| Bph25 | 6S | ADR52 | S00310, RM8101 | 0.21 | Antibiosis | ND | [55] |
| Bph26* | 12L | ADR52 | DS72B4, DS173B | 22.87–22.89 | Antibiosis | 1, 2 | [36,56] |
| Bph27 | 4L | GX2183 (O. rufipogon) | RM16846, RM16853 | 19.12–19.50 | Antibiosis, antixenosis | 1, 2 | [57] |
| Bph27(t) | 4L | Balamawee | Q52, Q20 | 20.79–21.33 | Antibiosis, antixenosis | ND | [14] |
| Bph28(t) | 11L | DV85 | InDel55, InDel66 | 16.90–16.96 | Tolerance | ND | [58] |
| Bph29* | 6S | RBPH54 (O. rufipogon) | BYL8, BID2 | 0.48–0.49 | ND | 1, 2 | [21] |
| Bph30* | 4S | AC-1613 | SSR28, SSR69 | 0.92–0.95 | Antibiosis | 1, 2, 3 | [59] |
| Bph31 | 3L | CR2711-76 | PA26, RM2334 | 26.26–26.74 | Antibiosis, antixenosis, Tolerance | 4 | [60] |
| Bph32* | 6S | Ptb33 | RM19291, RM8072 | 1.21–1.40 | Antibiosis | ND | [61] |
| Bph33 | 4S | KOLAYAL, PPLIYAL | H99, H101 | 0.91–0.97 | Antibiosis, antixenosis | ND | [62] |
| Bph33(t) | 1L | RP2068 | RM488, RM11522 | 24.80–28.00 | Antibiosis | ND | [63] |
| Bph34 | 4L | IRGC104646 (O. nivara) | RM16994, RM17007 | 21.32–21.47 | ND | 4 | [64] |
| Bph35 | 4S | RBPH660 (O. rufipogon) | PSM16, RM413 | 6.28–6.94 | ND | ND | [65] |
| Bph36 | 4S | GX2183 (O. rufipogon) | S13, X48 | 6.46–6.50 | Antibiosis, antixenosis | 1, 2 | [66] |
| Bph37-1 | 1L | IR64 | RM302, YM35 | 19.10–19.20 | Tolerance | ND | [67] |
| Bph37-2* | 6S | SE382 | ND | 1.20–1.50 | ND | NDS | [68] |
| Bph38 | 4L | GX2183 (O. rufipogon) | YM112, YM190 | 15.00–15.10 | Antibiosis, antixenosis | ND | [69] |
| Bph38(t) | 1L | Khazar | SNP693369, id10112165 | 20.71–21.23 | ND | 3 | [70] |
| Bph39 | 6S | Paedai Kalibungga | I7494, I1540 | 1.07–1.15 | Antibiosis, antixenosis | ND | [71] |
| bph39 | 4S | RPBio4918-230S (O. nivara) | RM8213, RM5953 | 4.44–9.38 | Antibiosis, tolerance | 4S | [72] |
| bph40 | 4S | RPBio4918-230S (O. nivara) | RM5953, R4M17 | 9.38–11.4 | Antibiosis, tolerance | 4S | [72] |
| Bph40* | 4S | SE232, SE67, C334 | rs4_4486223 | 4.48–4.49 | Antibiosis | 1, 2, 3 | [59] |
| Bph41-1 | 4S | GXU202(O. rufipogon) | W4_4_3, W1_6_3 | 4.68–4.78 | ND | ND | [73] |
| Bph41-2 | 4S | SWD10 | SWRm_01617, SWRm_01522 | 0.90–1.10 | Antibiosis, antixenosis | 4 | [74] |
| Bph42 | 4L | SWD10 | SWRm_01695, SWRm_00328 | 20.60–21.80 | Antibiosis, antixenosis | 4 | [74] |
| bph42 | 4S | O. rufipogon | RM16282, RM16335 | 9.07–9.58 | ND | 4 | [75] |
| Bph43 | 11L | IRGC 8678 | InDel16_22, InDel16_30 | 16.79–16.90 | Antibiosis, antixenosis | 3 | [76] |
| Bph44 | 4L | Balamawee | Q31, RM17007 | 21.38–21.47 | Tolerance | 1 | [77] |
| Bph45 | 4L | TNG71(O. nivara) | RM16655, RM3317 | 13.70–13.80 | Antixenosis | 1 | [10] |
| Bph46 | 12L | CL45 | 12M16.983, 12M19.042 | 16.99–19.04 | Antibiosis | 2 | [78] |
| qBph3 | 3L | IR02W101 (O. officinalis) | t6, f3 | 35.47–35.63 | ND | 2 | [79] |
| qBph4 | 4S | IR02W101 (O. officinalis) | P17, xc4_27 | 6.70–6.90 | ND | 2 | [79] |
| qBph6 | 6S | IR71033-121-15 | RM8120, RM8200 | 5.64–5.71 | Antibiosis | ND | [80] |
| qBph8 | 8L | Swarnalata | RM339, RM515 | 17.94–20.28 | Antixenosis | 2 | [81] |
| qbph11 | 11L | DV85 | XNpb202, C1172 | 17.43–19.56 | Tolerance | 2 | [82] |
| qBph11.3 | 11L | CL48 | RM26567, 11MA104 | 16.80–16.90 | Antibiosis | 2 | [78] |
| qBph12 | 12L | ASD7 | RM3326, RM28597 | 21.80–24.70 | Antibiosis | ND | [80] |
| qBph4.1 | 4S | Rathu Heenati | ND | 5.78–7.78 | ND | ND | [83] |
| qBph4.2-1 | 4S | IR65482-17 (O. australiensis) | RM261, XC4_27 | 6.58–6.89 | Antibiosis | 2 | [84] |
| qBph4.2-2 | 4L | Rathu Heenati | ND | 15.22–17.22 | Antixenosis | ND | [83] |
| qBph4.3 | 4S | Salkathi | RM551, RM335 | 0.18~0.69 | Antibiosis, antixenosis | 4 | [85] |
| qBph4.4 | 4S | Salkathi | RM335, RM5633 | 0.69~13.07 | Antibiosis, antixenosis | 4 | [85] |
| Gene | Chr. | Germplasm | Encoded Protein | Subcellular Localization | Reference |
|---|---|---|---|---|---|
| Bph1 | 12L | Mudgo | CC-NB-NB-LRR | Endomembrane system | [36] |
| Bph2 | 12L | ASD7 | CC-NB-NB-LRR | Endomembrane system | [36] |
| Bph3 | 4S | Rathu Heenati | LRK | Plasma membrane | [28] |
| Bph6 | 4L | Swarnalata | Atypical LRD | Exocyst | [32] |
| Bph7 | 12L | T12 | CC-NB-NB-LRR | Endomembrane system | [36] |
| Bph9 | 12L | Pokkali | CC-NB-NB-LRR | Endomembrane system | [36] |
| Bph10 | 12L | IR65482-4-136-2-2 (O. australiensis) | CC-NB-NB-LRR | Endomembrane system | [36] |
| Bph14 | 3L | B5 | CC-NB-LRR | Nucleus, cytoplasm | [11] |
| Bph15 | 4S | B5 | LRK | Plasma membrane | [45] |
| Bph18 | 12L | IR65482-7-216-1-2 (O. australiensis) | CC-NB-NB-LRR | Endomembrane system | [47] |
| Bph21 | 12L | IR71033-121-15 (O. minuta) | CC-NB-NB-LRR | Endomembrane system | [36] |
| Bph26 | 6S | ADR52 | CC-NB-NB-LRR | Endomembrane system | [36,56] |
| Bph29 | 6S | RBPH54 | B3 DNA-binding | Nucleus | [21] |
| Bph30 | 4S | AC-1613 | LRD | Endomembrane system | [59] |
| Bph32 | 6S | Ptb33 | SCR | Plasma membrane | [61] |
| Bph37 | 6L | SE382 | ND | CC-NB | [68] |
| Bph40 | 4S | SE232, SE67, C334 | ND | LRD | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Gu, D.; Liu, D.; Hassan, M.A.; Yu, C.; Wu, X.; Huang, S.; Bian, S.; Wei, P.; Li, J. Recent Advances in Gene Mining and Hormonal Mechanism for Brown Planthopper Resistance in Rice. Int. J. Mol. Sci. 2024, 25, 12965. https://doi.org/10.3390/ijms252312965
Zhang X, Gu D, Liu D, Hassan MA, Yu C, Wu X, Huang S, Bian S, Wei P, Li J. Recent Advances in Gene Mining and Hormonal Mechanism for Brown Planthopper Resistance in Rice. International Journal of Molecular Sciences. 2024; 25(23):12965. https://doi.org/10.3390/ijms252312965
Chicago/Turabian StyleZhang, Xiao, Dongfang Gu, Daoming Liu, Muhammad Ahmad Hassan, Cao Yu, Xiangzhi Wu, Shijie Huang, Shiquan Bian, Pengcheng Wei, and Juan Li. 2024. "Recent Advances in Gene Mining and Hormonal Mechanism for Brown Planthopper Resistance in Rice" International Journal of Molecular Sciences 25, no. 23: 12965. https://doi.org/10.3390/ijms252312965
APA StyleZhang, X., Gu, D., Liu, D., Hassan, M. A., Yu, C., Wu, X., Huang, S., Bian, S., Wei, P., & Li, J. (2024). Recent Advances in Gene Mining and Hormonal Mechanism for Brown Planthopper Resistance in Rice. International Journal of Molecular Sciences, 25(23), 12965. https://doi.org/10.3390/ijms252312965