
Structure of Genes Encoding Oxidosqualene Cyclases—Key Enzymes of Triterpenoid Biosynthesis from Sea Cucumber Eupentacta fraudatrix

 , , , ,
, , , , 
Abstract
1. Introduction
2. Results and Discussion
2.1. E. fraudatrix Genome Codes Two Different OSCs
2.1.1. cDNA Determination of OSC mRNAs
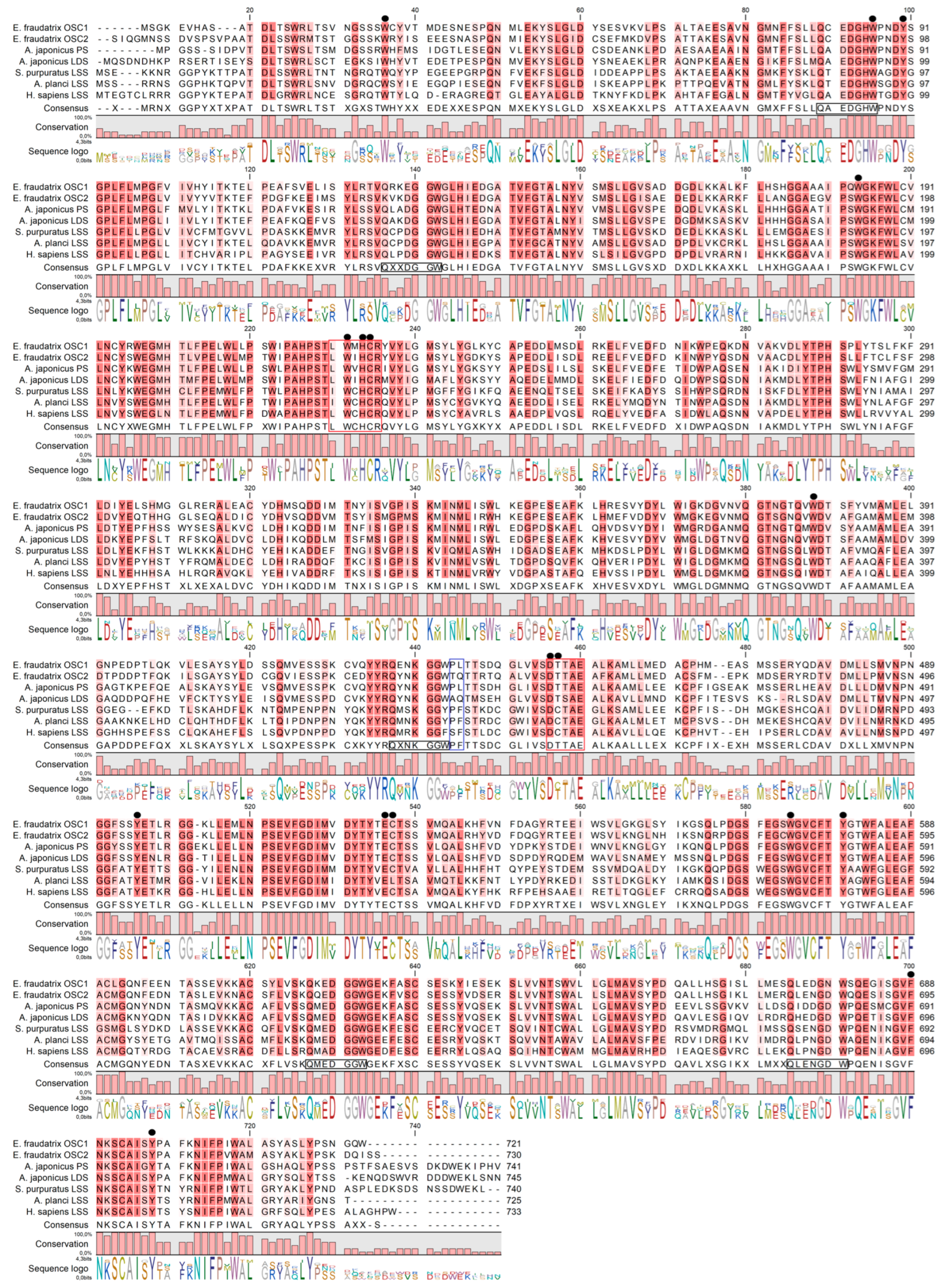
2.1.2. Sequence Analyses of OSC1 and OSC2 Proteins
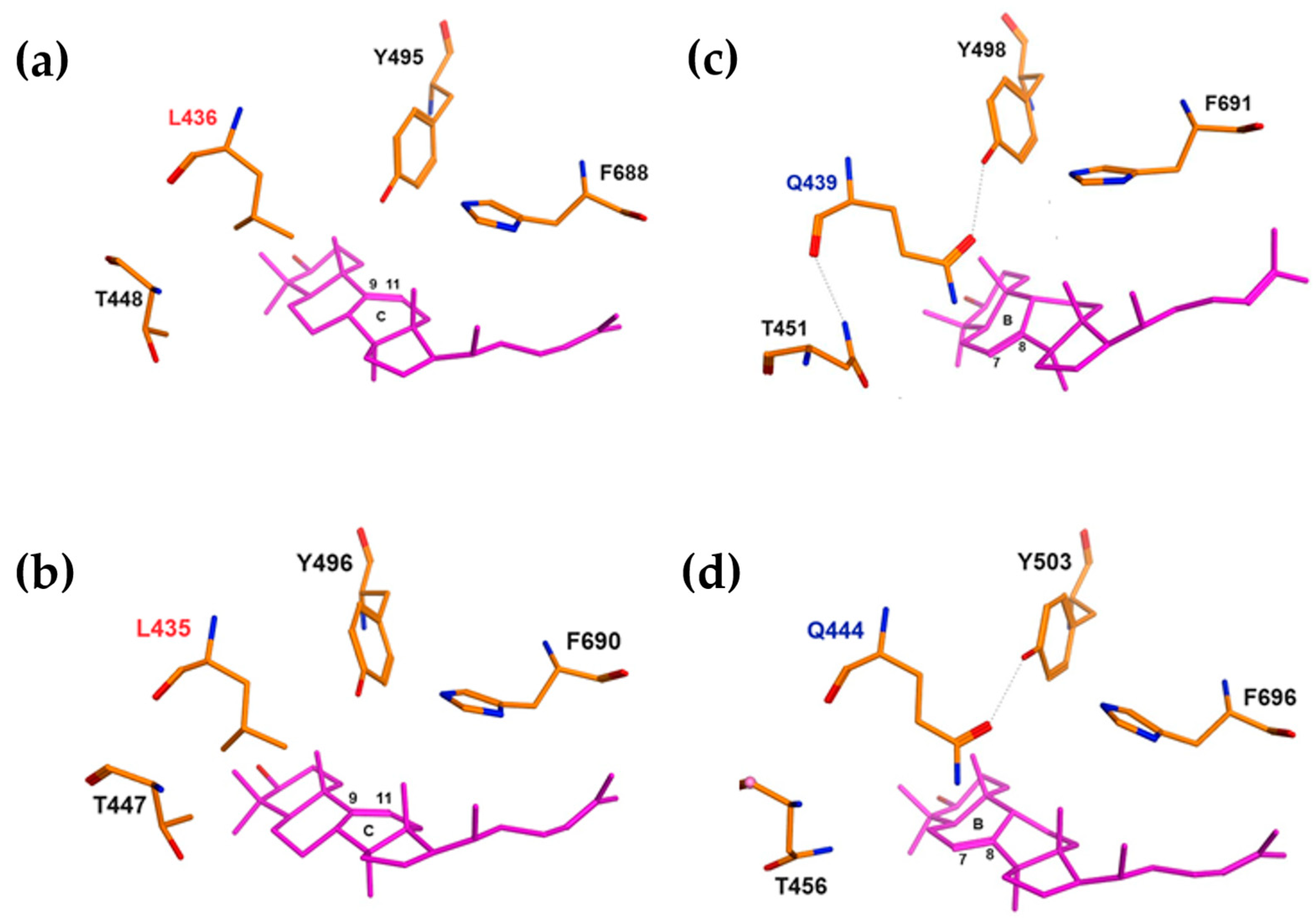
2.2. Confirmation of Key L and Q Residues Distinguishing Parkeol and Lanostadienol Synthases by Molecular Docking
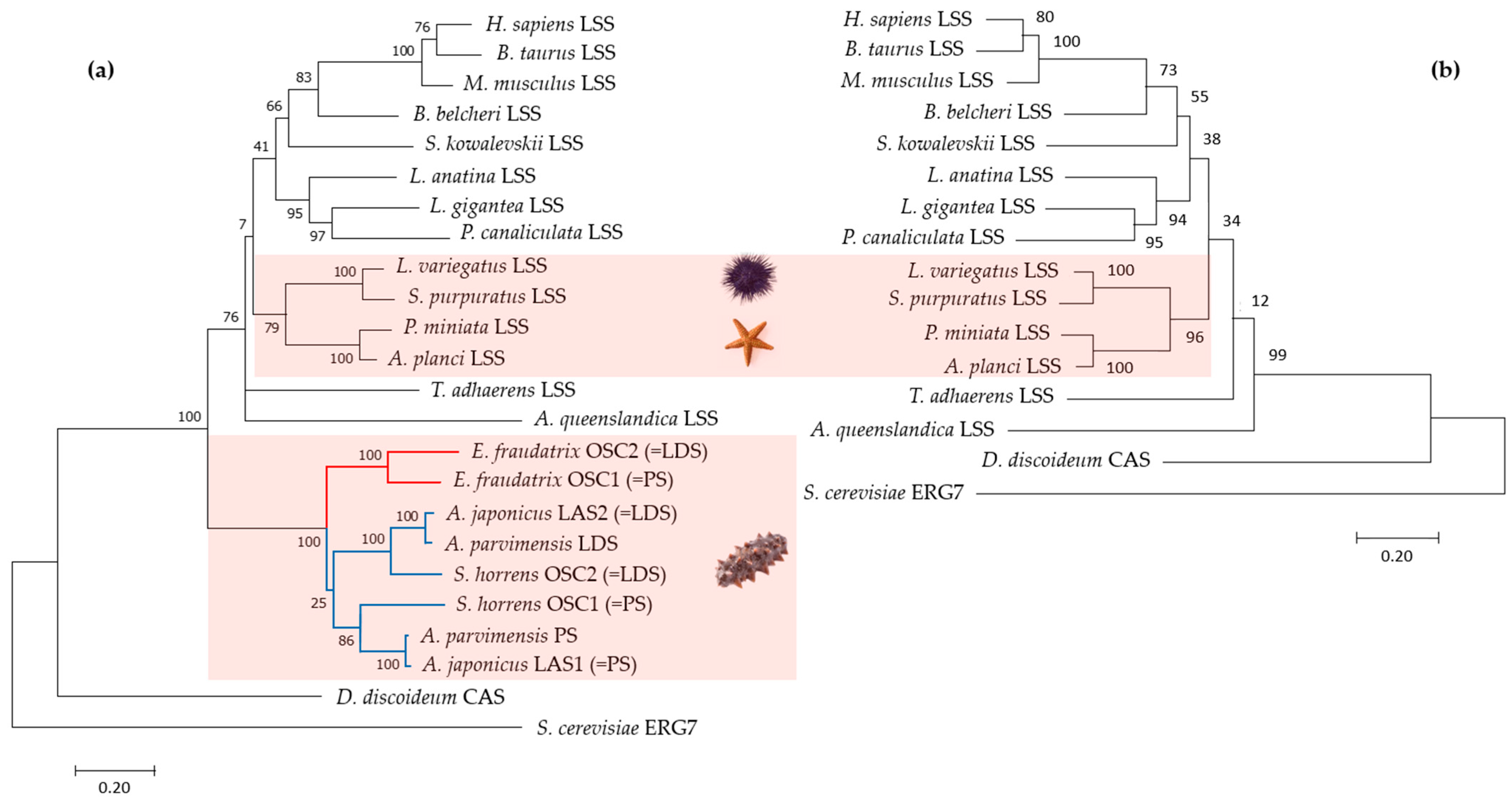
2.3. E. fraudatrix OSC1 and OSC2 form a Phylogenetically Distinct Branch
2.4. Gene Structure Determination and Analysis of OSC Genes
3. Materials and Methods
3.1. Sample Collection and Nucleic Acids Extraction
3.2. cDNA Synthesis, RACE, and PCR Amplification
3.3. Gene Structure Determination
3.4. Sequence Analysis, Homology Modeling, and Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dyakonov, A.; Baranova, Z.; Saveleva, T. Note on Holothurioidea of the South Sakhalin and South Kurile Islands area. Investig. Seas USSR 1958, 5, 358–380. [Google Scholar]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Menchinskaya, E.S.; Kalinin, V.I. Structure of cucumarioside I2 from the sea cucumber Eupentacta fraudatrix (Djakonov et Baranova) and cytotoxic and immunostimulatory activities of this saponin and relative compounds. Nat. Prod. Res. 2013, 27, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Kalinin, V.I.; Stonik, V.A. 3β-O-Glycosylated 16β-acetoxy-9β-H-lanosta-7,24-diene-3β,18,20β-triol, an intermediate metabolite from the sea cucumber Eupentacta fraudatrix and its biosynthetic significance. Biochem. Syst. Ecol. 2012, 44, 53–60. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Martyyas, E.A.; Kalinin, V.I. Triterpene glycosides from the sea cucumber Eupentacta fraudatrix. Structure and biological action of cucumariosides A1, A3, A4, A5, A6, A12 and A15, seven new minor non-sulfated tetraosides and unprecedented 25-keto, 27-norholostane aglycone. Nat. Prod. Commun. 2012, 7, 517–525. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Martyyas, E.A.; Kalinin, V.I. Triterpene Glycosides from the Sea Cucumber Eupentacta fraudatrix. Structure and Biological Action of Cucumariosides I1, I3, I4, Three New Minor Disulfated Pentaosides. Nat. Prod. Commun. 2013, 8, 1053–1058. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Stonik, V.A.; Kapustina, I.I.; Boguslavsky, V.M.; Dmitrenoik, A.S.; Kalinin, V.I.; Cordeiro, M.L.; Djerassi, C. Biosynthetic studies of marine lipids. 42. Biosynthesis of steroid and triterpenoid metabolites in the sea cucumber Eupentacta fraudatrix. Steroids 1993, 58, 508–517. [Google Scholar] [CrossRef]
- Popov, A.M. Comparative Study of Effects of Various Sterols and Triterpenoids on Permeability of Model Lipid Membranes. J. Evol. Biochem. Physiol. 2003, 39, 314–320. [Google Scholar] [CrossRef]
- Popov, R.S.; Ivanchina, N.V.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Dolmatov, I.Y.; Dmitrenok, P.S. Metabolite profiling of triterpene glycosides of the Far Eastern sea cucumber Eupentacta fraudatrix and their distribution in various body components using LC-ESI QTOF-MS. Mar. Drugs 2017, 15, 302. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Popov, R.S.; Kalinin, V.I.; Andrijaschenko, P.V.; Dmitrenok, P.S.; Yurchenko, E.A. Triterpene Glycosides from the Sea Cucumber Eupentacta fraudatrix. Structure and Cytotoxic action of Cucumarioside D with a Terminal 3-O-Me-Glucose Residue Unique for this Species. Nat. Prod. Commun. 2018, 13, 1137–1140. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Stonik, V.A. Triterpene glycosides from sea cucumbers (Holothurioidea, Echinodermata): Biological activities and functions. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2008; Volume 35, pp. 135–196. [Google Scholar]
- Mitu, S.A.; Bose, U.; Suwansa-Ard, S.; Turner, L.H.; Zhao, M.; Elizur, A.; Ogbourne, S.M.; Shaw, P.N.; Cummins, S.F. Evidence for a saponin biosynthesis pathway in the body wall of the commercially significant sea cucumber Holothuria scabra. Mar. Drugs 2017, 15, 349. [Google Scholar] [CrossRef]
- Thimmappa, R.; Wang, S.; Zheng, M.; Misra, R.C.; Huang, A.C.; Saalbach, G.; Chang, Y.; Zhou, Z.; Hinman, V.; Bao, Z.; et al. Biosynthesis of saponin defensive compounds in sea cucumbers. Nat. Chem. Biol. 2022, 18, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, R.; Xun, X.; Wang, J.; Bao, L.; Thimmappa, R.; Ding, J.; Jiang, J.; Zhang, L.; Li, T.; et al. Sea cucumber genome provides insights into saponin biosynthesis and aestivation regulation. Cell Discov. 2018, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kong, X.; Chen, J.; Zhang, H. De novo sequencing and transcriptome analysis of Stichopus horrens to reveal genes related to biosynthesis of triterpenoids. Aquaculture 2018, 491, 358–367. [Google Scholar] [CrossRef]
- Boyko, A.V.; Girich, A.S.; Tkacheva, E.S.; Dolmatov, I.Y. The Eupentacta fraudatrix transcriptome provides insights into regulation of cell transdifferentiation. Sci. Rep. 2020, 10, 1522. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; A Salazar, G.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2022, 50, D16–D25. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; A Yamashita, R.; et al. The Conserved Domain Database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef]
- Sato, T.; Hoshino, T. Functional Analysis of the DXDDTA Motif in Squalene-Hopene Cyclase by Site-Directed Mutagenesis Experiments: Initiation Site of the Polycyclization Reaction and Stabilization Site of the Carbocation Intermediate of the Initially Cyclized A-Rin. Biosci. Biotechnol. Biochem. 1999, 63, 2189–2198. [Google Scholar] [CrossRef]
- Poralla, K.; Hewelt, A.; Prestwich, G.D.; Abe, I.; Reipen, I.; Sprenger, G. A specific amino acid repeat in squalene and oxidosqualene cyclases. Trends Biochem. Sci. 1994, 19, 157–158. [Google Scholar] [CrossRef]
- Kushiro, T.; Shibuya, M.; Ebizuka, Y. β-Amyrin synthase: Cloning of oxidosqualene cyclase that catalyzes the formation of the most popular triterpene among higher plants. Eur. J. Biochem. 1998, 256, 238–244. [Google Scholar] [CrossRef]
- Godio, R.P.; Martín, J.F. Modified oxidosqualene cyclases in the formation of bioactive secondary metabolites: Biosynthesis of the antitumor clavaric acid. Fungal Genet. Biol. 2009, 46, 232–242. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lee, Y.R.; Tsao, N.W.; Wang, S.Y.; Shaw, J.F.; Chu, F.H. Characterization of the 2,3-Oxidosqualene Cyclase Gene from Antrodia cinnamomea and Enhancement of Cytotoxic Triterpenoid Compound Production. J. Nat. Prod. 2015, 78, 1556–1562. [Google Scholar] [CrossRef] [PubMed]
- Siedenburg, G.; Jendrossek, D. Squalene-Hopene Cyclases. Appl. Environ. Microbiol. 2011, 77, 3905–3915. [Google Scholar] [CrossRef] [PubMed]
- Corey, E.J.; Cheng, H.; Baker, C.H.; Matsuda, S.P.T.; Li, D.; Song, X. Studies on the Substrate Binding Segments and Catalytic Action of Lanosterol Synthase. Affinity Labeling with Carbocations Derived from Mechanism-Based Analogs of 2,3-Oxidosqualene and Site-Directed Mutagenesis Probes. J. Am. Chem. Soc. 1997, 119, 1289–1296. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, M.; Ye, M.; Qiao, X. Site-Directed Mutagenesis and Substrate Compatibility to Reveal the Structure–Function Relationships of Plant Oxidosqualene Cyclases. Nat. Prod. Rep. 2021, 38, 2261–2275. [Google Scholar] [CrossRef]
- Thoma, R.; Schulz-Gasch, T.; D’Arcy, B.; Benz, J.; Aebi, J.; Dehmlow, H.; Hennig, M.; Stihle, M.; Ruf, A. Insight into Steroid Scaffold Formation from the Structure of Human Oxidosqualene Cyclase. Nature 2004, 432, 118–122. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef]
- Mongiardino, K.N.; Tilic, E.; Miller, A.K.; Stiller, J.; Rouse, G.W. Confusion will be my epitaph: Genome-scale discordance stifles phylogenetic resolution of Holothuroidea. Proc. R. Soc. B 2023, 290, 20230988. [Google Scholar] [CrossRef]
- Hoff, K.J.; Lomsadze, A.; Borodovsky, M.; Stanke, M. Whole-Genome Annotation with BRAKER. In Gene Prediction: Methods and Protocols; Kollmar, M., Ed.; Springer: New York, NY, USA, 2019; pp. 65–95. [Google Scholar] [CrossRef]
- Kuznetsov, D.; Tegenfeldt, F.; Manni, M.; Seppey, M.; Berkeley, M.; Kriventseva, E.V.; Zdobnov, E.M. OrthoDB v11: Annotation of orthologs in the widest sampling of organismal diversity. Nucleic Acids Res. 2023, 51, D445–D451. [Google Scholar] [CrossRef]
- Sun, L.; Jiang, C.; Su, F.; Cui, W.; Yang, H. Chromosome-level genome assembly of the sea cucumber Apostichopus japonicus. Sci. Data 2023, 10, 454. [Google Scholar] [CrossRef]
- Chen, T.; Ren, C.; Wong, N.-K.; Yan, A.; Sun, C.; Fan, D.; Luo, P.; Jiang, X.; Zhang, L.; Ruan, Y.; et al. The Holothuria leucospilota genome elucidates sacrificial organ expulsion and bio-adhesive trap enriched with amyloid-patterned proteins. Proc. Natl. Acad. Sci. USA 2023, 120, e2213512120. [Google Scholar] [CrossRef]
- Hall, M.R.; Kocot, K.M.; Baughman, K.W.; Fernandez-Valverde, S.L.; Gauthier, M.E.A.; Hatleberg, W.L.; Krishnan, A.; McDougall, C.; Motti, C.A.; Shoguchi, E.; et al. The crown-of-thorns starfish genome as a guide for biocontrol of this coral reef pest. Nature 2017, 544, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Sea Urchin Genome Sequencing Consortium; Sodergren, E.; Wright, R. The genome of the sea urchin Strongylocentrotus purpuratus. Science 2006, 314, 941–952. [Google Scholar] [CrossRef]
- Davidson, P.L.; Guo, H.; Wang, L.; Berrio, A.; Zhang, H.; Chang, Y.; Soborowski, A.L.; McClay, D.R.; Fan, G.; Wray, G.A. Chromosomal-level genome assembly of the sea urchin Lytechinus variegatus substantially improves functional genomic analyses. Genome Biol. Evol. 2020, 12, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef] [PubMed]
- Telmer, C.A.; Karimi, K.; Chess, M.M.; Agalakov, S.; Arshinoff, B.I.; Lotay, V.; Wang, D.Z.; Chu, S.; Pells, T.J.; Vize, P.D.; et al. Echinobase: A resource to support the echinoderm research community. Genetics 2024, 227, iyae002. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Madden, T.L. The BLAST Sequence Analysis Tool. In The NCBI Handbook; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013; pp. 425–436. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OSC1 | OSC2 | AjLAS1 | AjLAS2 | ApPS | ApLDS | ShOSC1 | ShOSC2 | SpLSS | ApLSS | |
|---|---|---|---|---|---|---|---|---|---|---|
| E. fraudatrix OSC1 | 100 | 87.18 | 82.11 | 81.97 | 82.66 | 81.69 | 82.25 | 81.84 | 73.38 | 72.65 |
| E. fraudatrix OSC2 | 77.5 | 100 | 81.17 | 80.62 | 81.31 | 80.76 | 79.44 | 80.85 | 71.89 | 71.81 |
| A. japonicus LAS1 * | 70.6 | 67.2 | 100 | 84.86 | 98.79 | 84.89 | 88.48 | 83.47 | 72.73 | 73.71 |
| A. japonicus LAS2 * | 69.9 | 67.3 | 72.4 | 100 | 85.27 | 98.27 | 83.83 | 90.21 | 73.00 | 75.71 |
| A. parvimensis PS * | 71.0 | 67.5 | 98.3 | 72.4 | 100 | 85.30 | 88.31 | 83.62 | 72.86 | 74.02 |
| A. parvimensis LDS * | 69.7 | 67.8 | 72.4 | 96.0 | 73.1 | 100 | 83.54 | 90.62 | 72.73 | 75.14 |
| S. horrens OSC1 | 68.7 | 64.7 | 76.6 | 68.5 | 76.1 | 69.1 | 100 | 86.02 | 73.52 | 74.75 |
| S. horrens OSC2 | 68.1 | 67.5 | 71.3 | 80.1 | 71.6 | 81.3 | 71.6 | 100 | 73.68 | 75.32 |
| S. purpuratus LSS * | 57.75 | 55.65 | 58.89 | 57.30 | 58.75 | 56.75 | 57.46 | 57.52 | 100 | 80.77 |
| A. planci LSS * | 57.92 | 56.37 | 59.11 | 58.19 | 58.94 | 57.63 | 57.55 | 57.97 | 69.43 | 100 |
| Primers Name | 5′ -> 3′ Sequence | Objectives |
|---|---|---|
| OSC1_start | ATGTCTGGCAAAGAKGTSCATGCAAGTG | OSC1 cDNA amplification and gDNA amplicons sequencing. |
| OSC1_stop | ATCACTGGCCATTTGAAGGATAGAGCCTG | |
| OSC2_start | AGAATTCAAGGAATGAATTCCAGTGACGTCTC | OSC2 cDNA amplification and gDNA amplicons sequencing. |
| OSC2_stop | TCTCGAGATCTGGTCCTTCGATGGGTAGAGCT | |
| OSS_838For | GCCTACTCATACCTTGATTCTTCACAGA | OSC1 RACE and gDNA amplicons sequencing. |
| OSS_865Rev | TCTGTGAAGAATCAAGGTATGAGTAGGC | |
| OSC2_F2 | AGGACCAAAGAGATAATGTTGCTGCATGT | OSC2 RACE and gDNA amplicons sequencing. |
| OSC2_2RR | TGTTGGATCATCGGGCGTAT | |
| OSS_1143Rev | CAACATCTCCAAGAGTTTGCCWCCTCGG | OSC1 RACE and Control PCR of gDNA with OSS_838F. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldaev, S.N.; Chausova, V.E.; Isaeva, K.V.; Boyko, A.V.; Stonik, V.A.; Isaeva, M.P. Structure of Genes Encoding Oxidosqualene Cyclases—Key Enzymes of Triterpenoid Biosynthesis from Sea Cucumber Eupentacta fraudatrix. Int. J. Mol. Sci. 2024, 25, 12881. https://doi.org/10.3390/ijms252312881
Baldaev SN, Chausova VE, Isaeva KV, Boyko AV, Stonik VA, Isaeva MP. Structure of Genes Encoding Oxidosqualene Cyclases—Key Enzymes of Triterpenoid Biosynthesis from Sea Cucumber Eupentacta fraudatrix. International Journal of Molecular Sciences. 2024; 25(23):12881. https://doi.org/10.3390/ijms252312881
Chicago/Turabian StyleBaldaev, Sergey N., Viktoria E. Chausova, Ksenia V. Isaeva, Alexey V. Boyko, Valentin A. Stonik, and Marina P. Isaeva. 2024. "Structure of Genes Encoding Oxidosqualene Cyclases—Key Enzymes of Triterpenoid Biosynthesis from Sea Cucumber Eupentacta fraudatrix" International Journal of Molecular Sciences 25, no. 23: 12881. https://doi.org/10.3390/ijms252312881
APA StyleBaldaev, S. N., Chausova, V. E., Isaeva, K. V., Boyko, A. V., Stonik, V. A., & Isaeva, M. P. (2024). Structure of Genes Encoding Oxidosqualene Cyclases—Key Enzymes of Triterpenoid Biosynthesis from Sea Cucumber Eupentacta fraudatrix. International Journal of Molecular Sciences, 25(23), 12881. https://doi.org/10.3390/ijms252312881