
Site Matters: Differences in Gene Expression Profiles Along the Bovine Rumen Papilla During Subacute Rumen Acidosis

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
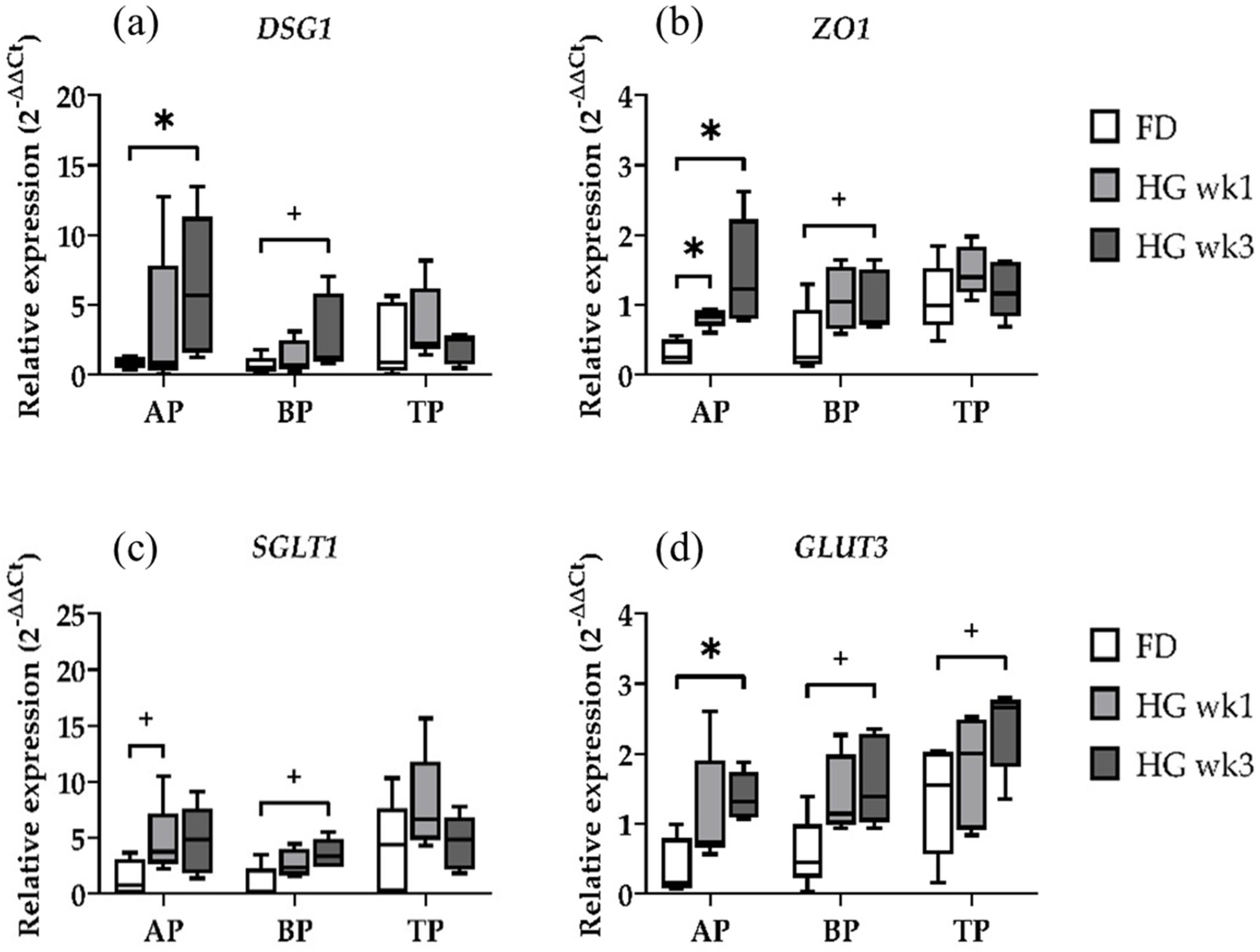
2.1. Gene Expression of Epithelial Integrity and Nutrient Transport Genes
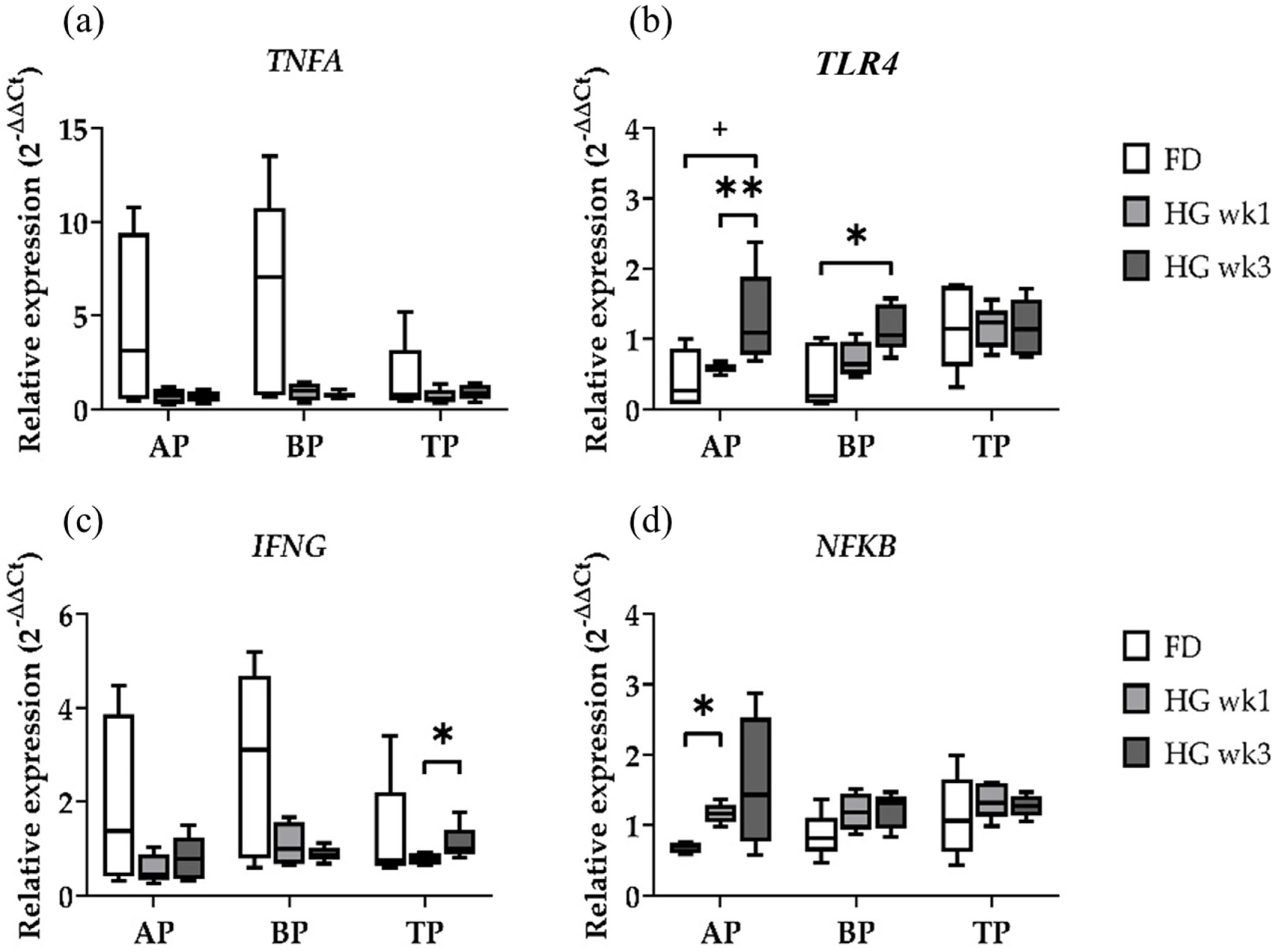
2.2. Gene Expression of Immune Response Genes
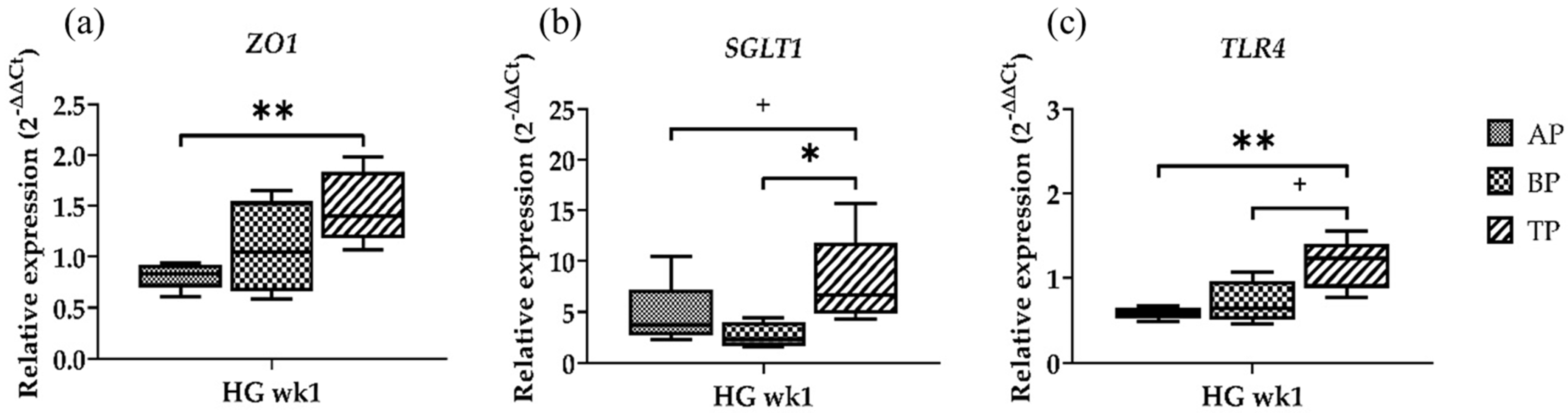
2.3. Gene Expression at Different Sampling Locations
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Tissue Collection
4.3. Histology of Papillae Biopsies
4.4. Tissue Homogenization, RNA Isolation, and RNA Integrity and Quantity
4.5. Primers: Selection, Design, and Validation
4.6. RT-PCR and RT-qPCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ross, E.M.; Moate, P.J.; Bath, C.R.; Davidson, S.E.; Sawbridge, T.I.; Guthridge, K.M.; Cocks, B.G.; Hayes, B.J. High throughput whole rumen metagenome profiling using untargeted massively parallel sequencing. BMC Genet. 2012, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef]
- Dijkstra, J.; Ellis, J.L.; Kebreab, E.; Strathe, A.B.; López, S.; France, J.; Bannink, A. Ruminal pH regulation and nutritional consequences of low pH. Anim. Feed Sci. Technol. 2012, 172, 22–33. [Google Scholar] [CrossRef]
- Lehr- und Forschungszentrum für Landwirtschaft Raumberg-Gumpenstein. In Proceedings of the 39 Viehwirtschaftliche Fachtagung gemäß Fortbildungsplan des Bundes: Milchproduktion—Status quo und Anpassung an Zukünftige Herausforderungen, Irdning, Austria, 25–26 April 2012.
- Humer, E.; Petri, R.M.; Aschenbach, J.R.; Bradford, B.J.; Penner, G.B.; Tafaj, M.; Südekum, K.H.; Zebeli, Q. Invited review: Practical feeding management recommendations to mitigate the risk of subacute ruminal acidosis in dairy cattle. J. Dairy Sci. 2018, 101, 872–888. [Google Scholar] [CrossRef]
- Stauder, A.; Humer, E.; Neubauer, V.; Reisinger, N.; Kaltenegger, A.; Zebeli, Q. Distinct responses in feed sorting, chewing behavior, and ruminal acidosis risk between primiparous and multiparous Simmental cows fed diets differing in forage and starch levels. J. Dairy Sci. 2020, 103, 8467–8481. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [CrossRef]
- Penner, G.B.; Oba, M.; Gäbel, G.; Aschenbach, J.R. A single mild episode of subacute ruminal acidosis does not affect ruminal barrier function in the short term. J. Dairy Sci. 2010, 93, 4838–4845. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Lopez, E.; Petri, R.M.; Ricci, S.; Rivera-Chacon, R.; Sener-Aydemir, A.; Sharma, S.; Reisinger, N.; Zebeli, Q. Dynamic changes in salivation, salivary composition, and rumen fermentation associated with duration of high-grain feeding in cows. J. Dairy Sci. 2021, 104, 4875–4892. [Google Scholar] [CrossRef]
- Tsuba, T.; Sasaki, J.; Kawashima, R. (Eds.) Physiological Aspects of Digestion and Metabolism in Ruminants: Proceedings of the 7th International Symposium on Ruminant Physiology, 28 September 1989 in Sendai Japan; Academic Press: London, UK, 1991. [Google Scholar]
- Kern, R.J.; Lindholm-Perry, A.K.; Freetly, H.C.; Kuehn, L.A.; Rule, D.C.; Ludden, P.A. Rumen papillae morphology of beef steers relative to gain and feed intake and the association of volatile fatty acids with kallikrein gene expression. Livest. Sci. 2016, 187, 24–30. [Google Scholar] [CrossRef]
- Meissner, S.; Hagen, F.; Deiner, C.; Günzel, D.; Greco, G.; Shen, Z.; Aschenbach, J.R. Key role of short-chain fatty acids in epithelial barrier failure during ruminal acidosis. J. Dairy Sci. 2017, 100, 6662–6675. [Google Scholar] [CrossRef]
- Baumgartner, W.; Braun, U. Innere Medizin und Chirurgie des Rindes; Dirksen, G., Gründer, H.-D., Stöber, M., Eds.; Parey: Stuttgart, Germany, 2006. [Google Scholar]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Technol. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Oetzel, G.R. Diagnosis and Management of Subacute Ruminal Acidosis in Dairy Herds. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 463–480. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Seyfert, H.-M.; Löhrke, B.; Schneider, F.; Zitnan, R.; Chudy, A.; Kuhla, S.; Hammon, H.M.; Blum, J.W.; Martens, H.; et al. An energy-rich diet causes rumen papillae proliferation associated with more IGF type 1 receptors and increased plasma IGF-1 concentrations in young goats. J. Nutr. 2004, 134, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.A.; Schiestel, C.; AlZahal, O.; Dionissopoulos, L.; Laarman, A.H.; Matthews, J.C.; McBride, B.W. The periparturient period is associated with structural and transcriptomic adaptations of rumen papillae in dairy cattle. J. Dairy Sci. 2015, 98, 2583–2595. [Google Scholar] [CrossRef] [PubMed]
- Zebeli, Q.; Dijkstra, J.; Tafaj, M.; Steingass, H.; Ametaj, B.N.; Drochner, W. Modeling the adequacy of dietary fiber in dairy cows based on the responses of ruminal pH and milk fat production to composition of the diet. J. Dairy Sci. 2008, 91, 2046–2066. [Google Scholar] [CrossRef]
- Hua, D.; Hendriks, W.H.; Xiong, B.; Pellikaan, W.F. Starch and Cellulose Degradation in the Rumen and Applications of Metagenomics on Ruminal Microorganisms. Animals 2022, 12, 3020. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.; Simmons, N.L. Functional organization of the bovine rumen epithelium. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R173–R181. [Google Scholar] [CrossRef]
- Ferraris, R.P.; Diamond, J.M. Crypt/villus site of substrate-dependent regulation of mouse intestinal glucose transporters. Proc. Natl. Acad. Sci. USA 1993, 90, 5868–5872. [Google Scholar] [CrossRef]
- Lescale-Matys, L.; Dyer, J.; Scott, D.; Freeman, T.C.; Wright, E.M.; Shirazi-Beechey, S.P. Regulation of the ovine intestinal Na+/glucose co-transporter (SGLT1) is dissociated from mRNA abundance. Biochem. J. 1993, 291, 435–440. [Google Scholar] [CrossRef]
- Shirazi-Beechey, S.P.; Gribble, S.M.; Wood, I.S.; Tarpey, P.S.; Beechey, R.B.; Dyer, J.; Scott, D.; Barker, P.J. Dietary regulation of the intestinal sodium-dependent glucose cotransporter (SGLT1). Biochem. Soc. Trans. 1994, 22, 655–658. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Wehning, H.; Kurze, M.; Schaberg, E.; Nieper, H.; Burckhardt, G.; Gäbel, G. Functional and molecular biological evidence of SGLT-1 in the ruminal epithelium of sheep. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G20–G27. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Q.; Okine, E.K.; Cheeseman, C.I.; Shirazi-Beechey, S.P.; Kennelly, J.J. Glucose transporter gene expression in lactating bovine gastrointestinal tract. J. Anim. Sci. 1998, 76, 2921–2929. [Google Scholar] [CrossRef] [PubMed]
- Aschenbach, J.R.; Bhatia, S.K.; Pfannkuche, H.; Gäbel, G. Glucose is absorbed in a sodium-dependent manner from forestomach contents of sheep. J. Nutr. 2000, 130, 2797–2801. [Google Scholar] [CrossRef]
- Molteni, M.; Gemma, S.; Rossetti, C. The Role of Toll-Like Receptor 4 in Infectious and Noninfectious Inflammation. Mediators Inflamm. 2016, 2016, 6978936. [Google Scholar] [CrossRef]
- Rivera-Chacon, R.; Castillo-Lopez, E.; Ricci, S.; Petri, R.M.; Reisinger, N.; Zebeli, Q. Supplementing a Phytogenic Feed Additive Modulates the Risk of Subacute Rumen Acidosis, Rumen Fermentation and Systemic Inflammation in Cattle Fed Acidogenic Diets. Animals 2022, 12, 1201. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Xie, F.; Wang, J.; Zhang, B. RefFinder: A web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 23, 125. [Google Scholar] [CrossRef]
- Petri, R.M.; Wetzels, S.U.; Qumar, M.; Khiaosa-Ard, R.; Zebeli, Q. Adaptive responses in short-chain fatty acid absorption, gene expression, and bacterial community of the bovine rumen epithelium recovered from a continuous or transient high-grain feeding. J. Dairy Sci. 2019, 102, 5361–5378. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.A.; Dionissopoulos, L.; AlZahal, O.; Doelman, J.; McBride, B.W. Rumen epithelial adaptation to ruminal acidosis in lactating cattle involves the coordinated expression of insulin-like growth factor-binding proteins and a cholesterolgenic enzyme. J. Dairy Sci. 2012, 95, 318–327. [Google Scholar] [CrossRef]
- Liu, J.; Xu, T.; Liu, Y.; Zhu, W.; Mao, S. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R232–R241. [Google Scholar] [CrossRef]
- Jin, D.; Chang, G.; Zhang, K.; Guo, J.; Xu, T.; Shen, X. Rumen-derived lipopolysaccharide enhances the expression of lingual antimicrobial peptide in mammary glands of dairy cows fed a high-concentrate diet. BMC Vet. Res. 2016, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Metzler-Zebeli, B.U.; Hollmann, M.; Sabitzer, S.; Podstatzky-Lichtenstein, L.; Klein, D.; Zebeli, Q. Epithelial response to high-grain diets involves alteration in nutrient transporters and Na+/K+-ATPase mRNA expression in rumen and colon of goats. J. Anim. Sci. 2013, 91, 4256–4266. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequence (5′ to 3′) | Annealing Temperature (°C) | Amplicon Size (bp) | References | |
|---|---|---|---|---|---|---|
| Reference genes | ||||||
| HPRT1 | NW_005397637.1 | F: | TTGTATACCCAATCATTATGCTGAG | 58 | 109 | This study |
| R: | ACCCATCTCCTTCATCACATCT | |||||
| OAZ1 | NM_001127243.2 | F: | CACAAGAACCGTGATGATCGA | 58 | 69 | Petri et al., 2018 [31] |
| R: | TCTCACAATCTCAAAGCCCAAA | |||||
| Epithelial barrier function | ||||||
| DSG1 | NM_174045.1 | F: | AGACAGAGAGCAATATGGCCAGT | 60 | 88 | Steele et al., 2010 [32] |
| R: | TTCACACTCTGCTGACATACCATCT | |||||
| ZO1 | XM_024982012.1 | F: | CGACCAGATCCTCAGGGTAA | 60 | 161 | Liu et al., 2013 [33] |
| R: | AATCACCCACATCGGATTCT | |||||
| Innate immune response | ||||||
| TNFA | NM_173966.3 | F: | AGCCCTCTRGTTCARACACT | 58 | 159 | This study |
| R: | GCTGGTTGTCTTCCAGCTTC | |||||
| TLR4 | XM_005210586.3 | F: | TGGGACCCTTGCGTACAG | 60 | 159 | This study |
| R: | ACGGCCACCAGCTTCTG | |||||
| Adaptive immune response | ||||||
| IFNG | NM_174086.1 | F: | GCAGCTCTGAGAAACTGGAGGA | 58 | 79 | This study |
| R: | ATGGCTTTGCGCTGGATCT | |||||
| NFKB | NM_001076409.1 | F: | ATACGTCGGCCGTGTCTAT | 58 | 144 | Jin et al., 2016 [34] |
| R: | GGAACTGTGATCCGTGTAG | |||||
| Glucose transport | ||||||
| SGLT1 | XM_005218131.3 | F: | ACATCGCCTACCCGACCTT | 64 | 74 | Metzler-Zebeli et al., 2014 [35] |
| R: | CAGCATGACCGACAGCATCA | |||||
| GLUT3 | NM_174603.3 | F: | ATGTCGCAGGAGAAGCAAGT | 64 | 64 | This study |
| R: | AGACACCACGGTGAAGATG | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sener-Aydemir, A.; Dengler, F.; Larsberg, F.; Rivera-Chacon, R.; Castillo-Lopez, E.; Zebeli, Q.; Kreuzer-Redmer, S. Site Matters: Differences in Gene Expression Profiles Along the Bovine Rumen Papilla During Subacute Rumen Acidosis. Int. J. Mol. Sci. 2024, 25, 12303. https://doi.org/10.3390/ijms252212303
Sener-Aydemir A, Dengler F, Larsberg F, Rivera-Chacon R, Castillo-Lopez E, Zebeli Q, Kreuzer-Redmer S. Site Matters: Differences in Gene Expression Profiles Along the Bovine Rumen Papilla During Subacute Rumen Acidosis. International Journal of Molecular Sciences. 2024; 25(22):12303. https://doi.org/10.3390/ijms252212303
Chicago/Turabian StyleSener-Aydemir, Arife, Franziska Dengler, Filip Larsberg, Raul Rivera-Chacon, Ezequias Castillo-Lopez, Qendrim Zebeli, and Susanne Kreuzer-Redmer. 2024. "Site Matters: Differences in Gene Expression Profiles Along the Bovine Rumen Papilla During Subacute Rumen Acidosis" International Journal of Molecular Sciences 25, no. 22: 12303. https://doi.org/10.3390/ijms252212303
APA StyleSener-Aydemir, A., Dengler, F., Larsberg, F., Rivera-Chacon, R., Castillo-Lopez, E., Zebeli, Q., & Kreuzer-Redmer, S. (2024). Site Matters: Differences in Gene Expression Profiles Along the Bovine Rumen Papilla During Subacute Rumen Acidosis. International Journal of Molecular Sciences, 25(22), 12303. https://doi.org/10.3390/ijms252212303