
6′-O-Caffeoylarbutin from Quezui Tea: A Highly Effective and Safe Tyrosinase Inhibitor

 ,
,
Abstract
1. Introduction
2. Results and Discussion
2.1. mTyr Inhibition Mechanism of 6′-O-Caffeoylarbutin
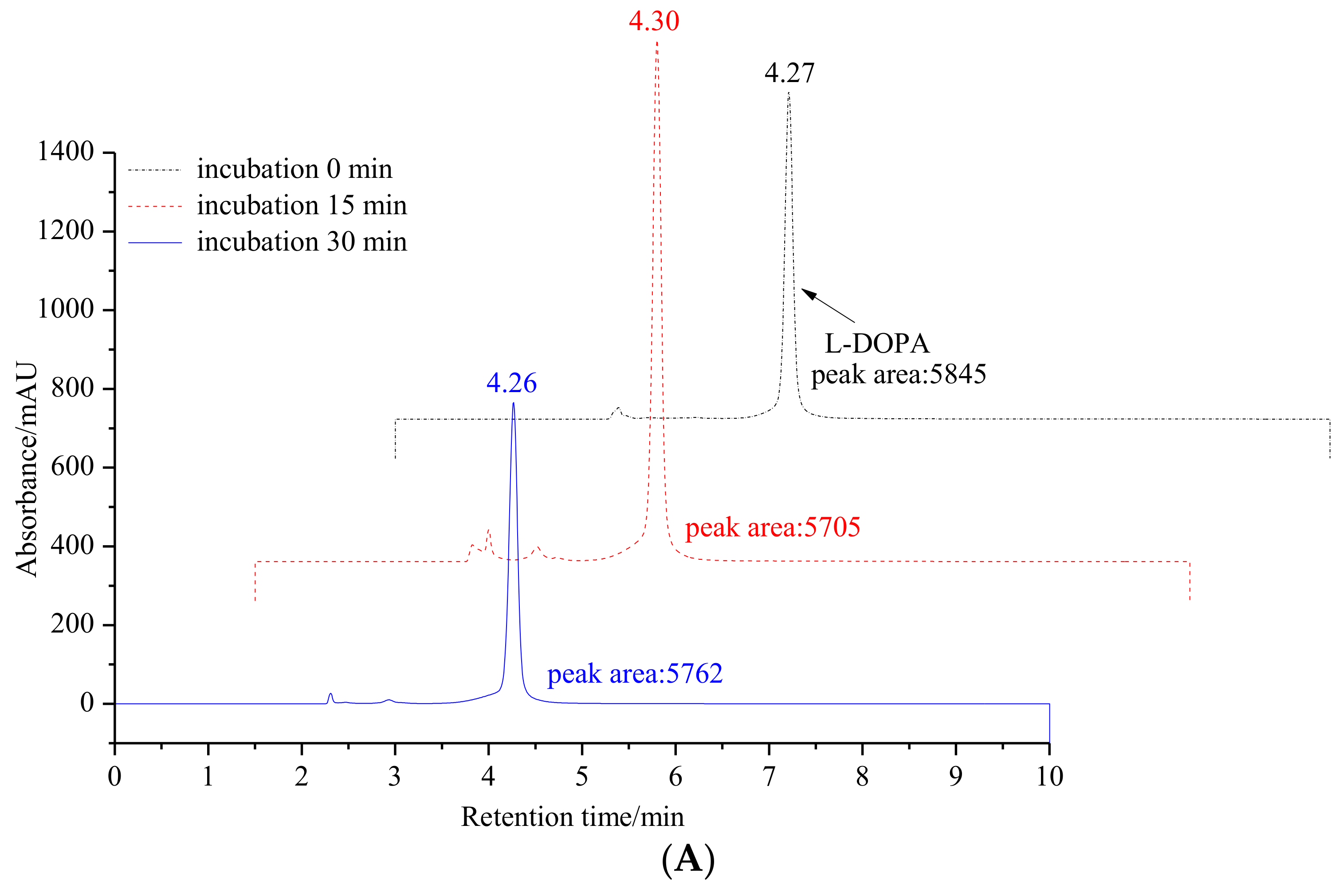
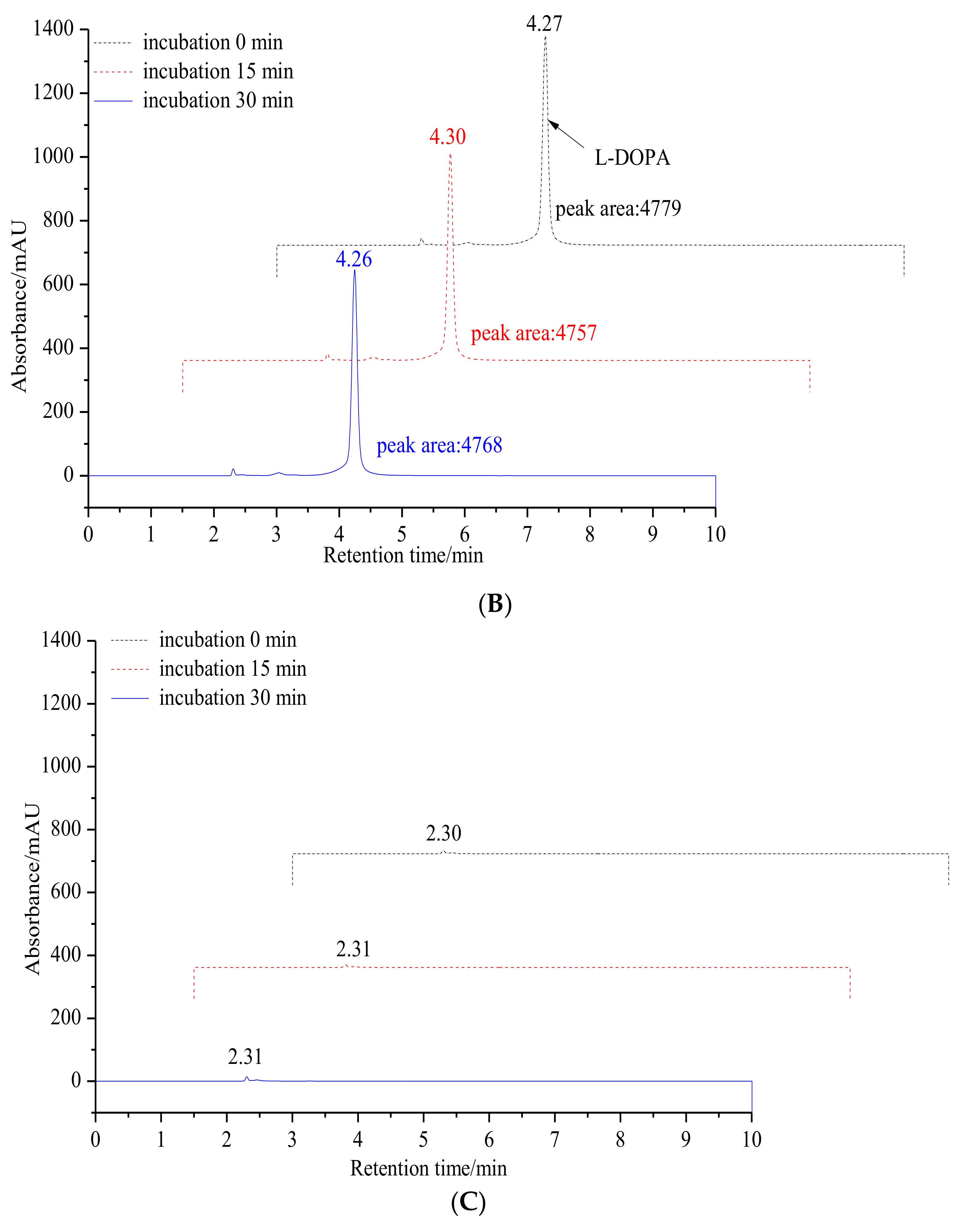
2.1.1. Inhibitory Effect of 6′-O-Caffeoylarbutin on Oxidation of L-DOPA Catalyzed by mTyr
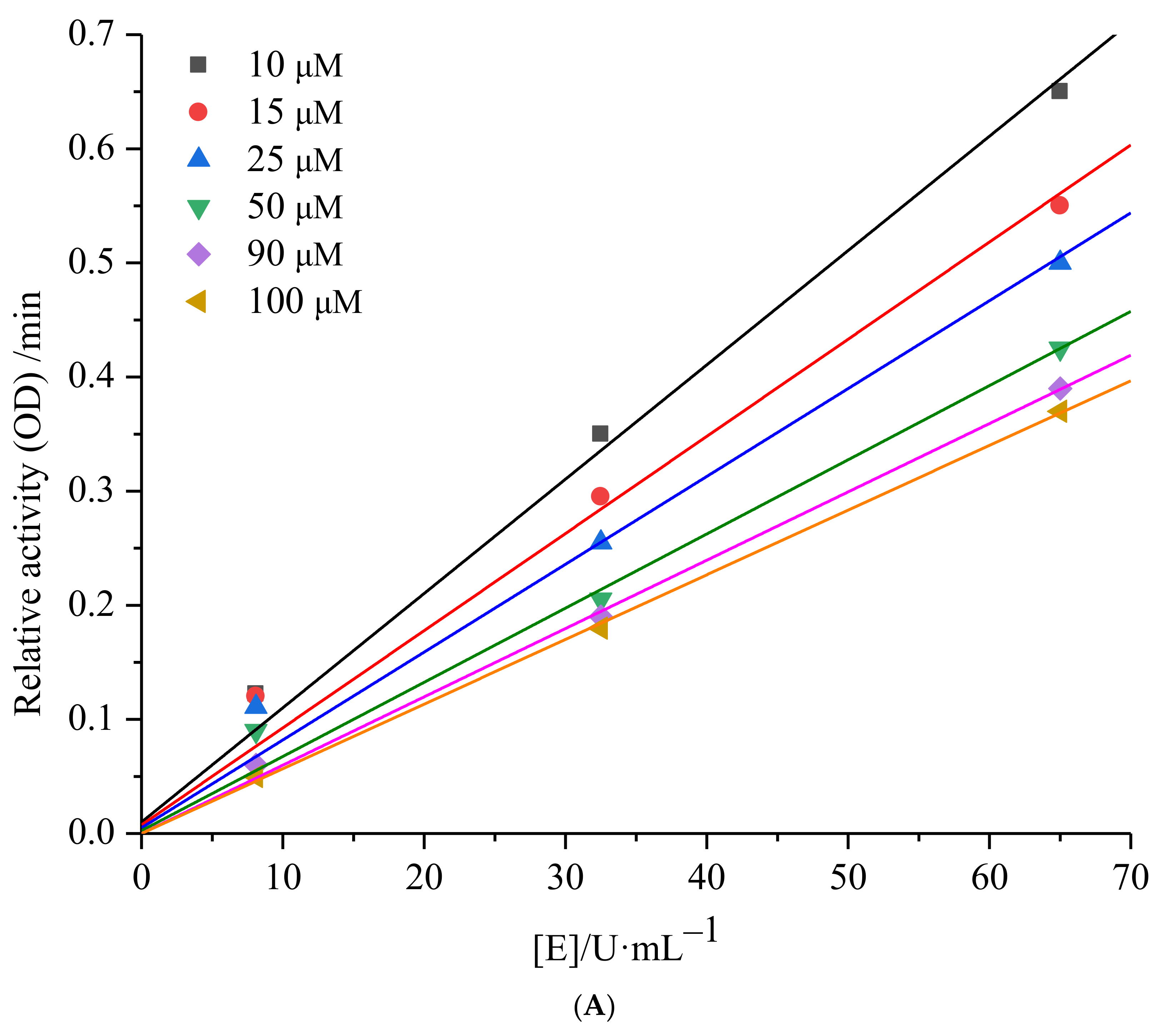
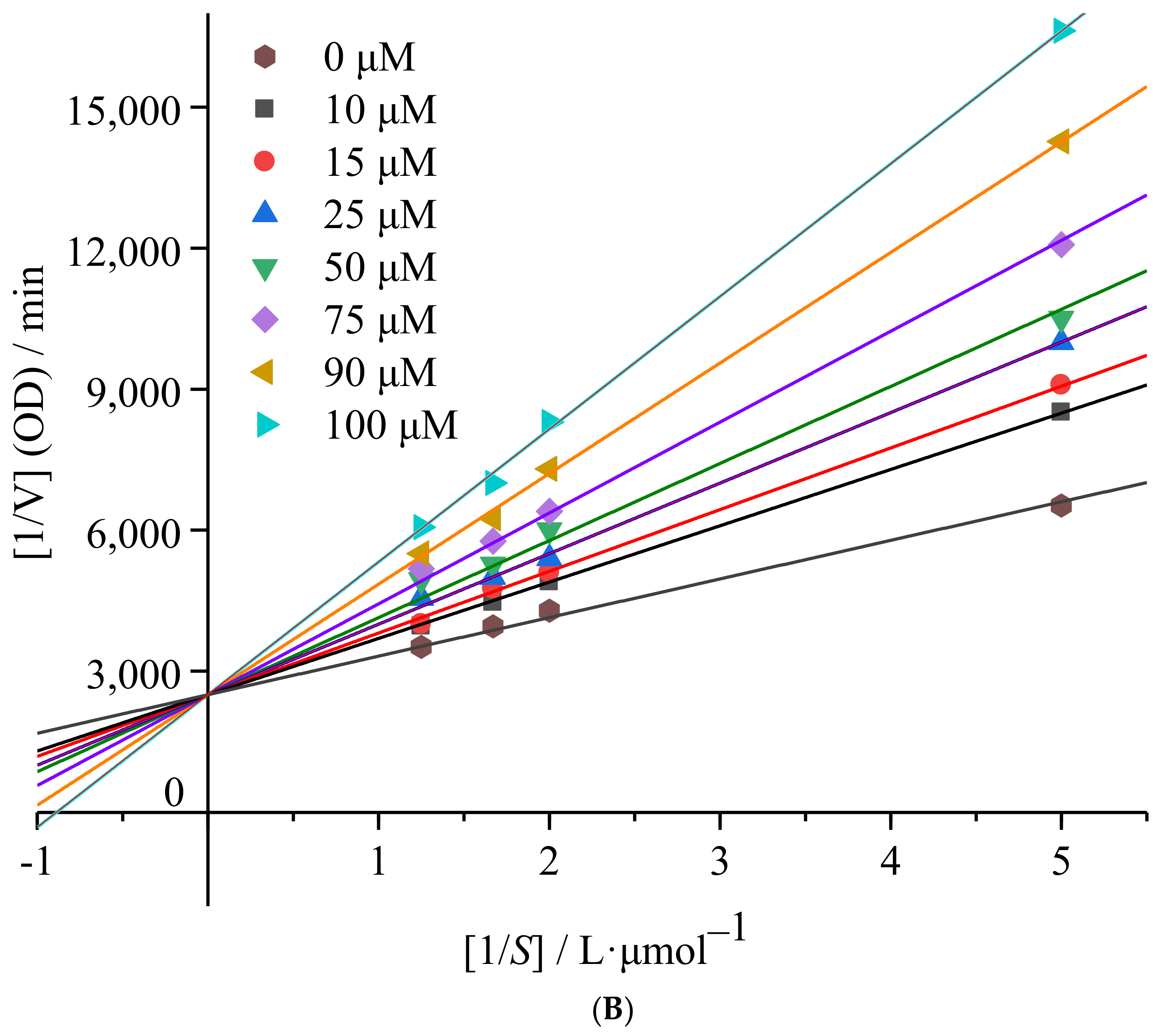
2.1.2. mTyr Inhibition Kinetics of 6′-O-Caffeoylarbutin
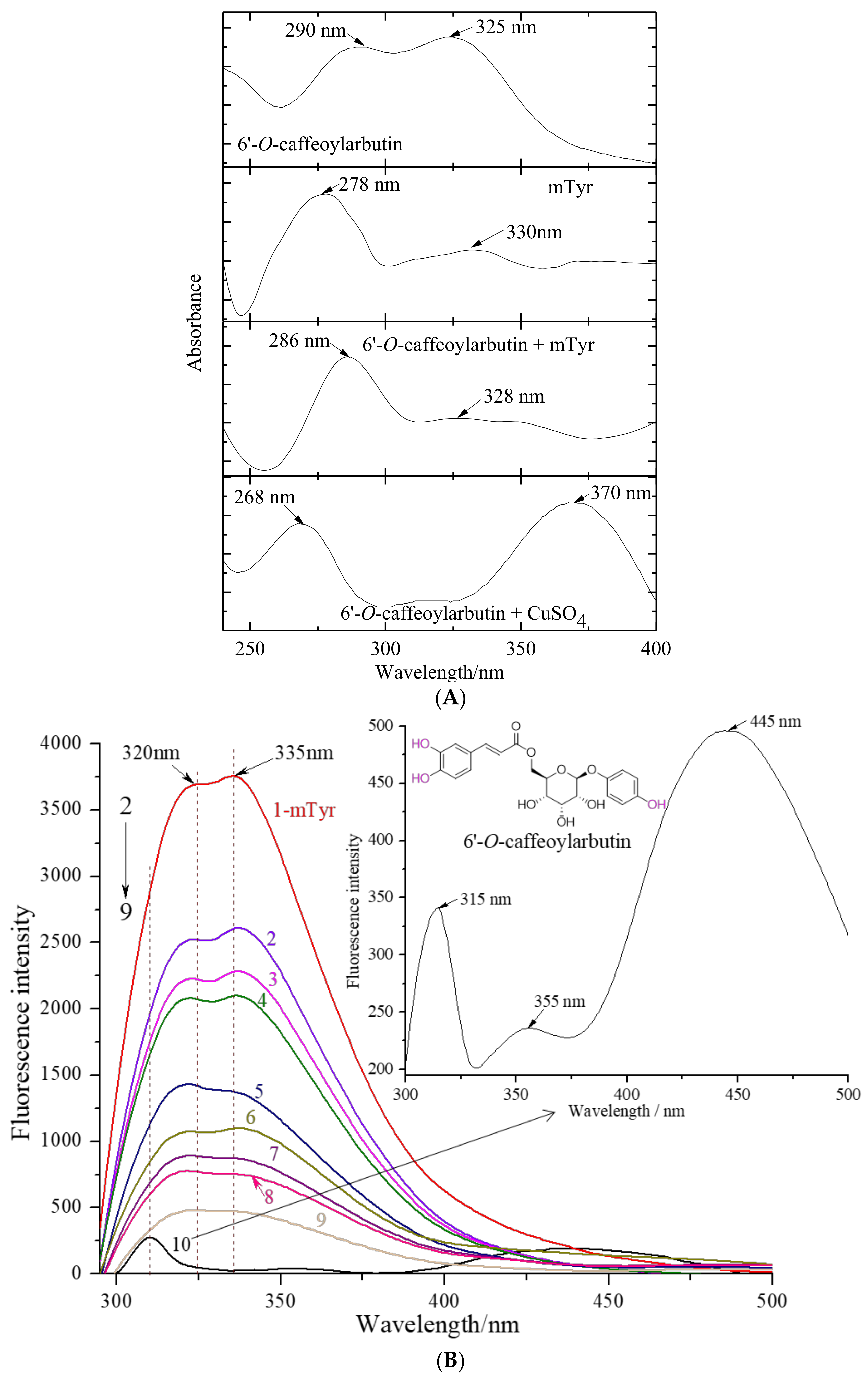
2.1.3. The Chelation of 6′-O-Caffeoylarbutin with Copper (II) Ion in mTyr
2.1.4. The Fluorescence Quenching Effect of 6′-O-Caffeoylarbutin on mTyr
2.2. Computational Simulation of Interaction between 6′-O-Caffeoylarbutin and mTyr
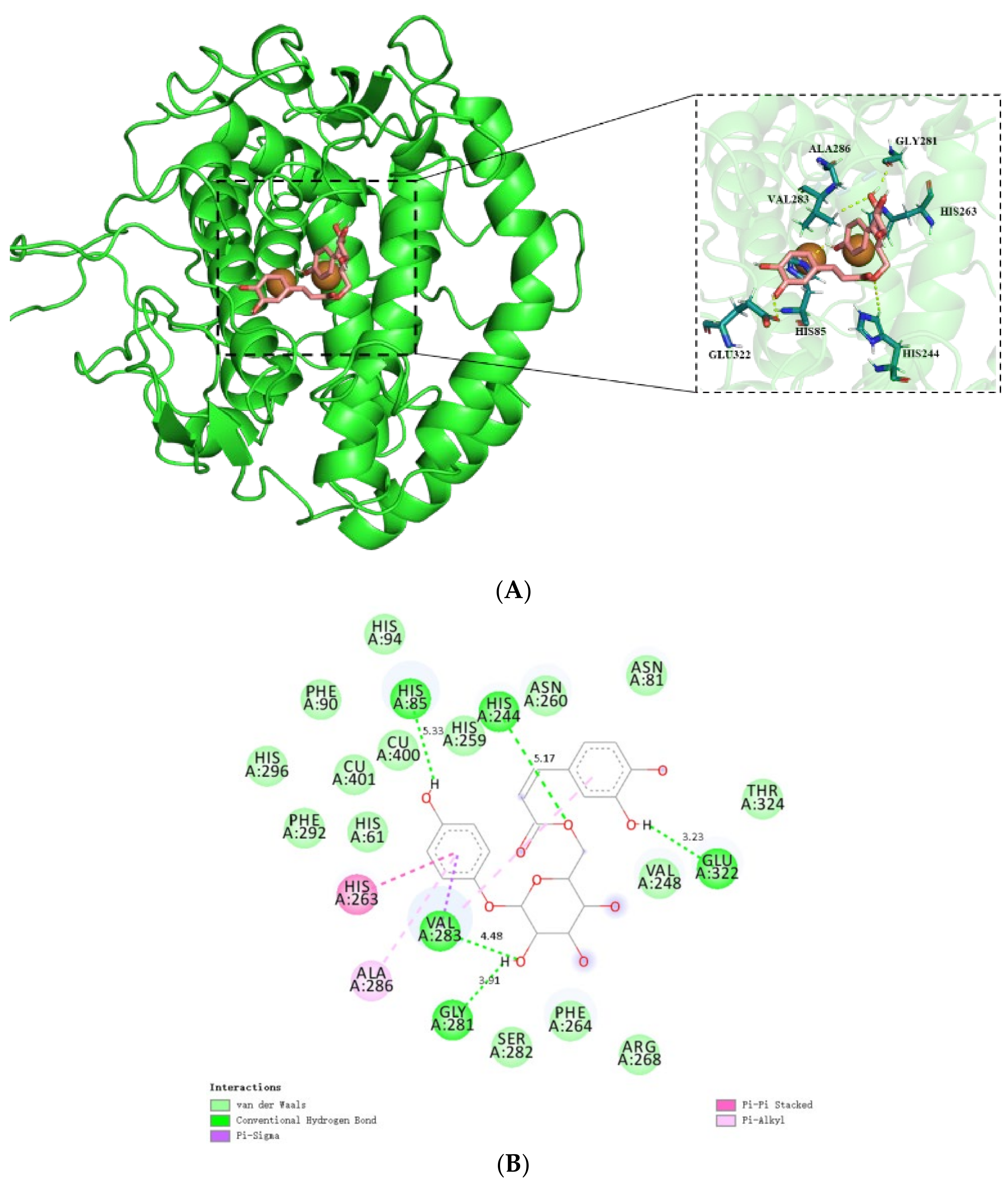
2.2.1. Molecular Docking
2.2.2. MD Simulation
2.3. Safety Evaluation
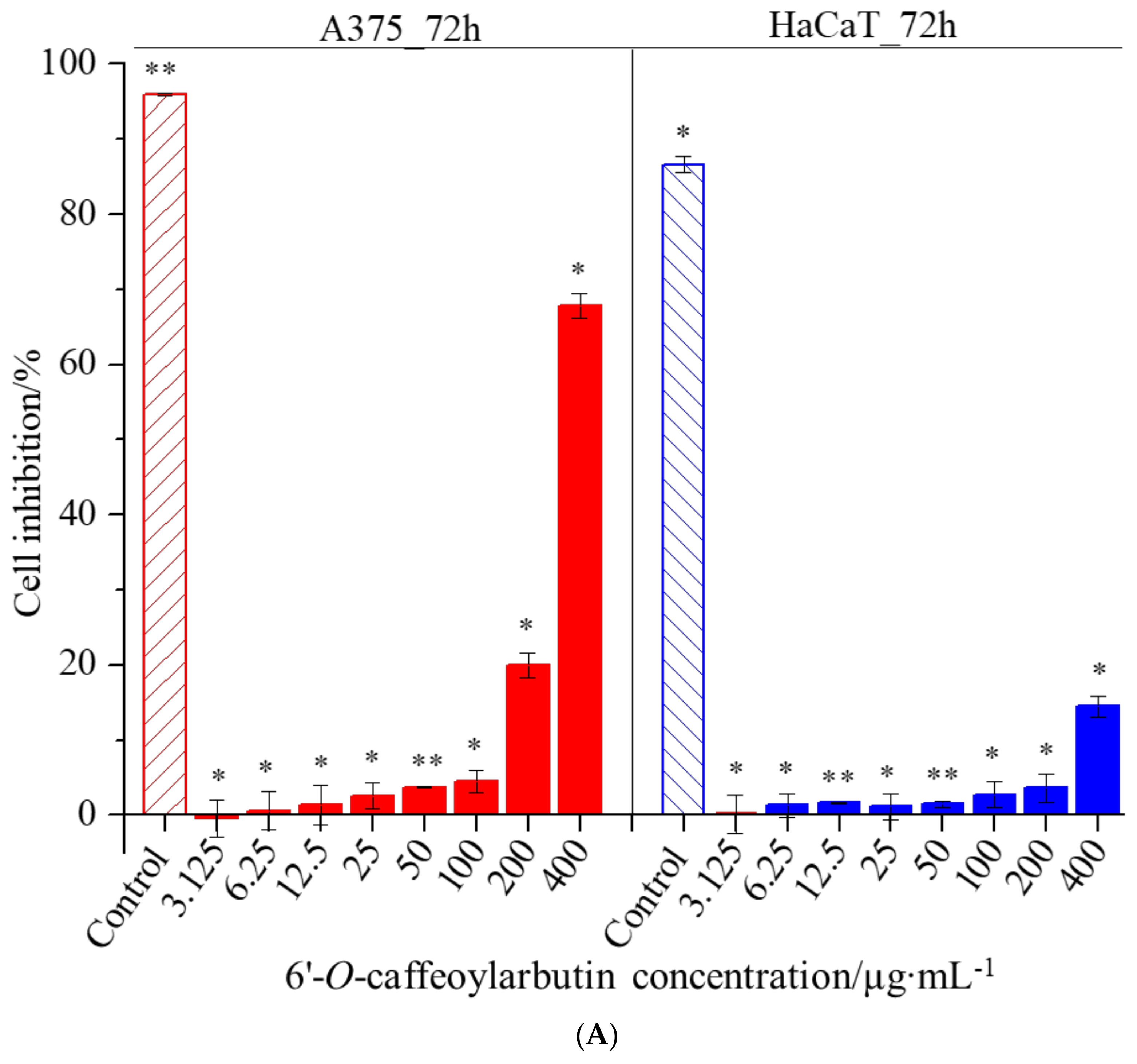
2.3.1. Cellular Assays
2.3.2. Acute Oral Toxicity Assays
2.4. A Preliminary Investigation of the Browning Inhibitory Effect of 6′-O-Caffeoylarbutin on Apple Juice
3. Materials and Methods
3.1. Reagents
3.2. mTyr Inhibition Mechanism of 6′-O-Caffeoylarbutin
3.2.1. Determination of Inhibitory Effect of 6′-O-Caffeoylarbutin on Oxidation of L-DOPA Catalyzed by mTyr
3.2.2. Determination of mTyr Inhibition Kinetics of 6′-O-Caffeoylarbutin
3.2.3. Determination of the Chelation of 6′-O-Caffeoylarbutin with Copper (II) Ion in mTyr
3.2.4. Determination of the Fluorescence Quenching Effect of 6′-O-Caffeoylarbutin on mTyr
3.3. Computational Simulation of Interaction between 6′-O-Caffeoylarbutin and mTyr
3.3.1. Molecular Docking
3.3.2. MD Simulation
3.4. Safety Evaluation
3.4.1. Cellular Assays
3.4.2. Acute Oral Toxicity Assays
3.5. A Preliminary Investigation of the Browning Inhibitory Effect of 6′-O-Caffeoylarbutin on Apple Juice
3.6. Data Processing and Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Xiong, B.; Xing, S.; Chen, Y.; Liao, Q.; Mo, J.; Chen, Y.; Li, Q.; Sun, H. Medicinal prospects of targeting tyrosinase: A feature review. Curr. Med. Chem. 2023, 30, 2638–2671. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Recent advances of polyphenol oxidases in plants. Molecules 2023, 28, 2158. [Google Scholar] [CrossRef] [PubMed]
- Zolghadri, S.; Bahrami, A.; Hassan Khan, M.T.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.L.; Li, W.Y.; Gu, Z.Y.; Wang, L.T.; Guo, L.; Ma, S.B.; Li, C.Y.; Sun, J.H.; Han, B.Q.; Chang, J. Recent advances and progress on melanin: From source to application. Int. J. Mol. Sci. 2023, 24, 4360. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.M.; Kwon, E.B.; Lee, B.; Kim, C.Y. Recent trends in controlling the enzymatic browning of fruit and vegetable products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Meng, Z.; Dong, T.; Fan, X.; Wang, Q. Enzymatic browning and polyphenol oxidase control strategies. Curr. Opin. Biotech. 2023, 81, 102921. [Google Scholar] [CrossRef] [PubMed]
- Panzella, L.; Napolitano, A. Natural and bioinspired phenolic compounds as tyrosinase inhibitors for the treatment of skin hyperpigmentation: Recent advances. Cosmetics 2019, 6, 57. [Google Scholar] [CrossRef]
- Tian, J.L.; Liu, T.L.; Xue, J.J.; Hong, W.; Niu, S.L. Flavanoids derivatives from the root bark of Broussonetia papyrifera as a tyrosinase inhibitor. Ind. Crop. Prod. 2019, 138, 111445. [Google Scholar] [CrossRef]
- Taruscio, T.G.; Barney, D.L.; Exon, J. Content and profile of flavanoid and phenolic acid compounds in conjunction with the antioxidant capacity for a variety of northwest Vaccinium berries. J. Agric. Food Chem. 2004, 52, 3169–3176. [Google Scholar] [CrossRef]
- Li, N.; Zeng, W.L.; Luo, X.L.; Yang, C.R.; Zhang, Y.J.; Ding, Y.; Zhao, P. A new arbutin derivative from the leaves of Vaccinium dunalianum wight. Nat. Prod. Res. 2017, 32, 65–70. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, L.; Wang, Y.; Zhao, T.; Khan, A.; Wang, Y.; Cao, J.; Cheng, G. Protective effect of Que Zui tea hot-water and aqueous ethanol extract against acetaminophen-induced liver injury in mice via inhibition of oxidative stress, inflammation, and apoptosis. Food Funct. 2021, 12, 2468–2480. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Tanaka, T.; Hirabayashi, K.; Zhang, Y.J.; Yang, C.R.; Kouno, I. Caffeoyl arbutin and related compounds from the buds of Vaccinium dunalianum. Phytochemistry 2008, 69, 3087–3094. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lao, Q.C.; Zhao, P.; Zhu, X.Y.; Zhu, H.T.; Luo, X.L.; Yang, C.R.; He, J.H.; Li, C.Q.; Zhang, Y.J. 6′-O-Caffeoylarbutin inhibits melanogenesis in zebrafish. Nat. Prod. Res. 2014, 28, 932–934. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Su, W.C.; Wang, Q.; Lin, Y.F.; Shi, Y. Antityrosinase and antioxidant activities of guanidine compounds and effect of guanylthiourea on melanogenesis. Process Biochem. 2019, 85, 84–96. [Google Scholar] [CrossRef]
- Wang, W.; Gao, Y.; Wang, W.W.; Zhang, J.Y.; Yin, J.F.; Le, T.; Xue, J.J.; Engelhardt, U.H.; Jiang, H.Y. Kojic acid showed consistent inhibitory activity on tyrosinase from mushroom and in cultured b16f10 cells compared with arbutins. Antioxidants 2022, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Funayama, M.; Arakawa, H.; Yamamoto, R.; Nishino, T.; Shin, T.; Murao, S.; Murao, S. Effects of α-Arbutin and β-Arbutin on Activity of Tyrosinases from Mushroom and Mouse Melanoma. Biosci. Biotech. Biochem. 1995, 59, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Zhang, Y.T.; Yan, G.J.; Chen, T.Y. Dissection of Interaction Kinetics through Single-Molecule Interaction Simulation. Anal. Chem. 2020, 92, 11582–11589. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.E. Basic principles of fluorescence and energy transfer applied to real-time PCR. Mol. Biotechnol. 2010, 44, 168–176. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, C. Excited state energy transfer in metal-organic frameworks. Adv. Mater. 2021, 33, e2005819. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, G.; He, Y.; Wang, J.J.; Zhao, Y. Tyrosinase inhibitory mechanism and anti-browning properties of novel kojic acid derivatives bearing aromatic aldehyde moiety. Curr. Res. Food Sci. 2022, 6, 100421. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jang, D.H.; Lee, K.W.; Kim, K.D.; Shah, A.B.; Zhumanova, K.; Park, K.H. Tyrosinase inhibition and kinetic details of puerol A having but-2-enolide structure from Amorpha fruticosa. Molecules 2020, 25, 2344. [Google Scholar] [CrossRef] [PubMed]
- Khade, B.S.; Mathe, V.L.; Dongre, P.M. α-amylase binding to thermal plasma synthesized zinc oxide nanosheets: A fluorescence study. J. Lumin. 2017, 187, 449–456. [Google Scholar] [CrossRef]
- Kong, F.; Kang, S.; Tian, J.; Li, M.; Yue, X. Interaction of xylitol with whey proteins: Multi-spectroscopic techniques and docking studies. Food Chem. 2020, 326, 126804. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Sun, X.; Ni, H.; Du, X.P.; Li, Q. Identification and characterization of the tyrosinase inhibitory activity of caffeine from camellia pollen. J. Agric. Food Chem. 2019, 67, 12741–12751. [Google Scholar] [CrossRef]
- Guterres, H.; Im, W. Improving protein-ligand docking results with high-throughput molecular dynamics simulations. J. Chem. Inf. Model. 2020, 60, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Haider, S.; Barakat, A.; Ul-Haq, Z. Discovery of potential chemical probe as inhibitors of CXCL12 using ligand-based virtual screening and molecular dynamic simulation. Molecules 2020, 25, 4829. [Google Scholar] [CrossRef]
- Pantiora, P.; Furlan, V.; Matiadis, D.; Mavroidi, B.; Perperopoulou, F.; Papageorgiou, A.C.; Sagnou, M.; Bren, U.; Pelecanou, M.; Labrou, N.E. Monocarbonyl Curcumin Analogues as Potent Inhibitors against Human Glutathione Transferase P1-1. Antioxidants 2023, 12, 63. [Google Scholar] [CrossRef]
- Furlan, V.; Bren, U. Insight into inhibitory mechanism of pde4d by dietary polyphenols using molecular dynamics simulations and free energy calculations. Biomolecules 2021, 11, 479. [Google Scholar] [CrossRef]
- Bhat, I.A.; Kausar, T.; Dar, A.A.; Nayeem, S.M.; Akram, M. Exclusive behaviour of asymmetric zwitterionic gemini surfactants towards lysozyme. J. Mol. Liq. 2021, 336, 116583. [Google Scholar] [CrossRef]
- Yang, X.Q.; Wang, W.B.; Jiang, Q.; Xie, S.D.; Zhao, P.; Liu, Z.; Zhu, G.L.; Xu, J.M.; Wang, J.Y.; Li, Y.H. Subcritical water extraction of phenolic compounds from Vaccinium dunalianum wight leaves and their antioxidant and tyrosinase inhibitory activities in vitro. Chem. Biodivers. 2023, 20, e202201099. [Google Scholar] [CrossRef]
- Garcia-Jimenez, A.; Teruel-Puche, J.A.; Berna, J.; Rodriguez-Lopez, J.N.; Tudela, J.; Garcia-Canovas, F. Action of tyrosinase on alpha and beta-arbutin: A kinetic study. PLoS ONE 2017, 12, e0177330. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.L.; Lim, G.T.; Yin, S.J.; Lee, J.Y.; Si, Y.X.; Yang, J.M.; Park, Y.D.; Qian, G.Y. The inhibitory effect of pyrogallol on tyrosinase activity and structure: Integration study of inhibition kinetics with molecular dynamics simulation. Int. J. Biol. Macromol. 2019, 121, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wang, C.; Shang, C.; You, X.; Zhang, L.; Liu, W. Integrated study of the mechanism of tyrosinase inhibition by baicalein using kinetic, multispectroscopic and computational simulation analyses. Int. J. Biol. Macromol. 2018, 118, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, A.; Mohammadsadeghi, N.; Mohammadi, F.; Saadati, F.; Nikfard, S. Evaluation of inhibitory effects of some novel phenolic derivatives on the mushroom tyrosinase activity: Insights from spectroscopic analyses, molecular docking and in vitro assays. Food Chem. 2022, 387, 132938. [Google Scholar] [CrossRef]
- Shang, C.; Zhang, Y.; You, X.; Guo, N.; Wang, Y.; Fan, Y.; Liu, W. The effect of 7, 8, 4′-trihydroxyflavone on tyrosinase activity and conformation: Spectroscopy and docking studies. Luminescence 2018, 33, 681–691. [Google Scholar] [CrossRef]
- Gou, L.; Lee, J.H.; Yang, J.M.; Park, Y.D.; Zhou, H.M.; Zhan, Y.; Lv, Z.R. The effect of alpha-ketoglutaric acid on tyrosinase activity and conformation: Kinetics and molecular dynamics simulation study. Int. J. Biol. Macromol. 2017, 105, 1654–1662. [Google Scholar] [CrossRef]
- Yang, B.F. Pharmacology; People’s Medical Publishing House: Beijing, China, 2003; pp. 1–27. [Google Scholar]
- Klimczak, I.; Gliszczyńska-Świgło, A. Green tea extract as an anti-browning agent for cloudy apple juice. J. Sci. Food Agric. 2017, 97, 1420–1426. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Final Dose (mg·kg−1) | Toxic Symptoms (♀/♂) | |||
|---|---|---|---|---|---|
| Reduced Activities | Perianal Uncleanliness | Prone State | Shortness of Breath | ||
| Vehicle | - | 0 | 0 | 0 | 0 |
| CA a -1 | 28,056 | 8 (2/6) | 0 | 2 (0/2) | 4 (2/2) |
| CA-2 | 23,848 | 1 (0/1) | 0 | 0 | 0 |
| CA-3 | 20,272 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, D.; Fu, W.; Yuan, T.; Han, K.; Lv, Y.; Wang, Q.; Jiang, Q.; Zhang, Y.; Zhu, G.; Xu, J.; et al. 6′-O-Caffeoylarbutin from Quezui Tea: A Highly Effective and Safe Tyrosinase Inhibitor. Int. J. Mol. Sci. 2024, 25, 972. https://doi.org/10.3390/ijms25020972
Xie D, Fu W, Yuan T, Han K, Lv Y, Wang Q, Jiang Q, Zhang Y, Zhu G, Xu J, et al. 6′-O-Caffeoylarbutin from Quezui Tea: A Highly Effective and Safe Tyrosinase Inhibitor. International Journal of Molecular Sciences. 2024; 25(2):972. https://doi.org/10.3390/ijms25020972
Chicago/Turabian StyleXie, Dong, Wengan Fu, Tiantian Yuan, Kangjia Han, Yuxiu Lv, Qi Wang, Qian Jiang, Yingjun Zhang, Guolei Zhu, Junming Xu, and et al. 2024. "6′-O-Caffeoylarbutin from Quezui Tea: A Highly Effective and Safe Tyrosinase Inhibitor" International Journal of Molecular Sciences 25, no. 2: 972. https://doi.org/10.3390/ijms25020972
APA StyleXie, D., Fu, W., Yuan, T., Han, K., Lv, Y., Wang, Q., Jiang, Q., Zhang, Y., Zhu, G., Xu, J., Zhao, P., & Yang, X. (2024). 6′-O-Caffeoylarbutin from Quezui Tea: A Highly Effective and Safe Tyrosinase Inhibitor. International Journal of Molecular Sciences, 25(2), 972. https://doi.org/10.3390/ijms25020972