
Posttranslational Acylations of the Rat Brain Transketolase Discriminate the Enzyme Responses to Inhibitors of ThDP-Dependent Enzymes or Thiamine Transport
 , and
, and 
Abstract
1. Introduction
2. Results
2.1. Effect of Inhibitors of Thiamine-Dependent Metabolism on TKT Acylations in the Rat Brain
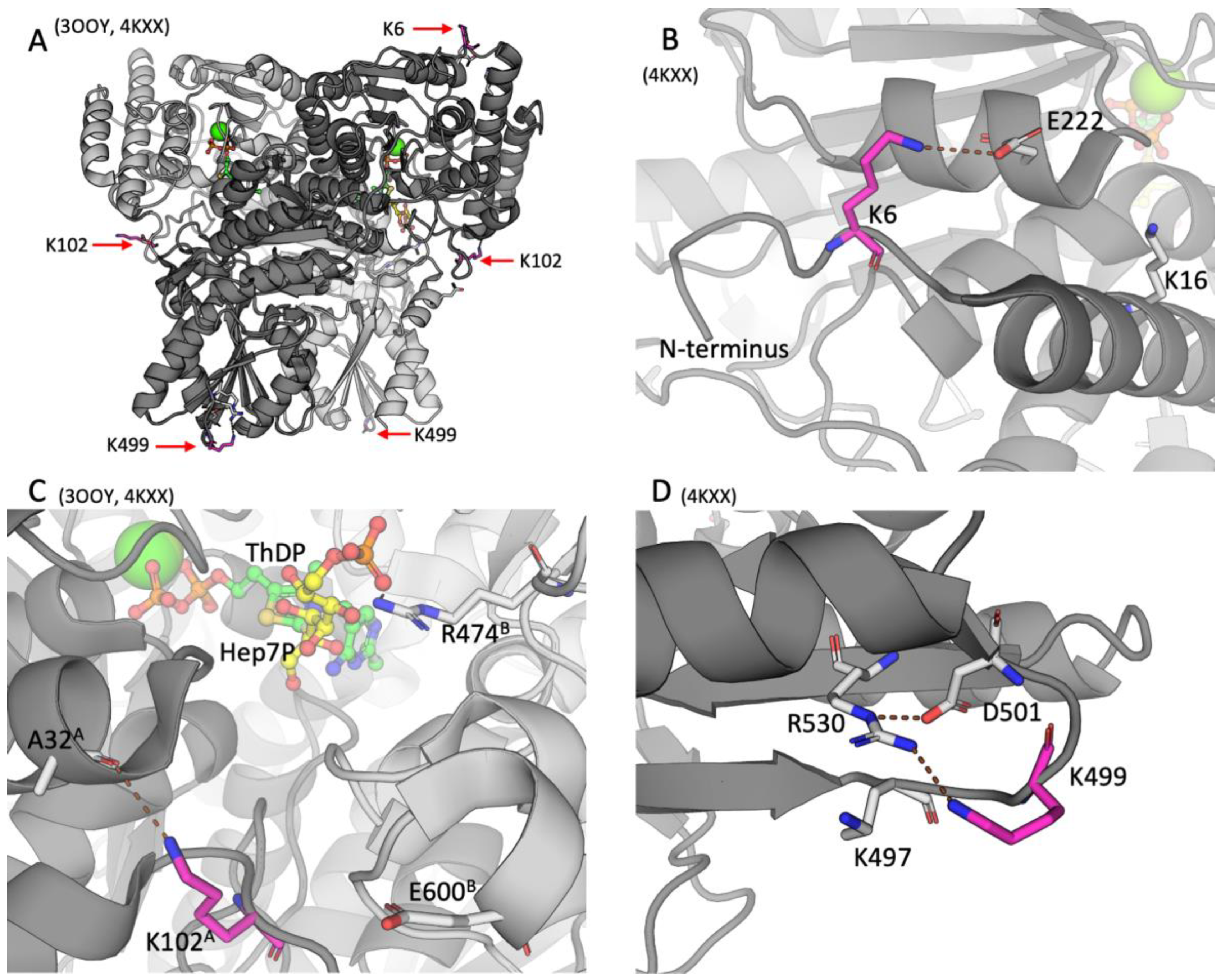
2.2. Positions of the Brain TKT Acylation Sites in the Resolved Structures of Mammalian TKT
2.3. Correlation Analysis of the Interplay between the Levels of TKT Expression, Activity, Acylations, and Sirtuin 5
3. Discussion
3.1. The Brain TKT Regulation by Malonylation
3.2. The Brain TKT Regulation by Acetylation
3.3. Biological Implications of TKT Acylation
4. Materials and Methods
4.1. Reagents
4.2. Animal Experiments
4.3. Homogenization of Rat Brain Tissue
4.4. TKT Activity Assay

4.5. Procedure of LC-MS/MS
4.6. Quantification of the TKT Acylations, Expression, and Sirtuin 5 Protein Level
4.7. Structural Visualization
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TKT | transketolase |
| ThDP | thiamine diphosphate |
| PPP | pentose phosphate pathway |
| OT | oxythiamine |
| M + A | metformin and amprolium |
| MS | mass spectrometry |
References
- Wang, H.L.; Chen, Y.; Wang, Y.Q.; Tao, E.W.; Tan, J.; Liu, Q.Q.; Li, C.M.; Tong, X.M.; Gao, Q.Y.; Hong, J.; et al. Sirtuin5 protects colorectal cancer from DNA damage by keeping nucleotide availability. Nat. Commun. 2022, 13, 6121. [Google Scholar] [CrossRef] [PubMed]
- Nishida, Y.; Rardin, M.J.; Carrico, C.; He, W.; Sahu, A.K.; Gut, P.; Najjar, R.; Fitch, M.; Hellerstein, M.; Gibson, B.W.; et al. SIRT5 Regulates both Cytosolic and Mitochondrial Protein Malonylation with Glycolysis as a Major Target. Mol. Cell 2015, 59, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Boros, L.G.; Puigjaner, J.; Cascante, M.; Lee, W.N.; Brandes, J.L.; Bassilian, S.; Yusuf, F.I.; Williams, R.D.; Muscarella, P.; Melvin, W.S.; et al. Oxythiamine and dehydroepiandrosterone inhibit the nonoxidative synthesis of ribose and tumor cell proliferation. Cancer Res. 1997, 57, 4242–4248. [Google Scholar] [PubMed]
- Pinson, B.; Moenner, M.; Saint-Marc, C.; Granger-Farbos, A.; Daignan-Fornier, B. On-demand utilization of phosphoribosyl pyrophosphate by downstream anabolic pathways. J. Biol. Chem. 2023, 299, 105011. [Google Scholar] [CrossRef]
- Lindqvist, Y.; Schneider, G.; Ermler, U.; Sundstrom, M. Three-dimensional structure of transketolase, a thiamine diphosphate dependent enzyme, at 2.5 A resolution. EMBO J. 1992, 11, 2373–2379. [Google Scholar] [CrossRef] [PubMed]
- Blass, J.P.; Gibson, G.E. Abnormality of a thiamine-requiring enzyme in patients with Wernicke-Korsakoff syndrome. N. Engl. J. Med. 1977, 297, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.B.; Svoronos, S.; Ghazanfari, A.; Martin, P.R.; Fisher, A.; Roecklein, B.; Rodbard, D.; Staton, R.; Behar, D.; Berg, C.J.; et al. Transketolase abnormality in cultured fibroblasts from familial chronic alcoholic men and their male offspring. J. Clin. Investig. 1987, 79, 1039–1043. [Google Scholar] [CrossRef]
- Martin, P.R.; McCool, B.A.; Singleton, C.K. Molecular genetics of transketolase in the pathogenesis of the Wernicke-Korsakoff syndrome. Metab. Brain Dis. 1995, 10, 45–55. [Google Scholar] [CrossRef]
- Ou, T.; Yang, W.; Li, W.; Lu, Y.; Dong, Z.; Zhu, H.; Sun, X.; Dong, Z.; Weng, X.; Chang, S.; et al. SIRT5 deficiency enhances the proliferative and therapeutic capacities of adipose-derived mesenchymal stem cells via metabolic switching. Clin. Transl. Med. 2020, 10, e172. [Google Scholar] [CrossRef]
- Pandya, N.J.; Meier, S.; Tyanova, S.; Terrigno, M.; Wang, C.; Punt, A.M.; Mientjes, E.J.; Vautheny, A.; Distel, B.; Kremer, T.; et al. A cross-species spatiotemporal proteomic analysis identifies UBE3A-dependent signaling pathways and targets. Mol. Psychiatry 2022, 27, 2590–2601. [Google Scholar] [CrossRef]
- Pinson, A.; Xing, L.; Namba, T.; Kalebic, N.; Peters, J.; Oegema, C.E.; Traikov, S.; Reppe, K.; Riesenberg, S.; Maricic, T.; et al. Human TKTL1 implies greater neurogenesis in frontal neocortex of modern humans than Neanderthals. Science 2022, 377, eabl6422. [Google Scholar] [CrossRef] [PubMed]
- Bunik, V.I.; Tylicki, A.; Lukashev, N.V. Thiamin diphosphate-dependent enzymes: From enzymology to metabolic regulation, drug design and disease models. FEBS J. 2013, 280, 6412–6442. [Google Scholar] [CrossRef]
- Zhang, F.; Masania, J.; Anwar, A.; Xue, M.; Zehnder, D.; Kanji, H.; Rabbani, N.; Thornalley, P.J. The uremic toxin oxythiamine causes functional thiamine deficiency in end-stage renal disease by inhibiting transketolase activity. Kidney Int. 2016, 90, 396–403. [Google Scholar] [CrossRef]
- Golan, O.; Dyer, R.; Sinclair, G.; Blydt-Hansen, T. Investigating oxythiamine levels in children undergoing kidney transplantation and the risk of immediate post-operative metabolic and hemodynamic decompensation. Pediatr. Nephrol. 2021, 36, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Aleshin, V.A.; Mkrtchyan, G.V.; Bunik, V.I. Mechanisms of Non-coenzyme Action of Thiamine: Protein Targets and Medical Significance. Biochem. Biokhimiia 2019, 84, 829–850. [Google Scholar] [CrossRef]
- Rogers, E.F. General discussion of antithiamin compounds and thiamin antagonists. Ann. N. Y. Acad. Sci. 1982, 378, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Al-Otaibi, T.; Hawsah, M.A.; Alojayri, G.; Mares, M.M.; Aljawdah, H.M.A.; Maodaa, S.N.; Al-Shaebi, E.M.; Dkhil, M.A.; Thagfan, F.A.; Al-Quraishy, S.; et al. In vivo anticoccidial, antioxidant, and anti-inflammatory activities of avocado fruit, Persea americana (Lauraceae), against Eimeria papillata infection. Parasitol. Int. 2023, 95, 102741. [Google Scholar] [CrossRef]
- Bunik, V.I.; Aleshin, V.A. Analysis of the Protein Binding Sites for Thiamin and Its Derivatives to Elucidate the Molecular Mechanisms of the Noncoenzyme Action of Thiamin (Vitamin B1). Stud. Nat. Prod. Chem. 2017, 53, 375–429. [Google Scholar] [CrossRef]
- Aleshin, V.A.; Mkrtchyan, G.V.; Kaehne, T.; Graf, A.V.; Maslova, M.V.; Bunik, V.I. Diurnal regulation of the function of the rat brain glutamate dehydrogenase by acetylation and its dependence on thiamine administration. J. Neurochem. 2020, 153, 80–102. [Google Scholar] [CrossRef]
- Aleshin, V.A.; Artiukhov, A.V.; Kaehne, T.; Graf, A.V.; Bunik, V.I. Daytime Dependence of the Activity of the Rat Brain Pyruvate Dehydrogenase Corresponds to the Mitochondrial Sirtuin 3 Level and Acetylation of Brain Proteins, All Regulated by Thiamine Administration Decreasing Phosphorylation of PDHA Ser293. Int. J. Mol. Sci. 2021, 22, 8006. [Google Scholar] [CrossRef]
- Bunik, V.; Aleshin, V.; Nogues, I.; Kahne, T.; Parroni, A.; Contestabile, R.; Salvo, M.L.; Graf, A.; Tramonti, A. Thiamine-dependent regulation of mammalian brain pyridoxal kinase in vitro and in vivo. J. Neurochem. 2022, 161, 20–39. [Google Scholar] [CrossRef] [PubMed]
- Zavileyskiy, L.G.; Aleshin, V.A.; Kaehne, T.; Karlina, I.S.; Artiukhov, A.V.; Maslova, M.V.; Graf, A.V.; Bunik, V.I. The Brain Protein Acylation System Responds to Seizures in the Rat Model of PTZ-Induced Epilepsy. Int. J. Mol. Sci. 2022, 23, 12302. [Google Scholar] [CrossRef] [PubMed]
- Ludtke, S.; Neumann, P.; Erixon, K.M.; Leeper, F.; Kluger, R.; Ficner, R.; Tittmann, K. Sub-angstrom-resolution crystallography reveals physical distortions that enhance reactivity of a covalent enzymatic intermediate. Nat. Chem. 2013, 5, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Artiukhov, A.V.; Graf, A.V.; Kazantsev, A.V.; Boyko, A.I.; Aleshin, V.A.; Ksenofontov, A.L.; Bunik, V.I. Increasing Inhibition of the Rat Brain 2-Oxoglutarate Dehydrogenase Decreases Glutathione Redox State, Elevating Anxiety and Perturbing Stress Adaptation. Pharmaceuticals 2022, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Aleshin, V.A.; Zhou, X.; Krishnan, S.; Karlsson, A.; Bunik, V.I. Interplay Between Thiamine and p53/p21 Axes Affects Antiproliferative Action of Cisplatin in Lung Adenocarcinoma Cells by Changing Metabolism of 2-Oxoglutarate/Glutamate. Front. Genet. 2021, 12, 658446. [Google Scholar] [CrossRef]
- Zolg, D.P.; Wilhelm, M.; Schmidt, T.; Medard, G.; Zerweck, J.; Knaute, T.; Wenschuh, H.; Reimer, U.; Schnatbaum, K.; Kuster, B. ProteomeTools: Systematic Characterization of 21 Post-translational Protein Modifications by Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS) Using Synthetic Peptides. Mol. Cell. Proteom. MCP 2018, 17, 1850–1863. [Google Scholar] [CrossRef]
- Simon, G.M.; Cheng, J.; Gordon, J.I. Quantitative assessment of the impact of the gut microbiota on lysine epsilon-acetylation of host proteins using gnotobiotic mice. Proc. Natl. Acad. Sci. USA 2012, 109, 11133–11138. [Google Scholar] [CrossRef]
- Weinert, B.T.; Scholz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef]
- Akimov, V.; Barrio-Hernandez, I.; Hansen, S.V.F.; Hallenborg, P.; Pedersen, A.K.; Bekker-Jensen, D.B.; Puglia, M.; Christensen, S.D.K.; Vanselow, J.T.; Nielsen, M.M.; et al. UbiSite approach for comprehensive mapping of lysine and N-terminal ubiquitination sites. Nat. Struct. Mol. Biol. 2018, 25, 631–640. [Google Scholar] [CrossRef]
- Lumpkin, R.J.; Gu, H.; Zhu, Y.; Leonard, M.; Ahmad, A.S.; Clauser, K.R.; Meyer, J.G.; Bennett, E.J.; Komives, E.A. Site-specific identification and quantitation of endogenous SUMO modifications under native conditions. Nat. Commun. 2017, 8, 1171. [Google Scholar] [CrossRef]
- Udeshi, N.D.; Svinkina, T.; Mertins, P.; Kuhn, E.; Mani, D.R.; Qiao, J.W.; Carr, S.A. Refined preparation and use of anti-diglycine remnant (K-epsilon-GG) antibody enables routine quantification of 10,000s of ubiquitination sites in single proteomics experiments. Mol. Cell. Proteom. MCP 2013, 12, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.A.; Beli, P.; Weinert, B.T.; Scholz, C.; Kelstrup, C.D.; Young, C.; Nielsen, M.L.; Olsen, J.V.; Brakebusch, C.; Choudhary, C. Proteomic analyses reveal divergent ubiquitylation site patterns in murine tissues. Mol. Cell. Proteom. MCP 2012, 11, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Nikkola, M.; Lindqvist, Y.; Schneider, G. Refined structure of transketolase from Saccharomyces cerevisiae at 2.0 A resolution. J. Mol. Biol. 1994, 238, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Solovjeva, O.N. Isolation and Properties of Noncovalent Complex of Transketolase with RNA. Biochemistry 2002, 67, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Tikhomirova, N.K.; Merchan, A.Y.; Kochetov, G.A. A new form of baker’s yeast transketolase. An enzyme-RNA complex. FEBS Lett. 1990, 274, 27–29. [Google Scholar] [CrossRef]
- Kochetov, G.A.; Nikitushkina, L.I.; Chernov, N.N. A complex of functionally-bound enzymes: Transketolase and glyceraldehydephosphate dehydrogenase. Biochem. Biophys. Res. Commun. 1970, 40, 873–879. [Google Scholar] [CrossRef]
- Wood, T.; Muzariri, C.C.; Malaba, L. Complex formation between transketolase, transaldolase, and glyceraldehyde phosphate dehydrogenase. Int. J. Biochem. 1985, 17, 1109–1115. [Google Scholar] [CrossRef]
- Li, M.; Zhao, X.; Yong, H.; Xu, J.; Qu, P.; Qiao, S.; Hou, P.; Li, Z.; Chu, S.; Zheng, J.; et al. Transketolase promotes colorectal cancer metastasis through regulating AKT phosphorylation. Cell Death Dis. 2022, 13, 99. [Google Scholar] [CrossRef]
- Zecha, J.; Gabriel, W.; Spallek, R.; Chang, Y.C.; Mergner, J.; Wilhelm, M.; Bassermann, F.; Kuster, B. Linking post-translational modifications and protein turnover by site-resolved protein turnover profiling. Nat. Commun. 2022, 13, 165. [Google Scholar] [CrossRef]
- Rikova, K. CST Curation Set: 16030. 2012. Available online: https://www.phosphosite.org/curatedInfoAction.action?record=40643606 (accessed on 23 October 2023).
- Rikova, K. CST Curation Set: 16031. 2012. Available online: https://www.phosphosite.org/curatedInfoAction.action?record=41382100 (accessed on 23 October 2023).
- Lundby, A.; Lage, K.; Weinert, B.T.; Bekker-Jensen, D.B.; Secher, A.; Skovgaard, T.; Kelstrup, C.D.; Dmytriyev, A.; Choudhary, C.; Lundby, C.; et al. Proteomic analysis of lysine acetylation sites in rat tissues reveals organ specificity and subcellular patterns. Cell Rep. 2012, 2, 419–431. [Google Scholar] [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef]
- Sevostyanova, I.; Solovjeva, O.; Selivanov, V.; Kochetov, G. Half-of-the-sites reactivity of transketolase from Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2009, 379, 851–854. [Google Scholar] [CrossRef]
- Kovina, M.V.; Kochetov, G.A. Cooperativity and flexibility of active sites in homodimeric transketolase. FEBS Lett. 1998, 440, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, C.J. An enzyme in the test tube, and a transcription factor in the cell: Moonlighting proteins and cellular factors that affect their behavior. Protein Sci. A Publ. Protein Soc. 2019, 28, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Domain, F.; Bina, X.R.; Levy, S.B. Transketolase A, an enzyme in central metabolism, derepresses the marRAB multiple antibiotic resistance operon of Escherichia coli by interaction with MarR. Mol. Microbiol. 2007, 66, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.S.; Kim, J.G.; Shin, M.H.; Lee, Y.A.; Kong, Y. Comparison of Secretome Profile of Pathogenic and Non-Pathogenic Entamoeba histolytica. Proteomics 2018, 18, e1700341. [Google Scholar] [CrossRef]
- Grundel, A.; Pfeiffer, M.; Jacobs, E.; Dumke, R. Network of Surface-Displayed Glycolytic Enzymes in Mycoplasma pneumoniae and Their Interactions with Human Plasminogen. Infect. Immun. 2015, 84, 666–676. [Google Scholar] [CrossRef]
- Wang, Z.; Qiu, Z.; Hua, S.; Yang, W.; Chen, Y.; Huang, F.; Fan, Y.; Tong, L.; Xu, T.; Tong, X.; et al. Nuclear Tkt promotes ischemic heart failure via the cleaved Parp1/Aif axis. Basic Res. Cardiol. 2022, 117, 18. [Google Scholar] [CrossRef]
- Gubler, C.J.; Johnson, L.R.; Wittorf, J.H. Yeast transketolase (sedoheptulose-7-phosphate:d-glyceraldehyde-3-phosphate dihydroxyacetonetransferase, EC 2.2.1.1) assay of thiamine diphosphate. In Vitamins and Coenzymes; Academic Press: Cambridge, MA, USA, 1970; pp. 120–125. [Google Scholar] [CrossRef]
- Brin, M. Effects of thiamine deficiency and of oxythiamine on rat tissue transketolase. J. Nutr. 1962, 78, 179–183. [Google Scholar] [CrossRef]
- Moraes, J.O.; Rodrigues, S.D.C.; Pereira, L.M.; Medeiros, R.C.N.; de Cordova, C.A.S.; de Cordova, F.M. Amprolium exposure alters mice behavior and metabolism in vivo. Anim. Model Exp. Med. 2018, 1, 272–281. [Google Scholar] [CrossRef]
- Oliveira, W.H.; Nunes, A.K.; Franca, M.E.; Santos, L.A.; Los, D.B.; Rocha, S.W.; Barbosa, K.P.; Rodrigues, G.B.; Peixoto, C.A. Effects of metformin on inflammation and short-term memory in streptozotocin-induced diabetic mice. Brain Res. 2016, 1644, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Kalariya, N.M.; Shoeb, M.; Ansari, N.H.; Srivastava, S.K.; Ramana, K.V. Antidiabetic drug metformin suppresses endotoxin-induced uveitis in rats. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3431–3440. [Google Scholar] [CrossRef] [PubMed]
- de la Haba, G.; Leder, I.G.; Racker, E. Crystalline Transketolase from Bakers’ Yeast: Isolation and Properties. J. Biol. Chem. 1955, 214, 409–426. [Google Scholar] [CrossRef] [PubMed]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef]
- Motulsky, H.J.; Brown, R.E. Detecting outliers when fitting data with nonlinear regression—A new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef]
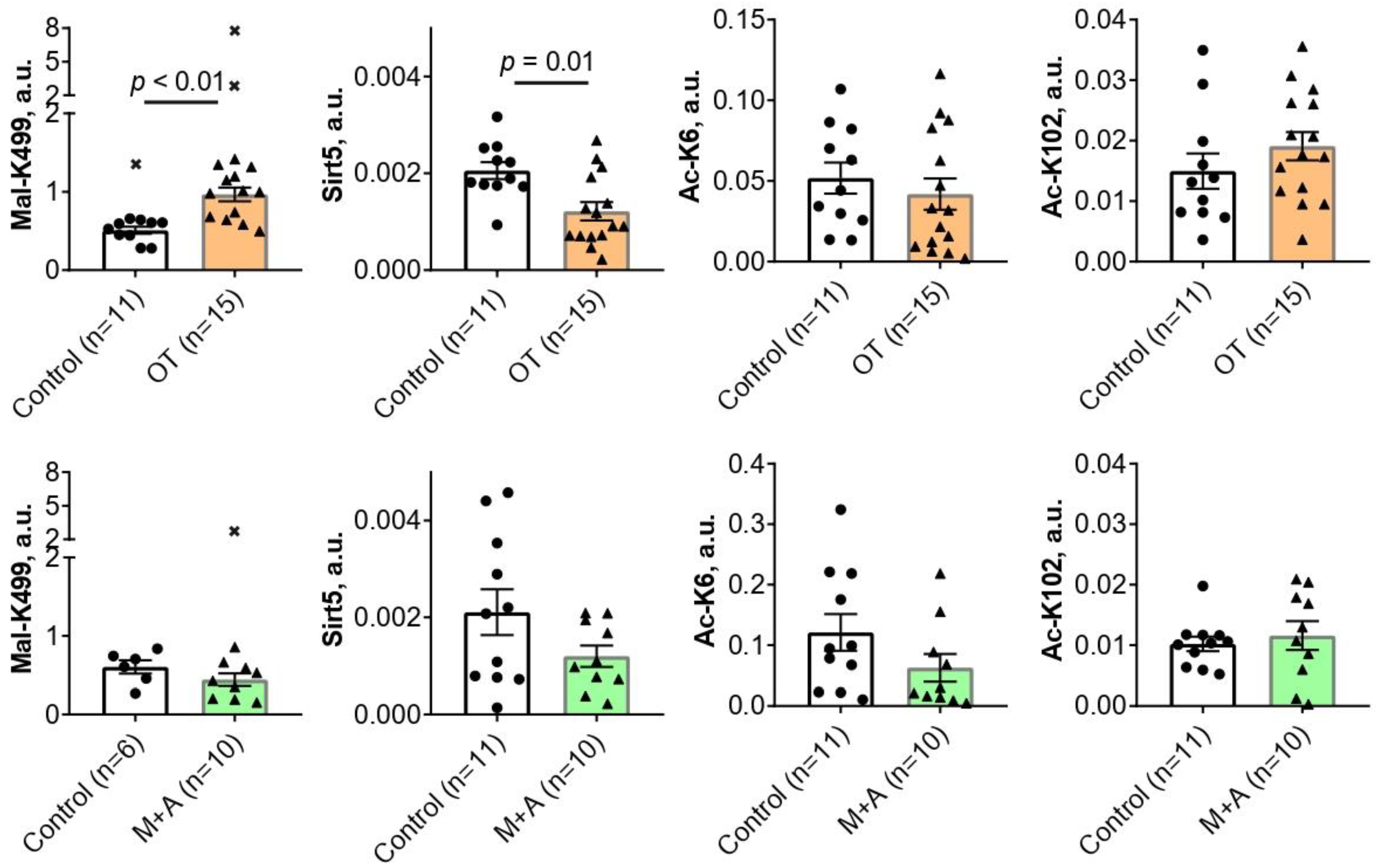

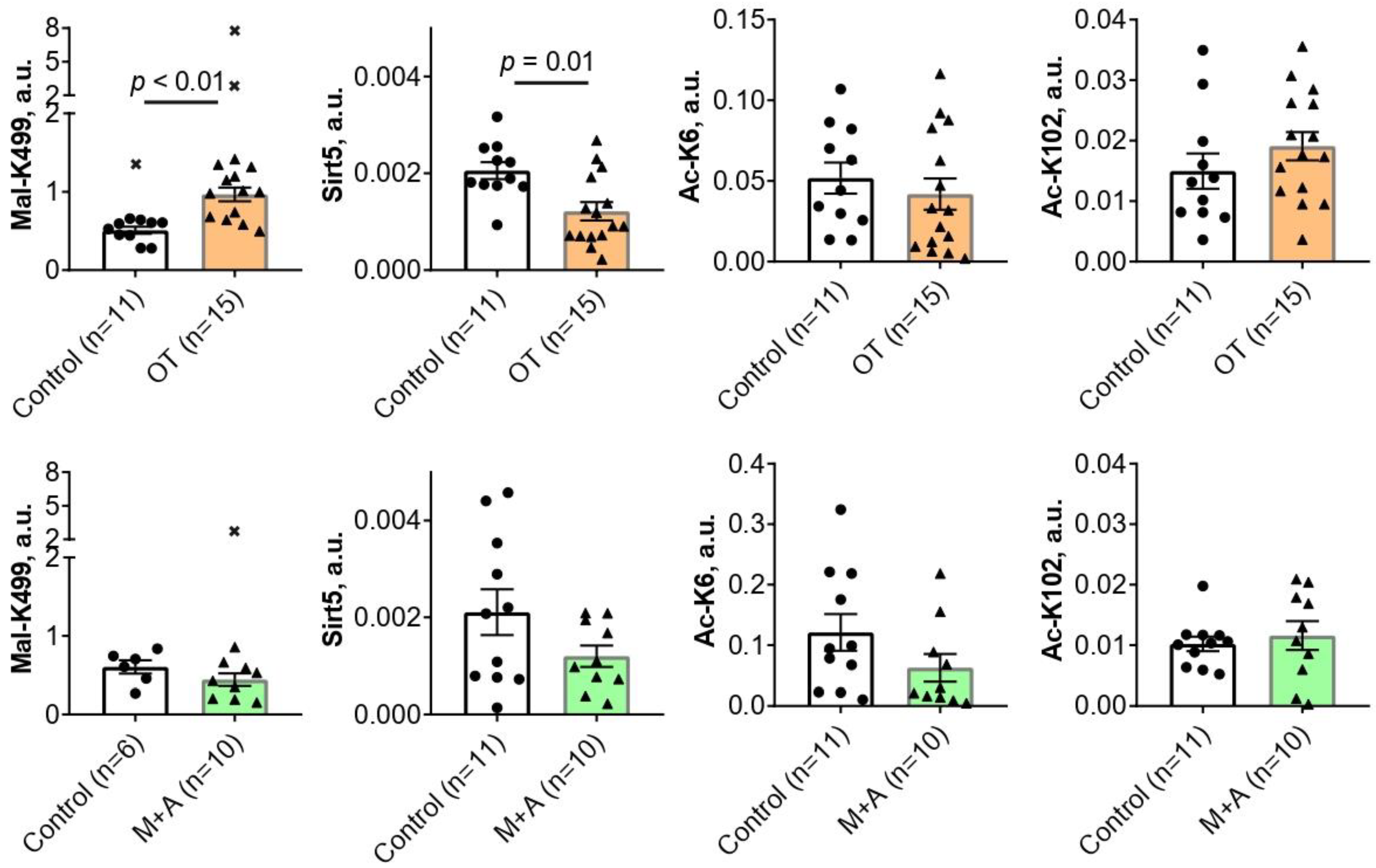{kind=link}
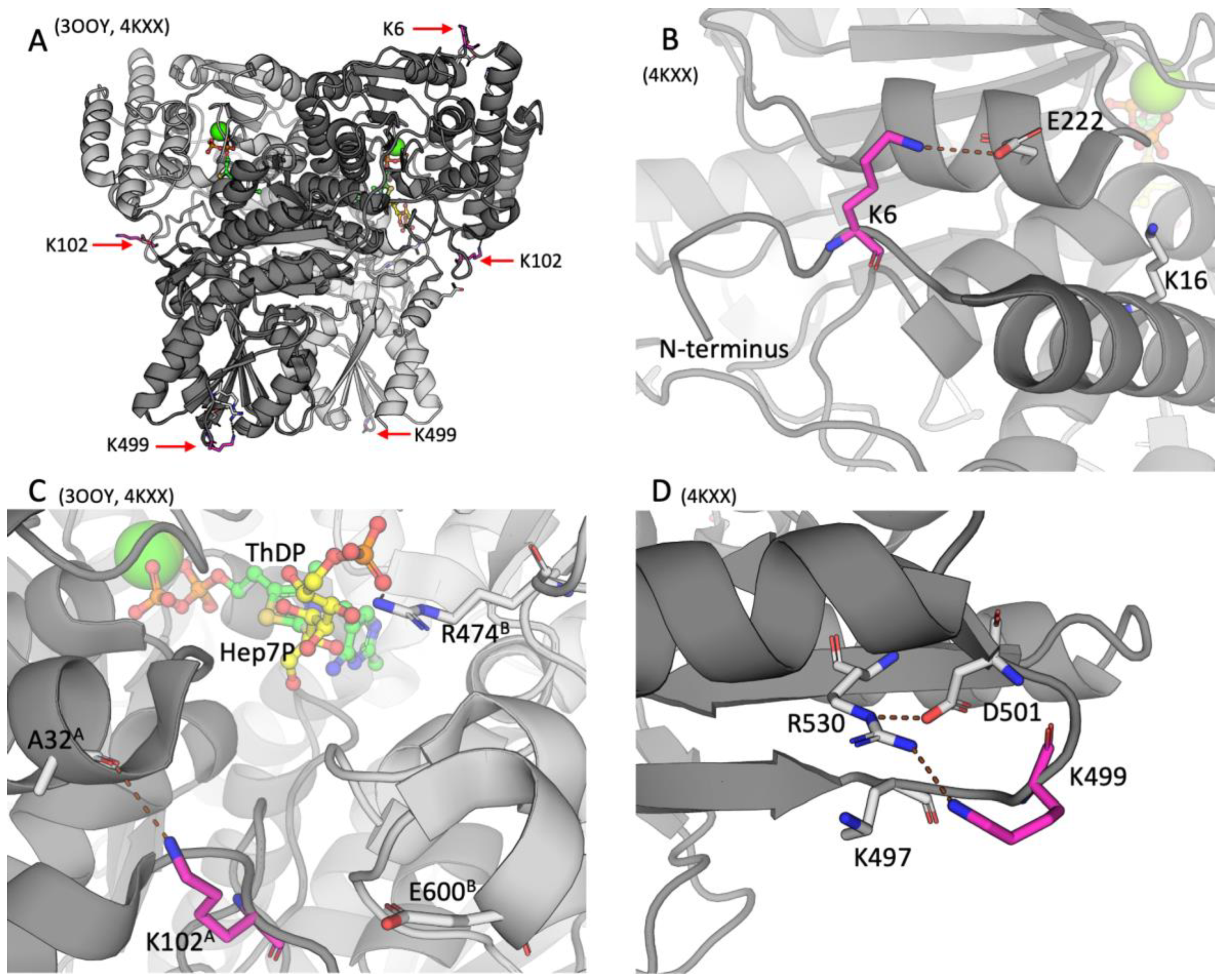{kind=link}
| A. Perturbations in Thiamine Metabolism | ||||||||
| M+A | Mal-K499 | Ac-K6 | Ac-K102 | TKT-expr | Act -ThDP | Act +ThDP | Sirt5 | |
| OT | ||||||||
| Mal-K499 | −0.14 | −0.15 | 0.20 | −0.07 | −0.22 | −0.19 | ||
| 0.66 | 0.62 | 0.51 | 0.80 | 0.43 | 0.53 | |||
| Ac-K6 | −0.22 | −0.08 | 0.22 | −0.35 | −0.38 | 0.76 | ||
| 0.31 | 0.74 | 0.35 | 0.12 | 0.09 | 0.00 | |||
| Ac-K102 | 0.03 | 0.13 | 0.26 | −0.45 | −0.41 | −0.11 | ||
| 0.88 | 0.54 | 0.25 | 0.04 | 0.07 | 0.65 | |||
| TKT-expr | −0.08 | −0.04 | 0.12 | −0.45 | −0.47 | 0.59 | ||
| 0.73 | 0.84 | 0.55 | 0.04 | 0.03 | 0.01 | |||
| Act -ThDP | −0.52 | −0.47 | −0.10 | 0.15 | 0.95 | −0.26 | ||
| 0.01 | 0.02 | 0.62 | 0.47 | 0.00 | 0.26 | |||
| Act +ThDP | −0.40 | −0.50 | −0.20 | −0.08 | 0.97 | −0.30 | ||
| 0.07 | 0.01 | 0.35 | 0.72 | 0.00 | 0.19 | |||
| Sirt5 | −0.16 | 0.60 | −0.18 | 0.51 | −0.21 | −0.30 | ||
| 0.47 | 0.00 | 0.38 | 0.01 | 0.31 | 0.15 | |||
| B. Control State | ||||||||
| Mal-K499 | Ac-K6 | Ac-K102 | TKT-expr | Act -ThDP | Act +ThDP | Sirt5 | ||
| Mal-K499 | −0.12 | −0.26 | 0.32 | −0.31 | −0.36 | 0.18 | ||
| 0.68 | 0.34 | 0.24 | 0.23 | 0.19 | 0.52 | |||
| Ac-K6 | −0.36 | −0.20 | −0.16 | −0.25 | 0.61 | |||
| 0.10 | 0.37 | 0.49 | 0.28 | 0.00 | ||||
| Ac-K102 | 0.04 | 0.07 | 0.02 | −0.52 | ||||
| 0.88 | 0.76 | 0.94 | 0.01 | |||||
| TKT-expr | 0.07 | −0.10 | 0.40 | |||||
| 0.76 | 0.67 | 0.06 | ||||||
| Act -ThDP | 0.93 | −0.10 | ||||||
| 0.00 | 0.67 | |||||||
| Act +ThDP | −0.14 | |||||||
| 0.53 | ||||||||
| Sirt5 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleshin, V.A.; Kaehne, T.; Maslova, M.V.; Graf, A.V.; Bunik, V.I. Posttranslational Acylations of the Rat Brain Transketolase Discriminate the Enzyme Responses to Inhibitors of ThDP-Dependent Enzymes or Thiamine Transport. Int. J. Mol. Sci. 2024, 25, 917. https://doi.org/10.3390/ijms25020917
Aleshin VA, Kaehne T, Maslova MV, Graf AV, Bunik VI. Posttranslational Acylations of the Rat Brain Transketolase Discriminate the Enzyme Responses to Inhibitors of ThDP-Dependent Enzymes or Thiamine Transport. International Journal of Molecular Sciences. 2024; 25(2):917. https://doi.org/10.3390/ijms25020917
Chicago/Turabian StyleAleshin, Vasily A., Thilo Kaehne, Maria V. Maslova, Anastasia V. Graf, and Victoria I. Bunik. 2024. "Posttranslational Acylations of the Rat Brain Transketolase Discriminate the Enzyme Responses to Inhibitors of ThDP-Dependent Enzymes or Thiamine Transport" International Journal of Molecular Sciences 25, no. 2: 917. https://doi.org/10.3390/ijms25020917
APA StyleAleshin, V. A., Kaehne, T., Maslova, M. V., Graf, A. V., & Bunik, V. I. (2024). Posttranslational Acylations of the Rat Brain Transketolase Discriminate the Enzyme Responses to Inhibitors of ThDP-Dependent Enzymes or Thiamine Transport. International Journal of Molecular Sciences, 25(2), 917. https://doi.org/10.3390/ijms25020917