


Novel Fermentation Strategies of Strawberry Tree Arbutus unedo Fruits to Obtain High Nutritional Value Products
 , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
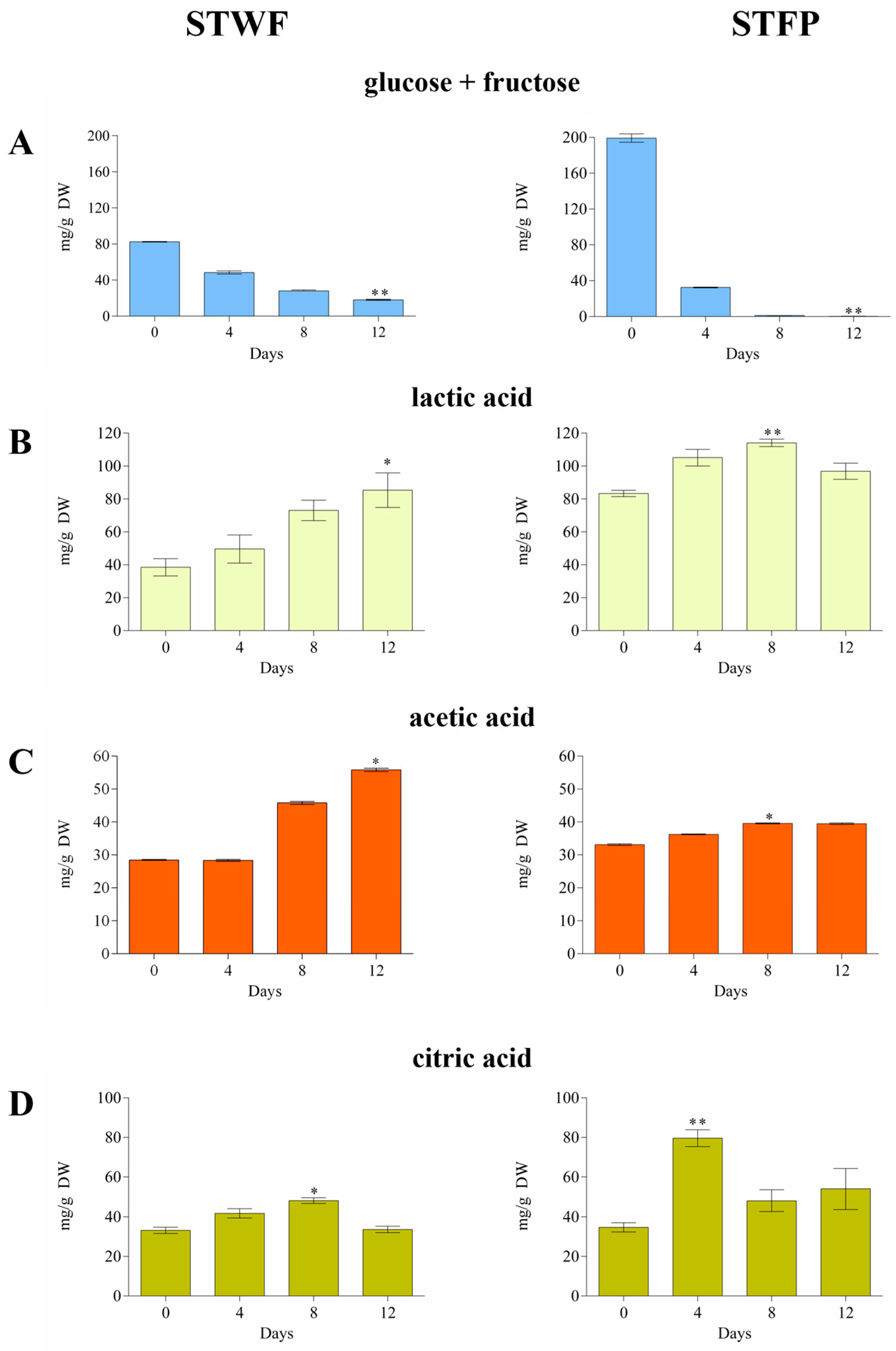
2.1. Strawberry Tree Fermentation Strategies
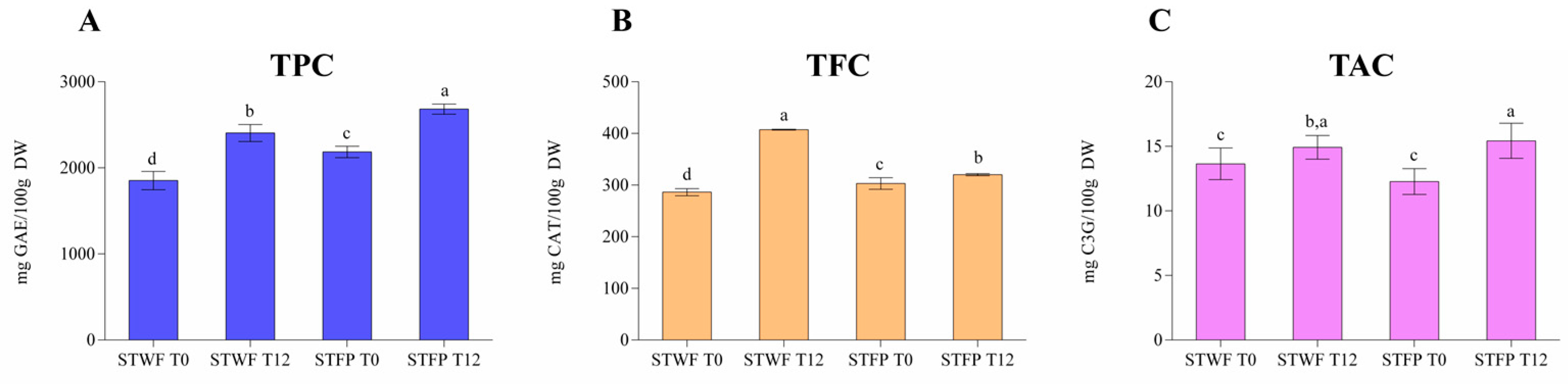
2.2. Phenolic Compound Content
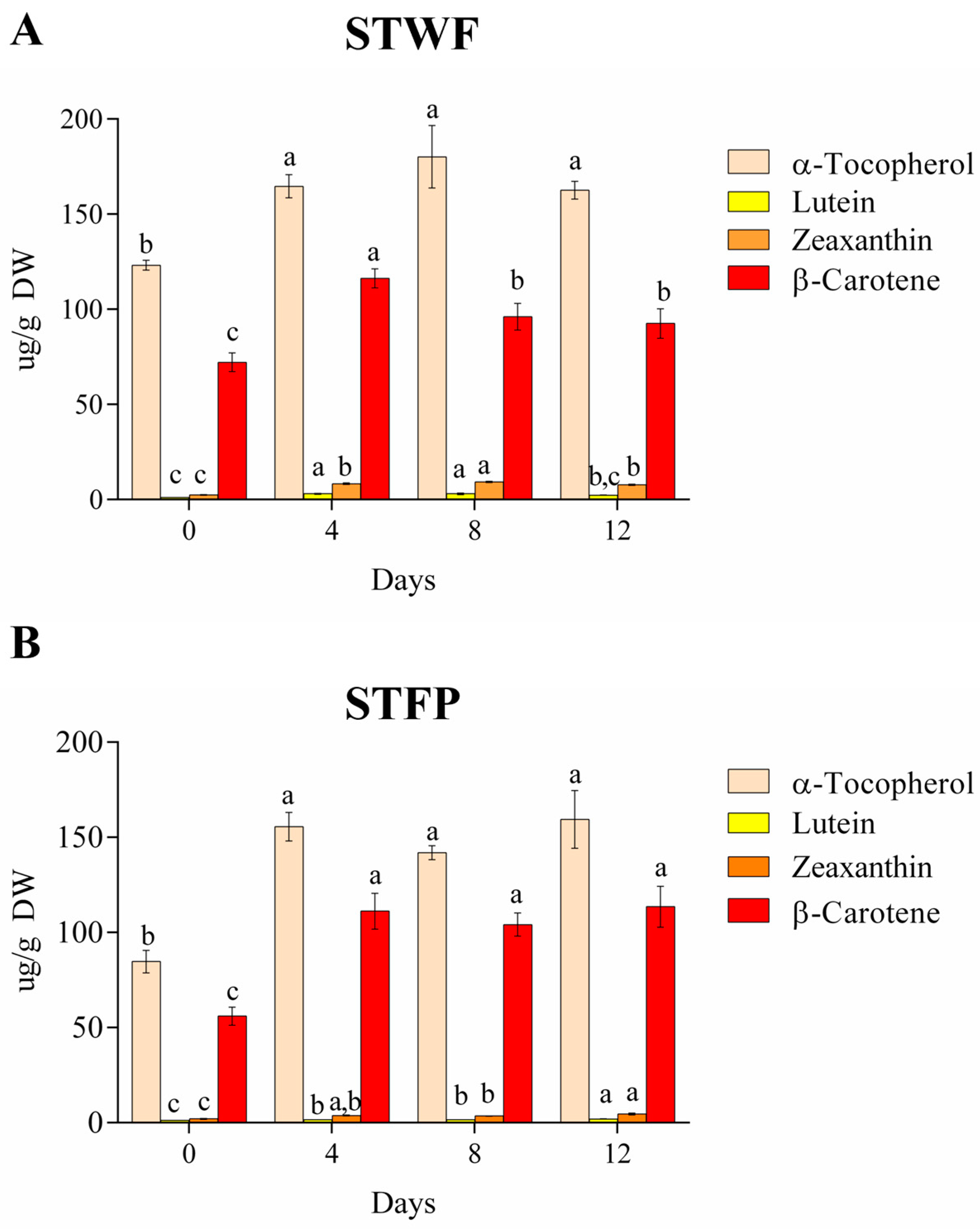
2.3. Isoprenoids Content during Fermentation of Strawberry Tree Fruit Samples
2.4. Enzyme Activities
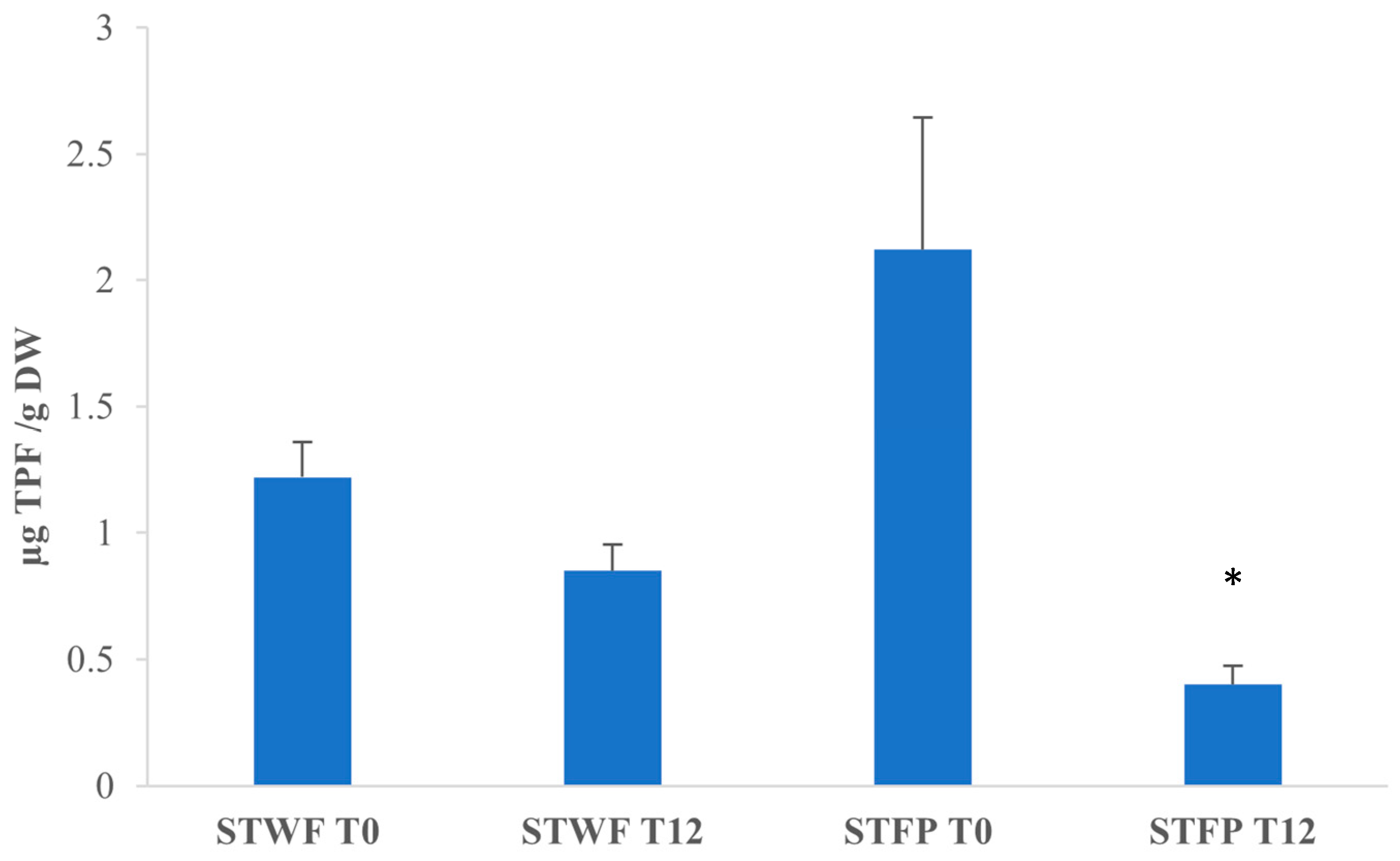
2.5. Antioxidant Activity
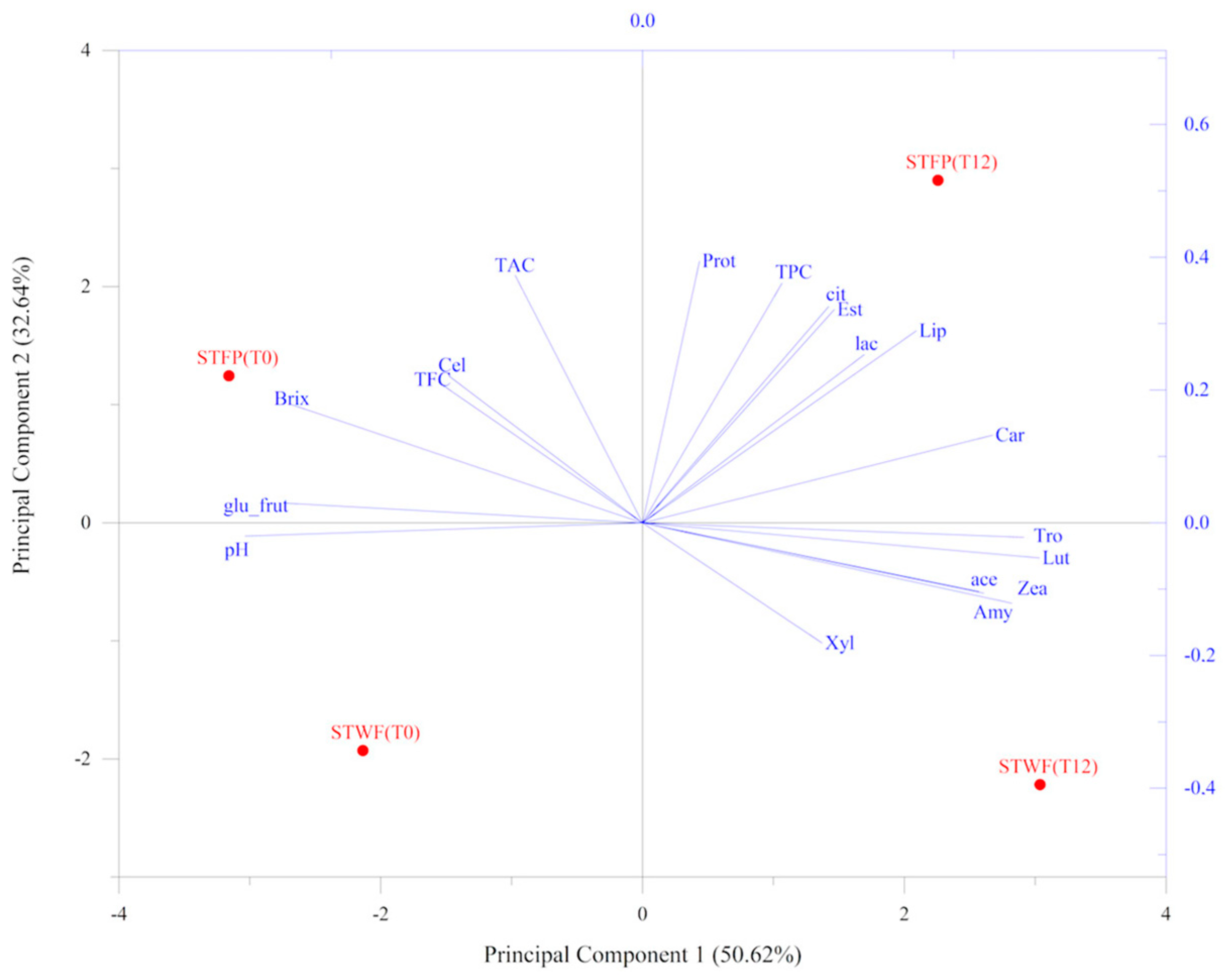
2.6. Principal Component Analysis
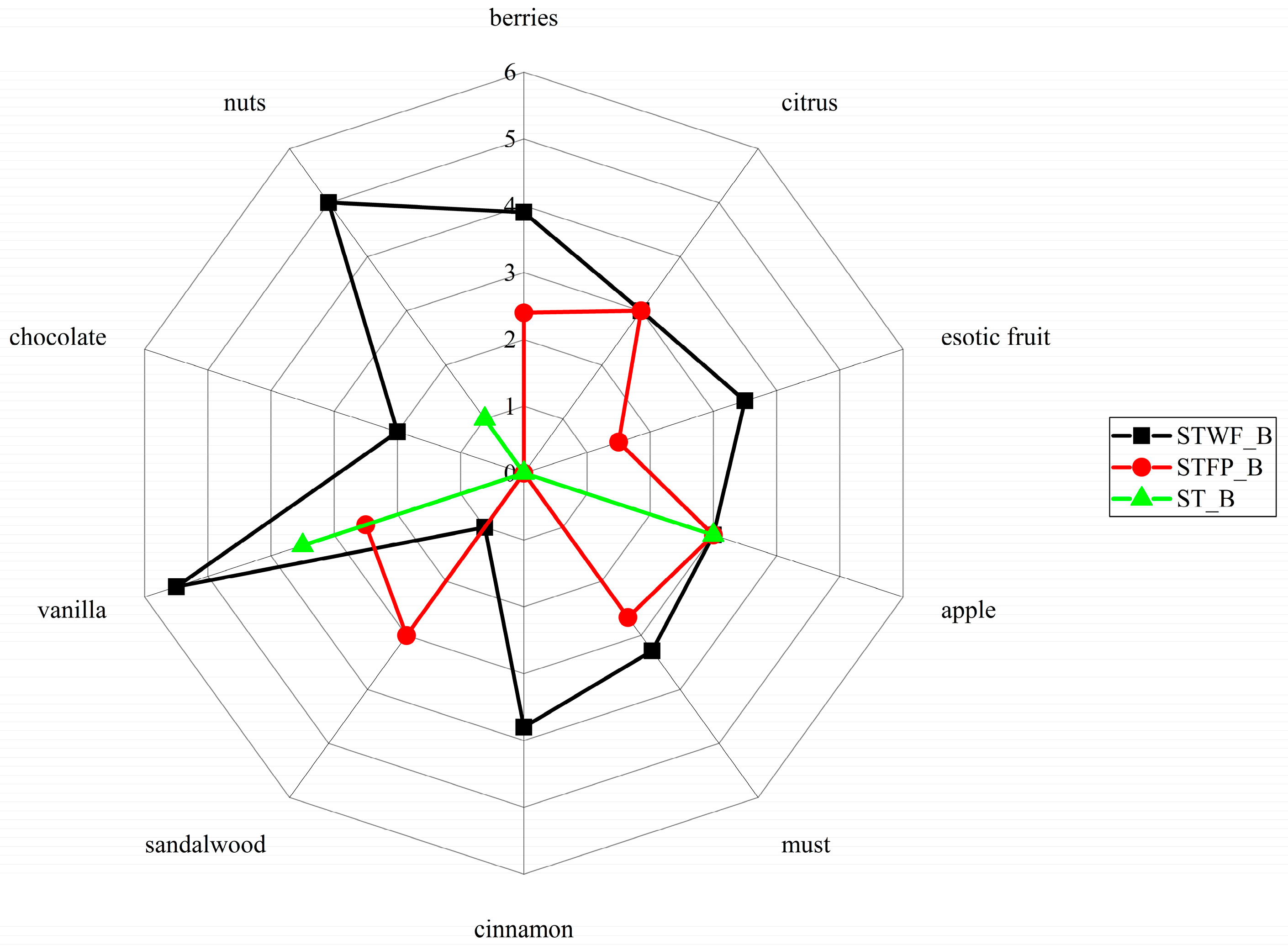
2.7. Preparation and Descriptive Sensory Analysis of an Enriched Food Prototype
3. Materials and Methods
3.1. Fruit Sampling
3.2. Microbial Strain
3.3. Preparation of Samples and Fermentation Conditions
3.4. Physical, Chemical and Microbiological Analyses
3.5. Extraction and Analysis of Phenolic Compounds
3.6. Extraction and Analysis of Isoprenoids (Tocopherols and Carotenoids)
3.7. Evaluation of Enzyme Activities
3.8. Antioxidant Activity
3.9. Preparation and Descriptive Sensory Analysis of the Food Prototypes Enriched in Fermented Strawberry Fruit Fermented Products
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martins, J.; Pinto, G.; Canhoto, J. Biotechnology of the multipurpose tree species Arbutus unedo: A review. J. For. Res. 2022, 33, 377–390. [Google Scholar] [CrossRef]
- Heywood, V.H. Flowering Plants of the World; B.T. Batsford: London, UK, 1993. [Google Scholar]
- Stevens, P.F. Angiosperm Phylogeny Website. Version 14 July 2017. 2001. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 10 July 2020).
- Villa, R.S. Ricerche sulla biologia di Arbutus unedo L. (Ericaceae): Ciclo di sviluppo. Boll. Della Soc. Sarda Di Sci. Nat. 1982, 21, 309–317. [Google Scholar]
- Garrido, N.; Silveira, T. Medronho: O Fruto Vermelho Que Portugal Está a (Re)Descobrir. In Público. 2020. Available online: https://www.publico.pt/2020/11/20/sociedade/noticia/medronho-fruto-vermelho-futuro-portugal-redescobrir-1939941 (accessed on 7 January 2021).
- Ait lhaj, Z.; Bchitou, R.; Gaboun, F.; Abdelwahd, R.; Benabdelouahab, T.; Kabbour, M.R.; Pare, P.; Diria, G.; Bakhy, K. Moroccan strawberry tree (Arbutus unedo L.) fruits: Nutritional value and mineral composition. Foods 2021, 10, 2263. [Google Scholar] [CrossRef]
- Oliveira, I.; Baptista, P.; Malheiro, R.; Casal, S.; Bento, A.; Pereira, J.A. Influence of strawberry tree (Arbutus unedo L.) fruit ripening stage on chemical composition and antioxidant activity. Food Res. Int. 2011, 44, 1401–1407. [Google Scholar] [CrossRef]
- Nunes, R.J.D.S. Micromedronho: Design of Microencapsulated Arbutus Unedo Leaves and Fruits by Spray Drying for Supplements and Functional Foods; Universidade do Algarve: Faro, Portugal, 2017. [Google Scholar]
- Pallauf, K.; Rivas-Gonzalo, J.C.; del Castillo, M.D.; Cano, M.P.; de Pascual-Teresa, S. Characterization of the antioxidant composition of strawberry tree (Arbutus unedo L.) fruits. J. Food Comp. Anal. 2008, 21, 273–281. [Google Scholar] [CrossRef]
- Zitouni, H.; Hssaini, L.H.; Ouaabou, R.; Viuda-Martos, M.; Hernandez, F.; Ercisli, S.; Hanine, H. Functional and technological properties of five strawberry (Arbutus unedo L.) fruit as bioactive ingredients in functional foods. Int. J. Food Prop. 2021, 24, 380–399. [Google Scholar] [CrossRef]
- Fortalezas, S.; Tavares, L.; Pimpão, R.; Tyagi, M.; Pontes, V.; Alves, P.M.; Santos, C.N. Antioxidant properties and neuroprotective capacity of strawberry tree fruit (Arbutus unedo). Nutrients 2010, 2, 214–229. [Google Scholar] [CrossRef]
- Ge, H.; Liu, G.; Xiang, Y.F.; Wang, Y.; Guo, C.W.; Chen, N.H.; Xu, J. The mechanism of poly-galloyl-glucoses preventing influenza a virus entry into host cells. PLoS ONE 2014, 9, e94392. [Google Scholar] [CrossRef]
- Saha, R.K.; Takahashi, T.; Kurebayashi, Y.; Fukushima, K.; Minami, A.; Kinbara, N. Antiviral effect of strictinin on influenza virus replication. Antivir. Res. 2010, 88, 10–18. [Google Scholar] [CrossRef]
- Junior, C.O.R.; Verde, S.C.; Rezende, C.A.M.; Caneschi, W.; Couri, M.R.C.; McDougall, B.R.; De Almeida, M.V. Synthesis and HIV-1 inhibitory activities of dicaffeoyl and digalloyl esters of quinic acid derivatives. Curr. Med. Chem. 2013, 20, 724–733. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J. Stability and metabolism of Arbutus unedo bioactive compounds (phenolics and antioxidants) under in vitro digestion and colonic fermentation. Food Chem. 2016, 201, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Migas, P.; Krauze-Baranowska, M. The significance of arbutin and its derivatives in therapy and cosmetics. Phytochem. Lett. 2015, 13, 35–40. [Google Scholar] [CrossRef]
- Jurica, K.; Gobin, I.; Kremer, D.; Cepo, D.V.; Grubesić, R.J.; Karaconji, I.B.; Kosalec, I. Arbutin and its metabolite hydroquinone as the main factors in the antimicrobial effect of strawberry tree (Arbutus unedo L.) leaves. J. Herb. Med. 2017, 8, 17–23. [Google Scholar] [CrossRef]
- Trichopoulou, A.; Vasilopoulou, E.; Hollman, P.; Chamalides, C.; Foufa, E.; Kaloudis, T.; Kromhout, D.; Miskaki, P.; Petrochilou, I.; Poulima, E. Nutritional composition and flavonoid content of edible wild greens and green pies: A potential rich source of antioxidant nutrients in the Mediterranean diet. Food Chem. 2000, 70, 319–323. [Google Scholar] [CrossRef]
- Miguel, M.G.; Faleiro, M.L.; Guerreiro, A.C.; Antunes, M.D. Arbutus unedo L.: Chemical and biological properties. Molecules 2014, 19, 15799–15823. [Google Scholar] [CrossRef] [PubMed]
- Morgado, S.; Morgado, M.; Plácido, A.I.; Roque, F.; Duarte, A.P. Arbutus unedo L.: From traditional medicine to potential uses in modern pharmacotherapy. J. Ethnopharmacol. 2018, 225, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO). The contribution of plant genetic resources for food and agriculture to food security and sustainable agricultural development. In The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture; Food and Agriculture Organization (FAO): Rome, Italy, 2010; pp. 182–201. [Google Scholar]
- Macchioni, V.; Santarelli, V.; Carbone, K. Phytochemical profile, antiradical capacity and α-glucosidase inhibitory potential of wild Arbutus unedo L. fruits from Central Italy: A chemometric approach. Plants 2020, 9, 1785. [Google Scholar] [CrossRef]
- Cavaco, T.; Longuinho, C.; Quintas, C.; Saraiva, I. Chemical and microbial changes during the natural fermentation of strawberry tree (Arbutus unedo L.) fruits. J. Food Biochem. 2007, 31, 715–725. [Google Scholar] [CrossRef]
- González, E.A.; Agrasar, A.T.; Pastrana Castro, L.M.; Fernández, I.O.; Pérez Guerra, N. Solid-state fermentation of red raspberry (Rubus ideaus L.) and arbutus berry (Arbutus unedo, L.) and characterization of their distillates. Food Res. Int. 2011, 44, 1419–1426. [Google Scholar] [CrossRef]
- Tejedor-Calvo, E.; Morales, D. Chemical and Aromatic Changes during Fermentation of Kombucha Beverages Produced Using Strawberry Tree (Arbutus unedo) Fruits. Fermentation 2023, 9, 326. [Google Scholar] [CrossRef]
- Santo, D.E.; Galego, L.; Gonçalves, T.; Quintas, C. Yeast diversity in the Mediterranean strawberry tree (Arbutus unedo L.) fruits’ fermentations. Food Res. Int. 2012, 47, 45–50. [Google Scholar] [CrossRef]
- Callewaert, R.; De Vuyst, L. Bacteriocin production with Lactobacillus amylovorus DCE 471 is improved and stabilized by fed-batch fermentation. Appl. Environ. Microb. 2000, 66, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, M.; Durante, M.; Ramires, F.A.; Grieco, F.; Tommasi, L.; Perbellini, E.; Falco, V.; Tasioula-Margari, M.; Logrieco, A.F.; Mita, G.; et al. New process for production of fermented black table olives using selected autochthonous microbial resources. Front. Microbiol. 2015, 6, 1007. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, M.; Durante, M.; Veneziani, G.; Taticchi, A.; Servili, M.; Bleve, G.; Mita, G. Patè olive cake: Possible exploitation of a by-product for food applications. Front. Nut. 2019, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Ramires, F.A.; Bleve, G.; De Domenico, S.; Leone, A. Combination of Solid State and Submerged Fermentation Strategies to Produce a New Jellyfish-Based Food. Foods 2022, 11, 3974. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Rodríguez, B.M.; Morales, P.; Fernández-Ruiz, V.; Sánchez-Mata, M.C.; Camara, M.; Díez-Marqués, C.; Pardo-de-Santayana, M.; Molina, M.; Tardío, J. Valorization of wild strawberry-tree fruits (Arbutus unedo L.) through nutritional assessment and natural production data. Food Res. Int. 2011, 44, 1244–1253. [Google Scholar] [CrossRef]
- Rodríguez, M.A.R.; Odériz, M.L.V.; Lozano, J.S.; Hernández, J.L. Estudio de la composición química de pequeños frutos: Arándano, frambuesa, grosella blanca, grosella negra, grosella roja y zarzamora producidos en Galicia. Ind. Conserve 1992, 67, 29–33. [Google Scholar]
- Tzamourani, A.P.; Di Napoli, E.; Paramithiotis, S.; Economou-Petrovits, G.; Panagiotidis, S.; Panagou, E.Z. Microbiological and physicochemical characterisation of green table olives of Halkidiki and Conservolea varieties processed by the spanish method on industrial scale. Int. J. Food Sci. Technol. 2021, 56, 3845–3857. [Google Scholar] [CrossRef]
- Izcara, S.; Morante-Zarcero, S.; Casado, N.; Sierra, I. Study of the Phenolic Compound Profile of Arbutus unedo L. Fruits at Different Ripening Stages by HPLC-TQ-MS/MS. Appl. Sci. 2021, 11, 11616. [Google Scholar] [CrossRef]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. 2005, 579, 200–213. [Google Scholar] [CrossRef]
- Yang, F.; Chen, C.; Ni, D.; Yang, Y.; Tian, J.; Li, Y.; Wang, L. Effects of Fermentation on Bioactivity and the Composition of Polyphenols Contained in Polyphenol-Rich Foods: A Review. Foods 2023, 12, 3315. [Google Scholar] [CrossRef] [PubMed]
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Khaneghah, A.M.; Meléndez-Martínez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Technol. 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Maiorano, G.; Ramires, F.A.; Durante, M.; Palamà, I.E.; Blando, F.; De Rinaldis, G.; Bleve, G. The Controlled Semi-Solid Fermentation of Seaweeds as a Strategy for Their Stabilization and New Food Applications. Foods 2022, 11, 2811. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, E.; Medina, E.; Brenes, M.; Romero, C. Endogenous enzymes involved in the transformation of oleuropein in Spanish table olive varieties. J. Agric.Food Chem. 2014, 62, 9569–9575. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Gallego, J.; Rodríguez-Gómez, F.; Barrio, E.; Querol, A.; Garrido-Fernández, A.; Arroyo-López, F.N. Exploring the yeast biodiversity of green table olive industrial fermentations for technological applications. Int. J. Food Microb. 2011, 147, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, N.; Kahar, P.; Lisdiyanti, P.; Lee, J.; Yopi Prasetya, B.; Ogino, C.; Kondo, A. GH-10 and GH-11 Endo-1,4-β-xylanase enzymes from Kitasatospora sp. produce xylose and xylooligosaccharides from sugarcane bagasse with no xylose inhibition. Biores. Technol. 2019, 272, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, P. Sources, production, and classification of xylanases. In Xylanolytic Enzymes; Academic Press: Tokyo, Japan; Elsevier: Amsterdam, The Netherlands, 2014; pp. 43–52. [Google Scholar]
- Chen, Y.; Wang, Y.; Chen, J.; Tang, H.; Wang, C.; Liab, Z.; Xiao, Y. Bioprocessing of soybeans (Glycine max L.) by solid-state fermentation with Eurotium cristatum YL-1 improves total phenolic content, isoflavone aglycones, and antioxidant activity. SC Adv. 2020, 10, 16928. [Google Scholar] [CrossRef]
- Bei, Q.; Chen, G.; Lu, F.; Wu, S.; Wu, Z. Enzymatic action mechanism of phenolic mobilization in oats (Avena sativa L.) during solid-state fermentation with Monascus anka. Food Chem. 2018, 245, 297–304. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Li, G.; Chai, X.; Zhong, Z.; Feng, Y.; Sun, H.; Wang, B. Penicillium fermentation combined with enzyme treatment to enhance the release of phenolic acids from wheat bran. Food Sci. Technol. 2023, 43, e100822. [Google Scholar] [CrossRef]
- McCue, P.P.; Shetty, K. A role for amylase and peroxidase-linked poly- merization in phenolic antioxidant mobilization in dark-germinated soybean and implications for health. Process Biochem. 2004, 39, 1785–1791. [Google Scholar] [CrossRef]
- Markovinović, A.B.; Brdar, D.; Putnik, P.; Bosiljkov, T.; Durgo, K.; Turković, A.H.; Kovačević, D.B. Strawberry tree fruits (Arbutus unedo L.): Bioactive composition, cellular antioxidant activity, and 3D printing of functional foods. Food Chem. 2024, 433, 137287. [Google Scholar] [CrossRef] [PubMed]
- Tenuta, M.C.; Deguin, B.; Loizzo, M.R.; Dugay, A.; Acquaviva, R.; Malfa, G.A.; Bonesi, M.; Bouzidi, C.; Tundis, R. Contribution of flavonoids and iridoids to the hypoglycaemic, antioxidant, and nitric oxide (NO) inhibitory activities of Arbutus unedo L. Antioxidants 2020, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Guzzon, R.; Nardin, T.; Larcher, R. The controversial relationship between chitosan and the microorganisms involved in the production of fermented beverages. Eur. Food. Res.Technol. 2022, 248, 751–765. [Google Scholar] [CrossRef]
- Catalano, F.; Romaniello, R.; Orsino, M.; Perone, C.; Bianchi, B.; Giametta, F. Experimental Tests in Production of Ready-to-Drink Primitive Wine with Different Modes of Circulation of the Fermenting Must. Appl. Sci. 2023, 13, 5941. [Google Scholar] [CrossRef]
- Ramires, F.A.; Bavaro, A.R.; D’Antuono, I.; Linsalata, V.; D’Amico, L.; Baruzzi, F.; Bleve, G. Liquid submerged fermentation by selected microbial strains for onion skins valorization and its effects on polyphenols. World J. Microb. Biotech. 2023, 39, 258. [Google Scholar] [CrossRef]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Giusti, M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV–visible Spectroscopy. Curren. Protoc. Food Chem. 2001, 1042, F1.2.1–F1.2.13. [Google Scholar]
- D’Antuono, I.; Bruno, A.; Linsalata, V.; Minervini, F.; Garbetta, A.; Tufariello, M.; Cardinali, A. Fermented Apulian table olives: Effect of selected microbial starters on polyphenols composition, antioxidant activities and bioaccessibility. Food Chem. 2018, 248, 137–145. [Google Scholar] [CrossRef]
- Blando, F.; Russo, R.; Negro, C.; De Bellis, L.; Frassinetti, S. Antimicrobial and antibiofilm activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. cladode polyphenolic extracts. Antioxidants 2019, 8, 117. [Google Scholar] [CrossRef]
- Durante, M.; Lenucci, M.S.; Marrese, P.P.; Rizzi, V.; De Caroli, M.; Piro, G.; Mita, G. α-Cyclodextrin encapsulation of supercritical CO2 extracted oleoresins from different plant matrices: A stability study. Food Chem. 2016, 199, 684–693. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, J.C.; Choi, J.S. The antioxidant activity of Ecklonia stolonifera. Arch. Pharm. Res. 1996, 19, 223–227. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Garbetta, A.; Nicassio, L.; D’Antuono, I.; Cardinali, A.; Linsalata, V.; Attolico, G.; Minervini, F. Influence of In Vitro digestion process on polyphenolic profile of skin grape (cv. Italia) and on antioxidant activity in basal or stressed conditions of human intestinal cell line (HT-29). Food. Res. Int. 2018, 106, 878–884. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Fermentation (Days) | STWF | STFP | ||
|---|---|---|---|---|
| pH | Brix (%) | pH | Brix (%) | |
| 0 | 3.61 ± 0.51 a | 10.5 ± 0.6 a | 3.61 ± 0.57 a | 15.9 ± 0.6 a |
| 4 | 3.45 ± 0.40 a | 8 ± 0.5 b | 3.49 ± 0.61 a | 12 ± 0.6 b |
| 8 | 3.23 ± 0.43 a | 6.8 ± 0.3 b,c | 3.28 ± 0.32 a | 11 ± 0.4 b |
| 12 | 3.14 ± 0.49 a | 5.2 ± 0.3 c | 3.21 ± 0.44 a | 9.2 ± 0.4 c |
| Microorganisms | Medium | Days | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 4 | 8 | 12 | ||||||
| STWF | STFP | STWF | STFP | STWF | STFP | STWF | STFP | ||
| Yeasts | DRBC | 2.56 ± 0.99 b | 3.58 ± 2.51 b | 6.82 ± 3.58 a | 7.67 ± 3.13 a | 7.07 ± 3.16 a | 7.65 ± 4.33 a | 7.06 ± 3.33 a | 7.65 ± 4.59 a |
| Moulds | DRBC | 3.08 ± 0.72 a | 2.68 ± 1.88 a | 0 b | 0 b | 0 b | 0 b | 0 b | 0 b |
| Aerobic colony count | PCA | 3.75 ± 1.23 b | 3.66 ± 2.09 a | 6.63 ± 3.09 a | 2.08 ± 1.48 a | 7.85 ± 3.12 a | 0 b | 6.65 ± 3.51 a | 0 b |
| Lactic acid bacteria | MRS | 0 b | 0 a | 7.19 ± 3.41 a | 0 a | 7.81 ± 3.87 a | 0 a | 7.85± 3.83 a | 0 a |
| Clostridium spp. | SPS | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| Enterobacteriaceae | VRBGA | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| coli-aerogenes bacteria | VRBA | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| Putative coagulase positive staphylococci | BPA | 1.98 ± 0.81 a | 2.03 ± 1.18 a | 0 b | 0 b | 0 b | 0 b | 0 b | 0 b |
| Polyphenols | STWF | STFP | ||
|---|---|---|---|---|
| T0 | T12 | T0 | T12 | |
| mg/100 g dw | ||||
| Gallic acid der | 149.66 ± 10.37 c | 117.00 ± 9.30 a | 122.45 ± 3.11 a | 137.91 ± 4.52 b |
| Gallic acid | 30.03 ± 3.41 b | 23.86 ± 7.67 b | 16.71 ± 3.70 a | 219.87 ± 1.48 c |
| Ellagic acid der 1 | 3.12 ± 0.33 a | 4.18 ± 0.96 ab | 4.51 ± 1.34 b | nd |
| Ellagic acid der 2 | 7.04 ± 0.36 b | 9.85 ± 2.39 c | 10.97 ± 0.61 c | 1.94 ± 0.20 a |
| Ellagic acid | 20.61 ± 3.37 a | 25.81 ± 2.10 b | 17.57 ± 2.61 a | 43.67 ± 0.79 c |
| Quercetin glycoside | 2.29 ± 0.42 a | 2.83 ± 0.83 ab | 3.36 ± 0.78 b | 2.81 ± 0.26 ab |
| Myricetin | 0.91 ± 0.11 a | 0.81 ± 0.22 a | nd | 1.28 ± 0.33 b |
| Quercetin 3-O-ramnoside | 6.06 ± 0.70 a | 7.53 ± 0.35 b | 7.20 ± 1.32 b | 9.71 ± 0.65 c |
| Ellagic acid der 3 | 0.79 ± 0.24 b | 0.97 ± 0.03 b | 0.47 ± 0.17 a | 0.87 ± 0.01 b |
| Quercetin | 5.07 ± 1.10 b | 5.58 ± 0.95 b | 0.84 ± 0.20 a | 10.03 ± 2.42 c |
| Kaempferide | 0.53 ± 0.11 b | 0.33 ± 0.06 a | nd | 0.85 ± 0.18 c |
| Anthocyanin 1 | 0.32 ± 0.10 a | 1.74 ± 0.11 b | 1.42 ± 0.60 b | nd |
| Cyanidin 3-O-glucoside | 4.10 ± 0.40 a | 13.31 ± 4.68 ab | 19.15 ± 17.71 b | 1.60 ± 0.26 a |
| Anthocyanin 1 | 0.41 ± 0.15 b | nd | 0.26 ± 0.05 a | nd |
| Anthocyanin 2 | 1.24 ± 0.30 a | 3.94 ± 0.01 b | 4.61 ± 0.97 b | 1.46 ± 0.56 a |
| Anthocyanin 3 | 1.05 ± 0.09 c | 0.43 ± 0.08 a | 0.46 ± 0.09 a | 0.63 ± 0.11 b |
| Total | 233.23 ± 3.10 b | 218.17 ± 9.89 a | 209.96 ± 19.54 a | 432.63 ± 0.67 c |
| Lipase U/g | Esterase mU/g | Amylase U/g | Protease U/g | Cellulase U/g | Endo-Xylanase U/g | |
|---|---|---|---|---|---|---|
| STWF | ||||||
| T0 | 124.05 ± 9.02 a | 10.09 ± 0.89 a | 19.2 6 ± 1.2 a | 40.27 ± 2.80 a | 11.30 ± 0.19 a | 4.41 ± 0.09 a |
| T12 | 166.97 ± 7.64 b | 12.31 ± 0.25 b | 24.58 ± 2.79 a | 53.05 ± 13.23 a | 11.89 ± 1.02 a | 3.49 ± 0.16 b |
| STFP | ||||||
| T0 | 114.06 ± 4.70 a | 5.59 ± 0.89 a | 23.34 ± 0.14 a | 13.90 ± 2.27 a | 12.36 ± 1.58 a | 3.48 ± 0.13 a |
| T12 | 133.13 ± 12.36 a | 9.37 ± 0.22 b | 23.68 ± 3.23 a | 20.12 ± 2.57 a | 9.84 ± 0.95 a | 6.06 ± 0.08 b |
| Bioactive Compound | Spearman’s Rho (rs) | Lipase | Esterase | Amylase | Protease | Cellulase | Endo-Xylanase |
|---|---|---|---|---|---|---|---|
| Total phenol content | Spearman Corr. | 0.755240 | 0.929600 * | −0.083920 | 0.956220 * | 0.237760 | 0.069930 |
| p-value | 0.004510 | 0.000012 | 0.795410 | 0.000001 | 0.456800 | 0.829020 | |
| Total flavonoid content | Spearman Corr. | 0.391610 | 0.732410 * | −0.524480 | 0.718040 * | 0.132870 | 0.314690 |
| p-value | 0.208060 | 0.006750 | 0.080020 | 0.008540 | 0.680600 | 0.319140 | |
| Total anthocyanin content | Spearman Corr. | 0.237760 | 0.598610 * | −0.244760 | 0.605960 * | 0.594410 * | −0.531470 |
| p-value | 0.456800 | 0.039740 | 0.443260 | 0.036760 | 0.041520 | 0.075360 | |
| α-Tocopherol | Spearman Corr. | 0.584940 | 0.239860 | 0.889670 * | 0.263160 | −0.171630 | 0.017510 |
| p-value | 0.045740 | 0.452710 | 0.000107 | 0.408580 | 0.593790 | 0.956920 | |
| Lutein | Spearman Corr. | 0.447550 | 0.021130 | 0.874130 | 0.038530 | −0.447550 | 0.286710 |
| p-value | 0.144590 | 0.948040 | 0.000201 | 0.905370 | 0.144590 | 0.366250 | |
| Zeaxanthin | Spearman Corr. | 0.447550 | 0.021130 | 0.874130 * | 0.038530 | −0.447550 | 0.286710 |
| p-value | 0.144590 | 0.948040 | 0.000201 | 0.905370 | 0.144590 | 0.366250 | |
| β-carotene | Spearman Corr. | 0.711030 * | 0.380960 | 0.742560 * | 0.417540 | 0.073560 | −0.101580 |
| p-value | 0.009530 | 0.221790 | 0.005670 | 0.176840 | 0.820280 | 0.753430 |
| STWF | STFP | ||||||
|---|---|---|---|---|---|---|---|
| T0 | T12 | T0 | T12 | ||||
| TPF (µg/g DW) | CAA Unit | TPF (µg/g DW) | CAA Unit | TPF (µg/g DW) | CAA Unit | TPF (µg/g DW) | CAA Unit |
| 240 | 96 | 155 | 96 | 441 | 96 | 441 | 96 |
| 48 | 93 | 31 | 93 | 88 | 92 | 88 | 92 |
| 9.6 | 77 | 6.2 | 86 | 18 | 81 | 18 | 81 |
| 1.92 | 58 | 1.24 | 64 | 3.5 | 55 | 3.5 | 55 |
| 0.38 | 31 | 0.25 | 41 | 0.70 | 33 | 0.70 | 33 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramires, F.A.; Durante, M.; D’Antuono, I.; Garbetta, A.; Bruno, A.; Tarantini, A.; Gallo, A.; Cardinali, A.; Bleve, G. Novel Fermentation Strategies of Strawberry Tree Arbutus unedo Fruits to Obtain High Nutritional Value Products. Int. J. Mol. Sci. 2024, 25, 684. https://doi.org/10.3390/ijms25020684
Ramires FA, Durante M, D’Antuono I, Garbetta A, Bruno A, Tarantini A, Gallo A, Cardinali A, Bleve G. Novel Fermentation Strategies of Strawberry Tree Arbutus unedo Fruits to Obtain High Nutritional Value Products. International Journal of Molecular Sciences. 2024; 25(2):684. https://doi.org/10.3390/ijms25020684
Chicago/Turabian StyleRamires, Francesca Anna, Miriana Durante, Isabella D’Antuono, Antonella Garbetta, Angelica Bruno, Annamaria Tarantini, Antonia Gallo, Angela Cardinali, and Gianluca Bleve. 2024. "Novel Fermentation Strategies of Strawberry Tree Arbutus unedo Fruits to Obtain High Nutritional Value Products" International Journal of Molecular Sciences 25, no. 2: 684. https://doi.org/10.3390/ijms25020684
APA StyleRamires, F. A., Durante, M., D’Antuono, I., Garbetta, A., Bruno, A., Tarantini, A., Gallo, A., Cardinali, A., & Bleve, G. (2024). Novel Fermentation Strategies of Strawberry Tree Arbutus unedo Fruits to Obtain High Nutritional Value Products. International Journal of Molecular Sciences, 25(2), 684. https://doi.org/10.3390/ijms25020684