
Integrated Metabolomics Approach Reveals the Dynamic Variations of Metabolites and Bioactivities in Paeonia ostii ‘Feng Dan’ Leaves during Development

 ,
,
Abstract
1. Introduction
2. Results and Discussion
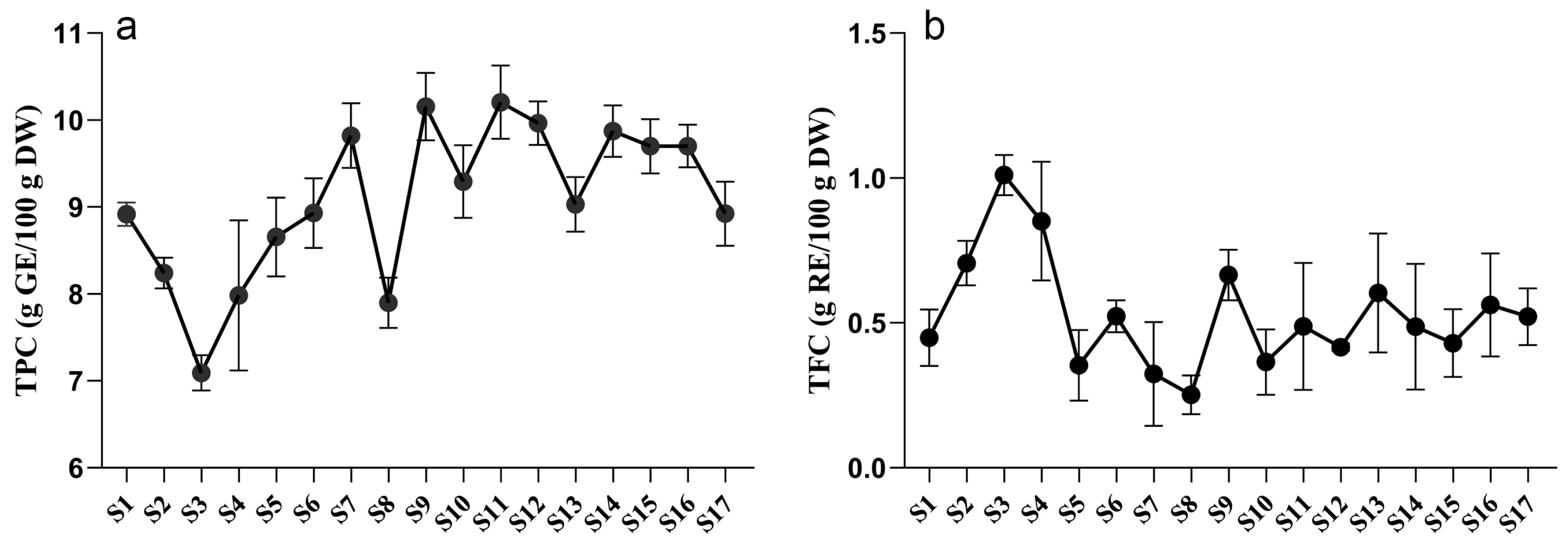
2.1. Tree Peony Leaves Are Rich in Phenols and Flavonoids
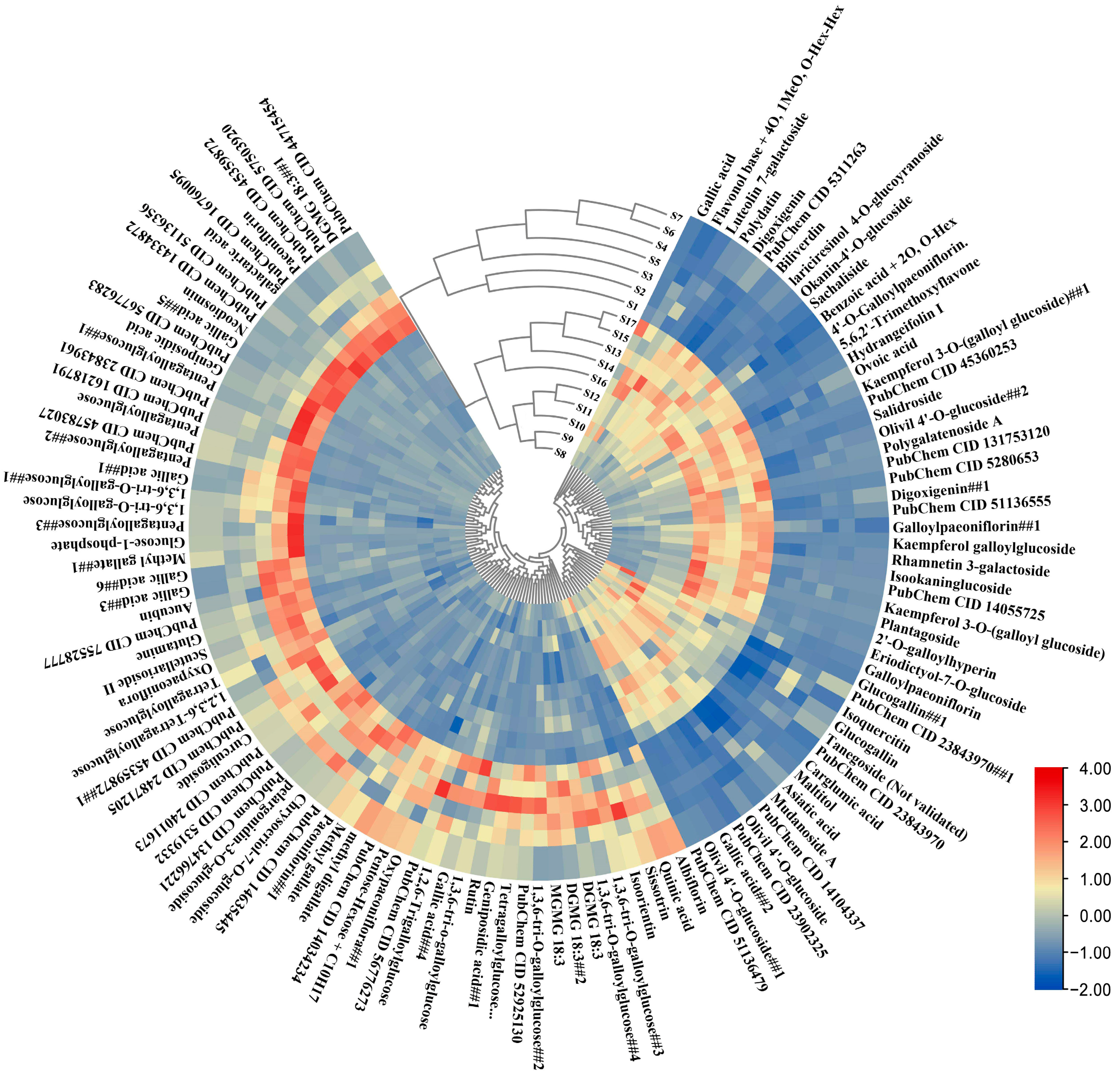
2.2. Annotation of Metabolites through Non-Targeted Metabolome Approach
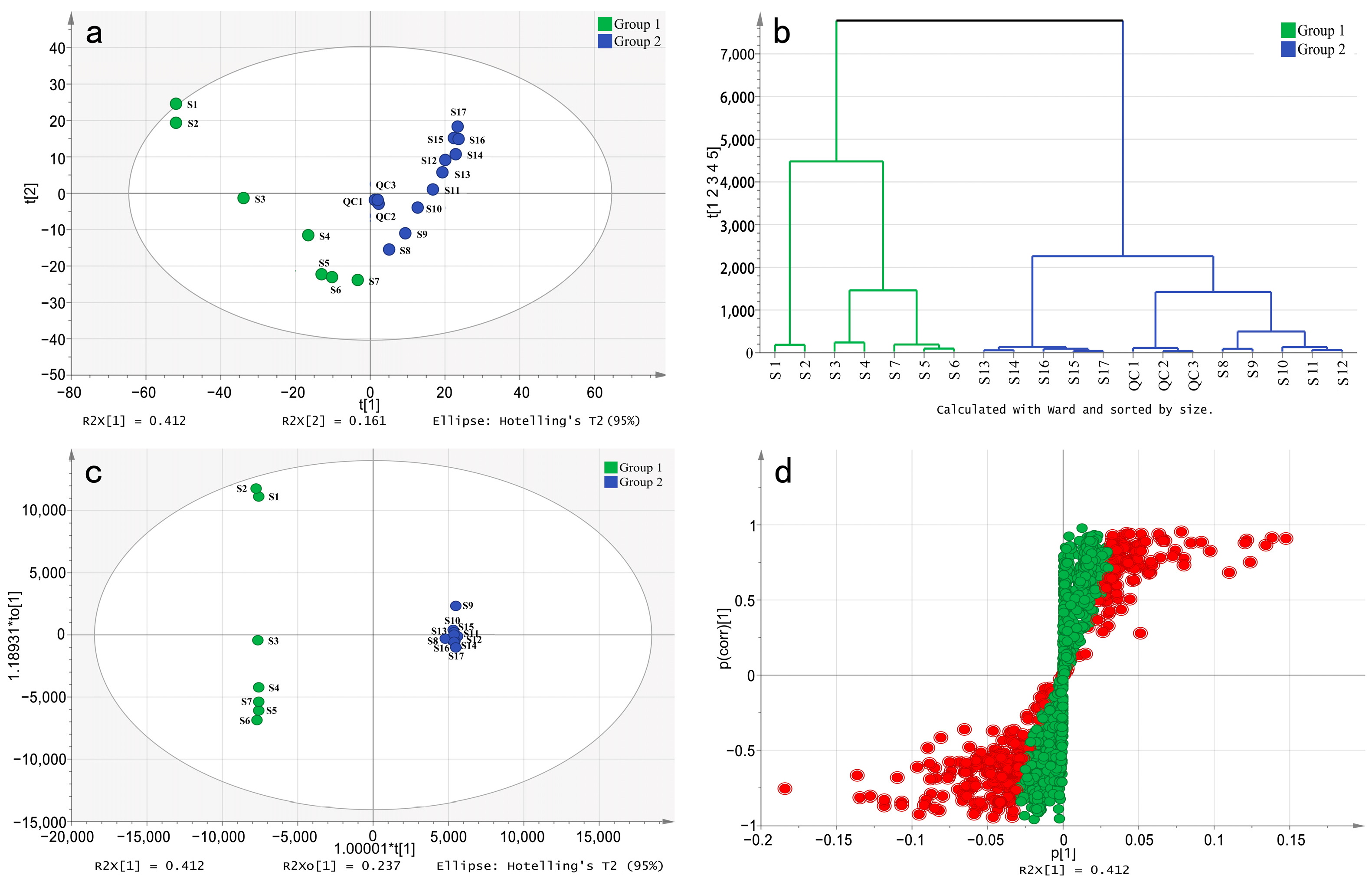
2.3. Multivariate Statistical Analysis of Non-Targeted Metabolome Data
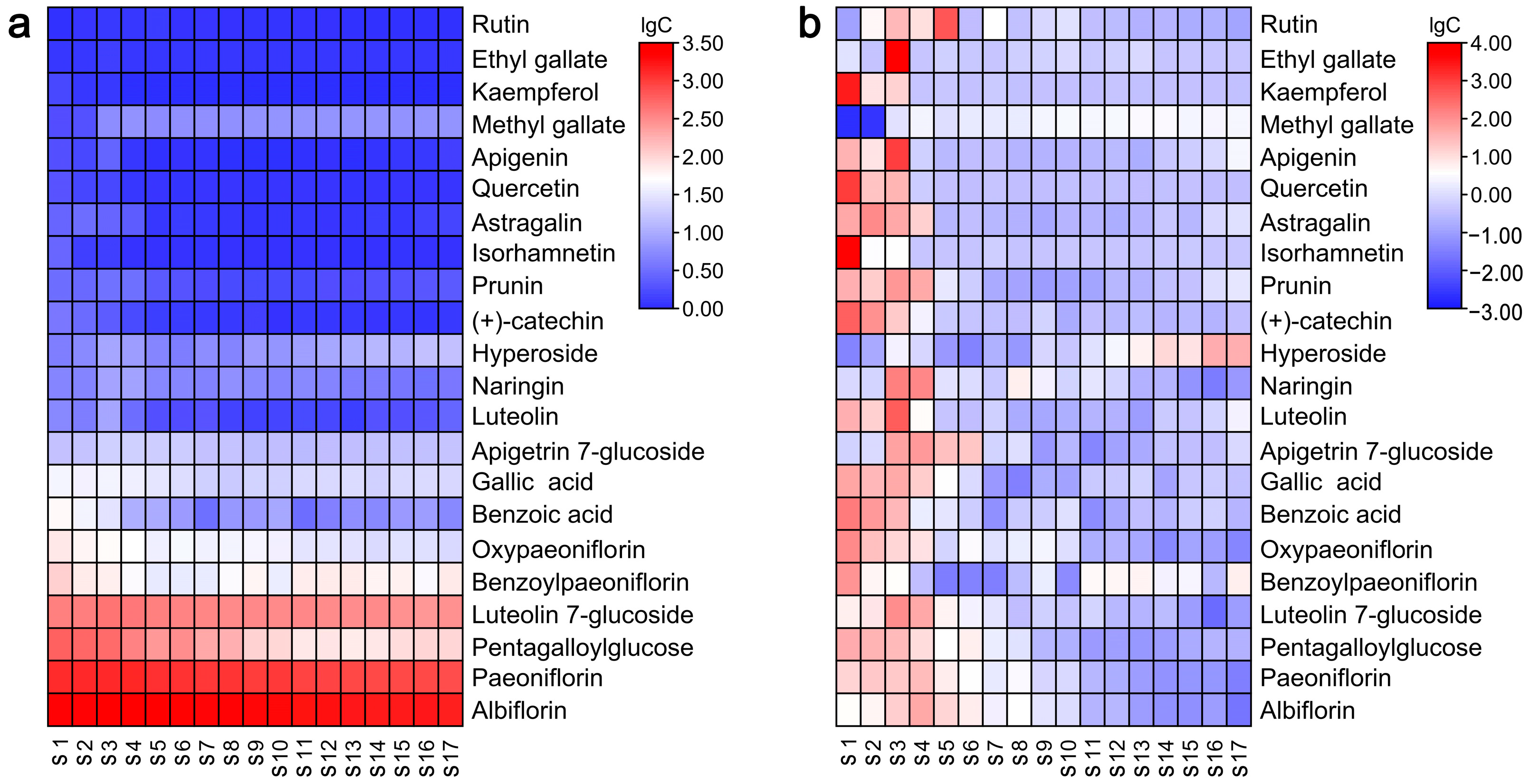
2.4. Quantification of Metabolites through Targeted LC-QQQ-MS Analysis
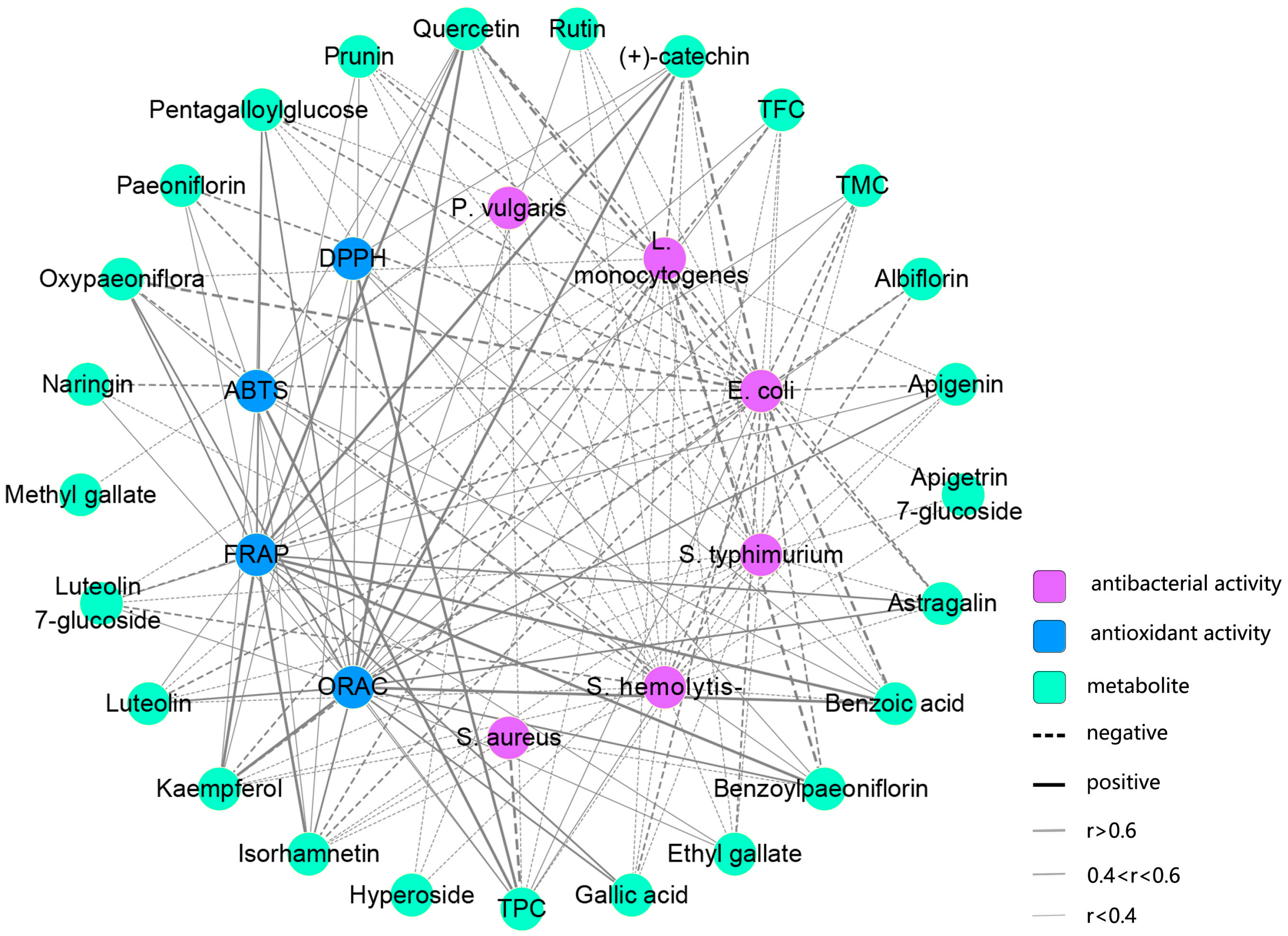
2.5. Antioxidant Activities of TPLs at Different Stages
2.6. Antibacterial Activities of TPLs at Different Stages
3. Materials and Methods
3.1. Chemicals and Reagents
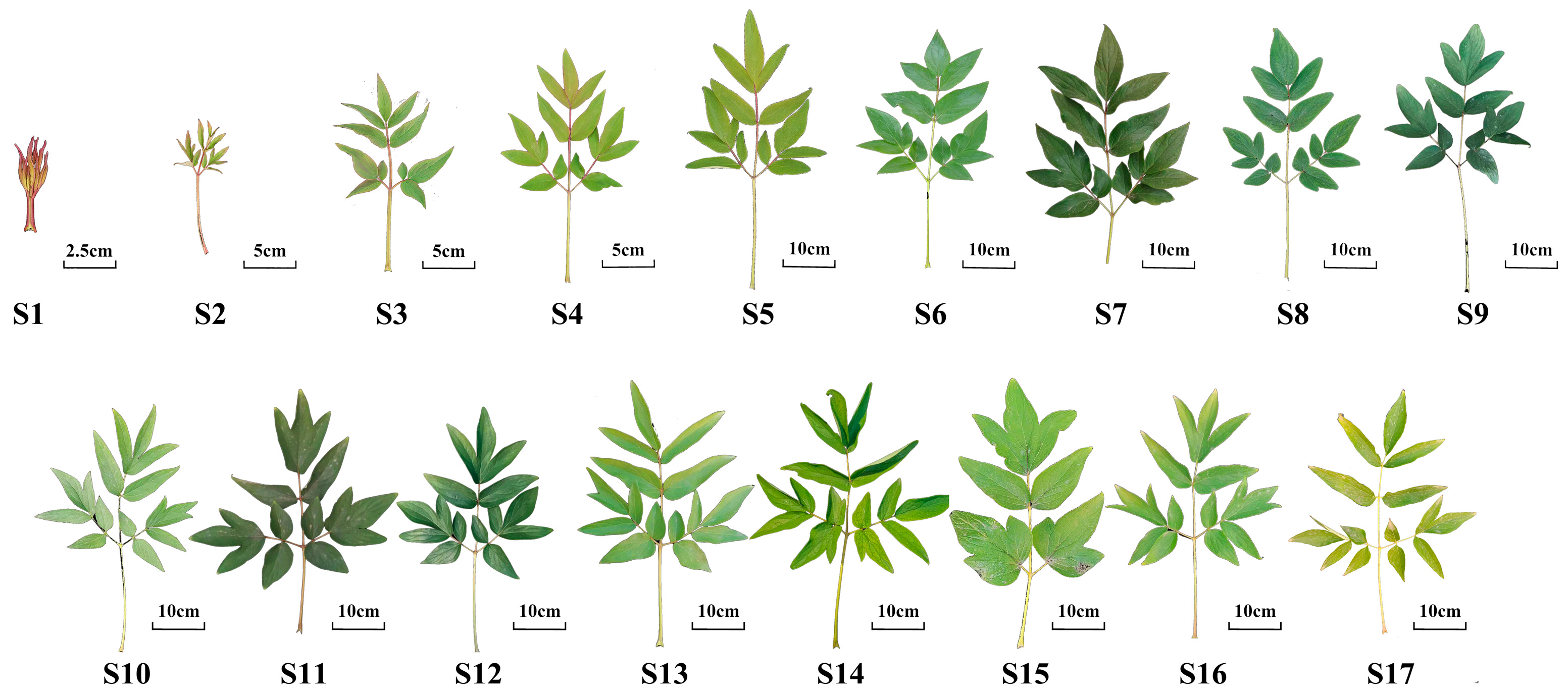
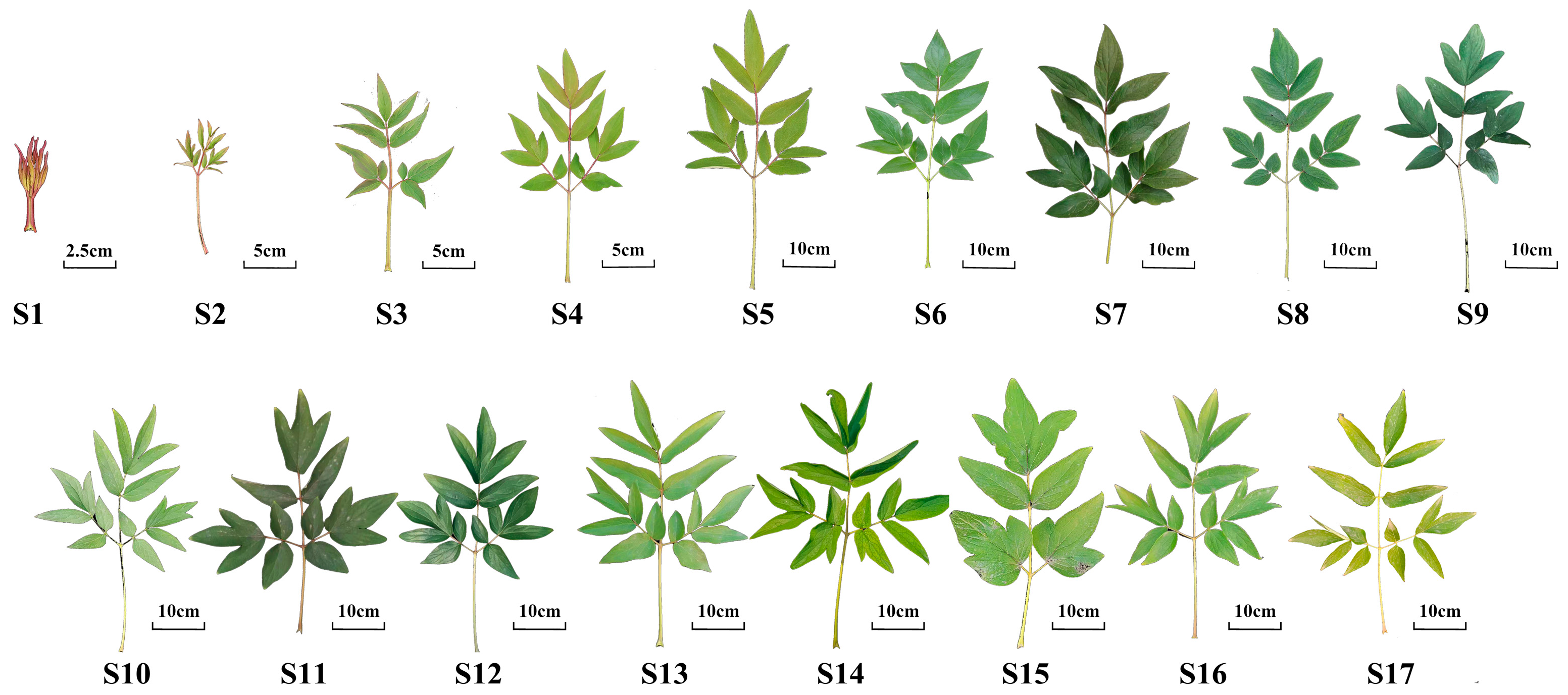
3.2. Plant Materials
3.3. Sample Preparation
3.4. Determination of Total Phenolic and Flavonoid Contents
3.5. UPLC-ESI-QTOF-MS Analysis
3.6. LC-ESI-QQQ-MS Analysis
3.7. Determination of Antioxidant Activity
3.8. Determination of Antibacterial Activity
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, S.L.; Xu, C.; Liu, J.; Yu, Y.; Wu, P.; Cheng, T.; Hong, D.Y. Out of the Pan-Himalaya: Evolutionary history of the Paeoniaceae revealed by phylogenomics. J. Syst. Evol. 2021, 59, 1170–1182. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Y.; Gao, J.Y.; Du, M.Z.; Zhang, K.; Zhang, J.L.; Xue, N.C.; Yan, M.; Qu, C.X.; Deng, R.X. HPLC-DAD analysis of 15 monoterpene glycosides in oil peony seed cakes sourced from different cultivation areas in China. Ind. Crop. Prod. 2018, 118, 259–270. [Google Scholar] [CrossRef]
- Deng, R.; Gao, J.; Yi, J.; Liu, P. Could peony seeds oil become a high-quality edible vegetable oil? The nutritional and phytochemistry profiles, extraction, health benefits, safety and value-added-products. Food Res. Int. 2022, 156, 111200. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Wang, L.S.; Shu, Q.Y.; Wu, J.; Chen, L.G.; Shao, S.; Yin, D.D. Fatty acid composition of developing tree peony (Paeonia section Moutan DC.) seeds and transcriptome analysis during seed development. BMC Genom. 2015, 16, 208. [Google Scholar] [CrossRef]
- Shahidi, F. Omega-3 fatty acids and marine oils in cardiovascular and general health: A critical overview of controversies and realities. J. Funct. Foods 2015, 19, 797–800. [Google Scholar] [CrossRef]
- Alseekh, S.; Scossa, F.; Wen, W.; Luo, J.; Yan, J.; Beleggia, R.; Klee, H.J.; Huang, S.; Papa, R.; Fernie, A.R. Domestication of crop metabolomes: Desired and unintended consequences. Trends Plant Sci. 2021, 26, 650–661. [Google Scholar] [CrossRef]
- He, C.N.; Peng, Y.; Zhang, Y.C.; Xu, L.J.; Gu, J.; Xiao, P.G. Phytochemical and biological studies of Paeoniaceae. Chem. Biodivers. 2010, 7, 805–838. [Google Scholar] [CrossRef]
- Li, P.; Shen, J.; Wang, Z.Q.; Liu, S.S.; Liu, Q.; Li, Y.; He, C.N.; Xiao, P.G. Genus Paeonia: A comprehensive review on traditional uses, phytochemistry, pharmacological activities, clinical application, and toxicology. J. Ethnopharmacol. 2021, 269, 113708. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Dalgaard, F.; Kyrø, C.; Murray, K.; Bondonno, C.P.; Lewis, J.R.; Croft, K.D.; Gislason, G.; Scalbert, A.; Cassidy, A.; et al. Flavonoid intake is associated with lower mortality in the Danish Diet Cancer and Health Cohort. Nat. Commun. 2019, 10, 3610–3651. [Google Scholar] [CrossRef]
- He, C.N.; Peng, B.; Dan, Y.; Peng, Y.; Xiao, P.G. Chemical taxonomy of tree peony species from China based on root cortex metabolic fingerprinting. Phytochemistry 2014, 107, 69–79. [Google Scholar] [CrossRef]
- Wu, S.H.; Wu, D.G.; Chen, Y.W. Chemical constituents and bioactivities of plants from the genus Paeonia. Chem. Biodivers. 2010, 7, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Wu, Q.; Yin, D.D.; Feng, C.Y.; Liu, Z.A.; Wang, L.S. Phytochemical variation among the traditional Chinese medicine Mu Dan Pi from Paeonia suffruticosa (tree peony). Phytochemistry 2018, 146, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.Z.; Tang, J.M.; Ni, J.; Zheng, T.T.; Zhou, Y.; Sun, D.Y.; Li, G.N.; Liu, P.; Niu, L.X.; Zhang, Y.L. Comprehensive metabolite profile of multi-bioactive extract from tree peony (Paeonia ostii and Paeonia rockii) fruits based on MS/MS molecular networking. Food Res. Int. 2021, 148, 110609. [Google Scholar] [CrossRef]
- He, J.Y.; Dong, Y.Q.; Liu, X.Y.; Wan, Y.L.; Gu, T.W.; Zhou, X.F.; Liu, M.H. Comparison of chemical compositions, antioxidant, and anti-photoaging activities of Paeonia suffruticosa flowers at different flowering stages. Antioxidants 2019, 8, 345. [Google Scholar] [CrossRef]
- Tong, N.N.; Zhou, X.Y.; Peng, L.P.; Liu, Z.A.; Shu, Q.Y. A comprehensive study of three species of Paeonia stem and leaf phytochemicals, and their antioxidant activities. J. Ethnopharmacol. 2021, 273, 113985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.C.; Geng, C.A.; Huang, X.Y.; Zhang, X.M.; Chen, J.J. Antidiabetic stilbenes from peony seeds with PTP1B, α-glucosidase, and DPPIV inhibitory activities. J. Agric. Food Chem. 2019, 67, 6765–6772. [Google Scholar] [CrossRef]
- Li, B.; Ge, J.; Liu, W.; Hu, D.; Li, P. Unveiling spatial metabolome of Paeonia suffruticosa and Paeonia lactiflora roots using MALDI MS imaging. New Phytol. 2021, 231, 892–902. [Google Scholar] [CrossRef]
- Zhang, X.X.; Zhai, Y.H.; Yuan, J.H.; Hu, Y.H. New insights into Paeoniaceae used as medicinal plants in China. Sci. Rep. 2019, 9, 18469. [Google Scholar] [CrossRef]
- Jiang, Y.; David, B.; Tu, P.; Barbin, Y. Recent analytical approaches in quality control of traditional Chinese medicines—A review. Anal. Chim. Acta 2010, 657, 9–18. [Google Scholar] [CrossRef]
- Neilson, E.H.; Goodger, J.Q.D.; Woodrow, I.E.; Møller, B.L. Plant chemical defense: At what cost? Trends Plant Sci. 2013, 18, 250–258. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and human health: The role of bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Zou, H.; Qiu, L.; Zheng, Y.; Yang, D.; Wang, Y. Effects of light on secondary metabolite biosynthesis in medicinal plants. Front. Plant Sci. 2021, 12, 781236. [Google Scholar] [CrossRef]
- Hao, J.; Lou, P.; Han, Y.; Zheng, L.; Lu, J.; Chen, Z.; Ni, J.; Yang, Y.; Xu, M. Ultraviolet-B irradiation increases antioxidant capacity of Pakchoi (Brassica rapa L.) by inducing flavonoid biosynthesis. Plants 2022, 11, 766. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Qu, Y.; Li, J.; Li, Y.; Ren, X.; Maffucci, K.; Li, R.; Wang, Z.; Zeng, R. In Vitro and in vivo antioxidant activities of the flowers and leaves from Paeonia rockii and identification of their antioxidant constituents by UHPLC-ESI-HRMSn via pre-column DPPH reaction. Molecules 2018, 23, 392. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, C.; Li, N.; Wu, L.; Huang, Z.; Hu, Z.; Li, X.; Qu, Z. Phytochemicals and anti-tyrosinase activities of Paeonia ostii leaves and roots. Plant Physiol. Biochem. 2022, 181, 50–60. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Modern analytical techniques in metabolomics analysis. Analyst 2012, 137, 293–300. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Chen, S.; Kong, H.; Lu, X.; Li, Y.; Yin, P.; Zeng, Z.; Xu, G. Pseudotargeted metabolomics method and its application in serum biomarker discovery for hepatocellular carcinoma based on ultra high-performance liquid chromatography/triple quadrupole mass spectrometry. Anal. Chem. 2013, 85, 8326–8333. [Google Scholar] [CrossRef]
- Bai, Z.Z.; Yu, R.; Tang, J.M.; Zhou, Y.; Zheng, T.T.; Ni, J.; Sun, D.Y.; Liu, P.; Niu, L.X.; Zhang, Y.L. Comparative investigation on metabolites and biological activities of Paeonia ostii stamens from different geographical regions of China. Ind. Crop. Prod. 2021, 172, 114038. [Google Scholar] [CrossRef]
- Cheng, J.; Chen, M.; Wan, H.; Chen, X.; Li, C.; Zhu, J.; Liu, Q.; Xu, G.; Yi, L. Paeoniflorin exerts antidepressant-like effects through enhancing neuronal FGF-2 by microglial inactivation. J. Ethnopharmacol. 2021, 274, 114046. [Google Scholar] [CrossRef] [PubMed]
- Manayi, A.; Omidpanah, S.; Barreca, D.; Ficarra, S.; Daglia, M.; Nabavi, S.F.; Nabavi, S.M. Neuroprotective effects of paeoniflorin in neurodegenerative diseases of the central nervous system. Phytochem. Rev. 2017, 16, 1173–1181. [Google Scholar] [CrossRef]
- Niu, K.; Liu, Y.; Zhou, Z.; Wu, X.; Wang, H.; Yan, J. Antitumor effects of Paeoniflorin on hippo signaling pathway in gastric cancer cells. J. Oncol. 2021, 2021, 4724938. [Google Scholar] [CrossRef]
- Qian, W.; Zhang, J.; Wang, W.; Wang, T.; Liu, M.; Yang, M.; Sun, Z.; Li, X.; Li, Y. Antimicrobial and antibiofilm activities of paeoniflorin against carbapenem-resistantKlebsiella pneumoniae. J. Appl. Microbiol. 2020, 128, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Torres-León, C.; Ventura-Sobrevilla, J.; Serna-Cock, L.; Ascacio-Valdés, J.A.; Contreras-Esquivel, J.; Aguilar, C.N. Pentagalloylglucose (PGG): A valuable phenolic compound with functional properties. J. Funct. Foods 2017, 37, 176–189. [Google Scholar] [CrossRef]
- Xin, Z.; Yang, W.; Niu, L.; Zhang, Y. Comprehensive metabolite profile uncovers the bioactive components, antioxidant and antibacterial activities in wild tree peony leaves. Int. J. Mol. Sci. 2023, 24, 10609. [Google Scholar] [CrossRef]
- Cetin Cakmak, K.; Gülçin, O. Anticholinergic and antioxidant activities of usnic acid-an activity-structure insight. Toxicol. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef]
- Fan, H.; Sun, L.; Yang, L.; Zhou, J.; Yin, P.; Li, K.; Li, X.; Xue, Q.; Liu, Y. Assessment of the bioactive phenolic composition of Acer truncatum seed coat as a byproduct of seed oil. Ind. Crop. Prod. 2018, 118, 11–19. [Google Scholar] [CrossRef]
- Fan, J.L.; Zhu, W.X.; Kang, H.B.; Ma, H.L.; Tao, G.J. Flavonoid constituents and antioxidant capacity in flowers of different Zhongyuan tree penoy cultivars. J. Funct. Foods 2012, 4, 147–157. [Google Scholar] [CrossRef]
- Blando, F.; Russo, R.; Negro, C.; De Bellis, L.; Frassinetti, S. Antimicrobial and antibiofilm activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. Cladode Polyphenolic Extracts. Antioxidants 2019, 8, 117. [Google Scholar] [CrossRef]
- Vilvert, J.C.; de Freitas, S.T.; Dos Santos, L.F.; Ribeiro, S.T.D.; Veloso, C.M. Phenolic compounds in acerola fruit and by-products: An overview on identification, quantification, influencing factors, and biological properties. J. Food Meas. Charact. 2023. [Google Scholar] [CrossRef]
- Bai, Z.Z.; Ni, J.; Tang, J.M.; Sun, D.Y.; Yan, Z.G.; Zhang, J.; Niu, L.X.; Zhang, Y.L. Bioactive components, antioxidant and antimicrobial activities of Paeonia rockii fruit during development. Food Chem. 2021, 343, 128444. [Google Scholar] [CrossRef] [PubMed]
- Taslimi, P.; Gulçin, I. Antioxidant and anticholinergic properties of olivetol. J. Food Biochem. 2018, 42, e12516. [Google Scholar] [CrossRef]
- Li, X.C.; Chen, D.F.; Mai, Y.; Wen, B.; Wang, X.Z. Concordance between antioxidant activities in vitro and chemical components of Radix Astragali (Huangqi). Nat. Prod. Res. 2012, 26, 1050. [Google Scholar] [CrossRef] [PubMed]
- Spínola, V.; Pinto, J.; Castilho, P.C. Identification and quantification of phenolic compounds of selected fruits from Madeira Island by HPLC-DAD–ESI-MSn and screening for their antioxidant activity. Food Chem. 2015, 173, 14–30. [Google Scholar] [CrossRef]
- Klančnik, A.; Piskernik, S.; Jeršek, B.; Možina, S.S. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J. Microbiol. Meth. 2010, 81, 121–126. [Google Scholar] [CrossRef]
- Trevino, V.; Yanez-Garza, I.L.; Rodriguez-Lopez, C.E.; Urrea-Lopez, R.; Garza-Rodriguez, M.L.; Barrera-Saldana, H.A.; Tamez-Pena, J.G.; Winkler, R.; Diaz, D.R. GridMass: A fast two-dimensional feature detection method for LC/MS. J. Mass Spectrom. 2015, 50, 165–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Gram-Negative Bacteria | Gram-Positive Bacteria | ||||
|---|---|---|---|---|---|---|
| E. coli | P. vulgaris | S. typhimurium | S. aureus | S. hemolytis-β | L. monocytogenes | |
| S1 | 0.39 | 6.25 | 3.13 | 3.13 | 0.39 | 3.13 |
| S2 | 0.39 | 6.25 | 3.13 | 3.13 | 0.39 | 6.25 |
| S3 | 0.39 | 6.25 | 3.13 | 12.50 | 0.39 | 6.25 |
| S4 | 1.56 | 6.25 | 3.13 | 12.50 | 0.39 | 6.25 |
| S5 | 1.56 | 6.25 | 3.13 | 6.25 | 0.39 | 12.50 |
| S6 | 3.13 | 3.13 | 3.13 | 6.25 | 0.20 | 12.50 |
| S7 | 1.56 | 3.13 | 1.56 | 3.13 | 0.39 | 6.25 |
| S8 | 1.56 | 6.25 | 6.25 | 6.25 | 0.39 | 12.50 |
| S9 | 1.56 | 6.25 | 6.25 | 6.25 | 0.39 | 6.25 |
| S10 | 1.56 | 6.25 | 3.13 | 3.13 | 0.39 | 12.50 |
| S11 | 1.56 | 3.13 | 3.13 | 3.13 | 0.39 | 6.25 |
| S12 | 1.56 | 6.25 | 3.13 | 6.25 | 0.78 | 6.25 |
| S13 | 1.56 | 6.25 | 3.13 | 3.13 | 0.39 | 6.25 |
| S14 | 1.56 | 3.13 | 3.13 | 3.13 | 0.78 | 6.25 |
| S15 | 1.56 | 6.25 | 3.13 | 3.13 | 0.39 | 6.25 |
| S16 | 3.13 | 6.26 | 3.13 | 6.26 | 0.78 | 6.26 |
| S17 | 3.14 | 3.14 | 3.14 | 6.28 | 0.39 | 6.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Z.; Tang, J.; Li, Y.; Li, Z.; Gu, S.; Deng, L.; Zhang, Y. Integrated Metabolomics Approach Reveals the Dynamic Variations of Metabolites and Bioactivities in Paeonia ostii ‘Feng Dan’ Leaves during Development. Int. J. Mol. Sci. 2024, 25, 1059. https://doi.org/10.3390/ijms25021059
Bai Z, Tang J, Li Y, Li Z, Gu S, Deng L, Zhang Y. Integrated Metabolomics Approach Reveals the Dynamic Variations of Metabolites and Bioactivities in Paeonia ostii ‘Feng Dan’ Leaves during Development. International Journal of Molecular Sciences. 2024; 25(2):1059. https://doi.org/10.3390/ijms25021059
Chicago/Turabian StyleBai, Zhangzhen, Junman Tang, Yajie Li, Zhuoning Li, Siyi Gu, Lu Deng, and Yanlong Zhang. 2024. "Integrated Metabolomics Approach Reveals the Dynamic Variations of Metabolites and Bioactivities in Paeonia ostii ‘Feng Dan’ Leaves during Development" International Journal of Molecular Sciences 25, no. 2: 1059. https://doi.org/10.3390/ijms25021059
APA StyleBai, Z., Tang, J., Li, Y., Li, Z., Gu, S., Deng, L., & Zhang, Y. (2024). Integrated Metabolomics Approach Reveals the Dynamic Variations of Metabolites and Bioactivities in Paeonia ostii ‘Feng Dan’ Leaves during Development. International Journal of Molecular Sciences, 25(2), 1059. https://doi.org/10.3390/ijms25021059