

Exercise Reduces Glucose Intolerance, Cardiac Inflammation and Adipose Tissue Dysfunction in Psammomys obesus Exposed to Short Photoperiod and High Energy Diet
 , , and
, , and
Abstract
1. Introduction
2. Results
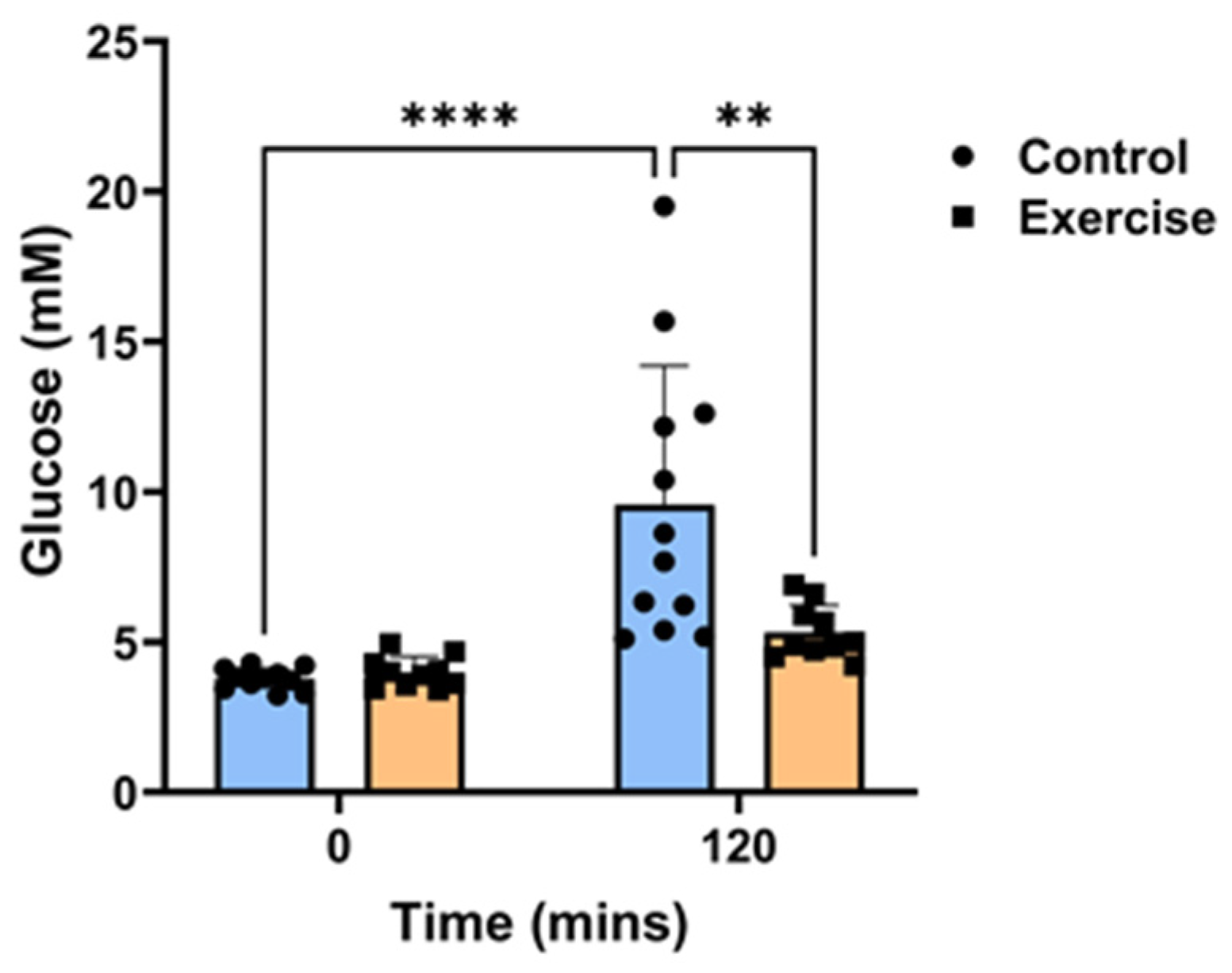
2.1. Exercise Prevented Circadian Disruption-Induced Glucose Intolerance
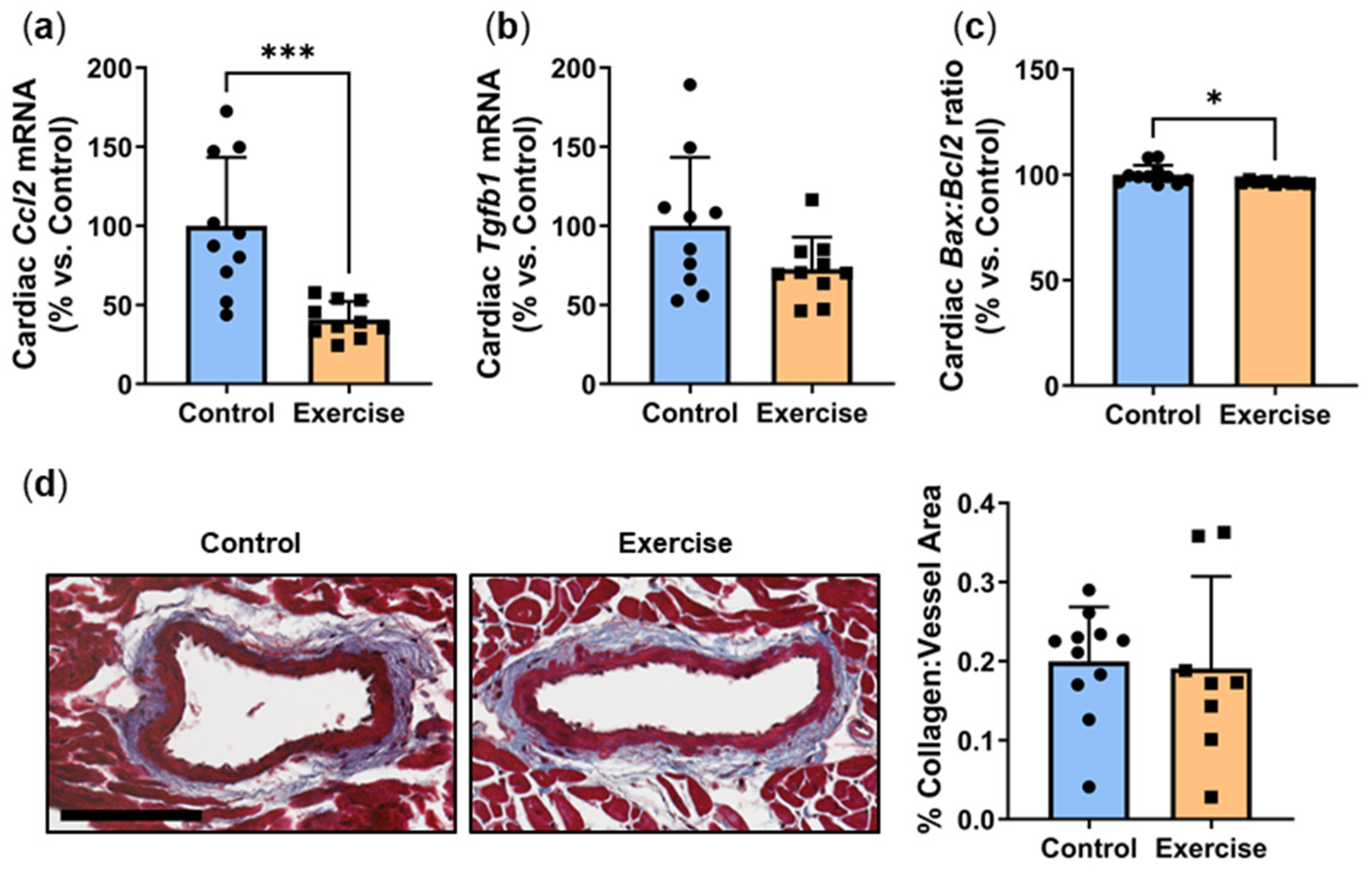
2.2. Exercise Reduced Inflammation and Apoptosis in Cardiac Tissue but Had No Effect on Myocardial Perivascular Fibrosis
2.3. Exercise Increased the Heart:Body Weight Ratio and Hypertrophy Marker Myh7:Myh6, Yet Reduced Transcription Factor Gata4
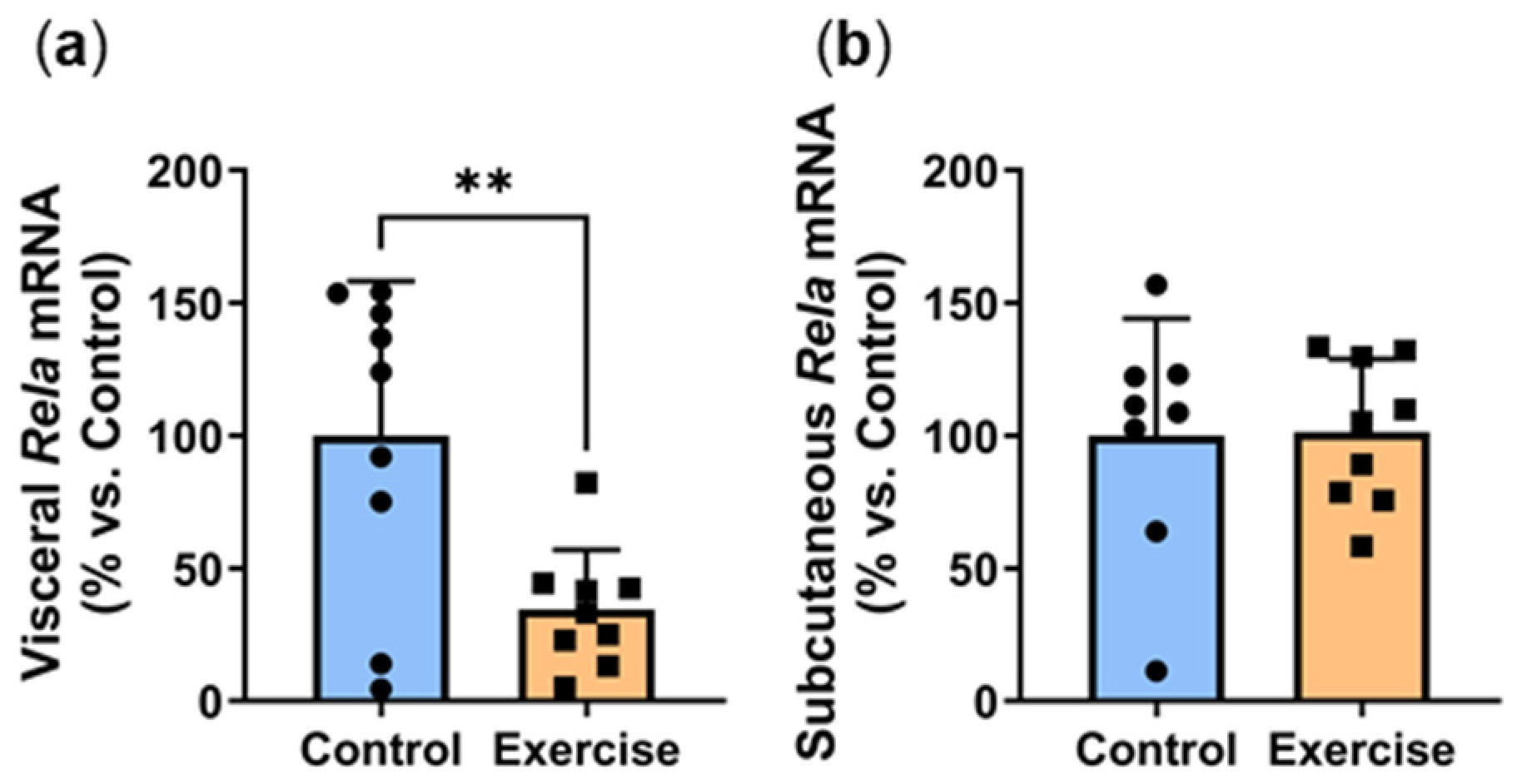
2.4. Exercise Reduced Visceral Adipose Expression of the Inflammatory Transcription Factor Rela
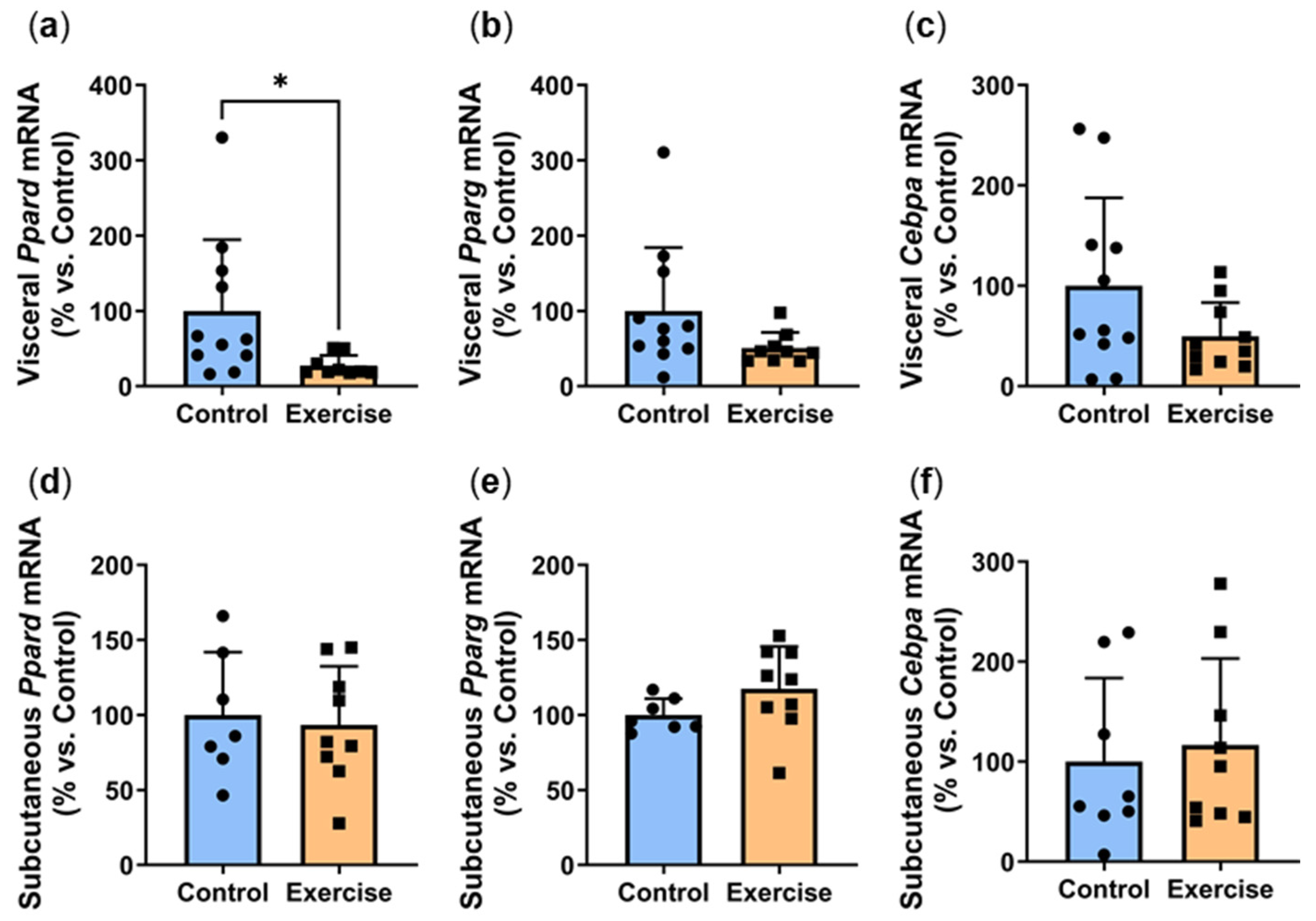
2.5. Exercise Reduced Visceral Adipose Expression of Adipogenesis Markers
2.6. Exercise Reduced Visceral Adipose Expression of Browning Markers
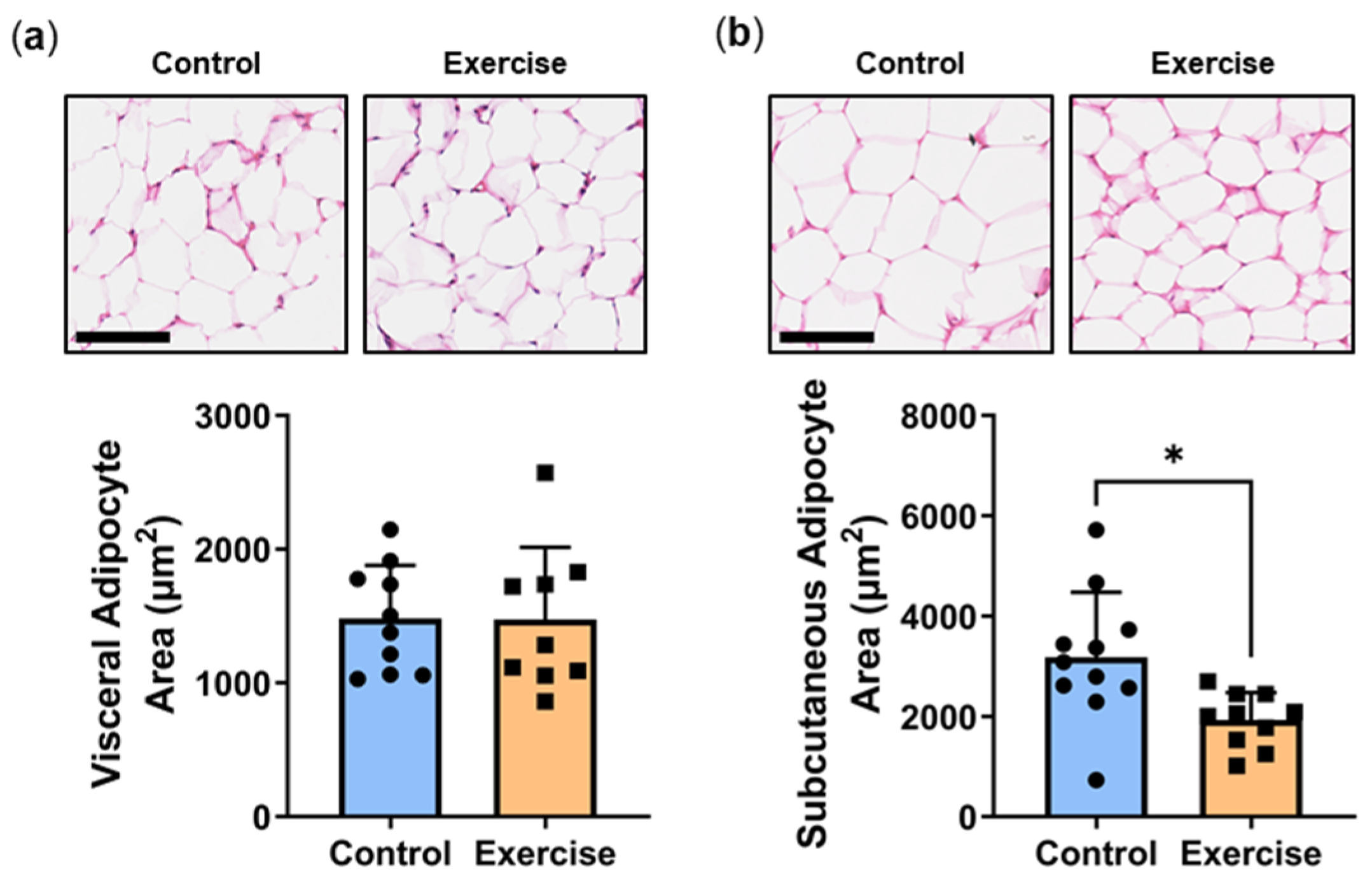
2.7. Exercise Reduced Subcutaneous Adipocyte Area
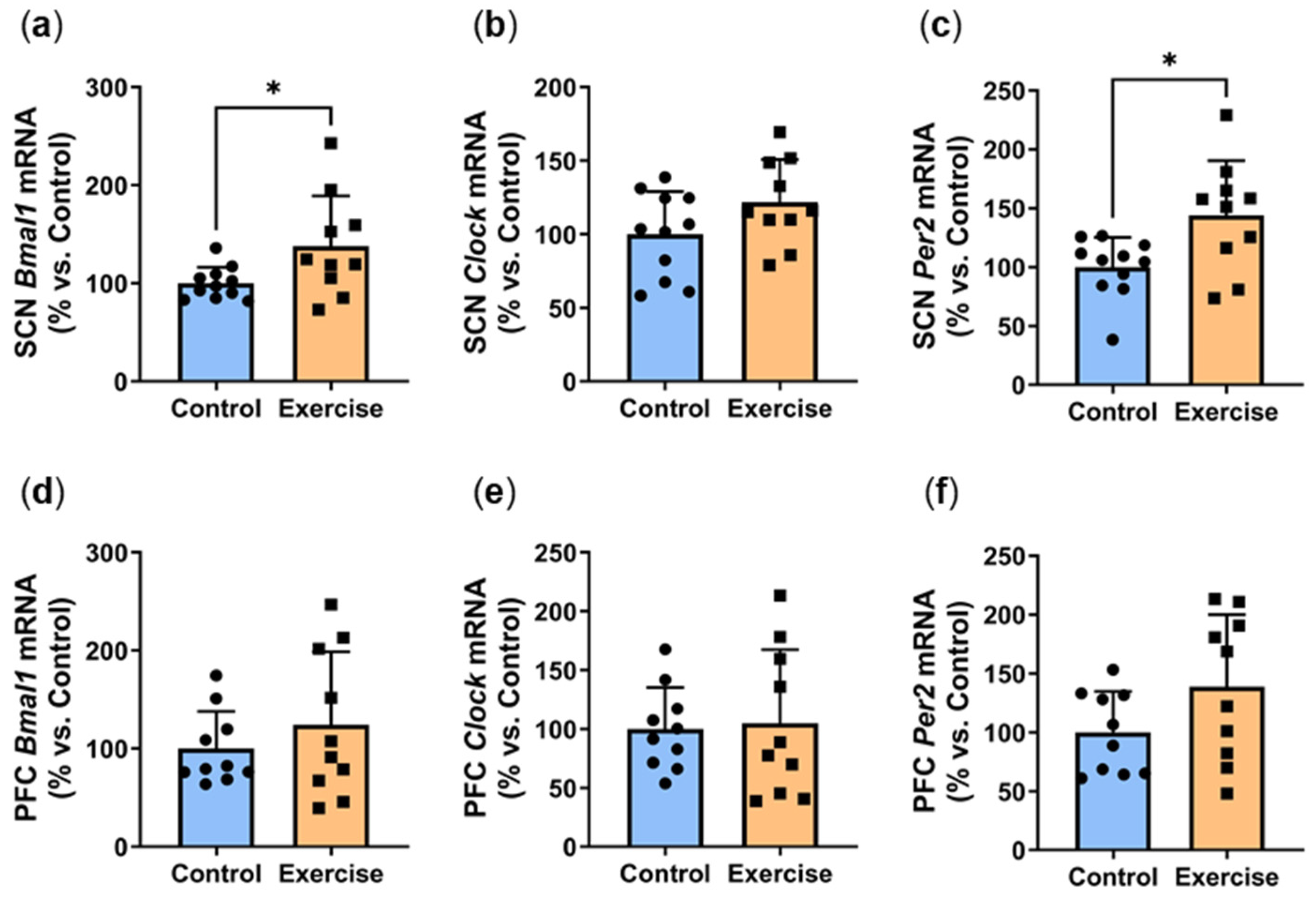
2.8. Exercise Increased Circadian Genes at ZT7
3. Discussion
4. Materials and Methods
4.1. Animal Studies
4.2. Gene Expression Analysis
4.3. Histological Analysis
4.4. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Remchak, M.E.; Piersol, K.L.; Bhatti, S.; Spaeth, A.M.; Buckman, J.F.; Malin, S.K. Considerations for Maximizing the Exercise “Drug” to Combat Insulin Resistance: Role of Nutrition, Sleep, and Alcohol. Nutrients 2021, 13, 1708. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.V.; Bhatt, D.L.; Barsness, G.W.; Beatty, A.L.; Deedwania, P.C.; Inzucchi, S.E.; Kosiborod, M.; Leiter, L.A.; Lipska, K.J.; Newman, J.D.; et al. Clinical Management of Stable Coronary Artery Disease in Patients With Type 2 Diabetes Mellitus: A Scientific Statement From the American Heart Association. Circulation 2020, 141, e779–e806. [Google Scholar] [CrossRef] [PubMed]
- Dunlay, S.M.; Givertz, M.M.; Aguilar, D.; Allen, L.A.; Chan, M.; Desai, A.S.; Deswal, A.; Dickson, V.V.; Kosiborod, M.N.; Lekavich, C.L.; et al. Type 2 Diabetes Mellitus and Heart Failure: A Scientific Statement From the American Heart Association and the Heart Failure Society of America: This statement does not represent an update of the 2017 ACC/AHA/HFSA heart failure guideline update. Circulation 2019, 140, e294–e324. [Google Scholar] [CrossRef] [PubMed]
- Nankivell, V.A.; Tan, J.T.M.; Wilsdon, L.A.; Morrison, K.R.; Bilu, C.; Psaltis, P.J.; Zimmet, P.; Kronfeld-Schor, N.; Nicholls, S.J.; Bursill, C.A.; et al. Circadian disruption by short light exposure and a high energy diet impairs glucose tolerance and increases cardiac fibrosis in Psammomys obesus. Sci. Rep. 2021, 11, 9673. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.T.M.; Nankivell, V.A.; Bilu, C.; Shemesh, T.; Nicholls, S.J.; Zimmet, P.; Kronfeld-Schor, N.; Brown, A.; Bursill, C.A. High-Energy Diet and Shorter Light Exposure Drives Markers of Adipocyte Dysfunction in Visceral and Subcutaneous Adipose Depots of Psammomys obesus. Int. J. Mol. Sci. 2019, 20, 6291. [Google Scholar] [CrossRef] [PubMed]
- Panda, S. The arrival of circadian medicine. Nat. Rev. Endocrinol. 2019, 15, 67–69. [Google Scholar] [CrossRef]
- Carroll, R.G.; Timmons, G.A.; Cervantes-Silva, M.P.; Kennedy, O.D.; Curtis, A.M. Immunometabolism around the Clock. Trends Mol. Med. 2019, 25, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.C.; Marin, T.C.; Azevedo, L.; Rosa-Silva, J.M.; Shea, S.A.; Thosar, S.S. Chronobiology of Exercise: Evaluating the Best Time to Exercise for Greater Cardiovascular and Metabolic Benefits. Compr. Physiol. 2022, 12, 3621–3639. [Google Scholar] [CrossRef] [PubMed]
- Crnko, S.; Cour, M.; Van Laake, L.W.; Lecour, S. Vasculature on the clock: Circadian rhythm and vascular dysfunction. Vascul. Pharmacol. 2018, 108, 1–7. [Google Scholar] [CrossRef]
- Allerton, T.D.; Savoie, J.J.; Fitch, M.D.; Hellerstein, M.K.; Stephens, J.M.; White, U. Exercise reduced the formation of new adipocytes in the adipose tissue of mice in vivo. PLoS ONE 2021, 16, e0244804. [Google Scholar] [CrossRef]
- Stanford, K.I.; Middelbeek, R.J.; Goodyear, L.J. Exercise Effects on White Adipose Tissue: Beiging and Metabolic Adaptations. Diabetes 2015, 64, 2361–2368. [Google Scholar] [CrossRef]
- Steidle-Kloc, E.; Schönfelder, M.; Müller, E.; Sixt, S.; Schuler, G.; Patsch, W.; Niebauer, J. Does exercise training impact clock genes in patients with coronary artery disease and type 2 diabetes mellitus? Eur. J. Prev. Cardiol. 2016, 23, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.T.L.; Samuels, R.E.; Bano-Otalora, B.; Belle, M.D.C.; Wegner, S.; Guilding, C.; Northeast, R.C.; Loudon, A.S.I.; Gigg, J.; Piggins, H.D. Timed daily exercise remodels circadian rhythms in mice. Commun. Biol. 2021, 4, 761. [Google Scholar] [CrossRef] [PubMed]
- Bilu, C.; Zimmet, P.; Vishnevskia-Dai, V.; Einat, H.; Agam, G.; Grossman, E.; Kronfeld-Schor, N. Diurnality, Type 2 Diabetes, and Depressive-Like Behavior. J. Biol. Rhythms 2019, 34, 69–83. [Google Scholar] [CrossRef]
- Russo, I.; Frangogiannis, N.G. Diabetes-associated cardiac fibrosis: Cellular effectors, molecular mechanisms and therapeutic opportunities. J. Mol. Cell Cardiol. 2016, 90, 84–93. [Google Scholar] [CrossRef]
- Bilu, C.; Einat, H.; Barak, O.; Zimmet, P.; Vishnevskia-Dai, V.; Govrin, A.; Agam, G.; Kronfeld-Schor, N. Linking type 2 diabetes mellitus, cardiac hypertrophy and depression in a diurnal animal model. Sci. Rep. 2019, 9, 11865. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Endocrine and signalling role of adipose tissue: New perspectives on fat. Acta Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Hsu, C.H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPalpha induces adipogenesis through PPARgamma: A unified pathway. Genes. Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef]
- Contreras, C.; Nogueiras, R.; Dieguez, C.; Medina-Gomez, G.; Lopez, M. Hypothalamus and thermogenesis: Heating the BAT, browning the WAT. Mol. Cell Endocrinol. 2016, 438, 107–115. [Google Scholar] [CrossRef]
- Lizcano, F. The Beige Adipocyte as a Therapy for Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 5058. [Google Scholar] [CrossRef]
- Laforest, S.; Labrecque, J.; Michaud, A.; Cianflone, K.; Tchernof, A. Adipocyte size as a determinant of metabolic disease and adipose tissue dysfunction. Crit. Rev. Clin. Lab. Sci. 2015, 52, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Bilu, C.; Einat, H.; Zimmet, P.; Vishnevskia-Dai, V.; Schwartz, W.J.; Kronfeld-Schor, N. Beneficial effects of voluntary wheel running on activity rhythms, metabolic state, and affect in a diurnal model of circadian disruption. Sci. Rep. 2022, 12, 2434. [Google Scholar] [CrossRef] [PubMed]
- Serin, Y.; Acar Tek, N. Effect of Circadian Rhythm on Metabolic Processes and the Regulation of Energy Balance. Ann. Nutr. Metab. 2019, 74, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Sklifasovskaya, A.P.; Blagonravov, M.L.; Ryabinina, A.Y.; Azova, M.M.; Goryachev, V.A. Expression of Bax and Bcl-2 Proteins in Left-Ventricular Cardiomyocytes in Wistar-Kyoto and SHR Rats with Insulin-Dependent Diabetes Mellitus. Bull. Exp. Biol. Med. 2021, 171, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Pei, Z.; Ji, J.; Gao, Y.; Wang, H.; Wu, Y.; Yang, J.; Yang, Q.; Zhang, L. Exercise reduces hyperlipidemia-induced cardiac damage in apolipoprotein E-deficient mice via its effects against inflammation and oxidative stress. Sci. Rep. 2023, 13, 9134. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tu, W.; Li, X.; Li, C.; Lu, J.; Dai, P.; Chen, Y.; Gu, M.; Li, M.; Jiang, S.; et al. Exercise improves cardiac function and attenuates myocardial inflammation and apoptosis by regulating APJ/STAT3 in mice with stroke. Life Sci. 2023, 332, 122041. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhou, Z.; Peng, X.; Sun, C.; Yang, D.; Li, C.; Zhu, R.; Zhang, P.; Zheng, L.; Tang, C. Cardioprotective responses to aerobic exercise-induced physiological hypertrophy in zebrafish heart. J. Physiol. Sci. 2021, 71, 33. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, O.; Bhardwaj Ratan, D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz Bruce, A.; Druid, H.; et al. Evidence for Cardiomyocyte Renewal in Humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef]
- Broderick, T.L.; Jankowski, M.; Wang, D.; Danalache, B.A.; Parrott, C.R.; Gutkowska, J. Downregulation in GATA4 and Downstream Structural and Contractile Genes in the db/db Mouse Heart. ISRN Endocrinol. 2012, 2012, 736860. [Google Scholar] [CrossRef]
- Han, M.; Yang, Z.; Sayed, D.; He, M.; Gao, S.; Lin, L.; Yoon, S.; Abdellatif, M. GATA4 expression is primarily regulated via a miR-26b-dependent post-transcriptional mechanism during cardiac hypertrophy. Cardiovasc. Res. 2012, 93, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Colby, J.K.; Zuo, X.; Jaoude, J.; Wei, D.; Shureiqi, I. The Role of PPAR-δ in Metabolism, Inflammation, and Cancer: Many Characters of a Critical Transcription Factor. Int. J. Mol. Sci. 2018, 19, 3339. [Google Scholar] [CrossRef] [PubMed]
- Christodoulides, C.; Vidal-Puig, A. PPARs and adipocyte function. Mol. Cell Endocrinol. 2010, 318, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, D.; Zhao, W.; Xu, L. Deciphering the Roles of PPARgamma in Adipocytes via Dynamic Change of Transcription Complex. Front. Endocrinol. 2018, 9, 473. [Google Scholar] [CrossRef] [PubMed]
- Gollisch, K.S.; Brandauer, J.; Jessen, N.; Toyoda, T.; Nayer, A.; Hirshman, M.F.; Goodyear, L.J. Effects of exercise training on subcutaneous and visceral adipose tissue in normal- and high-fat diet-fed rats. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E495–E504. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, Q.; Zheng, L.; Shou, J.; Zhuang, S.; Xiao, W.; Chen, P. Depot-specific adaption of adipose tissue for different exercise approaches in high-fat diet/streptozocin-induced diabetic mice. Front. Physiol. 2023, 14, 1189528. [Google Scholar] [CrossRef] [PubMed]
- Kolnes, K.J.; Petersen, M.H.; Lien-Iversen, T.; Hojlund, K.; Jensen, J. Effect of Exercise Training on Fat Loss-Energetic Perspectives and the Role of Improved Adipose Tissue Function and Body Fat Distribution. Front. Physiol. 2021, 12, 737709. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, B.R.; Martinez-Huenchullan, S.F.; McLennan, S.V.; Twigg, S.M.; Williams, P.F. Exercise induces favorable metabolic changes in white adipose tissue preventing high-fat diet obesity. Physiol. Rep. 2021, 9, e14929. [Google Scholar] [CrossRef]
- Takahashi, J.S.; Hong, H.K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Edery, I. Circadian rhythms in a nutshell. Physiol. Genom. 2000, 3, 59–74. [Google Scholar] [CrossRef]
- Latimer, M.N.; Young, M.E. Circadian Governance of Cardiac Growth. Cells 2022, 11, 1494. [Google Scholar] [CrossRef] [PubMed]
- Hogenesch, J.B.; Gu, Y.Z.; Jain, S.; Bradfield, C.A. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 1998, 95, 5474–5479. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Zhao, Z.; Lee, C.C. Interactive Organization of the Circadian Core Regulators PER2, BMAL1, CLOCK and PML. Sci. Rep. 2016, 6, 29174. [Google Scholar] [CrossRef] [PubMed]
- Civelek, E.; Ozturk Civelek, D.; Akyel, Y.K.; Kaleli Durman, D.; Okyar, A. Circadian Dysfunction in Adipose Tissue: Chronotherapy in Metabolic Diseases. Biology 2023, 12, 1077. [Google Scholar] [CrossRef] [PubMed]
- Viswambharan, H.; Carvas, J.M.; Antic, V.; Marecic, A.; Jud, C.; Zaugg, C.E.; Ming, X.F.; Montani, J.P.; Albrecht, U.; Yang, Z. Mutation of the circadian clock gene Per2 alters vascular endothelial function. Circulation 2007, 115, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Wen, M.S.; Wang, H.W.; Hsieh, I.C.; Li, Y.; Liu, P.Y.; Lin, F.C.; Liao, J.K. Increased vascular senescence and impaired endothelial progenitor cell function mediated by mutation of circadian gene Per2. Circulation 2008, 118, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Sahraoui, A.; Dewachter, C.; de Medina, G.; Naeije, R.; Aouichat Bouguerra, S.; Dewachter, L. Myocardial Structural and Biological Anomalies Induced by High Fat Diet in Psammomys obesus Gerbils. PLoS ONE 2016, 11, e0148117. [Google Scholar] [CrossRef]
- Spolding, B.; Connor, T.; Wittmer, C.; Abreu, L.L.; Kaspi, A.; Ziemann, M.; Kaur, G.; Cooper, A.; Morrison, S.; Lee, S.; et al. Rapid development of non-alcoholic steatohepatitis in Psammomys obesus (Israeli sand rat). PLoS ONE 2014, 9, e92656. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.T.; Prosser, H.C.; Dunn, L.L.; Vanags, L.Z.; Ridiandries, A.; Tsatralis, T.; Lecce, L.; Clayton, Z.E.; Yuen, S.C.; Robertson, S.; et al. High-Density Lipoproteins Rescue Diabetes-Impaired Angiogenesis via Scavenger Receptor Class B Type I. Diabetes 2016, 65, 3091–3103. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Exercise | |
|---|---|---|
| Body weight (g) | 217.9 ± 45.1 | 200.0 ± 32.9 |
| Fasting baseline glucose (mM) | 3.8 ± 0.4 | 4.0 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.T.M.; Price, K.J.; Fanshaw, S.-R.; Bilu, C.; Pham, Q.T.; Pham, A.; Sandeman, L.; Nankivell, V.A.; Solly, E.L.; Kronfeld-Schor, N.; et al. Exercise Reduces Glucose Intolerance, Cardiac Inflammation and Adipose Tissue Dysfunction in Psammomys obesus Exposed to Short Photoperiod and High Energy Diet. Int. J. Mol. Sci. 2024, 25, 7756. https://doi.org/10.3390/ijms25147756
Tan JTM, Price KJ, Fanshaw S-R, Bilu C, Pham QT, Pham A, Sandeman L, Nankivell VA, Solly EL, Kronfeld-Schor N, et al. Exercise Reduces Glucose Intolerance, Cardiac Inflammation and Adipose Tissue Dysfunction in Psammomys obesus Exposed to Short Photoperiod and High Energy Diet. International Journal of Molecular Sciences. 2024; 25(14):7756. https://doi.org/10.3390/ijms25147756
Chicago/Turabian StyleTan, Joanne T. M., Kiara J. Price, Sarah-Rose Fanshaw, Carmel Bilu, Quang Tuan Pham, Anthony Pham, Lauren Sandeman, Victoria A. Nankivell, Emma L. Solly, Noga Kronfeld-Schor, and et al. 2024. "Exercise Reduces Glucose Intolerance, Cardiac Inflammation and Adipose Tissue Dysfunction in Psammomys obesus Exposed to Short Photoperiod and High Energy Diet" International Journal of Molecular Sciences 25, no. 14: 7756. https://doi.org/10.3390/ijms25147756
APA StyleTan, J. T. M., Price, K. J., Fanshaw, S.-R., Bilu, C., Pham, Q. T., Pham, A., Sandeman, L., Nankivell, V. A., Solly, E. L., Kronfeld-Schor, N., & Bursill, C. A. (2024). Exercise Reduces Glucose Intolerance, Cardiac Inflammation and Adipose Tissue Dysfunction in Psammomys obesus Exposed to Short Photoperiod and High Energy Diet. International Journal of Molecular Sciences, 25(14), 7756. https://doi.org/10.3390/ijms25147756