
Oil Palm AP2 Subfamily Gene EgAP2.25 Improves Salt Stress Tolerance in Transgenic Tobacco Plants

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
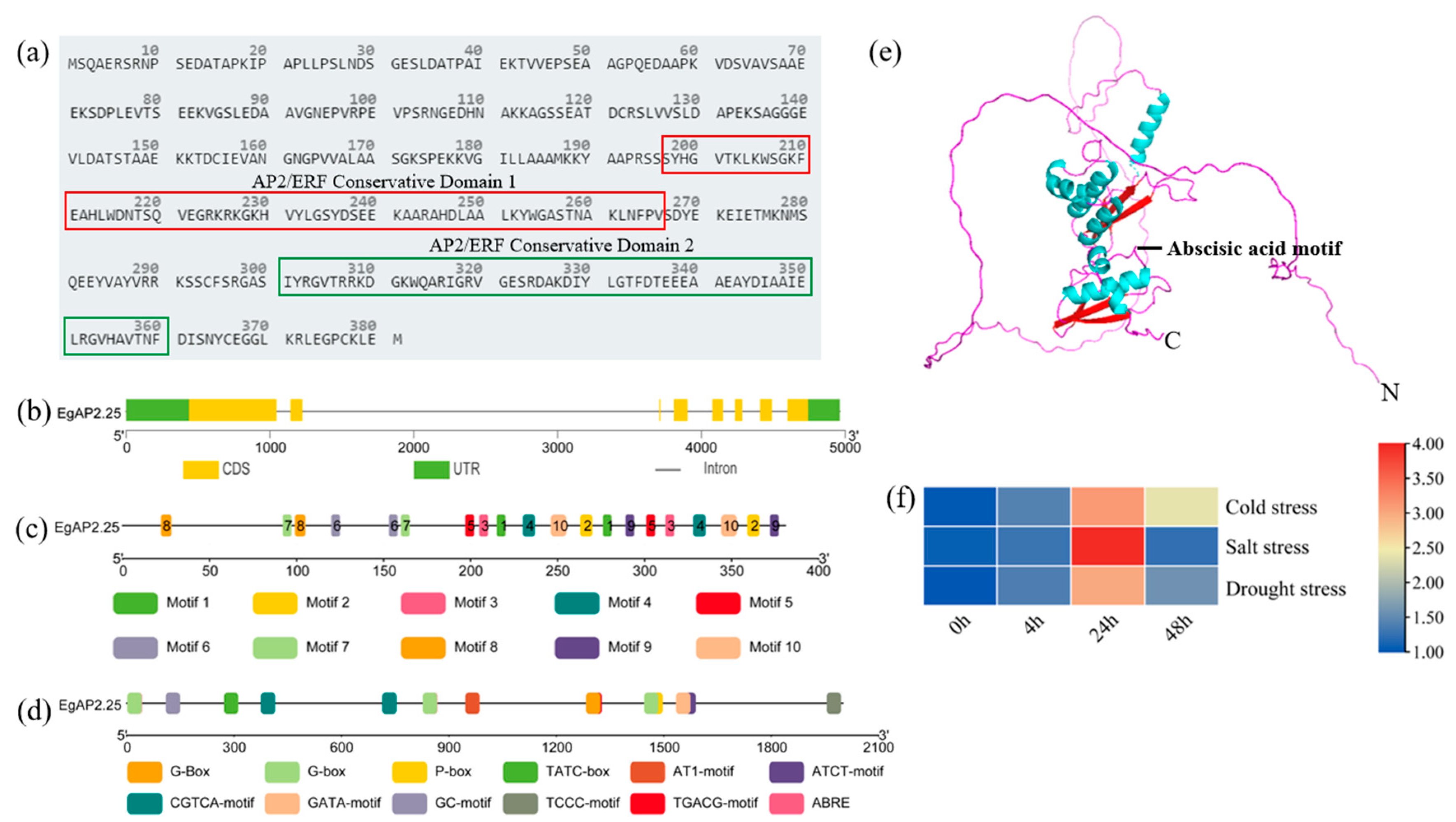
2.1. Sequence and Character of Oil Palm EgAP2.25
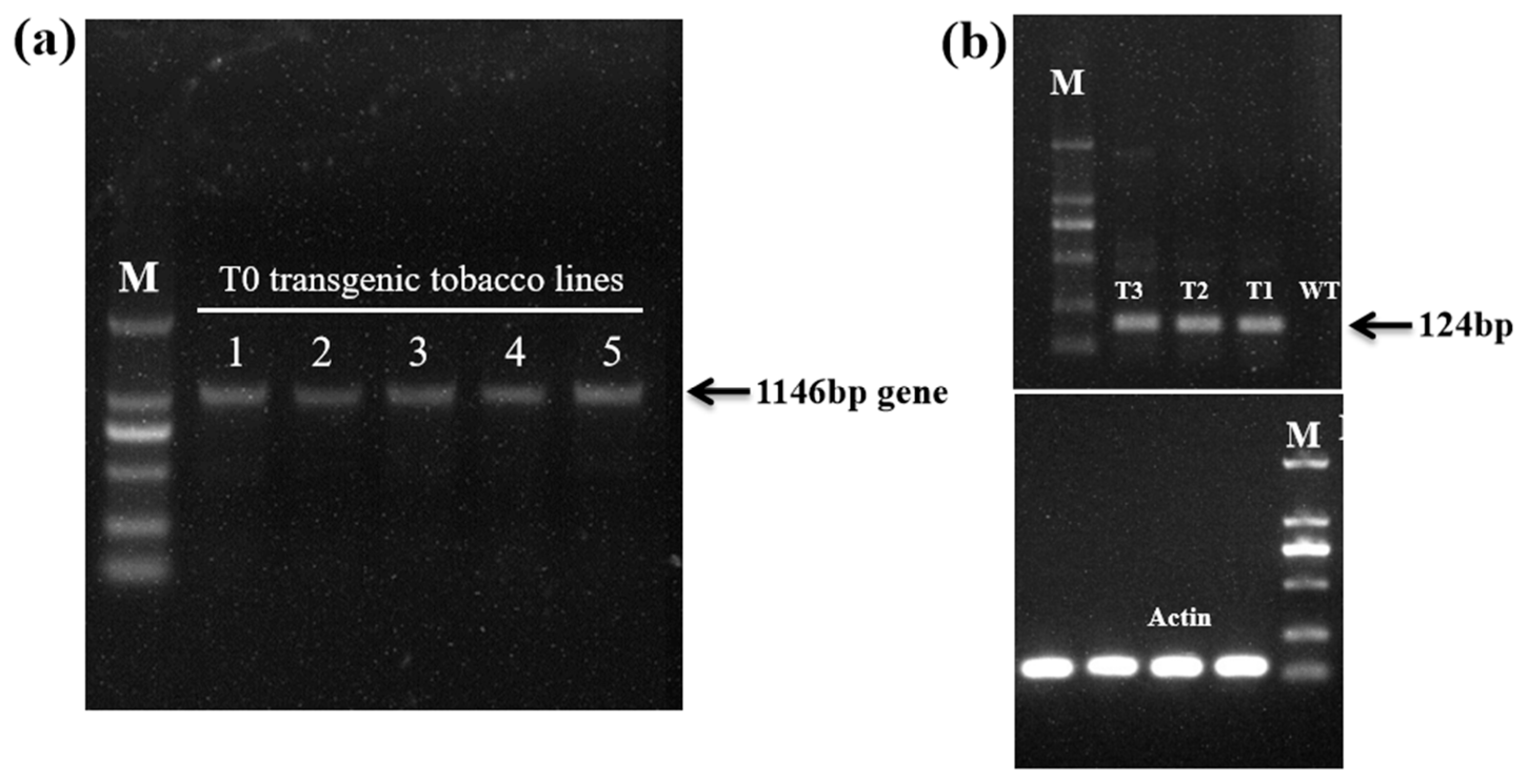
2.2. Generation of Transgenic Tobacco Plants Over-Expressing EgAP2.25
2.3. Analysis of Salinity Stress on the Germination Index and Phytomass of Tobacco Lines
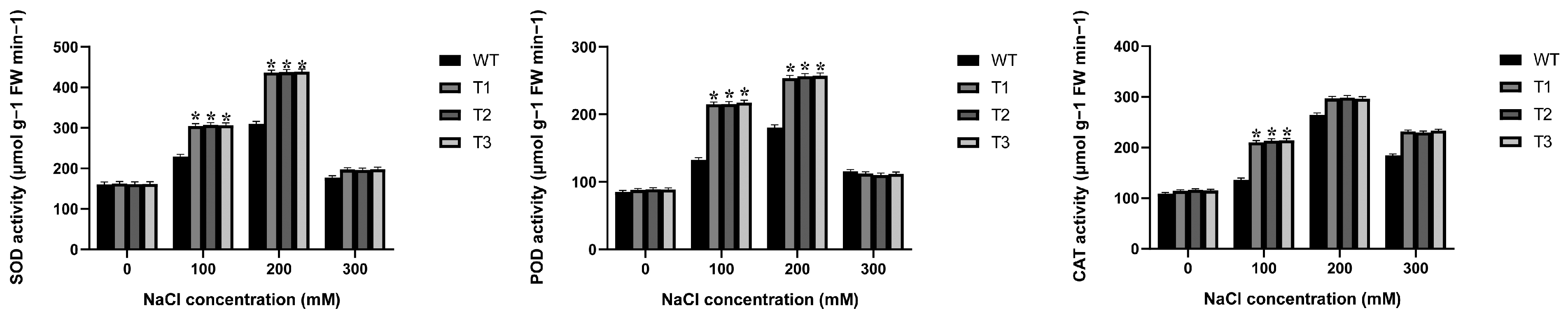
2.4. Antioxidant Enzyme Activities in EgAP2.25 Transgenic Tobacco
2.5. Physiological and Biochemical Variation in EgAP2.25 Transgenic Tobacco
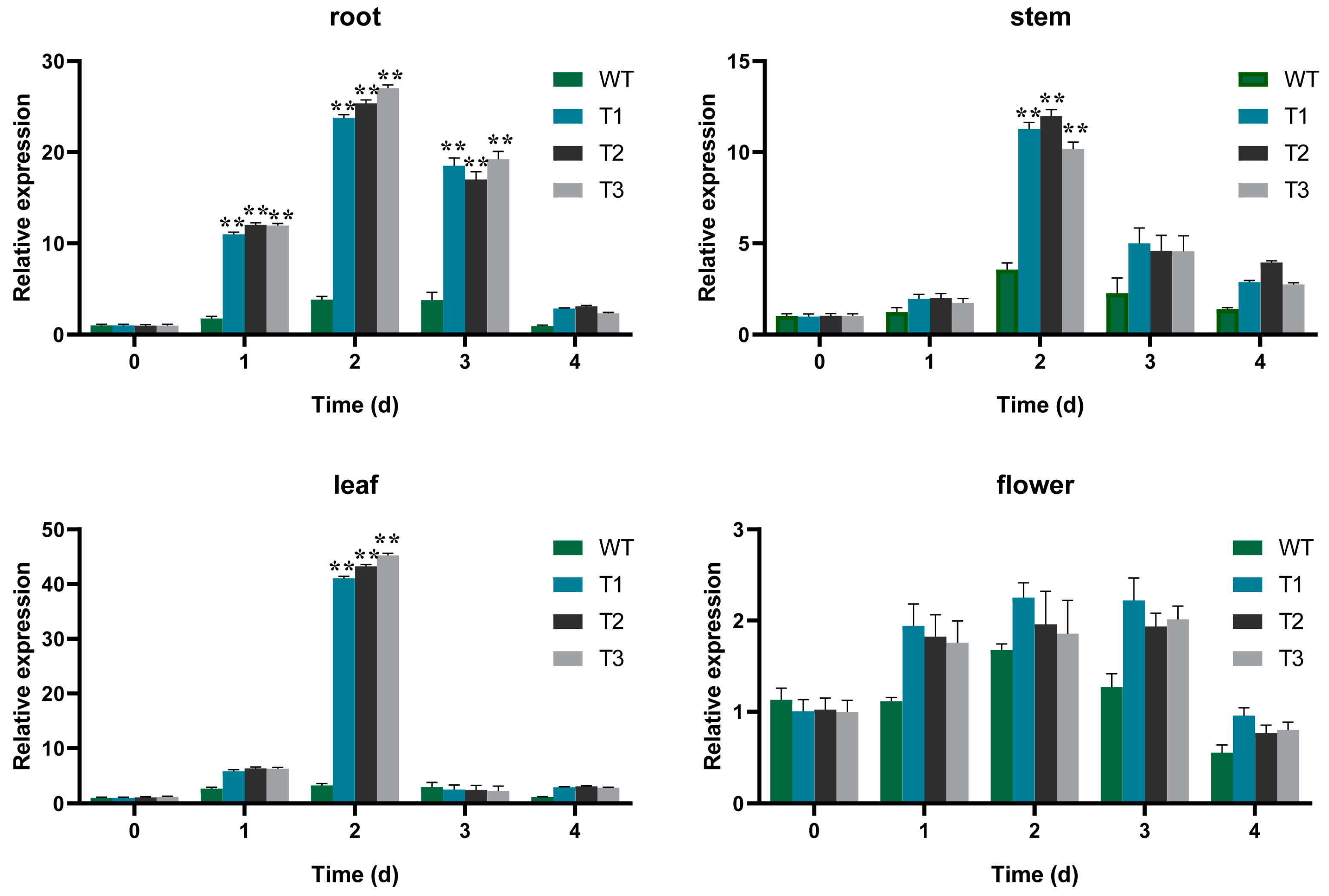
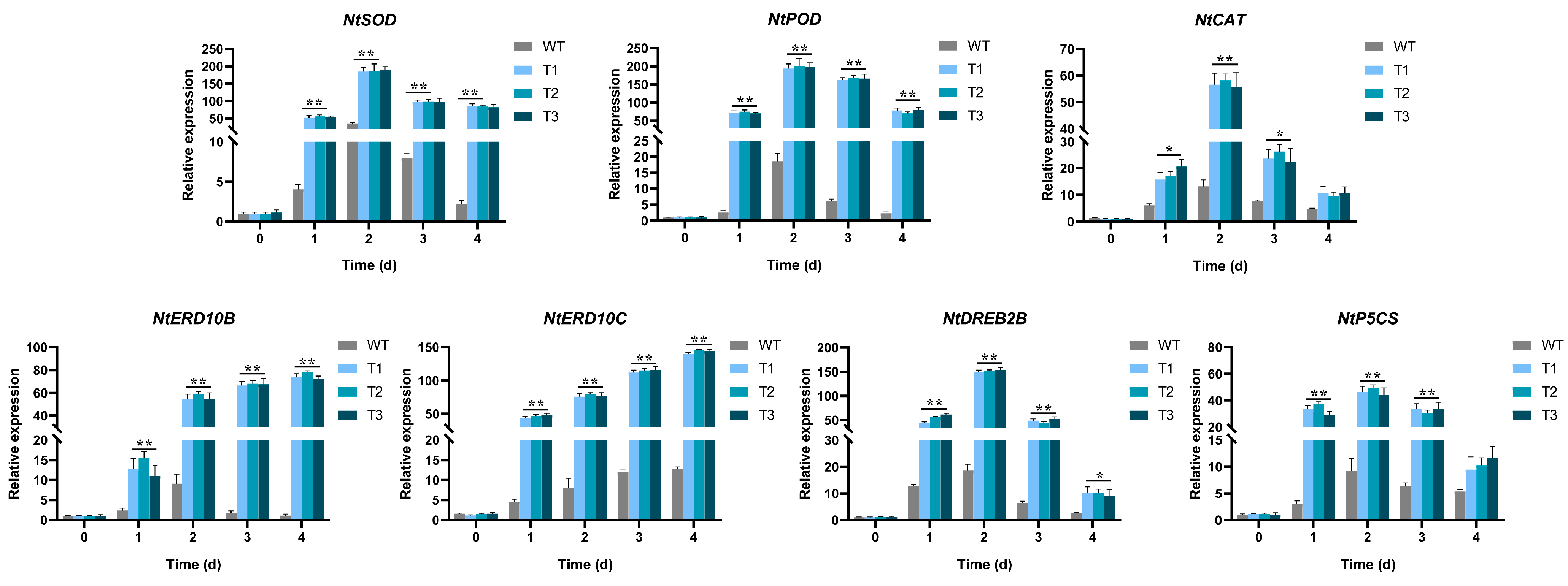
2.6. Expression Profiles of EgAP2.25 and Salinity Stress Marker Genes in Transgenic Tobacco Plants
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Condition, and Salinity Treatment
4.2. Cloning and Amplification of EgAP2.25 Gene
4.3. Tobacco Transformation
4.4. Salinity Stress Treatment to Transgenic Tobacco Plants
4.5. Physiological and Biochemical Analyses of WT and EgAP2.25 Transgenic Tobacco
4.6. Expression Analysis of EgAP2.25 and Salinity Stress Marker Genes via Quantitative Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Van-Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The role of melatonin in salt stress reponses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A new insight of salt stress signaling in plant. Mol. Cells. 2019, 2, 447–459. [Google Scholar]
- Avin-Wittenberg, T. Autophagy and its role in plant abiotic stress management. Plant Cell Environ. 2019, 42, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Avsian-Kretchmer, O.; Gueta-Dahan, Y.; Lev-Yadun, S.; Gollop, R.; Ben-Hayyim, G. The salt-stress signal transduction pathway that activates the gpx1 promoter is mediated by intracellular H2O2 different from the pathway induced by extracellular H2O2. Plant Physiol. 2004, 135, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Deal, R.B.; Henikoff, S. Histone variants and modifications in plant gene regulation. Curr. Opin. Plant Biol. 2011, 14, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Roychoudhury, A. Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress. Protoplasma 2017, 254, 3–16. [Google Scholar] [CrossRef]
- Mzid, R.; Zorrig, W.; Ben-Ayed, R.; Ben-Hamed, K.; Ayadi, M.; Damak, Y.; Lauvergeat, V.; Hanana, M. The grapevine VvWRKY2 gene enhances salt and osmotic stress tolerance in transgenic Nicotiana tabacum. 3 Biotech 2018, 8, 277. [Google Scholar] [CrossRef]
- Zhou, L.X.; Yarra, R.; Jin, L.F.; Cao, H.X. Genome-wide identification and expression analysis of MYB gene family in oil palm (Elaeis guineensis Jacq) under abiotic stress conditions. Environ. Exp. Bot. 2020, 180, 104245. [Google Scholar] [CrossRef]
- Shinde, H.; Dudhate, A.; Tsugama, D.; Gupta, S.K.; Liu, S.; Takano, T. Pearl millet stress-responsive NAC transcription factor PgNAC21 enhances salinity stress tolerance in Arabidopsis. Plant Physiol. Biochem. 2019, 135, 546–553. [Google Scholar] [CrossRef]
- Zhou, L.X.; Yarra, R.; Yang, Y.D.; Liu, Y.J.; Yang, M.D.; Cao, H.X. The oil palm R2R3-MYB subfamily genes EgMYB111 and EgMYB157 improve multiple abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Rep. 2022, 41, 377–393. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.X.; Yarra, R. Genome-wide identification and characterization of AP2/ERF transcription factor family genes in oil palm under abiotic stress conditions. Int. J. Mol. Sci. 2021, 22, 2821. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Jalmi, S.K.; Bhagat, P.K.; Verma, N.; Sinha, A.K. A bHLH transcription factor MYC2 imparts salt intolerance by regulating proline biosynthesis in Arabidopsis. FEBS J. 2020, 287, 2560–2576. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Vishal, B.; Khoo, K.; Rajappa, S.; Loh, C.S.; Kumar, P.P. Expression of AoNHX1 increases salt tolerance of rice and Arabidopsis and bHLH transcription factors regulate AtNHX1 and AtNHX6 in Arabidopsis. Plant Cell Rep. 2019, 38, 1299–1315. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.X.; Yarra, R. Genome-wide identification and expression analysis of bZIP transcription factors in oil palm (Elaeis guineensis Jacq) under abiotic stress. Protoplasma 2022, 259, 469–483. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishna, M.; SinghSidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Shoji, T.; Yuan, L. ERF Gene Clusters: Working Together to Regulate Metabolism. Trends Plant Sci. 2021, 26, 23–32. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Ngatia, J.N.; Wang, Y.; Khoso, M.A.; Farooq, U.; Chen, S. AP2/ERF an important cold stress-related transcription factor family in plants: A review. Physiol. Mol. Biol. Plants 2021, 27, 1953–1968. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, P.N.; Matias, F.; Martínez-Andújar, C.; Martinez-Melgarejo, P.A.; Prudencio, Á.S.; Galeano, E.; Pérez-Alfocea, F.; Carrer, H. Overexpression of TgERF1, a Transcription Factor from Tectona grandis, Increases Tolerance to Drought and Salt Stress in Tobacco. Int. J. Mol. Sci. 2023, 24, 4149. [Google Scholar] [CrossRef]
- Kavas, M.; Gökdemir, G.; Seçgin, Z.; Bakhsh, A. Ectopic expression of common bean ERF transcription factor PvERF35 promotes salt stress tolerance in tobacco. Plant Biol. 2020, 22, 1102–1112. [Google Scholar] [CrossRef]
- Kidokoro, S.; Watanabe, K.; Ohori, T.; Moriwaki, T.; Maruyama, K.; Mizoi, J.; Myint-Phyu, S.; Htwe, N.; Fujita, Y.; Sekita, S.; et al. Soybean DREB1/CBF-type transcription factors function in heat and drought as well as cold stress-responsive gene expression. Plant J. 2015, 81, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Jisha, V.; Dampanaboina, L.; Vadassery, J.; Mithöfer, A.; Kappara, S.; Ramanan, R. Over-expression of an AP2/ERF type transcription factor OsEREBP1 confers biotic and abiotic stress tolerance in rice. PLoS ONE 2015, 10, e0127831. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wei, X.; Yin, C.; Zhou, H.; Yan, J.; He, W.; Yan, J.; Li, H. ZmEREB57 regulates OPDA synthesis and enhances salt stress tolerance through two distinct signalling pathways in Zea mays. Plant Cell Environ. 2023, 46, 2867–2883. [Google Scholar] [CrossRef] [PubMed]
- Barcelos, E.; Rios, S.A.; Cunha, R.N.; Lopes, R.; Motoike, S.Y.; Babiychuk, E.; Skirycz, A.; Kushnir, S. Oil palm natural diversity and the potential for yield improvement. Front. Plant Sci. 2015, 6, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Mejia-Alvarado, F.S.; Botero-Rozo, D.; Araque, L.; Bayona, C.; Herrera-Corzo, M.; Montoya, C.; Ayala-Díaz, I.; Romero, H.M. Molecular network of the oil palm root response to aluminum stress. BMC Plant Biol. 2023, 23, 346. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, 874–878. [Google Scholar] [CrossRef]
- Wang, L.; Tanveer, M.; Wang, H.; Arnao, M.B. Melatonin as a key regulator in seed germination under abiotic stress. J. Pineal Res. 2024, 76, e12937. [Google Scholar] [CrossRef]
- Bahariah, B.; Parveez, G.K.A.; Masani, M.Y.A.; Khalid, N.A. The use of mannose selection system for gene transfer in tobacco plants (Nicotiana tabacum L.), a model plant for oil palm transformation. J. Oil Plam. Res. 2014, 26, 154–162. [Google Scholar]
- Masura, S.S.; Parveez, G.K.A.; Ti, L.L.E. Isolation and characterization of an oil palm constitutive promoter derived from a translationally control tumor protein (TCTP) gene. Plant Physiol. Biochem. 2011, 49, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Seok, H.Y.; Tran, H.T.; Lee, S.Y.; Moon, Y.H. AtERF71/HRE2 an Arabidopsis AP2/ERF transcription factor gene contains both positive and negative Cis-regulatory elements in its promoter region involved in hypoxia and salt stress responses. Int. J. Mol. Sci. 2022, 23, 5310. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, R.S.; Jangale, B.L.; Krishna, B.; Sane, P.V. Improved abiotic stress tolerance in Arabidopsis by constitutive active form of a banana DREB2 type transcription factor MaDREB20. CA, than its native form, MaDREB20. Protoplasma 2023, 260, 671–690. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van-Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 2, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cheng, Z.; Yao, W.; Gao, Y.; Fan, G.; Guo, Q.; Zhou, B.; Jiang, T. Over-expression of PagERF072 from poplar improves salt tolerance. Int. J. Mol. Sci. 2022, 23, 10707. [Google Scholar] [CrossRef]
- Shapiguzov, A.; Kangasjärvi, J. Studying plant stress reactions in vivo by PAM chlorophyll fluorescence imaging. Methods Mol. Biol. 2022, 2526, 43–61. [Google Scholar]
- Selim, S.; Abuelsoud, W.; Al-Sanea, M.M.; AbdElgawad, H. Elevated CO2 differently suppresses the arsenic oxide nanoparticles-induced stress in C3 (Hordeum vulgare) and C4 (Zea maize) plants via altered homeostasis in metabolites specifically proline and anthocyanin metabolism. Plant Physiol. Biochem. 2021, 166, 235–245. [Google Scholar] [CrossRef]
- Funck, D.; Baumgarten, L.; Stift, M.; Von-Wirén, N.; Schönemann, L. Differential contribution of P5CS isoforms to stress tolerance in Arabidopsis. Front. Plant Sci. 2020, 11, 565134. [Google Scholar] [CrossRef] [PubMed]
- Sanwal, S.K.; Kesh, H.; Kumar, A.; Dubey, B.K.; Khar, A.; Rouphael, Y.; Kumar, P. Salt tolerance potential in Onion: Confirmation through physiological and biochemical traits. Plants 2022, 11, 3325. [Google Scholar] [CrossRef] [PubMed]
- Fal, S.; Aasfar, A.; Rabie, R.; Smouni, A.; Arroussi, H.E. Salt induced oxidative stress alters physiological biochemical and metabolomic responses of green microalga Chlamydomonas reinhardtii. Heliyon 2022, 8, e08811. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Kang, Y.; Lee, S.; Jeon, D.H.; Seo, S.; Lee, T.H.; Kim, C. Physio-chemical and co-expression network analysis associated with salt stress in sorghum. Front. Biosci. (Landmark Ed.) 2022, 27, 55. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice (Oryza sativa L.) encode transcription activators that function in drought- high-salt- and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Daszkowska-Golec, A. Arabidopsis seed germination under abiotic stress as a concert of action of phytohormones. OMICS 2011, 15, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Foolad, M.R.; Subbiah, P.; Zhang, L. Common QTL affect the rate of tomato seed germination under different stress and nonstress conditions. Int. J. Plant Genom. 2007, 2007, 97386. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.; Lin, R. The role of light in regulating seed dormancy and germination. J. Integr. Plant Biol. 2020, 62, 1310–1326. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Zhou, J.H.; Li, Z.; Qiao, J.; Quan, R.; Wang, J.; Huang, R.; Qin, H. Salt and ABA response ERF1 improves seed germination and salt tolerance by repressing ABA signaling in rice. Plant Physiol. 2022, 189, 1110–1127. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.M.; Zhou, M.L.; Wang, D.; Tang, Y.X.; Lin, M.; Wu, Y.M. Gene LcAP2/ERF107 enhances tolerance to salt stress. Protein Pept. Lett. 2016, 23, 442–449. [Google Scholar] [CrossRef]
- Horch, R.B.; Fry, J.E.; Hoffmann, N.L.; Rogers, S.G.; Fraley, R.T. A simple method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhou, D.; Wang, J.; Li, Y.; Liu, Y.; Ning, Y. Evaluation of the toxicity effects of microplastics and cadmium on earthworms. Sci. Total Environ. 2022, 836, 155747. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Zeng, J.; He, Y. Analysis of quantitative trait loci affecting chlorophyll content of rice leaves in a double haploid population and two backcross populations. Gene 2014, 536, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Cao, H.; Zeng, X.; Wu, Q.; Li, Q.; Martin, J.J.J.; Fu, D.; Liu, X.; Li, X.; Li, R.; et al. Oil Palm AP2 Subfamily Gene EgAP2.25 Improves Salt Stress Tolerance in Transgenic Tobacco Plants. Int. J. Mol. Sci. 2024, 25, 5621. https://doi.org/10.3390/ijms25115621
Zhou L, Cao H, Zeng X, Wu Q, Li Q, Martin JJJ, Fu D, Liu X, Li X, Li R, et al. Oil Palm AP2 Subfamily Gene EgAP2.25 Improves Salt Stress Tolerance in Transgenic Tobacco Plants. International Journal of Molecular Sciences. 2024; 25(11):5621. https://doi.org/10.3390/ijms25115621
Chicago/Turabian StyleZhou, Lixia, Hongxing Cao, Xianhai Zeng, Qiufei Wu, Qihong Li, Jerome Jeyakumar John Martin, Dengqiang Fu, Xiaoyu Liu, Xinyu Li, Rui Li, and et al. 2024. "Oil Palm AP2 Subfamily Gene EgAP2.25 Improves Salt Stress Tolerance in Transgenic Tobacco Plants" International Journal of Molecular Sciences 25, no. 11: 5621. https://doi.org/10.3390/ijms25115621
APA StyleZhou, L., Cao, H., Zeng, X., Wu, Q., Li, Q., Martin, J. J. J., Fu, D., Liu, X., Li, X., Li, R., & Ye, J. (2024). Oil Palm AP2 Subfamily Gene EgAP2.25 Improves Salt Stress Tolerance in Transgenic Tobacco Plants. International Journal of Molecular Sciences, 25(11), 5621. https://doi.org/10.3390/ijms25115621