
Functional Genome Analyses Reveal the Molecular Basis of Oil Accumulation in Developing Seeds of Castor Beans



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
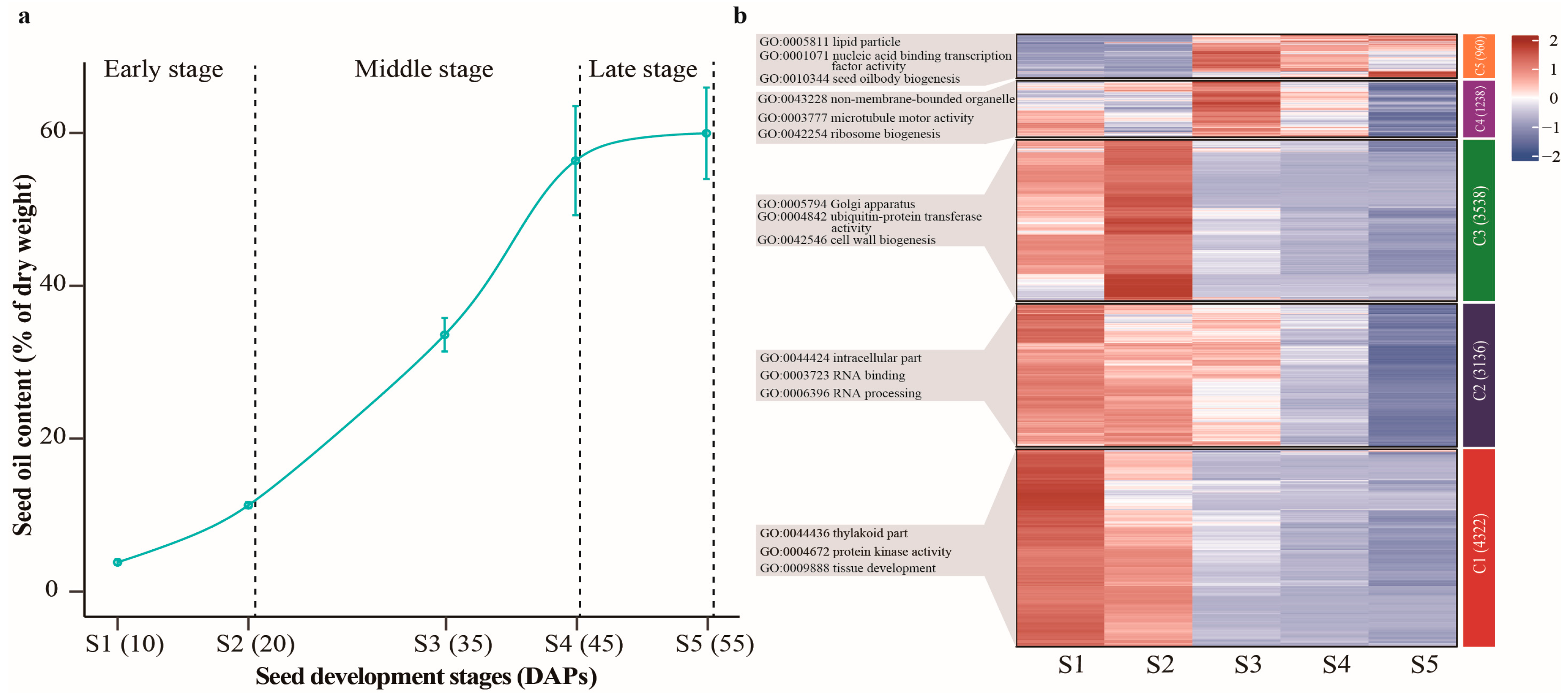
2.1. Dynamics of Oil Accumulation in Developing Seeds of Castor Beans
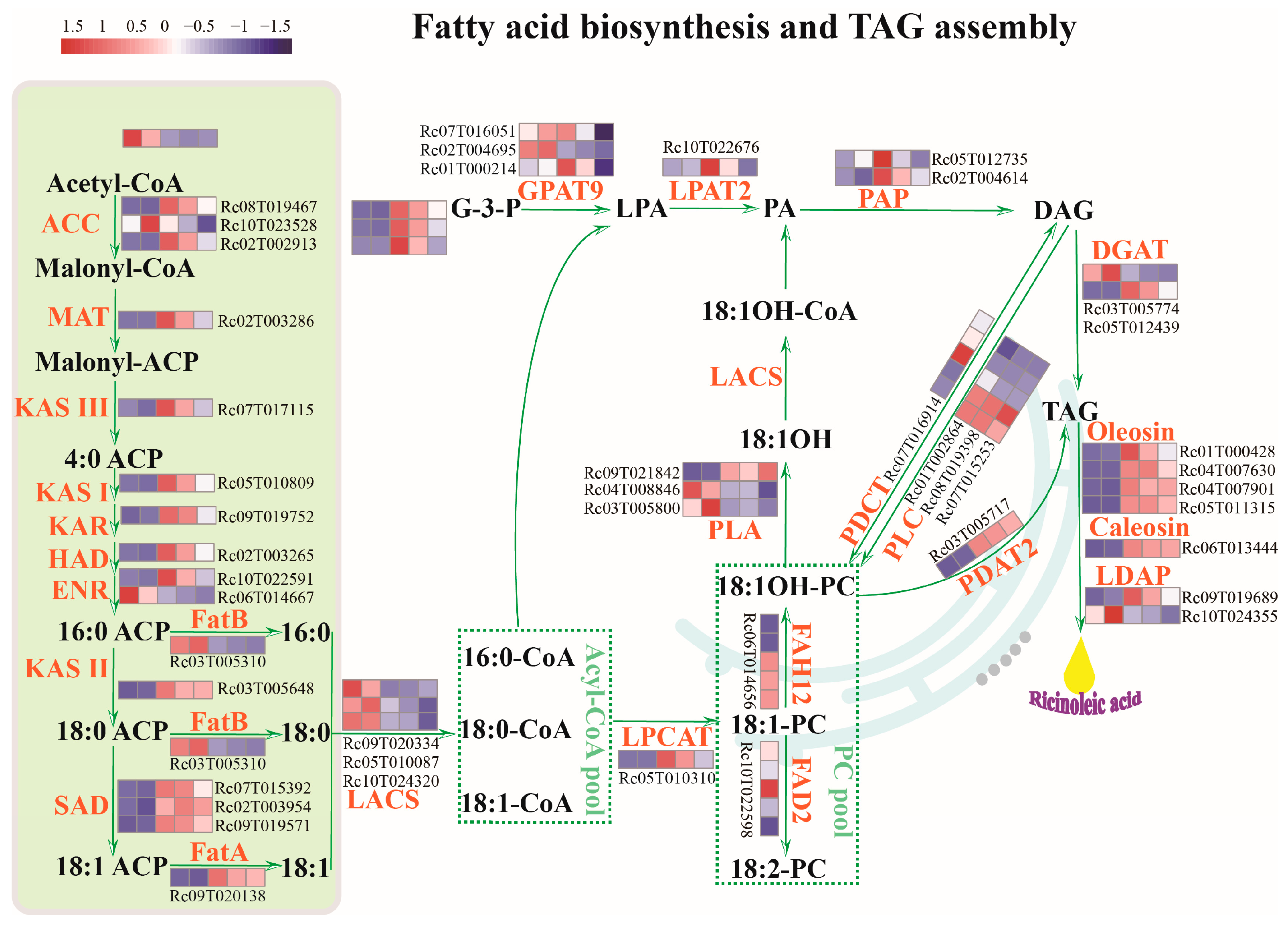
2.2. Identification of Key Enzyme Genes Involved in the Production of Substrates for Fatty Acid Biosynthesis in the Glycolysis Pathway
2.3. Identification of Key Rate-Limiting Enzyme Genes Involved in Fatty Acid and TAG Biosynthesis in Developing Castor Seeds
2.4. Detection of The Transcription Factors Involved in Oil Accumulation
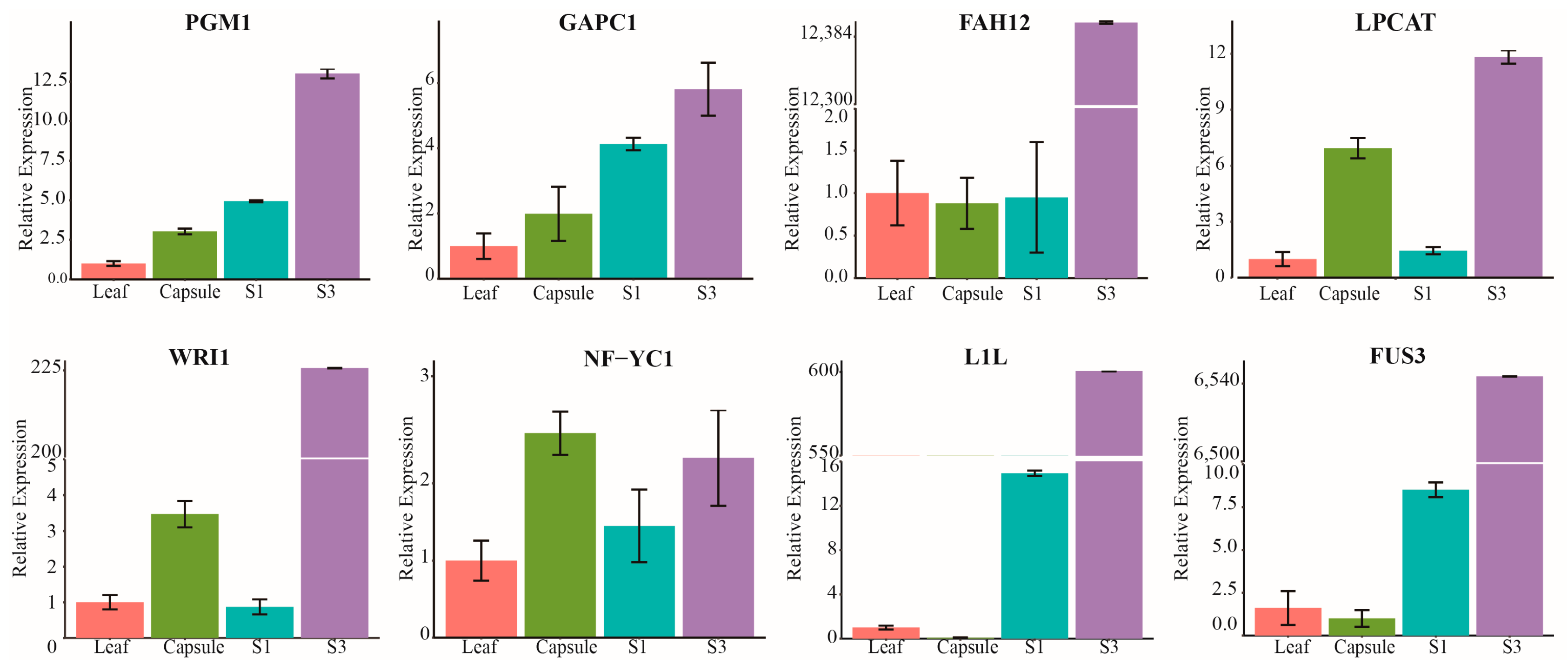
2.5. Detection of The Expression Patterns of Oil Accumulation-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Transcriptome Data Analysis
4.3. Identification of The Genes Related to Oil Accumulation in The Castor Genome
4.4. Promoter Analysis and Subcellular Localization Prediction
4.5. Gene Expression Pattern Analysis
4.6. qRT-PCR Validation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, J.; Nelson, E.; Tilman, D.; Polasky, S.; Tiffany, D. Environmental, economic, and energetic costs and benefits of biodiesel and ethanol biofuels. Proc. Natl. Acad. Sci. USA 2006, 103, 11206–11210. [Google Scholar] [CrossRef]
- Joshi, S.; Hadiya, P.; Shah, M.; Sircar, A. Techno-economical and experimental analysis of biodiesel production from used cooking oil. Biophys. Econ. Resour. Qual. 2019, 4, 2. [Google Scholar] [CrossRef]
- Randelli, F. An integrated analysis of production costs and net energy balance of biofuels. Reg. Environ. Chang. 2008, 9, 221–229. [Google Scholar] [CrossRef]
- Meira, M.; Quintella, C.M.; Ribeiro, E.M.O.; Silva, H.R.G.; Guimarães, A.K. Overview of the challenges in the production of biodiesel. Biomass Conv. Bioref. 2014, 5, 321–329. [Google Scholar] [CrossRef]
- Bhuiya, M.M.; Rasul, M.; Khan, M.M.; Ashwath, N.; Azad, A.K.; Hazrat, M.A. Second generation biodiesel: Potential alternative to-edible oil-derived biodiesel. Energy Procedia 2014, 61, 1969–1972. [Google Scholar] [CrossRef]
- Mubofu, E.B. Castor oil as a potential renewable resource for the production of functional materials. Sustain. Chem. Process. 2016, 4, 11. [Google Scholar] [CrossRef]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef]
- Oswalt, J.S.; Jacob, M.R.; Liv, S.S.; Dick, L.A.; Bednarz, C.W.; Ritchie, G.L. Plant height and seed yield of castor (Ricinus communis L.) sprayed with growth retardants and harvest aid chemicals. Ind. Crops Prod. 2014, 61, 272–277. [Google Scholar] [CrossRef]
- Carrino, L.; Visconti, D.; Fiorentino, N.; Fagnano, M. Biofuel production with castor bean: A win–win strategy for marginal land. Agronomy 2020, 10, 1690. [Google Scholar] [CrossRef]
- Bates, P.D.; Johnson, S.R.; Cao, X.; Li, J.; Nam, J.W.; Jaworski, J.G.; Ohlrogge, J.B.; Browse, J. Fatty acid synthesis is inhibited by inefficient utilization of unusual fatty acids for glycerolipid assembly. Proc. Natl. Acad. Sci. USA 2014, 111, 1204–1209. [Google Scholar] [CrossRef]
- Bates, P.D. Understanding the control of acyl flux through the lipid metabolic network of plant oil biosynthesis. Plant Lipid. Biolog. 2016, 1861, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Z.; Kong, Q.; Lim, A.R.Q.; Lu, S.P.; Zhao, H.; Guo, L.; Yuan, L.; Ma, W. Transcriptional regulation of oil biosynthesis in seed plants: Current understanding, applications, and perspectives. Plant Commun. 2022, 3, 100328. [Google Scholar] [CrossRef] [PubMed]
- Houston, N.L.; Hajduch, M.; Thelen, J.J. Quantitative proteomics of seed filling in castor: Comparison with soybean and rapeseed reveals differences between photosynthetic and nonphotosynthetic seed metabolism. Plant Physiol. 2009, 151, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Lee, K.R.; Go, Y.S.; Jung, J.H.; Suh, M.C.; Kim, J.B. Endoplasmic reticulum-located PDAT1-2 from castor bean enhances hydroxy fatty acid accumulation in transgenic plants. Plant Cell Physiol. 2011, 52, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wu, D.; Yang, T.Q.; Sun, C.; Wang, Z.Q.; Han, B.; Wu, S.B.; Yu, A.M.; Chapman, M.A.; Muraguri, S.; et al. Genomic insights into the origin, domestication and genetic basis of agronomic traits of castor bean. Genome Biol. 2021, 22, 113. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Wu, D.; Zhang, Y.Y.; Li, D.Z.; Xu, W.; Liu, A.Z. Epigenetic regulation of seed-specific gene expression by DNA methylation valleys in castor bean. BMC Biol. 2022, 20, 57. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.M.; Li, F.; Liu, A.Z. Comparative proteomic and transcriptomic analyses provide new insight into the formation of seed size in castor bean. BMC Plant Biol. 2020, 20, 48. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.S.; Bewley, J.D. Seed Development in Ricinus communis (Castor Bean). I. Descriptive Morphology. Can. J. Bot. 1982, 60, 1751–1760. [Google Scholar] [CrossRef]
- Jin, P.; Wu, D.; Dai, H.; Sun, R.; Liu, A.Z. Characterization and functional divergence of genes encoding sucrose transporters in oilseeds castor bean. Oil Crop Sci. 2022, 7, 31–39. [Google Scholar] [CrossRef]
- Zhang, Y.; Mulpuri, S.; Liu, A.Z. High light exposure on seed coat increases lipid accumulation in seeds of castor bean (Ricinus communis L.), a nongreen oilseed crop. Photosynth. Res. 2016, 128, 125–140. [Google Scholar] [CrossRef]
- Braun, D.M. Phloem Loading and Unloading of Sucrose: What a Long, Strange Trip from Source to Sink. Annu. Rev. Plant Biol. 2022, 73, 553–584. [Google Scholar] [CrossRef] [PubMed]
- Cabello, S.; Lorenz, C.; Crespo, S.; Cabrera, J.; Ludwig, R.; Escobar, C.; Hofmann, J. Altered sucrose synthase and invertase expression affects the local and systemic sugar metabolism of nematode-infected Arabidopsis thaliana plants. J. Exp. Bot. 2014, 65, 201–212. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, B.; Plaxton, W.C. Multifaceted functions of post-translational enzyme modifications in the control of plant glycolysis. Curr. Opin. Plant Biol. 2020, 55, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Periappuram, C.; Steinhauer, L.; Barton, D.L.; Taylor, D.C.; Chatson, B.; Zou, J. The plastidic phosphoglucomutase from Arabidopsis. A reversible enzyme reaction with an important role in metabolic control. Plant Physiol. 2000, 122, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Granot, D.; Kelly, G.; Stein, O.; David-Schwartz, R. Substantial roles of hexokinase and fructokinase in the effects of sugars on plant physiology and development. J. Exp. Bot. 2014, 65, 809–819. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, H.J.; Zhang, W.J.; Wang, N.; Zhang, S.J.; Chu, C.C.; Liu, C.M. Engineering of the cytosolic form of phosphoglucose isomerase into chloroplasts improves plant photosynthesis and biomass. New Phytol. 2021, 231, 315–325. [Google Scholar] [CrossRef]
- Carrera, D.Á.; George, G.M.; Fischer-Stettler, M.; Galbier, F.; Eicke, S.; Truernit, E.; Streb, S.; Zeeman, S.C.; Raines, C. Distinct plastid fructose bisphosphate aldolases function in photosynthetic and non-photosynthetic metabolism in Arabidopsis. J. Exp. Bot. 2021, 72, 3739–3755. [Google Scholar] [CrossRef]
- Yoshida, K.; Hisabori, T. Biochemical basis for redox regulation of chloroplast-localized phosphofructokinase from Arabidopsis thaliana. Plant Cell Physiol. 2021, 62, 401–410. [Google Scholar] [CrossRef]
- Zhao, Y.; Cao, P.; Cui, Y.F.; Liu, D.X.; Li, J.P.; Zhao, Y.B.; Yang, S.Q.; Zhang, B.; Zhou, R.N.; Sun, M.H.; et al. Enhanced production of seed oil with improved fatty acid composition by overexpressing NAD(+)-dependent glycerol-3-phosphate dehydrogenase in soybean. J. Integr. Plant Biol. 2021, 63, 1036–1053. [Google Scholar] [CrossRef]
- Myers, T.D.; Palladino, M.J. Newly discovered roles of triosephosphate isomerase including functions within the nucleus. Mol. Med. 2023, 29, 18. [Google Scholar] [CrossRef]
- Rawsthorne, S. Carbon flux and fatty acid synthesis in plants. Prog. Lipid Res. 2002, 41, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Tellez, S.; Anoman, A.D.; Flores-Tornero, M.; Toujani, W.; Alseek, S.; Fernie, A.R.; Nebauer, S.G.; Munoz-Bertomeu, J.; Segura, J.; Ros, R. Phosphoglycerate kinases are co-regulated to adjust metabolism and to optimize growth. Plant Physiol. 2018, 176, 1182–1198. [Google Scholar] [CrossRef] [PubMed]
- Andriotis, V.M.; Kruger, N.J.; Pike, M.J.; Smith, A.M. Plastidial glycolysis in developing Arabidopsis embryos. New Phytol. 2010, 185, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Flugge, U.I.; Hausler, R.E.; Ludewig, F.; Gierth, M. The role of transporters in supplying energy to plant plastids. J. Exp. Bot. 2011, 62, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.L.; Du, X.Q.; Zhang, F.X.; Zhang, F.; Hu, Y.; Liu, S.C.; Jiang, X.N.; Wang, G.D.; Liu, D. A mutation in the E2 subunit of the mitochondrial pyruvate dehydrogenase complex in Arabidopsis reduces plant organ size and enhances the accumulation of amino acids and intermediate products of the TCA cycle. Planta 2012, 236, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.P.; Kroon, J.T.; Swarbreck, D.; Febrer, M.; Larson, T.R.; Graham, I.A.; Caccamo, M.; Slabas, A.R. Tissue-specific whole transcriptome sequencing in castor, directed at understanding triacylglycerol lipid biosynthetic pathways. PLoS ONE 2012, 7, 30100. [Google Scholar] [CrossRef]
- Cagliari, A.; Margis-Pinheiro, M.; Loss, G.; Mastroberti, A.A.; de Araujo Mariath, J.E.; Margis, R. Identification and expression analysis of castor bean (Ricinus communis) genes encoding enzymes from the triacylglycerol biosynthesis pathway. Plant Sci. 2010, 179, 499–509. [Google Scholar] [CrossRef]
- Bates, P.D.; Browse, J. The Significance of Different Diacylgycerol Synthesis Pathways on Plant Oil Composition and Bioengineering. Front. Plant Sci. 2012, 3, 147. [Google Scholar] [CrossRef]
- Hatanaka, T.; Tomita, Y.; Matsuoka, D.; Sasayama, D.; Fukayama, H.; Azuma, T.; Soltani Gishini, M.F.; Hildebrand, D.; Nakamura, Y. Different acyl-CoA: Diacylglycerol acyltransferases vary widely in function, and a targeted amino acid substitution enhances oil accumulation. J. Exp. Bot. 2022, 73, 3030–3043. [Google Scholar] [CrossRef]
- Hayashi, M.; Tanaka, M.; Yamamoto, S.; Nakagawa, T.; Kanai, M.; Anegawa, A.; Ohnishi, M.; Mimura, T.; Nishimura, M. Plastidial folate prevents starch biosynthesis triggered by sugar influx into non-photosynthetic plastids of Arabidopsis. Plant Cell Physiol. 2017, 58, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Ren, Y.L.; Wang, C.M.; Zhang, H.; Wang, F.; Chen, J.; Liu, X.; Zheng, T.H.; Cai, M.H.; Zeng, Z.Q.; et al. OsVIN2 encodes a vacuolar acid invertase that affects grain size by altering sugar metabolism in rice. Plant Cell Rep. 2019, 38, 1273–1290. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ruan, Y.L. New insights into roles of cell wall invertase in early seed development revealed by comprehensive spatial and temporal expression patterns of GhCWIN1 in cotton. Plant Physiol. 2012, 160, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, H.; Zhang, Y.; Kang, T.; Zhang, L.; Tong, J.H.; Xiao, L.T.; Zhang, H.Z. Constitutive expression of cell wall invertase genes increases grain yield and starch content in maize. Plant Biotechnol. J. 2013, 11, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Weselake, R.J.; Taylor, D.C.; Rahman, M.H.; Shah, S.; Laroche, A.; McVetty, P.B.E.; Harwood, J.L. Increasing the flow of carbon into seed oil. Bio. Adv. 2009, 27, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Ruzlan, N.; Low, Y.S.J.; Win, W.; Azizah Musa, N.; Ong, A.L.; Chew, F.T.; Appleton, D.; Mohd, Y.H.; Kulaveerasingam, H. Key glycolytic branch influences mesocarp oil content in oil palm. Sci. Rep. 2017, 7, 9626. [Google Scholar] [CrossRef] [PubMed]
- Rius, S.P.; Casati, P.; Iglesias, A.A.; Gomez-Casati, D.F. Characterization of Arabidopsis lines deficient in GAPC-1, a cytosolic NAD-dependent glyceraldehyde-3-phosphate dehydrogenase. Plant Physiol. 2008, 148, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Troncoso-Ponce, M.A.; Rivoal, J.; Venegas-Calerón, M.; Dorion, S.; Sánchez, R.; Cejudo, F.J.; Garcés, R.; Martínez-Force, E. Molecular cloning and biochemical characterization of three phosphoglycerate kinase isoforms from developing sunflower (Helianthus annuus L.) seeds. Phytochemistry 2012, 79, 27–38. [Google Scholar] [CrossRef]
- van de Loo, F.J.; Broun, P.; Turner, S.; Somerville, C. An oleate 12-hydroxylase from Ricinus communis L. is a fatty acyl desaturase homolog. Proc. Natl. Acad. Sci. USA 1995, 92, 6743–6747. [Google Scholar] [CrossRef]
- Lunn, D.; Wallis, J.G.; Browse, J. A multigene approach secures hydroxy fatty acid production in Arabidopsis. J. Exp. Bot. 2022, 73, 2875–2888. [Google Scholar] [CrossRef]
- Lunn, D.; Le, A.; Wallis, J.G.; Browse, J. Castor LPCAT and PDAT1A Act in Concert to Promote Transacylation of Hydroxy-Fatty Acid onto Triacylglycerol. Plant Physiol. 2020, 184, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, A.; Stahl, U.; Lenman, M.; Banas, A.; Lee, M.; Sandager, L.; Ronne, H.; Stymne, S. Phospholipid:diacylglycerol acyltransferase: An enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. USA 2000, 97, 6487–6492. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.Z.; Mao, W.W.; Jia, M.R.; Xing, S.N.; Ali, U.; Zhao, Y.Y.; Chen, Y.T.; Cao, M.L.; Dai, Z.R.; Zhang, K.; et al. FaMYB44.2, a transcriptional repressor, negatively regulates sucrose accumulation in strawberry receptacles through interplay with FaMYB10. J. Exp. Bot. 2018, 69, 4805–4820. [Google Scholar] [CrossRef] [PubMed]
- van Erp, H.; Bates, P.D.; Burgal, J.; Shockey, J.; Browse, J. Castor phospholipid:diacylglycerol acyltransferase facilitates efficient metabolism of hydroxy fatty acids in transgenic Arabidopsis. Plant Physiol. 2011, 155, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Guo, W.; Li, J.C.; Yue, P.T.; Bu, H.D.; Jiang, J.; Liu, W.T.; Xu, Y.X.; Yuan, H.; Li, T.; et al. Histone Acetylation at the Promoter for the Transcription Factor PuWRKY31 Affects Sucrose Accumulation in Pear Fruit. Plant Physiol. 2020, 182, 2035–2046. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.Q.; Qin, H.; Zhou, J.H.; Quan, R.D.; Lu, X.Y.; Huang, R.F.; Zhang, H.W. OsERF2 controls rice root growth and hormone responses through tuning expression of key genes involved in hormone signaling and sucrose metabolism. Plant Mol. Biol. 2016, 90, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.M.; Kwong, R.W.; Park, S.; Le, B.H.; Baden, R.; Cagliari, A.; Hashimoto, M.; Munoz, M.D.; Fischer, R.L.; Goldberg, R.B.; et al. LEC1 sequentially regulates the transcription of genes involved in diverse developmental processes during seed development. Proc. Natl. Acad. Sci. USA 2017, 114, 6710–6719. [Google Scholar] [CrossRef]
- Zaret, K.S.; Carroll, J.S. Pioneer transcription factors: Establishing competence for gene expression. Genes Dev. 2011, 25, 2227–2241. [Google Scholar] [CrossRef]
- Kwong, R.W.; Bui, A.Q.; Lee, H.; Kwong, L.W.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell 2003, 15, 5–18. [Google Scholar] [CrossRef]
- Tan, H.L.; Yang, X.H.; Zhang, F.X.; Zheng, X.; Qu, C.M.; Mu, J.Y.; Fu, F.Y.; Li, J.N.; Guan, R.Z.; Zhang, H.S.; et al. Enhanced seed oil production in canola by conditional expression of Brassica napus LEAFY COTYLEDON1 and LEC1-LIKE in developing seeds. Plant Physiol. 2011, 156, 1577–1588. [Google Scholar] [CrossRef]
- Mendes, A.; Kelly, A.A.; van Erp, H.; Shaw, E.; Powers, S.J.; Kurup, S.; Eastmond, P.J. bZIP67 regulates the omega-3 fatty acid content of Arabidopsis seed oil by activating fatty acid desaturase3. Plant Cell 2013, 25, 3104–3116. [Google Scholar] [CrossRef] [PubMed]
- Jo, L.; Pelletier, J.M.; Hsu, S.W.; Baden, R.; Goldberg, R.B.; Harada, J.J. Combinatorial interactions of the LEC1 transcription factor specify diverse developmental programs during soybean seed development. Proc. Natl. Acad. Sci. USA 2020, 14, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Maeo, K.; Tokuda, T.; Ayame, A.; Mitsui, N.; Kawai, T.; Tsukagoshi, H.; Ishiguro, S.; Nakamura, K. An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis. Plant J. 2009, 60, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.J.; Zhang, X.F.; Xue, H.W. Rice aleurone layer specific OsNF-YB1 regulates grain filling and endosperm development by interacting with an ERF transcription factor. J. Exp. Bot. 2016, 67, 6399–6411. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Beisson, F.; Pollard, M.; Ohlrogge, J. Oil content of Arabidopsis seeds: The influence of seed anatomy, light and plant-to-plant variation. Phytochemistry 2006, 67, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Hernández-Plaza, A.; Szklarczyk, D.; Botas, J.; Cantalapiedra, C.P.; Giner-Lamia, J.; Mende, D.R.; Kirsch, R.; Rattei, T.; Letunic, I.; Jensen, L.J.; et al. eggNOG 6.0: Enabling comparative genomics across 12535 organisms. Nucleic Acids Res. 2023, 51, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.Z.; Hu, E.Q.; Xu, S.B.; Chen, M.J.; Guo, P.F.; Dai, Z.H.; Feng, T.Z.; Zhou, L.; Tang, W.L.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, A.; Zhou, Z.; Chen, Y.; Sun, J.; Li, P.; Gu, X.; Liu, A. Functional Genome Analyses Reveal the Molecular Basis of Oil Accumulation in Developing Seeds of Castor Beans. Int. J. Mol. Sci. 2024, 25, 92. https://doi.org/10.3390/ijms25010092
Yu A, Zhou Z, Chen Y, Sun J, Li P, Gu X, Liu A. Functional Genome Analyses Reveal the Molecular Basis of Oil Accumulation in Developing Seeds of Castor Beans. International Journal of Molecular Sciences. 2024; 25(1):92. https://doi.org/10.3390/ijms25010092
Chicago/Turabian StyleYu, Anmin, Zekun Zhou, Yizhong Chen, Jing Sun, Ping Li, Xu Gu, and Aizhong Liu. 2024. "Functional Genome Analyses Reveal the Molecular Basis of Oil Accumulation in Developing Seeds of Castor Beans" International Journal of Molecular Sciences 25, no. 1: 92. https://doi.org/10.3390/ijms25010092
APA StyleYu, A., Zhou, Z., Chen, Y., Sun, J., Li, P., Gu, X., & Liu, A. (2024). Functional Genome Analyses Reveal the Molecular Basis of Oil Accumulation in Developing Seeds of Castor Beans. International Journal of Molecular Sciences, 25(1), 92. https://doi.org/10.3390/ijms25010092