
Matrine Targets BTF3 to Inhibit the Growth of Canine Mammary Tumor Cells

 ,
,
Abstract
:1. Introduction
2. Results
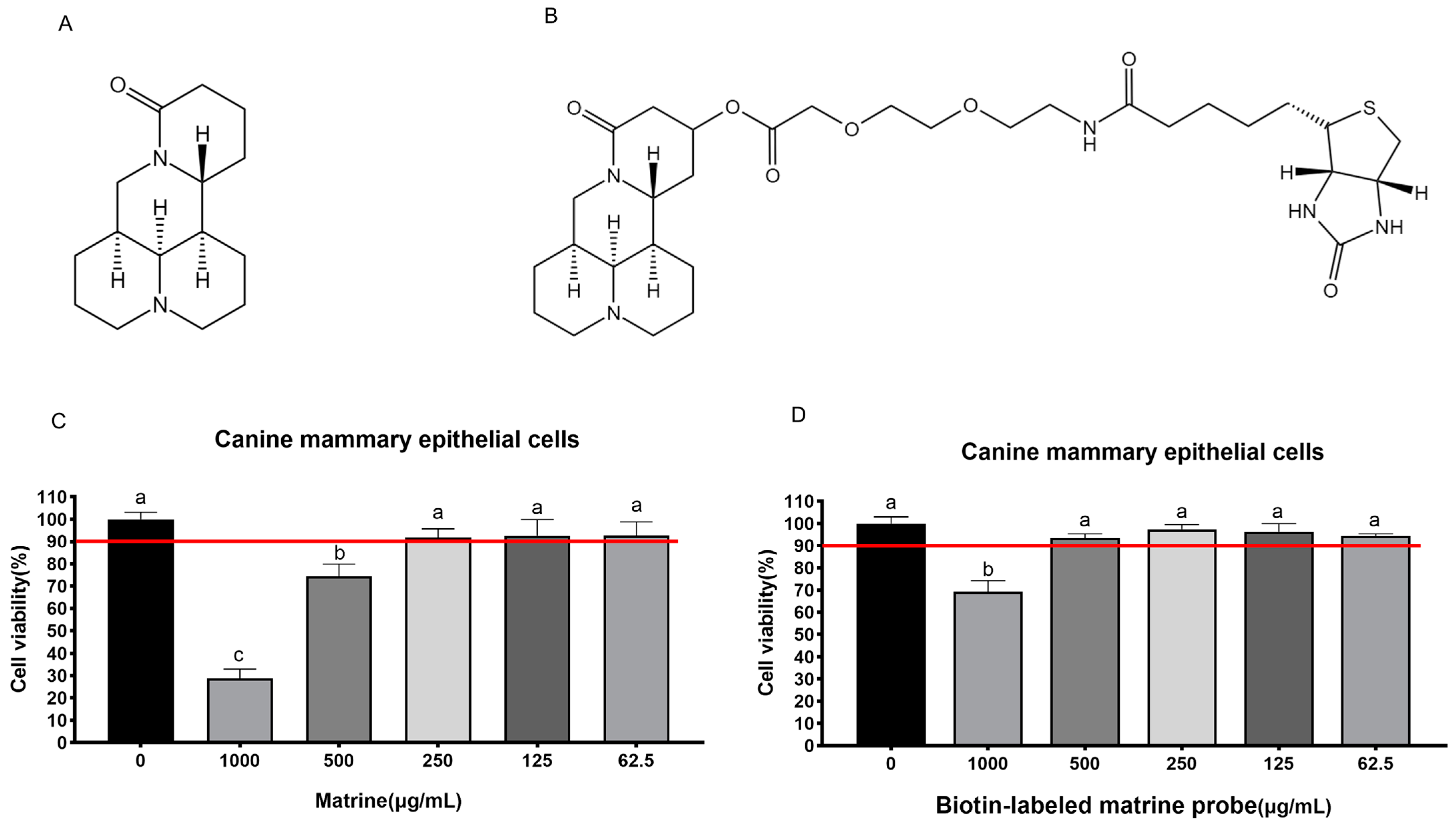
2.1. Maximum Safe Concentration of Matrine and Biotin-Labeled Matrine Probe
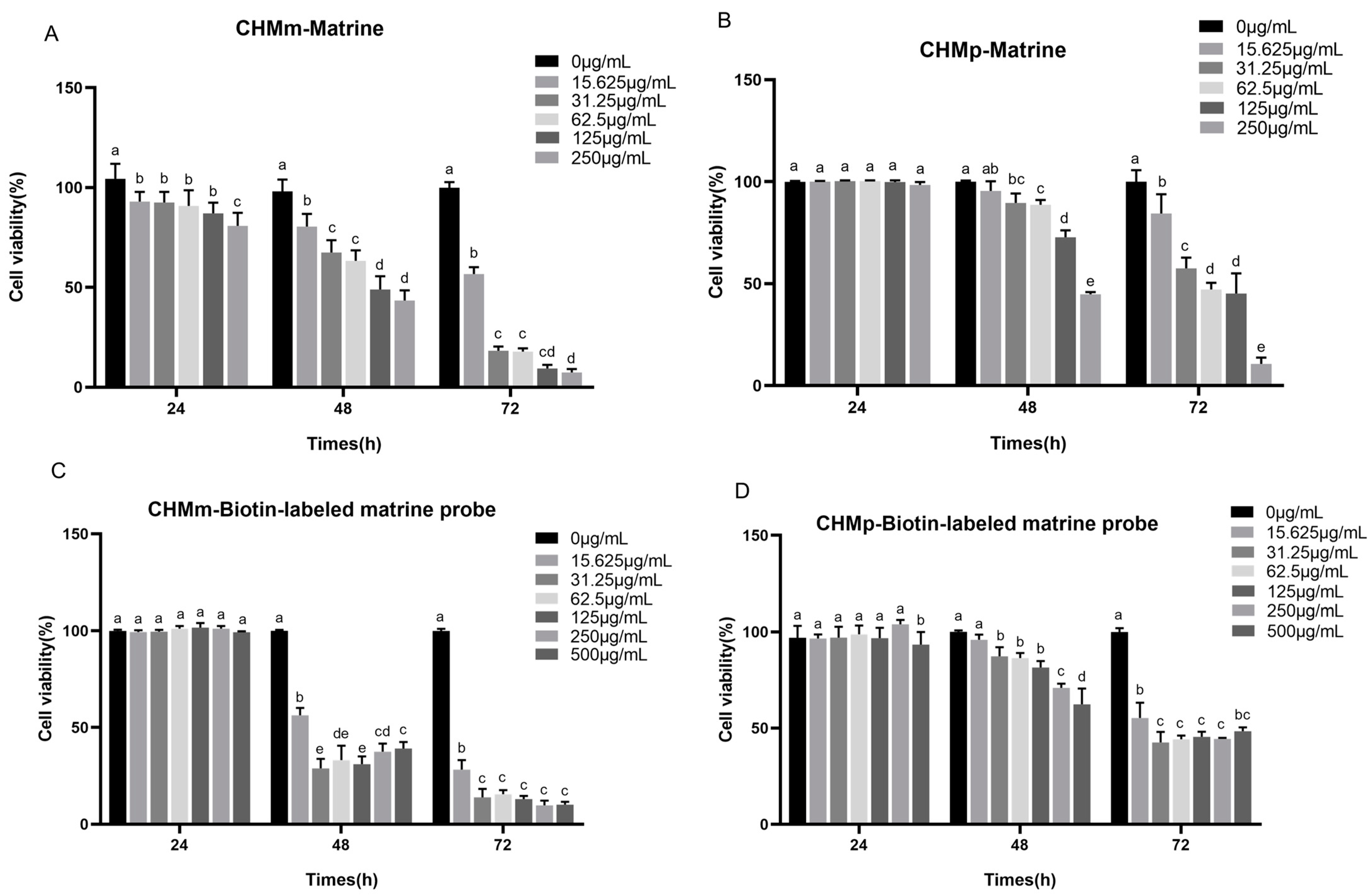
2.2. Cell Proliferation Assays
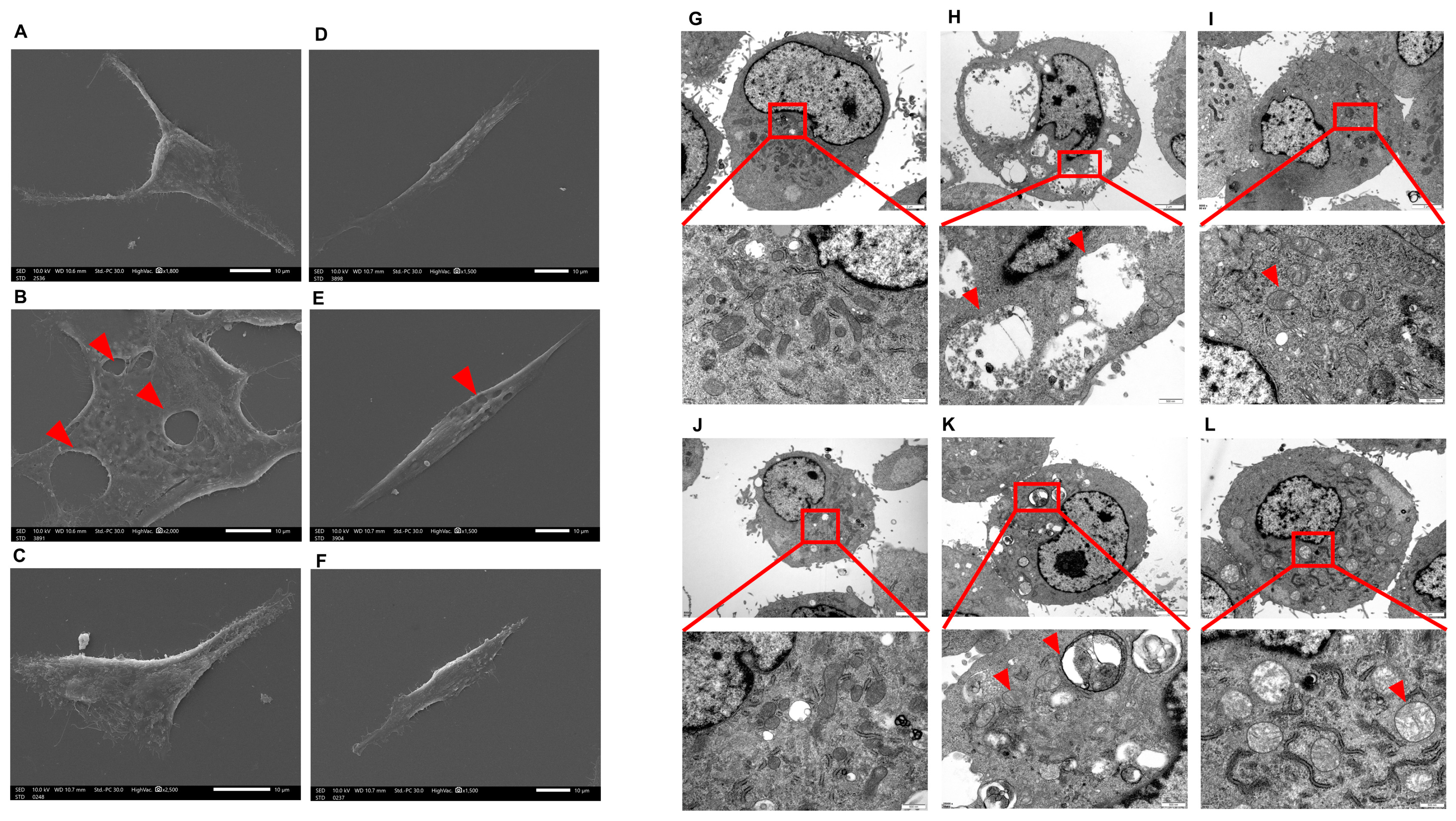
2.3. Scanning Electron Microscope (SEM) and Transmission Electron Microscope (TEM) Analysis
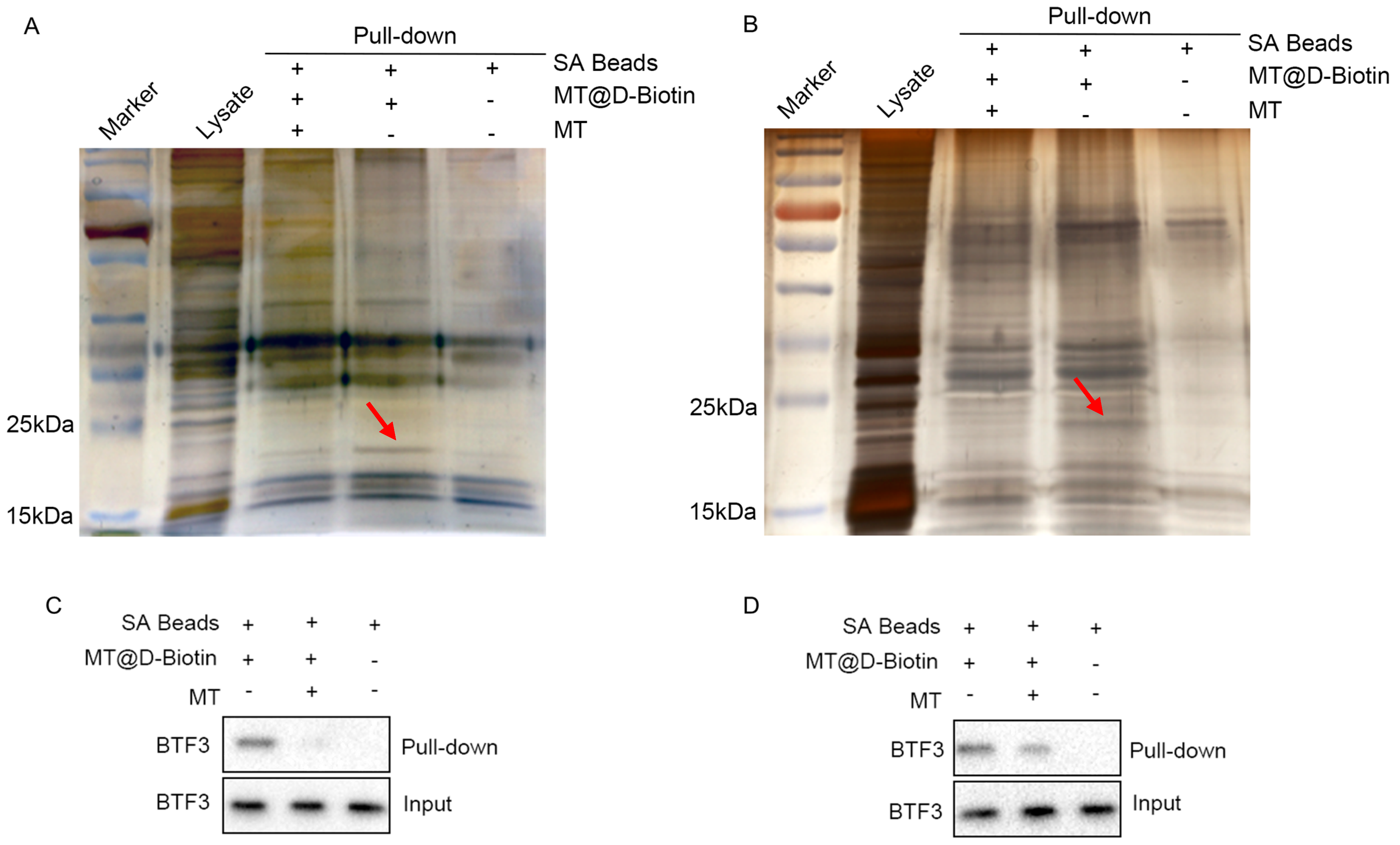
2.4. Enrichment of Matrine Targets
2.5. GO and KEGG Analysis of Potential Targets of MATRINE
2.6. PPI Protein Interaction Network Construction
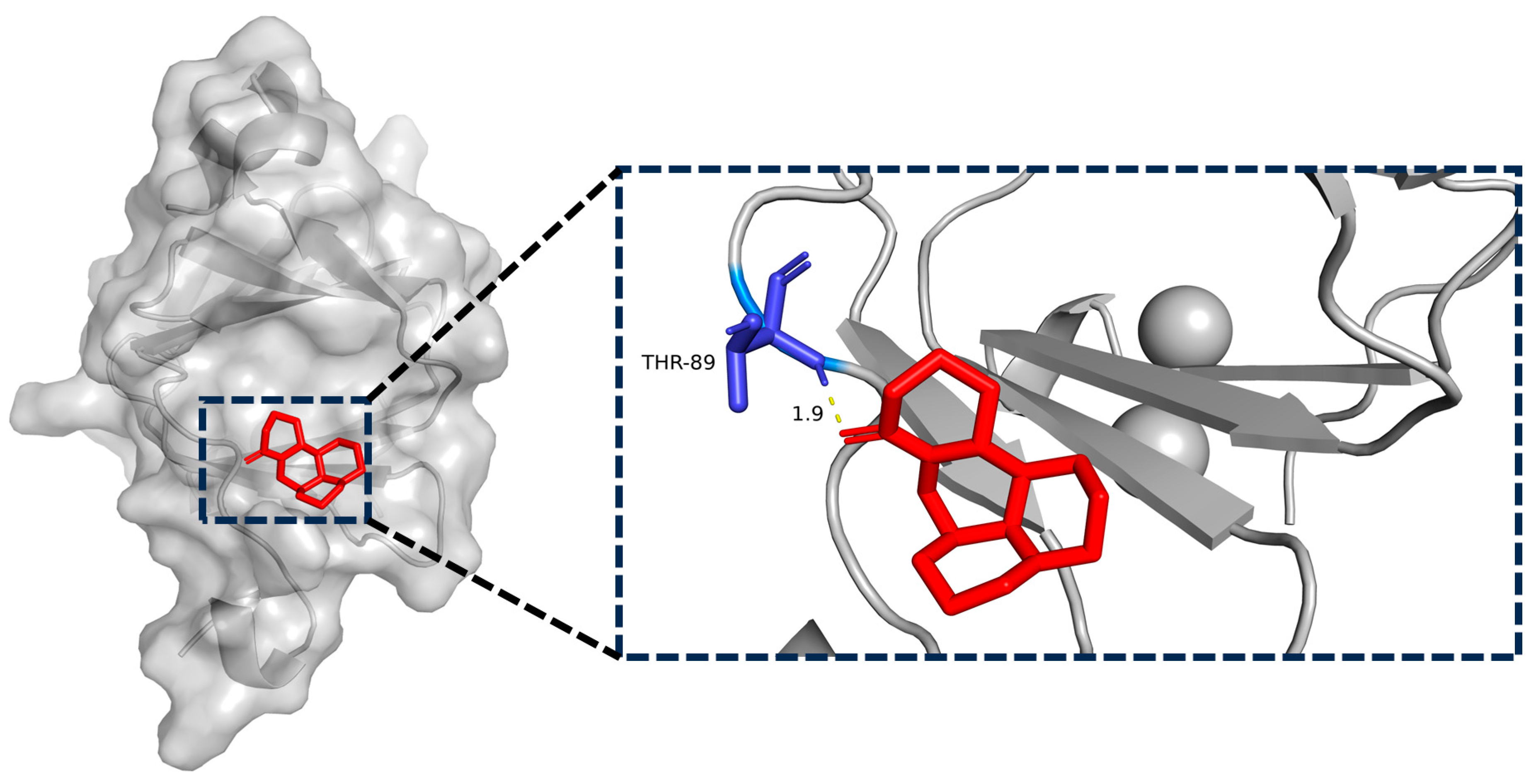
2.7. BTF3 Is the Specific Binding Protein of Matrine
2.8. Matrine and BTF3 Molecular Docking Simulations
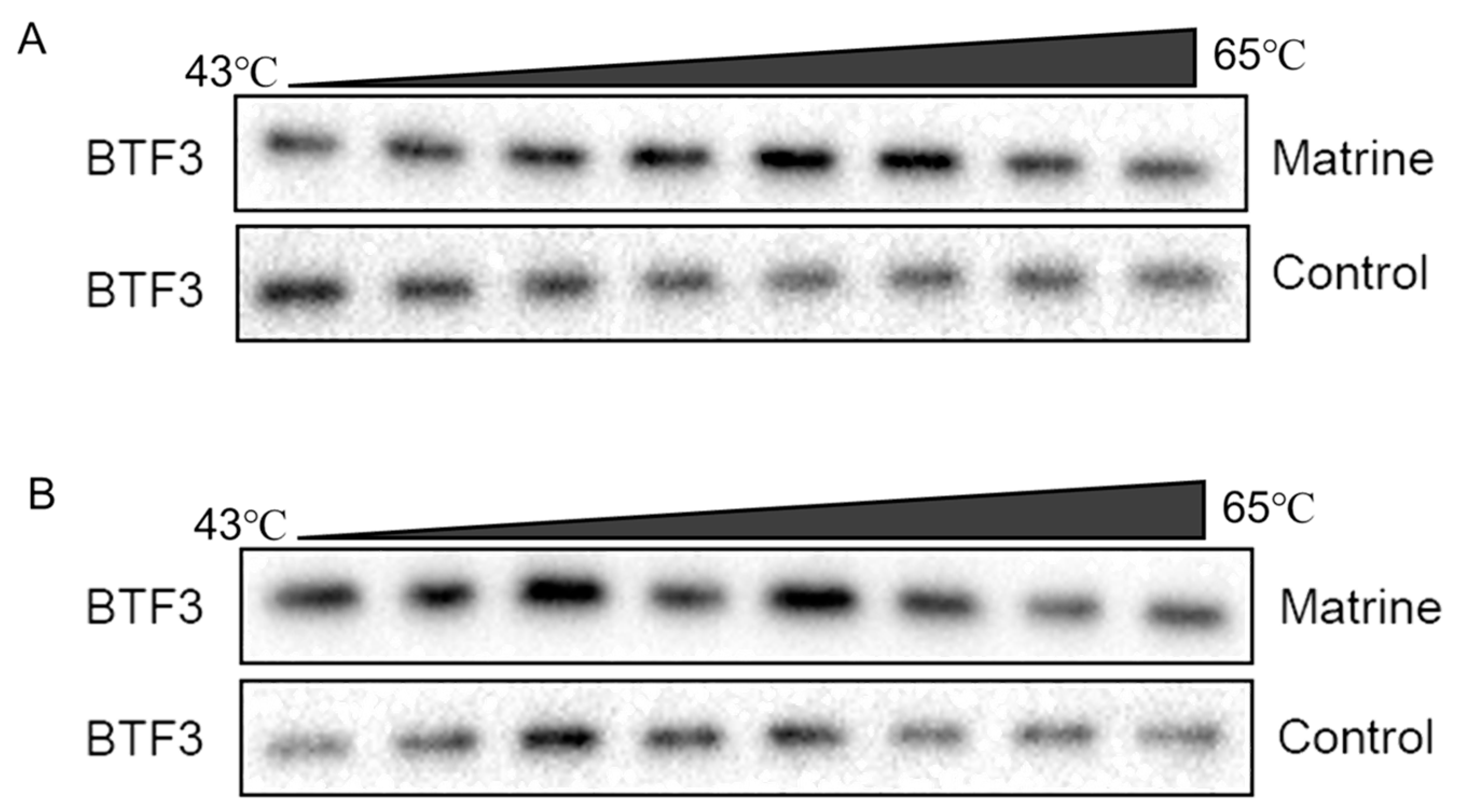
2.9. CETSA Identifies BTF3 as a Target of Matrine Action
2.10. Effects of Matrine on Btf3 Gene in Canine Mammary Tumor Cells
2.11. Effects of Matrine on BTF3 Protein in Canine Mammary Tumor Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Maximum Safe Concentration Detection
4.3. Cell Proliferation Assay
4.4. SEM and TEM Analysis
4.5. Pull-Down
4.6. GO and KEGG Analysis
4.7. PPI Network Construction
4.8. SDS-PAGE Silver Nitrate Staining
4.9. Molecular Docking
4.10. CETSA
4.11. qPCR
4.12. Western Blot
4.13. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
| ABPP | Activity-based protein profiling |
| CCK-8 | Cell Counting Kit-8 |
| CETSA | Cellular thermal shift assay |
References
- Mattiuzzi, C.; Lippi, G. Current Cancer Epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast Cancer Statistics, 2022. CA A Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Aleskandarany, M.A.; Vandenberghe, M.E.; Marchiò, C.; Ellis, I.O.; Sapino, A.; Rakha, E.A. Tumour Heterogeneity of Breast Cancer: From Morphology to Personalised Medicine. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2018, 85, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Vogell, A.; Evans, M.L. Cancer Screening in Women. Obstet. Gynecol. Clin. N. Am. 2019, 46, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Mintz, R.L.; Gao, M.A.; Lo, K.; Lao, Y.H.; Li, M.; Leong, K.W. CRISPR Technology for Breast Cancer: Diagnostics, Modeling, and Therapy. Adv. Biosyst. 2018, 2, 1800132. [Google Scholar] [CrossRef] [PubMed]
- Loibl, S.; Poortmans, P.; Morrow, M.; Denkert, C.; Curigliano, G. Breast cancer. Lancet 2021, 397, 1750–1769. [Google Scholar] [CrossRef]
- Swain, S.M.; Miles, D.; Kim, S.B.; Im, Y.H.; Im, S.A.; Semiglazov, V.; Ciruelos, E.; Schneeweiss, A.; Loi, S.; Monturus, E.; et al. Pertuzumab, trastuzumab, and docetaxel for HER2-positive metastatic breast cancer (CLEOPATRA): End-of-study results from a double-blind, randomised, placebo-controlled, phase 3 study. Lancet. Oncol. 2020, 21, 519–530. [Google Scholar] [CrossRef]
- Long, X.; Nephew, K.P. Fulvestrant (ICI 182,780)-dependent interacting proteins mediate immobilization and degradation of estrogen receptor-alpha. J. Biol. Chem. 2006, 281, 9607–9615. [Google Scholar] [CrossRef]
- Fidler, I.J. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef]
- Huber, K.E.; Carey, L.A.; Wazer, D.E. Breast cancer molecular subtypes in patients with locally advanced disease: Impact on prognosis, patterns of recurrence, and response to therapy. Semin. Radiat. Oncol. 2009, 19, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Han, B.; Siegel, E.; Cui, Y.; Giuliano, A.; Cui, X. Breast cancer lung metastasis: Molecular biology and therapeutic implications. Cancer Biol. Ther. 2018, 19, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Li, J.; Zhu, S.; Wu, J.; Chen, C.; Liu, Q.; Wei, W.; Zhang, Y.; Sun, S. Breast cancer subtypes predict the preferential site of distant metastases: A SEER based study. Oncotarget 2017, 8, 27990–27996. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.E. Naturally occurring cancers in dogs: Insights for translational genetics and medicine. ILAR J. 2014, 55, 16–45. [Google Scholar] [CrossRef] [PubMed]
- Rowell, J.L.; McCarthy, D.O.; Alvarez, C.E. Dog models of naturally occurring cancer. Trends Mol. Med. 2011, 17, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Sleeckx, N.; de Rooster, H.; Veldhuis Kroeze, E.J.; Van Ginneken, C.; Van Brantegem, L. Canine mammary tumours, an overview. Reprod. Domest. Anim. Zuchthyg. 2011, 46, 1112–1131. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.M.; Yang, I.S.; Seung, B.J.; Lee, S.; Kim, D.; Ha, Y.J.; Seo, M.K.; Kim, K.K.; Kim, H.S.; Cheong, J.H.; et al. Cross-species oncogenic signatures of breast cancer in canine mammary tumors. Nat. Commun. 2020, 11, 3616. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, L.; Sun, X.; Yang, Q.; Wan, L.; Guo, C. Matrine: A Promising Natural Product With Various Pharmacological Activities. Front. Pharmacol. 2020, 11, 588. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, H.; Sun, P.; Khan, A.; Guo, J.; Zheng, X.; Sun, Y.; Fan, K.; Yin, W.; Li, H. Matrine exhibits antiviral activity in a PRRSV/PCV2 co-infected mouse model. Phytomed. Int. J. Phytother. Phytopharm. 2020, 77, 153289. [Google Scholar] [CrossRef]
- Chu, Y.; Jing, Y.; Zhao, X.; Wang, M.; Zhang, M.; Ma, R.; Ma, W.; Lv, Y.; Zhu, L. Modulation of the HMGB1/TLR4/NF-κB signaling pathway in the CNS by matrine in experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2021, 352, 577480. [Google Scholar] [CrossRef]
- Niu, H.; Zhang, Y.; Wu, B.; Zhang, Y.; Jiang, H.; He, P. Matrine induces the apoptosis of lung cancer cells through downregulation of inhibitor of apoptosis proteins and the Akt signaling pathway. Oncol. Rep. 2014, 32, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Li, J.; Li, T.; Chen, G.; Zhang, Z.; Si, Z. Matrine-induced apoptosis in Hep3B cells via the inhibition of MDM2. Mol. Med. Rep. 2017, 15, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Zhou, D.; Wang, X.; Hu, Z.; Yan, Y.; Huang, J. Matrine Suppresses Proliferation and Invasion of SGC7901 Cells through Inactivation of PI3K/Akt/uPA Pathway. Ann. Clin. Lab. Sci. 2016, 46, 457–462. [Google Scholar] [PubMed]
- Zhou, B.G.; Wei, C.S.; Zhang, S.; Zhang, Z.; Gao, H.M. Matrine reversed multidrug resistance of breast cancer MCF-7/ADR cells through PI3K/AKT signaling pathway. J. Cell. Biochem. 2018, 119, 3885–3891. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Mo, W.; Wang, L.; Wang, X. Matrine suppresses breast cancer metastasis by targeting ITGB1 and inhibiting epithelial-to-mesenchymal transition. Exp. Ther. Med. 2020, 19, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.; Pries, V.; Hedberg, C.; Waldmann, H. Target identification for small bioactive molecules: Finding the needle in the haystack. Angew. Chem. (Int. Ed. Engl.) 2013, 52, 2744–2792. [Google Scholar] [CrossRef]
- Pichler, C.M.; Krysiak, J.; Breinbauer, R. Target identification of covalently binding drugs by activity-based protein profiling (ABPP). Bioorganic Med. Chem. 2016, 24, 3291–3303. [Google Scholar] [CrossRef]
- Zhang, T.; Li, J.; He, Y.; Yang, F.; Hao, Y.; Jin, W.; Wu, J.; Sun, Z.; Li, Y.; Chen, Y.; et al. A small molecule targeting myoferlin exerts promising anti-tumor effects on breast cancer. Nat. Commun. 2018, 9, 3726. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Moskwa, N.; Kang, W.; Fan, T.M.; Lee, C. Canine as a Comparative and Translational Model for Human Mammary Tumor. J. Breast Cancer 2023, 26, 1–13. [Google Scholar] [CrossRef]
- Pinho, S.S.; Carvalho, S.; Cabral, J.; Reis, C.A.; Gärtner, F. Canine tumors: A spontaneous animal model of human carcinogenesis. Transl. Res. J. Lab. Clin. Med. 2012, 159, 165–172. [Google Scholar] [CrossRef]
- Khanna, C.; Lindblad-Toh, K.; Vail, D.; London, C.; Bergman, P.; Barber, L.; Breen, M.; Kitchell, B.; McNeil, E.; Modiano, J.F.; et al. The dog as a cancer model. Nat. Biotechnol. 2006, 24, 1065–1066. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, F.; Peña, L.; Ibisch, C.; Loussouarn, D.; Gama, A.; Rieder, N.; Belousov, A.; Campone, M.; Abadie, J. Canine invasive mammary carcinomas as models of human breast cancer. Part 1: Natural history and prognostic factors. Breast Cancer Res. Treat. 2018, 167, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Jia, L.Y.; Rong, Z.; Zhou, X.; Cao, L.Q.; Li, A.H.; Guo, M.; Jin, J.; Wang, Y.D.; Huang, L.; et al. Research Advances on Matrine. Front. Chem. 2022, 10, 867318. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Gu, Y.; Zhang, Y. The discovery of multi-target compounds with anti-inflammation activity from traditional Chinese medicine by TCM-target effects relationship spectrum. J. Ethnopharmacol. 2022, 293, 115289. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, B.; Feng, S.; Wang, J.; Zhang, F. Target recognition and network pharmacology for revealing anti-diabetes mechanisms of natural product. J. Comput. Sci. 2020, 45, 101186. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, Q.; Yang, H.; Zhang, X.W.; Feng, N.; Wang, J.K.; Liu, T.T.; Zeng, K.W.; Tu, P.F. Allosteric Regulation of IGF2BP1 as a Novel Strategy for the Activation of Tumor Immune Microenvironment. ACS Cent. Sci. 2022, 8, 1102–1115. [Google Scholar] [CrossRef]
- Zhang, X.W.; Feng, N.; Liu, Y.C.; Guo, Q.; Wang, J.K.; Bai, Y.Z.; Ye, X.M.; Yang, Z.; Yang, H.; Liu, Y.; et al. Neuroinflammation inhibition by small-molecule targeting USP7 noncatalytic domain for neurodegenerative disease therapy. Sci. Adv. 2022, 8, eabo0789. [Google Scholar] [CrossRef]
- Harel, A.; Inger, A.; Stelzer, G.; Strichman-Almashanu, L.; Dalah, I.; Safran, M.; Lancet, D. GIFtS: Annotation landscape analysis with GeneCards. BMC Bioinform. 2009, 10, 348. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Chen, L.; Li, H. The dietary compound luteolin inhibits pancreatic cancer growth by targeting BCL-2. Food Funct. 2018, 9, 3018–3027. [Google Scholar] [CrossRef]
- Zheng, X.M.; Black, D.; Chambon, P.; Egly, J.M. Sequencing and expression of complementary DNA for the general transcription factor BTF3. Nature 1990, 344, 556–559. [Google Scholar] [CrossRef]
- Zhang, D.Z.; Chen, B.H.; Zhang, L.F.; Cheng, M.K.; Fang, X.J.; Wu, X.J. Basic Transcription Factor 3 Is Required for Proliferation and Epithelial-Mesenchymal Transition via Regulation of FOXM1 and JAK2/STAT3 Signaling in Gastric Cancer. Oncol. Res. 2017, 25, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, J.; Lu, T.; Fang, Z.; Huang, Z.; Lu, S.; Dai, C.; Li, M. Positive expression of basic transcription factor 3 predicts poor survival of colorectal cancer patients: Possible mechanisms involved. Cell Death Dis. 2019, 10, 509. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Bang, W.; Cho, J.H.; Lee, R.H.; Kim, S.H.; Kim, M.S.; Park, S.M.; Shin, J.C.; Chung, H.J.; Oh, K.B.; et al. Kahweol induces apoptosis by suppressing BTF3 expression through the ERK signaling pathway in non-small cell lung cancer cells. Int. J. Oncol. 2016, 49, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, W.; Wang, L.; Zhang, X.C.; Li, X.; Rao, Z. Crystal structures of NAC domains of human nascent polypeptide-associated complex (NAC) and its αNAC subunit. Protein Cell 2010, 1, 406–416. [Google Scholar] [CrossRef]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein-ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | kDa | Description | Matrine Combined Score | |
|---|---|---|---|---|
| CHMm | CHMp | |||
| BTF3 | 22.1 | Basic Transcription Factor 3 | 7.4 | 7.2 |
| MYH9 | 226.2 | Myosin Heavy Chain 9 | 6.5 | 2.8 |
| KPNB1 | 97.2 | Karyopherin Subunit Beta 1 | 1.7 | 5.9 |
| EIF3D | 65.4 | Eukaryotic Translation Initiation Factor 3 Subunit D | 2.2 | 2.1 |
| RBM39 | 58.8 | RNA-binding motif protein 39 | 2.0 | 1.7 |
| Gene | Sequence | Product Length |
|---|---|---|
| Btf3 | F: TGCGCTCCAACAAGATGAAAG | 105 bp |
| R: TCTTCTTTCGGCGAGCAGTT | ||
| Gapdh | F: GATCCCGCCAACATCAAATG | 166 bp |
| R: TCACGCCCATCACAAACATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Z.; Sun, N.; Noor, F.; Sun, P.; Zhang, H.; Zhong, J.; Yin, W.; Fan, K.; Yang, H.; Zhang, Z.; et al. Matrine Targets BTF3 to Inhibit the Growth of Canine Mammary Tumor Cells. Int. J. Mol. Sci. 2024, 25, 540. https://doi.org/10.3390/ijms25010540
Feng Z, Sun N, Noor F, Sun P, Zhang H, Zhong J, Yin W, Fan K, Yang H, Zhang Z, et al. Matrine Targets BTF3 to Inhibit the Growth of Canine Mammary Tumor Cells. International Journal of Molecular Sciences. 2024; 25(1):540. https://doi.org/10.3390/ijms25010540
Chicago/Turabian StyleFeng, Zijian, Na Sun, Fida Noor, Panpan Sun, Hua Zhang, Jia Zhong, Wei Yin, Kuohai Fan, Huizhen Yang, Zhenbiao Zhang, and et al. 2024. "Matrine Targets BTF3 to Inhibit the Growth of Canine Mammary Tumor Cells" International Journal of Molecular Sciences 25, no. 1: 540. https://doi.org/10.3390/ijms25010540
APA StyleFeng, Z., Sun, N., Noor, F., Sun, P., Zhang, H., Zhong, J., Yin, W., Fan, K., Yang, H., Zhang, Z., Sun, Y., & Li, H. (2024). Matrine Targets BTF3 to Inhibit the Growth of Canine Mammary Tumor Cells. International Journal of Molecular Sciences, 25(1), 540. https://doi.org/10.3390/ijms25010540